
Stand States Drive Disparities in the Carbon Storage Within a Masson Pine Forest Ecosystem


Abstract
1. Introduction
2. Materials and Methods
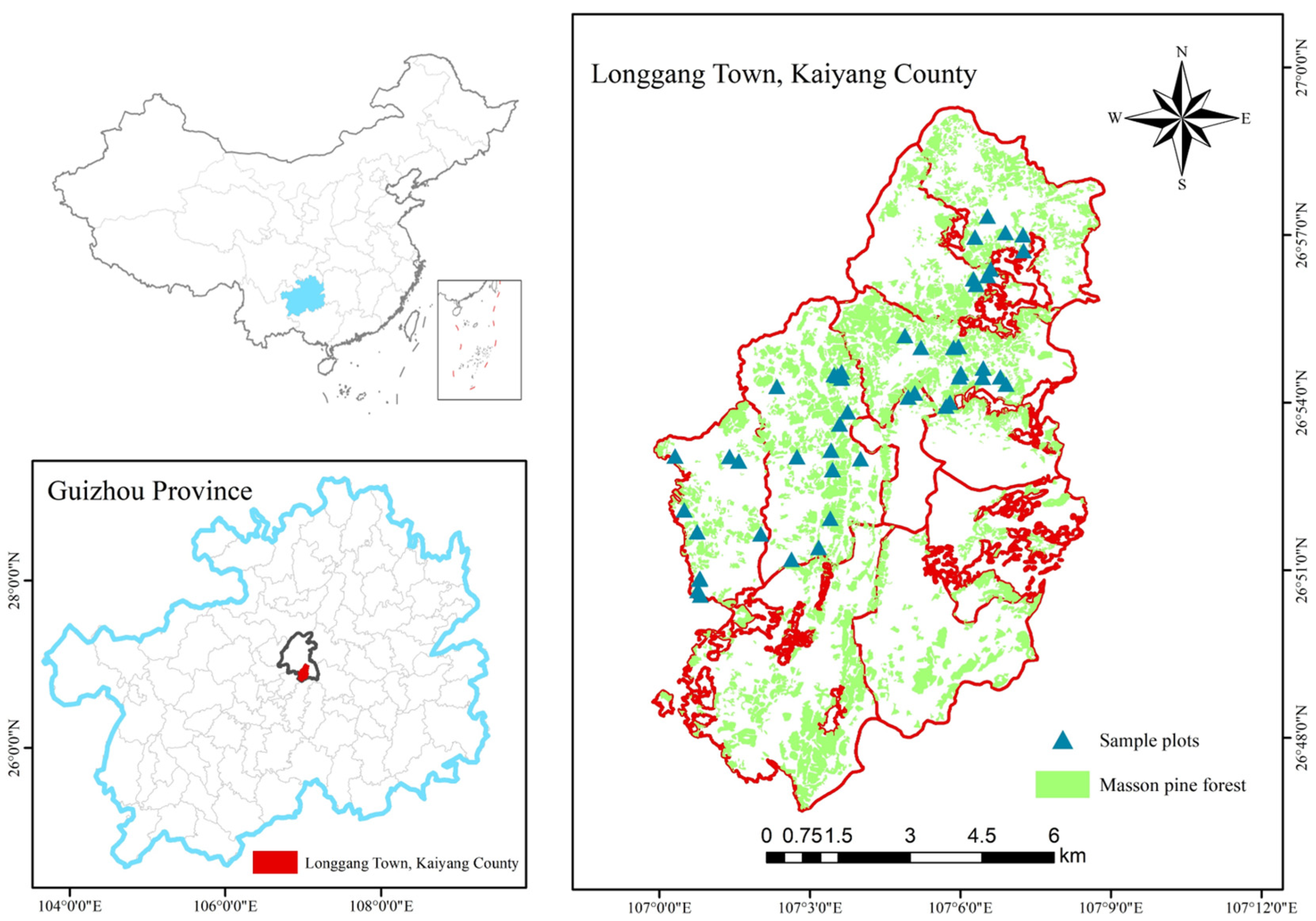
2.1. Study Area
2.2. Plot Establish and Survey
2.3. Sample Collection and Processing
2.4. Evaluation Index Measuring Forest Stand State
2.5. Calculation of C Storage in Forest Ecosystems
2.6. Mantel Test and Canonical Correspondence Analysis (CCA)
2.7. Statistical Analyses
3. Results
3.1. Characteristics of Stand State and C Storage in P. massoniana Forests
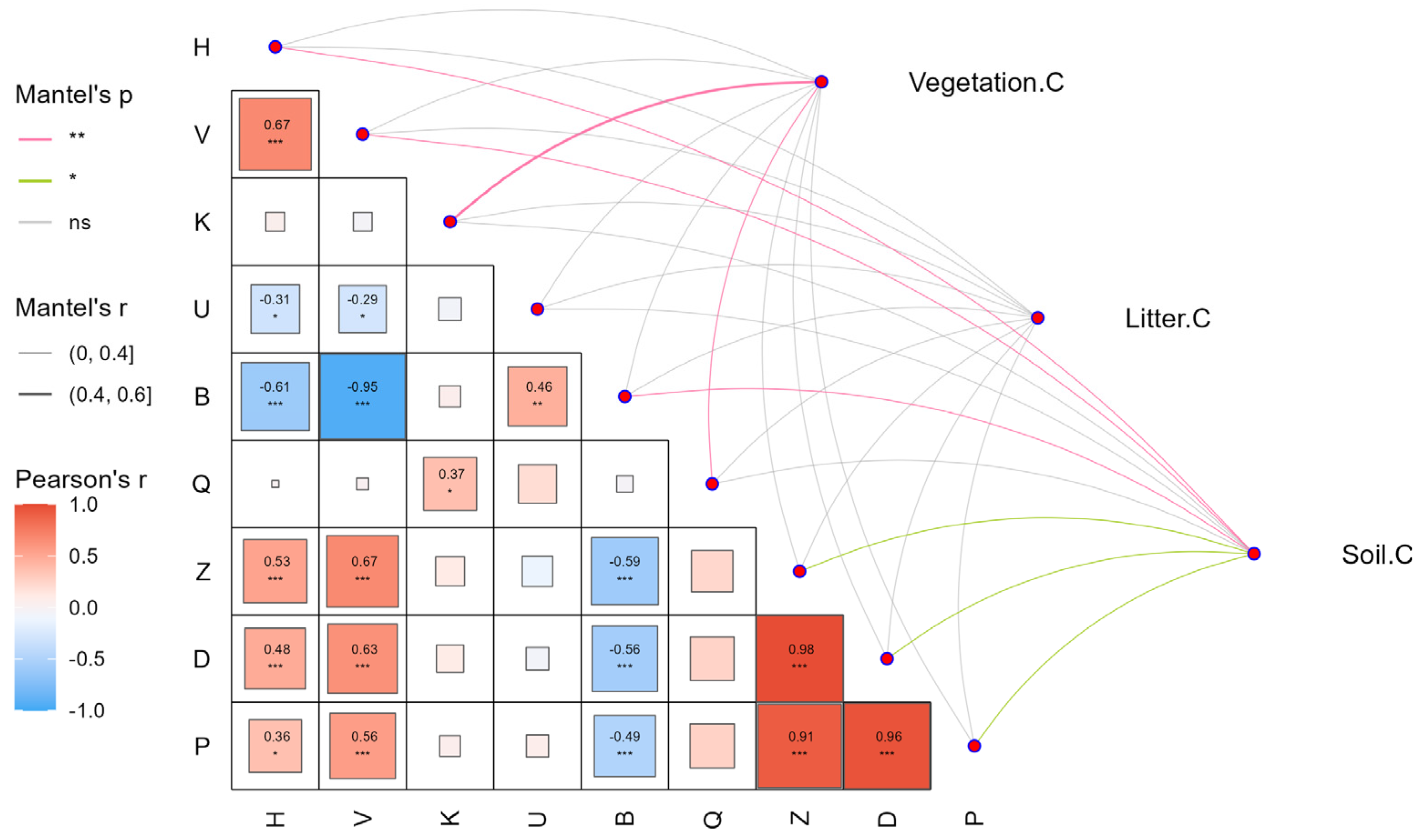
3.2. The Correlation Between Stand State and C Storage in P. massoniana Forests
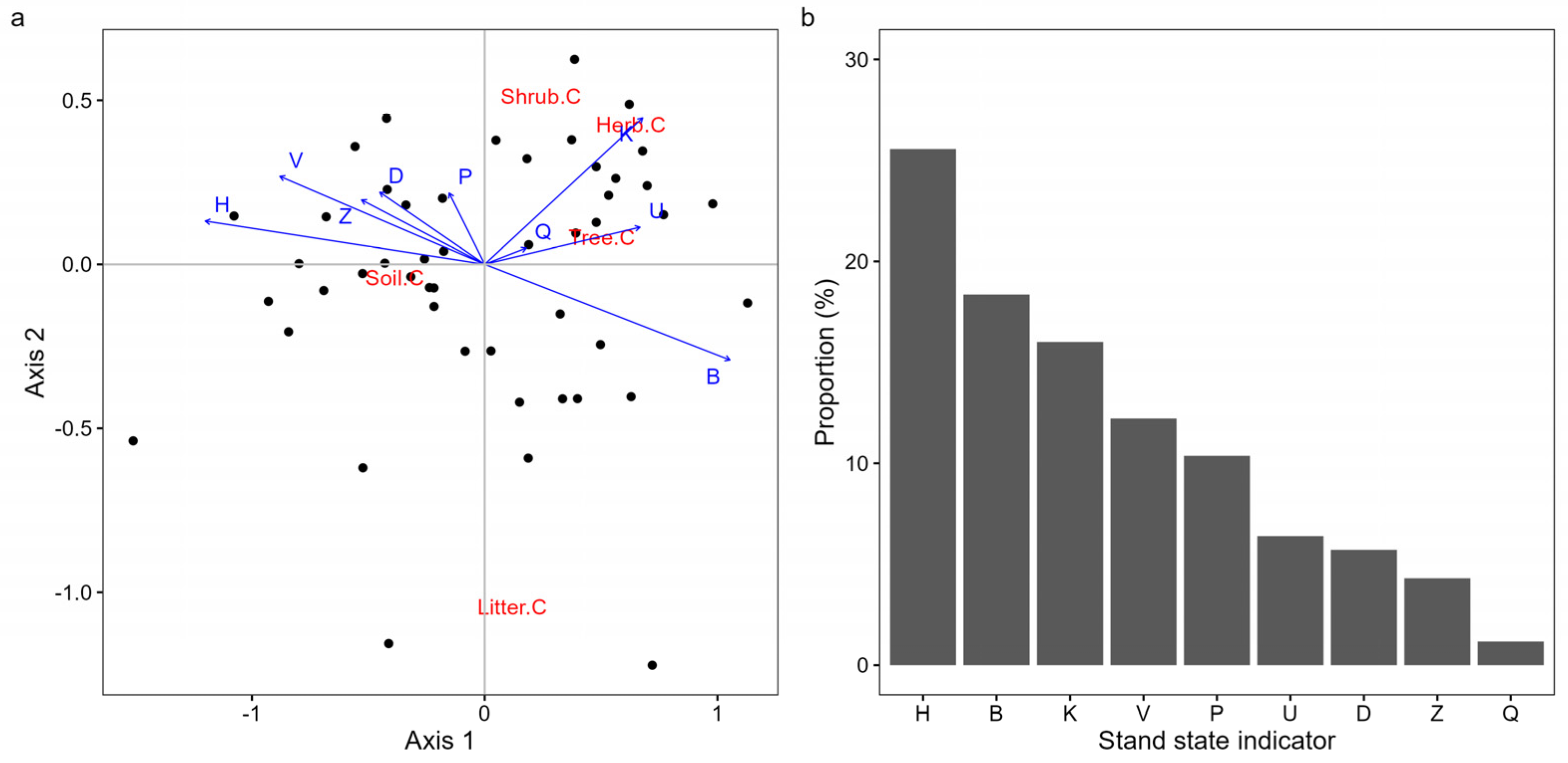
3.3. The Coupling Correlation Between the Stand State and C Storage in P. massoniana Forests
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ruiz–Benito, P.; Madrigal–Gonzalez, J.; Ratcliffe, S.; Coomes, D.A.; Kaendler, G.; Lehtonen, A.; Wirth, C.; Zavala, M.A. Stand Structure and Recent Climate Change Constrain Stand Basal Area Change in European Forests: A Comparison Across Boreal, Temperate, and Mediterranean Biomes. Ecosystems 2014, 17, 1439–1454. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). Global Forest Resources Assessment 2020: Main Report; FAO: Rome, Italy, 2021. [Google Scholar] [CrossRef]
- Ahtikoski, A.; Ahtikoski, R.; Haapanen, M.; Hynynen, J.; Karkkainen, K. Economic Performance of Genetically Improved Reforestation Material in Joint Production of Timber and Carbon Sequestration: A Case Study from Finland. Forests 2020, 11, 847. [Google Scholar] [CrossRef]
- Ouimette, A.P.; Ollinger, S.V.; Lepine, L.C.; Stephens, R.B.; Rowe, R.J.; Vadeboncoeur, M.A.; Tumber–Davila, S.J.; Hobbie, E.A. Accounting for Carbon Flux to Mycorrhizal Fungi May Resolve Discrepancies in Forest Carbon Budgets. Ecosystems 2020, 23, 715–729. [Google Scholar] [CrossRef]
- Straus, H.; Podvinsek, S.; Klopcic, M. Identifying Optimal Forest Management Maximizing Carbon Sequestration in Mountain Forests Impacted by Natural Disturbances: A Case Study in the Alps. Forests 2023, 14, 947. [Google Scholar] [CrossRef]
- Nilsen, P.; Strand, L.T. Carbon stores and fluxes in even- and uneven- aged Norway spruce stands. Silva Fenn. 2013, 47, 1024. [Google Scholar] [CrossRef]
- Sharma, A.; Bohn, K.K.; Jose, S.; Dwivedi, P. Even-Aged vs. Uneven-Aged Silviculture: Implications for Multifunctional Management of Southern Pine Ecosystems. Forests 2016, 7, 86. [Google Scholar] [CrossRef]
- Magnani, F.; Dewar, R.C.; Borghetti, M. Leakage and spillover effects of forest management on carbon storage: Theoretical insights from a simple model. Tellus Ser. B Chem. Phys. Meteorol. 2009, 61, 385–393. [Google Scholar] [CrossRef]
- Tu, J.; Zhao, Z.; Chai, Z. The Short-Term Impact of Logging Intensity on the Stand State of Middle-Aged Masson Pine (Pinus massoniana Lamb.) Plantations. Forests 2025, 16, 183. [Google Scholar] [CrossRef]
- Jagodzinski, A.M.; Dyderski, M.K.; Gesikiewicz, K.; Horodecki, P.; Cysewska, A.; Wierczynska, S.; Maciejczyk, K. How do tree stand parameters affect young Scots pine biomass?—Allometric equations and biomass conversion and expansion factors. For. Ecol. Manag. 2018, 409, 74–83. [Google Scholar] [CrossRef]
- Thom, D.; Keeton, W. Stand structure drives disparities in carbon storage in northern hardwood-conifer forests. For. Ecol. Manag. 2019, 442, 10–20. [Google Scholar] [CrossRef]
- Takagi, K.; Hirayama, K.; Hayashi, M.; Makoto, K.; Okada, K.; Oguma, H.; Saigusa, N. Forest structure explains spatial heterogeneity of decadal carbon dynamics in a cool–temperate forest. Environ. Res. Lett. 2024, 19, 114022. [Google Scholar] [CrossRef]
- Penne, C.; Ahrends, B.; Deurer, M.; Boettcher, J. The impact of the canopy structure on the spatial variability in forest floor carbon stocks. Geoderma 2010, 158, 282–297. [Google Scholar] [CrossRef]
- Feng, Q.; Chen, C.; Qin, L.; He, Y.; Wang, P.; Duan, Y.; Wang, Y.; He, Y. Effects of Different Management Models on Stand Structure and Plant Diversity of Natural Secondary Forests of Quercus Mongolica. Sci. Silvae Sin. 2018, 54, 12–21. [Google Scholar]
- Rota, E.; Caruso, T.; Bargagli, R. Community structure, diversity and spatial organization of enchytraeids in Mediterranean urban holm oak stands. Eur. J. Soil Biol. 2014, 62, 83–91. [Google Scholar] [CrossRef]
- Aussenac, R.; Bergeron, Y.; Ghotsa, M.C.; Gravel, D.; Pilch, K.; Drobyshev, I. Intraspecific variability in growth response to environmental fluctuations modulates the stabilizing effect of species diversity on forest growth. J. Ecol. 2017, 105, 1010–1020. [Google Scholar] [CrossRef]
- Haq, S.M.; Waheed, M.; Khoja, A.A.; Amjad, M.S.; Bussmann, R.W.; Ali, K.; Jones, D.A. Measuring forest health at stand level: A multi–indicator evaluation for use in adaptive management and policy. Ecol. Indic. 2023, 150, 110225. [Google Scholar] [CrossRef]
- Chai, Z.; Zhu, J.; Zhao, Z. Close-to-nature management alleviated microbial P limitation in middle–aged Masson pine plantations: Evidence derived from ecoenzymatic stoichiometry. For. Ecol. Manag. 2025, 580, 122543. [Google Scholar] [CrossRef]
- Fang, X.; Tan, W.; Gao, X.; Chai, Z. Close-to-nature management positively improves the spatial structure of Masson pine forest stands. Web Ecol. 2021, 21, 45–54. [Google Scholar] [CrossRef]
- Chai, Z.; Tan, W. Impacts of close-to-nature management on the stand states of Masson pine forests. Ecosyst. Health Sustain. 2020, 6, 1846461. [Google Scholar] [CrossRef]
- Zou, F.; Chai, Z. Effects of Close-to-Nature Management on Carbon Storage of Pinus massoniana Plantation Ecosystem. Guangxi Sci. 2024, 31, 405–415. [Google Scholar]
- Li, Y.; Mo, Y.; Qin, J.; Ma, J. Carbon Storage and Distribution Pattern of Pinus massoniana Plantations Ecosystem in Different Forest Ages in Guangxi. Guangxi Sci. 2024, 31, 346–356. [Google Scholar]
- Wei, M.; Wang, C.; Yang, Z.; Huang, Z.; Wang, H.; He, B. Carbon storage and distribution of Pinus massoniana plantation ecosystem in northwest Guangxi. Subtrop. Agric. Res. 2019, 15, 152–156. [Google Scholar]
- Zhang, K.; Gao, D.; Guo, H.; Zeng, J.; Liu, X. Forest structure characteristics on soil carbon and nitrogen storage of Pinus massoniana plantations in southern subtropic region. Front. For. Glob. Change 2022, 5, 1022221. [Google Scholar] [CrossRef]
- Hui, G.; Zhang, G.; Zhao, Z.; Yang, A. Methods of Forest Structure Research: A Review. Curr. For. Rep. 2019, 5, 142–154. [Google Scholar] [CrossRef]
- Wei, H.; Zhang, Y.; Dong, L.; Liu, Z.; Chen, Y. Comprehensive evaluation of spatial structure state for the main forest types in Maoershan mountain. J. Cent. South Univ. For. Technol. 2021, 41, 131–139. [Google Scholar]
- Zhang, G.; Hui, G.; Zhang, G.; Hu, Y.; Zhao, Z. A Novel Comprehensive Evaluation Method of Forest State Based on Unit Circle. Forests 2019, 10, 5. [Google Scholar] [CrossRef]
- Zhao, Z.; Hui, G.; Liu, W.; Hu, Y.; Zhang, G. A Novel Method for Calculating Stand Structural Diversity Based on the Relationship of Adjacent Trees. Forests 2022, 13, 343. [Google Scholar] [CrossRef]
- Guizhou Provincial Department of Natural Resources and Planning. Guizhou Province Individual Tree Carbon Sink Project Methodology (No. 201712–V1); Guizhou Provincial Department of Natural Resources and Planning: Guiyang, China, 2017. [Google Scholar]
- Huang, H. Everything Is Linkable, R Package Version 0.0.3. 2021. Available online: https://github.com/Hy4m/linkET (accessed on 1 September 2024).
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package, R Package Version 2.6-5. 2024. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 September 2024).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. Available online: https://ggplot2.tidyverse.org (accessed on 1 September 2024).
- Zhang, C.; Deng, Q.; Liu, A.; Liu, C.; Xie, G. Effects of Stand Structure and Topography on Forest Vegetation Carbon Density in Jiangxi Province. Forests 2021, 12, 1483. [Google Scholar] [CrossRef]
- He, X.; Zhou, C.; Lei, X.; Li, H. Stand carbon stock growth model system for Larix olgensis plantation. J. Beijing For. Univ. 2021, 43, 1–10. [Google Scholar]
- Li, T.; Wu, X.; Wu, Y.; Li, M. Forest Carbon Density Estimation Using Tree Species Diversity and Stand Spatial Structure Indices. Forests 2023, 14, 1105. [Google Scholar] [CrossRef]
- Duan, M.; Wang, G.; Shi, J.; Zhou, H. Long-term Effects of Thinning on Carbon Storage in Pinus Tabulaeformis Plantations. J. Soil Water Conserv. 2018, 32, 190–196. [Google Scholar]
- Liu, X.; Wang, C.; Gao, J.; Yuan, J.; Huang, Y.; Wang, B.; Peng, Y. Approaches to carbon sequestration enhancement in China’s plantation ecosystem for carbon peaking and carbon neutrality goals. Acta Ecol. Sin. 2023, 43, 5662–5673. [Google Scholar]
- Li, X.; Ramos, A.; Wu, D.; Lie, Z.; Xu, W.; Tang, X.; Liu, J. Carbon sequestration and storage capacity of Chinese fir at different stand ages. Sci. Total Environ. 2023, 904, 166962. [Google Scholar] [CrossRef]
- Denis, V. The Role of Biodiversity in Ecosystem Resilience. IOP Conf. Ser. Earth Environ. Sci. 2022, 1072, 012012. [Google Scholar]
- Dymond, C.; Tedder, S.; Spittlehouse, D.; Raymer, B.; Hopkins, K.; McCallion, K.; Sandland, J. Diversifying managed forests to increase resilience. Can. J. For. Res. 2014, 44, 1196–1205. [Google Scholar] [CrossRef]
- Dai, L.; Zhou, L.; Wu, L.; Liu, L.; Huang, Y.; Peng, T.; Qiu, J.; He, Z.; Cao, G. Carbon density and vertical spatial distribution characteristics of Cunninghamia lanceolata forest ecosystem with different stand density. Acta Ecol. Sin. 2022, 42, 710–719. [Google Scholar]
- Hu, H.; Luo, B.; Wei, S.; Wei, S.; Sun, L.; Luo, S.; Ma, H. Biomass carbon density and carbon sequestration capacity in seven typical forest types of the Xiaoxing’an Mountains. Chin. J. Plant Ecol. 2015, 39, 140–158. [Google Scholar]
- Lan, X.; Du, H.; Song, T.; Zeng, F.; Peng, W.; Liu, Y.; Fan, Z.; Zhang, J. Vegetation carbon storage in the main forest types in Guangxi and the related influencing factors. Acta Ecol. Sin. 2019, 39, 2043–2053. [Google Scholar]
- Chen, X.; Taylor, A.; Reich, P.; Hisano, M.; Chen, H.; Chang, S. Tree diversity increases decadal forest soil carbon and nitrogen accrual. Nature 2023, 618, 94–101. [Google Scholar] [CrossRef]
- Zhao, F.; Ouyang, X. Assessing relative contributions of various influencing factors to soil organic carbon in aerially–seeded Pinus massoniana plantations. Acta Ecol. Sin. 2016, 36, 2637–2645. [Google Scholar]
- Güner, Ş.; Güner, D. Changes in carbon stocks of soil and forest floor in black pine plantations in Turkey. J. For. Res. 2020, 32, 339–347. [Google Scholar] [CrossRef]
- Murdiyarso, D.; Sasmito, S.D.; Sillanpää, M.; MacKenzie, R.; Gaveau, D. Mangrove selective logging sustains biomass carbon recovery, soil carbon, and sediment. Sci. Rep. 2021, 11, 12325. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicators | Min | Max | Max−Min | Mean | SD | CV (%) |
|---|---|---|---|---|---|---|
| Stand density (treeshm−2) | 435 | 1965 | 1530 | 987.188 | 387.337 | 39.24 |
| Average tree height (m) | 11.240 | 21.473 | 10.233 | 15.738 | 2.634 | 16.74 |
| Average DBH (cm) | 11.843 | 27.939 | 16.096 | 18.905 | 4.313 | 22.81 |
| Shrub height (m) | 0.094 | 1.347 | 1.253 | 0.528 | 0.277 | 52.53 |
| Shrub cover (%) | 0.313 | 34.450 | 34.137 | 13.100 | 8.126 | 62.03 |
| Herb height (m) | 0.069 | 1.167 | 1.098 | 0.408 | 0.214 | 52.36 |
| Herb cover (%) | 0.242 | 38.521 | 38.279 | 17.317 | 8.854 | 51.13 |
| Item | Indicators | Min | Max | Max−Min | Mean | SD | CV (%) |
|---|---|---|---|---|---|---|---|
| Stand state | DBH distribution (H) | 0.130 | 0.605 | 0.475 | 0.207 | 0.096 | 46.25 |
| Tree height distribution (V) | 0.102 | 0.304 | 0.202 | 0.198 | 0.041 | 20.95 | |
| Stand density (K) | 0.735 | 1.000 | 0.265 | 0.963 | 0.059 | 6.09 | |
| Stand dominance (U) | 0.009 | 0.690 | 0.681 | 0.252 | 0.158 | 62.66 | |
| Stand growth (B) | 0.522 | 0.740 | 0.218 | 0.610 | 0.046 | 7.53 | |
| Stand health (Q) | 0.412 | 1.000 | 0.588 | 0.816 | 0.159 | 19.54 | |
| Species composition (Z) | 0.037 | 0.732 | 0.695 | 0.298 | 0.171 | 57.54 | |
| Species diversity (D) | 0.033 | 0.737 | 0.704 | 0.330 | 0.195 | 59.03 | |
| Species evenness (P) | 0.053 | 0.351 | 0.298 | 0.189 | 0.080 | 42.26 | |
| Carbon storage (t) | Tree | 1.297 | 8.705 | 7.408 | 5.385 | 1.647 | 30.58 |
| Shrub | 0.010 | 0.058 | 0.048 | 0.033 | 0.014 | 41.25 | |
| Herb | 0.006 | 0.070 | 0.064 | 0.032 | 0.018 | 57.74 | |
| Litter | 0.120 | 0.629 | 0.509 | 0.290 | 0.093 | 31.96 | |
| Soil (0–20 cm) | 2.291 | 6.251 | 3.960 | 3.840 | 0.954 | 24.85 | |
| Soil (20–40 cm) | 1.378 | 5.282 | 3.904 | 2.203 | 0.694 | 31.51 | |
| Soil (40–60 cm) | 0.736 | 3.788 | 3.052 | 1.679 | 0.615 | 36.63 | |
| Soil (0–60 cm) | 4.677 | 14.760 | 10.083 | 7.723 | 2.024 | 26.21 | |
| Total | 9.880 | 19.283 | 9.403 | 13.462 | 2.382 | 17.69 |
| Stand Variables | CCA1 | CCA2 | R2 | p |
|---|---|---|---|---|
| DBH distribution (H) | −0.9851 | 0.1719 | 0.1898 | 0.020 * |
| Tree height distribution (V) | −0.9421 | 0.3354 | 0.1265 | 0.054 |
| Stand density (K) | 0.8329 | 0.5535 | 0.1041 | 0.072 |
| Stand dominance (U) | 0.9944 | 0.1053 | 0.0560 | 0.269 |
| Stand growth (B) | 0.9493 | −0.3143 | 0.1748 | 0.018 * |
| Stand health (Q) | 0.9766 | 0.2152 | 0.0043 | 0.908 |
| Species composition (Z) | −0.9234 | 0.3838 | 0.0499 | 0.322 |
| Species diversity (D) | −0.8883 | 0.4592 | 0.0431 | 0.374 |
| Species evenness (P) | −0.6341 | 0.7733 | 0.0179 | 0.667 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Wen, W.; Chai, Z. Stand States Drive Disparities in the Carbon Storage Within a Masson Pine Forest Ecosystem. Forests 2025, 16, 499. https://doi.org/10.3390/f16030499
Hu J, Wen W, Chai Z. Stand States Drive Disparities in the Carbon Storage Within a Masson Pine Forest Ecosystem. Forests. 2025; 16(3):499. https://doi.org/10.3390/f16030499
Chicago/Turabian StyleHu, Jiamin, Weihua Wen, and Zongzheng Chai. 2025. "Stand States Drive Disparities in the Carbon Storage Within a Masson Pine Forest Ecosystem" Forests 16, no. 3: 499. https://doi.org/10.3390/f16030499
APA StyleHu, J., Wen, W., & Chai, Z. (2025). Stand States Drive Disparities in the Carbon Storage Within a Masson Pine Forest Ecosystem. Forests, 16(3), 499. https://doi.org/10.3390/f16030499