
Trade-Off Between Leaf Mass Area and Phosphorus Concentration Alters Resource-Use Strategy of Understory Plants Under Long-Term Nitrogen and Phosphorus Addition in a Subtropical Forest


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Fertilization Experiments
2.2. Plant Sample Collection and Measurement
2.3. Statistical Analysis
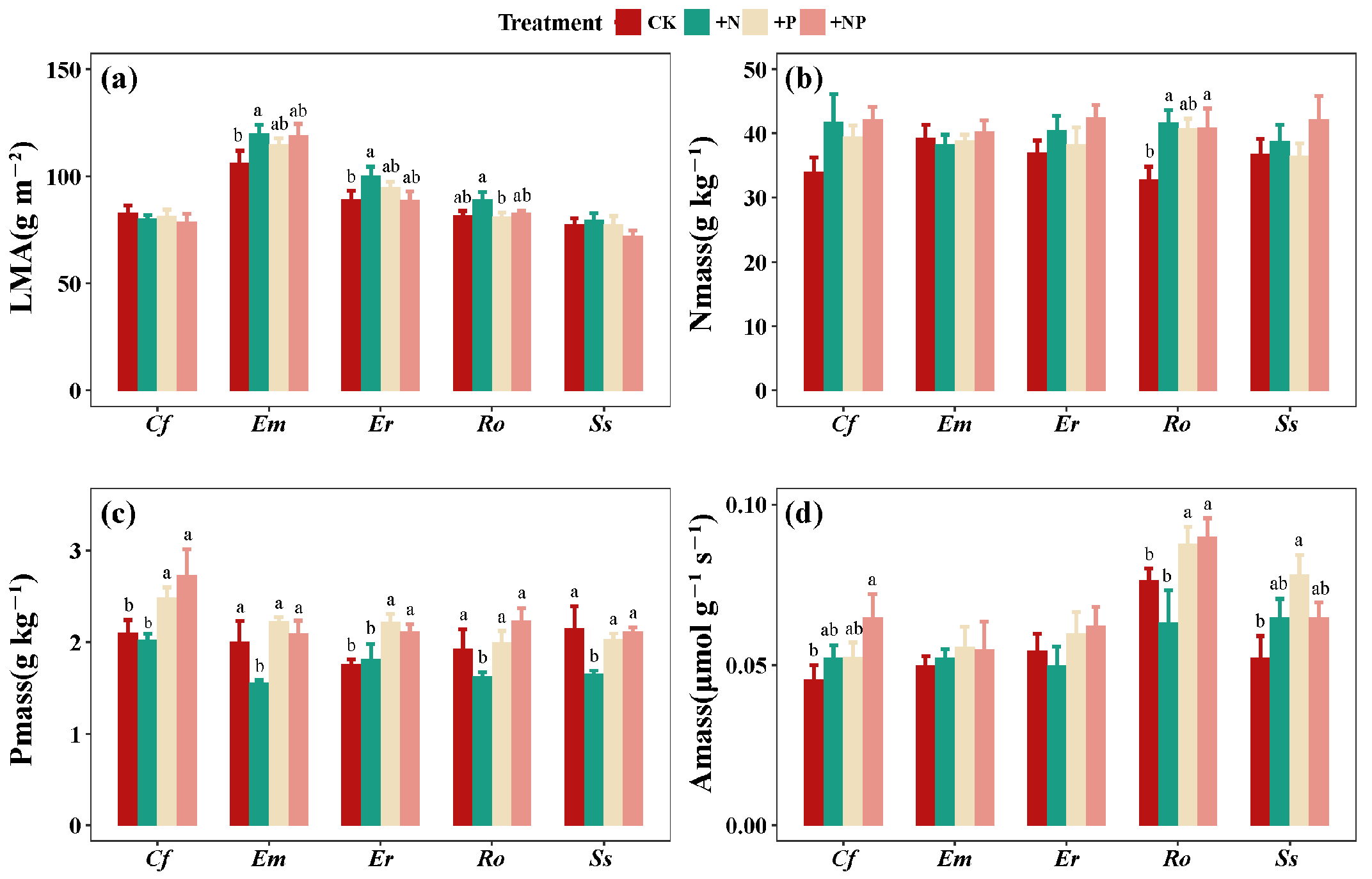
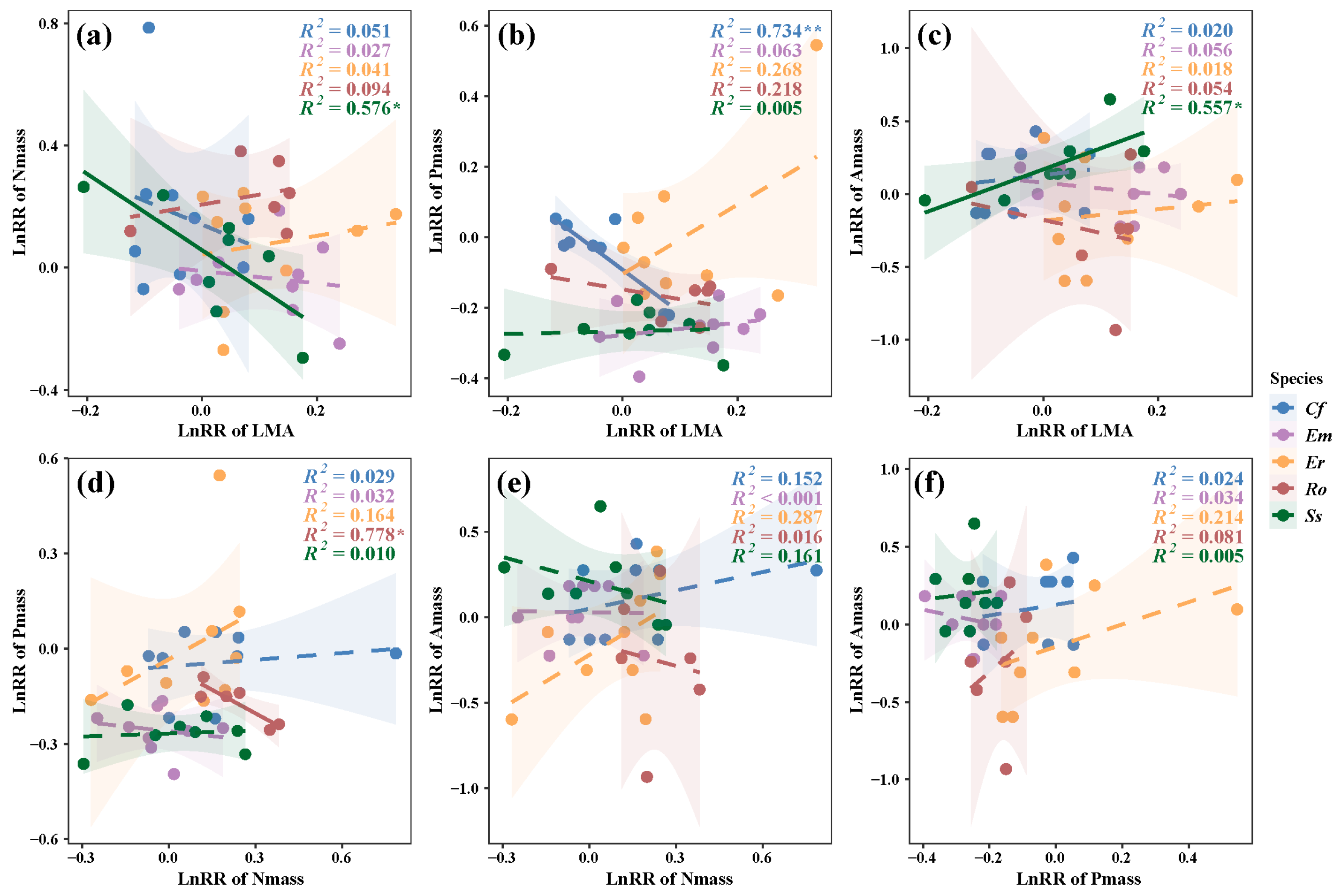
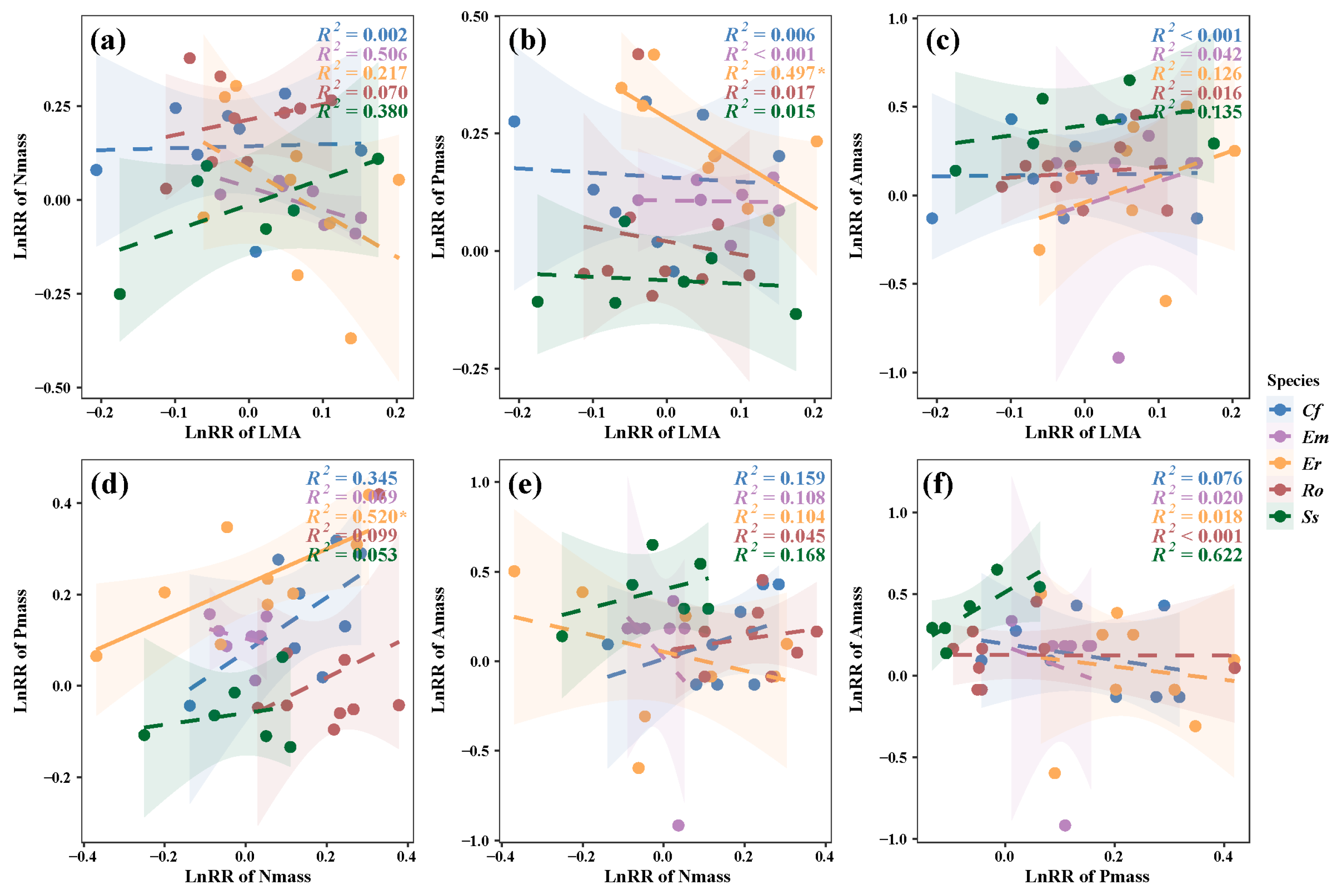
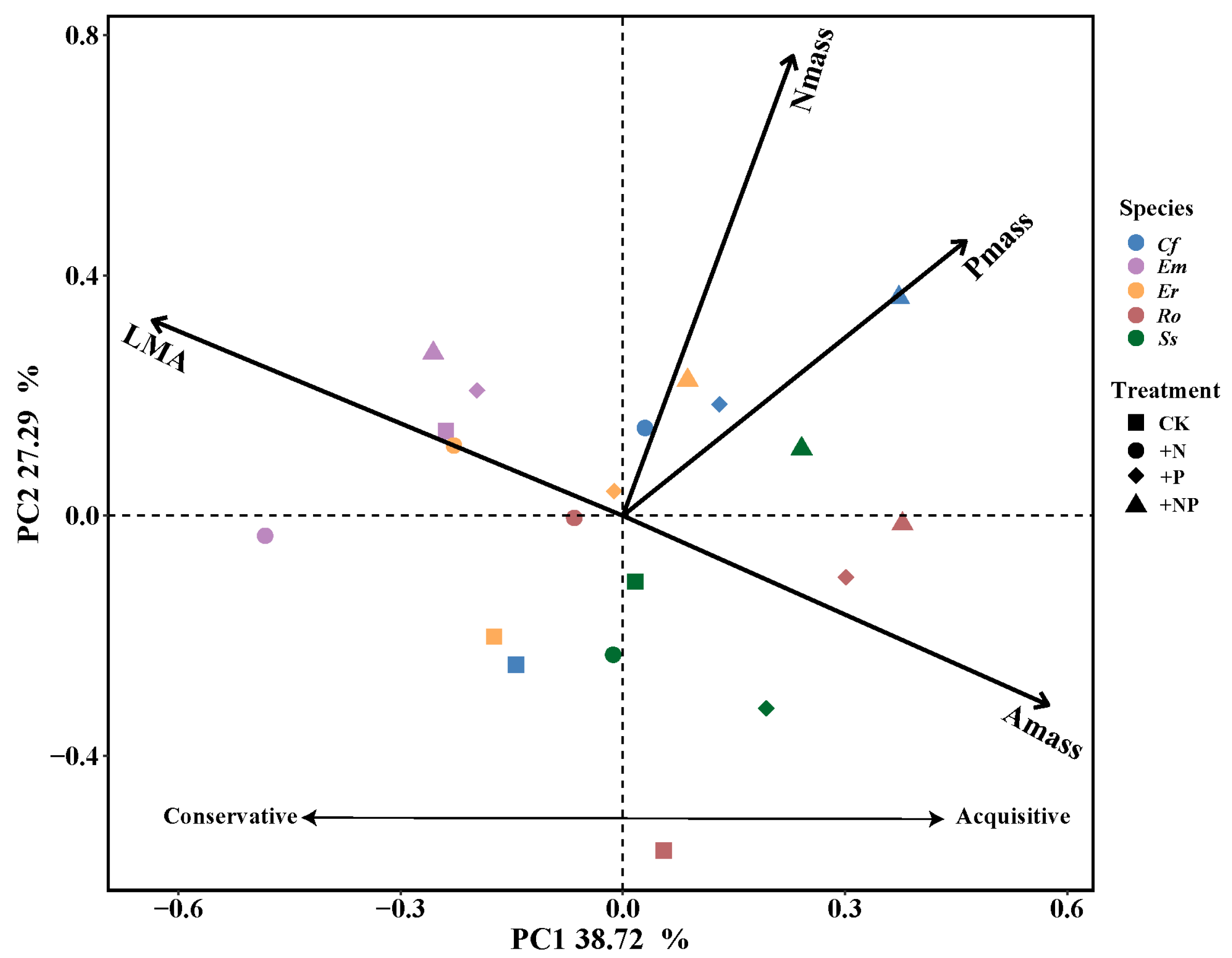
3. Results
3.1. Effects of N and P Additions on LES Traits of Understory Plants
3.2. Relationships Between LES Trait Responses and N and P Additions
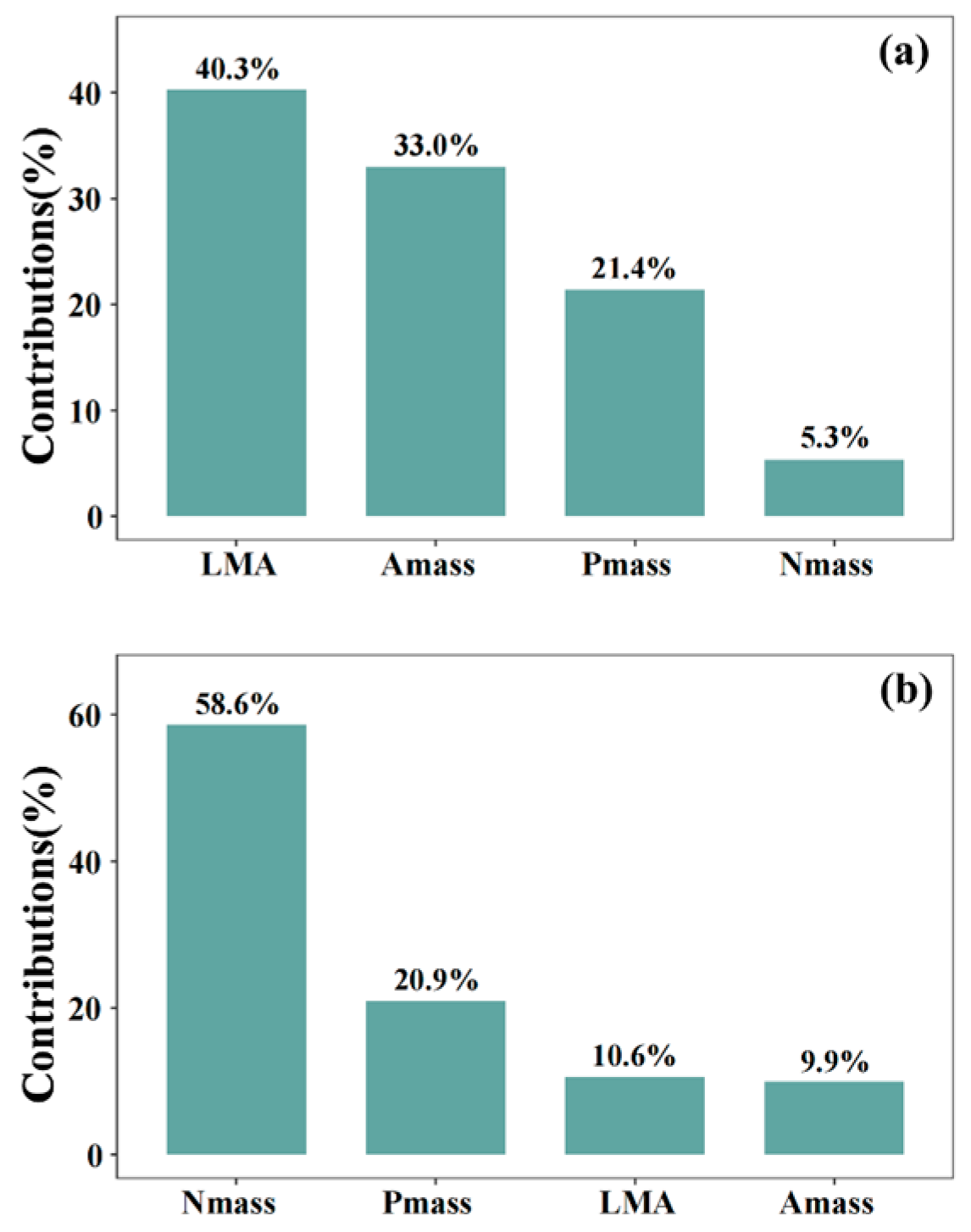
3.3. Shifts in LES Position and Plant Resource-Use Strategies Under N and P Additions
4. Discussions
4.1. Fertilization Alters Plant LES Traits and Their Relationships
4.2. Plants Adjust Resource-Use Strategies Through Trait Changes After Fertilization
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ackerman, D.; Millet, D.B.; Chen, X. Global Estimates of Inorganic Nitrogen Deposition across Four Decades. Glob. Biogeochem. Cycles 2019, 33, 100–107. [Google Scholar] [CrossRef]
- Pan, Y.; Liu, B.; Cao, J.; Liu, J.; Tian, S.; Du, E. Enhanced Atmospheric Phosphorus Deposition in Asia and Europe in the Past Two Decades. Atmos. Ocean. Sci. Lett. 2021, 14, 100051. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The Worldwide Leaf Economics Spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Poorter, L.; Bongers, F. Leaf Traits Are Good Predictors of Plant Performance Across 53 Rain Forest Species. Ecology 2006, 87, 1733–1743. [Google Scholar] [CrossRef]
- Fan, Y.; Lambers, H.; Sayer, E.J.; Li, Y.; Li, Y.; Zhang, J.; Li, Z.; Hu, Z.; Li, H.; Wang, J.; et al. Variation in Leaf Phosphorus Fractions Reflects Plant Adaptations and Distribution in Low-Phosphorus Tropical Forests. Funct. Ecol. 2025, 39, 621–634. [Google Scholar] [CrossRef]
- Zhang, X.; Li, B.; Peñuelas, J.; Sardans, J.; Cheng, D.; Yu, H.; Zhong, Q. Resource-Acquisitive Species Have Greater Plasticity in Leaf Functional Traits than Resource-Conservative Species in Response to Nitrogen Addition in Subtropical China. Sci. Total Environ. 2023, 903, 166177. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B. The World-Wide ‘Fast–Slow’ Plant Economics Spectrum: A Traits Manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Ye, X.; Bu, W.; Hu, X.; Wang, F.; Sun, R.; He, P.; Liang, X.; Chen, F. Are Small Trees More Responsive to Nutrient Addition than Large Trees in an Evergreen Broadleaved Forest? For. Ecol. Manag. 2023, 543, 121129. [Google Scholar] [CrossRef]
- Mo, Q.; Wang, W.; Chen, Y.; Peng, Z.; Zhou, Q. Response of Foliar Functional Traits to Experimental N and P Addition among Overstory and Understory Species in a Tropical Secondary Forest. Glob. Ecol. Conserv. 2020, 23, e01109. [Google Scholar] [CrossRef]
- Su, Y.; Tang, Y.; Hu, Y.; Liu, M.; Lu, X.; Duan, B. Exploring Plant Adaptation Strategies to Phosphorus Limitation Induced by Nitrogen Addition: Foliar Phosphorus Allocation and Root Functional Traits Analysis in Two Dominant Subalpine Tree Species. J. Plant Ecol. 2024, 17, rtae060. [Google Scholar] [CrossRef]
- Freschet, G.T.; Cornelissen, J.H.C.; Van Logtestijn, R.S.P.; Aerts, R. Evidence of the ‘Plant Economics Spectrum’ in a Subarctic Flora. J. Ecol. 2010, 98, 362–373. [Google Scholar] [CrossRef]
- Zhao, N.; Yu, G.; Wang, Q.; Wang, R.; Zhang, J.; Liu, C.; He, N. Conservative Allocation Strategy of Multiple Nutrients among Major Plant Organs: From Species to Community. J. Ecol. 2020, 108, 267–278. [Google Scholar] [CrossRef]
- Zhao, C.; Lin, Q.; Tian, D.; Ji, C.; Shen, H.; Fan, D.; Wang, X.; Fang, J. Nitrogen Addition Promotes Conservative Resource-Use Strategies via Aggravating Phosphorus Limitation of Evergreen Trees in Subtropical Forest. Sci. Total Environ. 2023, 889, 164047. [Google Scholar] [CrossRef]
- Ye, X.; Bu, W.; Hu, X.; Liu, B.; Liang, K.; Chen, F. Species Divergence in Seedling Leaf Traits and Tree Growth Response to Nitrogen and Phosphorus Additions in an Evergreen Broadleaved Forest of Subtropical China. J. For. Res. 2023, 34, 137–150. [Google Scholar] [CrossRef]
- Shi, X.-M.; Qi, J.-H.; Liu, A.-X.; Zakari, S.; Song, L. Leaf Phenotypic Plasticity Coupled with Integration Facilitates the Adaptation of Plants to Enhanced N Deposition. Environ. Pollut. 2023, 327, 121570. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Zhang, L.; Lambers, H.; Ren, W.; Lu, X.; Hou, E.; Fu, S.; Kuang, Y. Addition of Nitrogen to Canopy versus Understorey Has Different Effects on Leaf Traits of Understorey Plants in a Subtropical Evergreen Broad-Leaved Forest. J. Ecol. 2021, 109, 692–702. [Google Scholar] [CrossRef]
- Lambers, H.; Raven, J.A.; Shaver, G.R.; Smith, S.E. Plant Nutrient-Acquisition Strategies Change with Soil Age. Trends Ecol. Evol. 2008, 23, 95–103. [Google Scholar] [CrossRef]
- Scheller, R.M.; Mladenoff, D.J. Understory Species Patterns and Diversity in Old-Growth and Managed Northern Hardwood Forests. Ecol. Appl. 2002, 12, 1329–1343. [Google Scholar] [CrossRef]
- Mao, Q.; Lu, X.; Wang, C.; Zhou, K.; Mo, J. Responses of Understory Plant Physiological Traits to a Decade of Nitrogen Addition in a Tropical Reforested Ecosystem. For. Ecol. Manag. 2017, 401, 65–74. [Google Scholar] [CrossRef]
- Turner, B.L.; Brenes-Arguedas, T.; Condit, R. Pervasive Phosphorus Limitation of Tree Species but Not Communities in Tropical Forests. Nature 2018, 555, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Sun, J.; Zarif, N.; Khan, K.; Jamil, M.A.; Yang, L.; Clothier, B.; Rewald, B. Effects of Increased N Deposition on Leaf Functional Traits of Four Contrasting Tree Species in Northeast China. Plants 2020, 9, 1231. [Google Scholar] [CrossRef]
- Cárate-Tandalla, D.; Camenzind, T.; Leuschner, C.; Homeier, J. Contrasting Species Responses to Continued Nitrogen and Phosphorus Addition in Tropical Montane Forest Tree Seedlings. Biotropica 2018, 50, 234–245. [Google Scholar] [CrossRef]
- Zheng, Z.; Mamuti, M.; Liu, H.; Shu, Y.; Hu, S.; Wang, X.; Li, B.; Lin, L.; Li, X. Effects of Nutrient Additions on Litter Decomposition Regulated by Phosphorus-Induced Changes in Litter Chemistry in a Subtropical Forest, China. For. Ecol. Manag. 2017, 400, 123–128. [Google Scholar] [CrossRef]
- Gao, Q.; Hasselquist, N.J.; Palmroth, S.; Zheng, Z.; You, W. Short-Term Response of Soil Respiration to Nitrogen Fertilization in a Subtropical Evergreen Forest. Soil Biol. Biochem. 2014, 76, 297–300. [Google Scholar] [CrossRef]
- Zheng, Z.; Lu, J.; Su, Y.; Yang, Q.; Lin, Y.; Liu, H.; Yang, J.; Huang, H.; Wang, X. Differential Effects of N and P Additions on Foliar Stoichiometry between Species and Community Levels in a Subtropical Forest in Eastern China. Ecol. Indic. 2020, 117, 106537. [Google Scholar] [CrossRef]
- Song, Y. Vegetation Ecology, 2nd ed.; Higher Education Press: Beijing, China, 2017; ISBN 978-7-04-046159-6. [Google Scholar]
- Hedges, L.V.; Gurevitch, J.; Curtis, P.S. The Meta-Analysis of Response Ratios in Experimental Ecology. Ecology 1999, 80, 1150–1156. [Google Scholar] [CrossRef]
- Marklein, A.R.; Houlton, B.Z. Nitrogen Inputs Accelerate Phosphorus Cycling Rates across a Wide Variety of Terrestrial Ecosystems. New Phytol. 2012, 193, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Bai, T.; Liu, Y.-Y.; Muhammad, I.; Yang, X.; Yin, X.-J.; Bai, L.; Wang, Y.-J. Mixed Nitrogen Form Addition Facilitates the Growth Adaptation of Legume Plant to Heavy Metal Contamination in Degraded Mining Areas. Glob. Ecol. Conserv. 2020, 24, e01387. [Google Scholar] [CrossRef]
- Cernusak, L.A.; Winter, K.; Turner, B.L. Leaf Nitrogen to Phosphorus Ratios of Tropical Trees: Experimental Assessment of Physiological and Environmental Controls. New Phytol. 2010, 185, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Cui, E.; Lu, R.; Xu, X.; Sun, H.; Qiao, Y.; Ping, J.; Qiu, S.; Lin, Y.; Bao, J.; Yong, Y.; et al. Soil Phosphorus Drives Plant Trait Variations in a Mature Subtropical Forest. Glob. Change Biol. 2022, 28, 3310–3320. [Google Scholar] [CrossRef] [PubMed]
- Westoby, M.; Falster, D.S.; Moles, A.T.; Vesk, P.A.; Wright, I.J. Plant Ecological Strategies: Some Leading Dimensions of Variation Between Species. Annu. Rev. Ecol. Evol. Syst. 2002, 33, 125–159. [Google Scholar] [CrossRef]
- Cui, E.; Weng, E.; Yan, E.; Xia, J. Robust Leaf Trait Relationships across Species under Global Environmental Changes. Nat. Commun. 2020, 11, 2999. [Google Scholar] [CrossRef] [PubMed]
- Griffith, D.M.; Quigley, K.M.; Anderson, T.M. Leaf Thickness Controls Variation in Leaf Mass per Area (LMA) among Grazing-Adapted Grasses in Serengeti. Oecologia 2016, 181, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, X.; Niklas, K.J.; Sun, J.; Wang, Z.; Zhong, Q.; Hu, D.; Cheng, D. A Whole-Plant Economics Spectrum Including Bark Functional Traits for 59 Subtropical Woody Plant Species. J. Ecol. 2022, 110, 248–261. [Google Scholar] [CrossRef]
- Zhang, Q.; Hao, G.; Li, M.; Li, L.; Kang, B.; Yang, N.; Li, H. Transformation of Plant to Resource Acquisition Under High Nitrogen Addition Will Reduce Green Roof Ecosystem Functioning. Front. Plant Sci. 2022, 13, 894782. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Abbreviation | Family | Diameter at the Breast Height (DBH, cm) | Tree Density (Tree hm−2) | Important Value |
|---|---|---|---|---|---|
| Camellia fraternal | Cf | Theaceae | 2.1 | 575 | 0.28 |
| Eurya muricata | Em | Theaceae | 3.4 | 225 | 0.13 |
| Eurya rubiginosa | Er | Theaceae | 2.1 | 250 | 0.20 |
| Rhododendron ovatum | Ro | Ericaceae | 2.5 | 500 | 0.20 |
| Symplocos sumuntia | Ss | Symplocaceae | 2.1 | 450 | 0.22 |
| Variable | df | LMA | Nmass | Pmass | Amass |
|---|---|---|---|---|---|
| Fertilization | 3 | 3.167 * | 3.705 * | 8.589 ** | 4.290 * |
| Species | 4 | 79.865 ** | 0.098 | 2.920 * | 9.666 ** |
| Fertilization × Species | 12 | 1.287 | 0.593 | 0.639 | 0.924 |
| Treatments | Ranking | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| CK | Em | Er | Cf | Ss | Ro |
| +N | Em | Er | Ro | Ss | Cf |
| +P | Em | Er | Cf | Ss | Ro |
| +NP | Em | Er | Ss | Cf | Ro |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yong, Y.; Huang, P.; Xu, S.; Yang, X.; Bao, J.; Zheng, Z. Trade-Off Between Leaf Mass Area and Phosphorus Concentration Alters Resource-Use Strategy of Understory Plants Under Long-Term Nitrogen and Phosphorus Addition in a Subtropical Forest. Forests 2025, 16, 319. https://doi.org/10.3390/f16020319
Yong Y, Huang P, Xu S, Yang X, Bao J, Zheng Z. Trade-Off Between Leaf Mass Area and Phosphorus Concentration Alters Resource-Use Strategy of Understory Plants Under Long-Term Nitrogen and Phosphorus Addition in a Subtropical Forest. Forests. 2025; 16(2):319. https://doi.org/10.3390/f16020319
Chicago/Turabian StyleYong, Yutong, Pei Huang, Siyi Xu, Xiongfei Yang, Jiehuan Bao, and Zemei Zheng. 2025. "Trade-Off Between Leaf Mass Area and Phosphorus Concentration Alters Resource-Use Strategy of Understory Plants Under Long-Term Nitrogen and Phosphorus Addition in a Subtropical Forest" Forests 16, no. 2: 319. https://doi.org/10.3390/f16020319
APA StyleYong, Y., Huang, P., Xu, S., Yang, X., Bao, J., & Zheng, Z. (2025). Trade-Off Between Leaf Mass Area and Phosphorus Concentration Alters Resource-Use Strategy of Understory Plants Under Long-Term Nitrogen and Phosphorus Addition in a Subtropical Forest. Forests, 16(2), 319. https://doi.org/10.3390/f16020319