
Roles of Spatial Distance, Habitat Difference, and Community Age on Plant Diversity Patterns of Fragmented Castanopsis orthacantha Franch. Forests in Central Yunnan, Southwest China


Abstract
1. Introduction
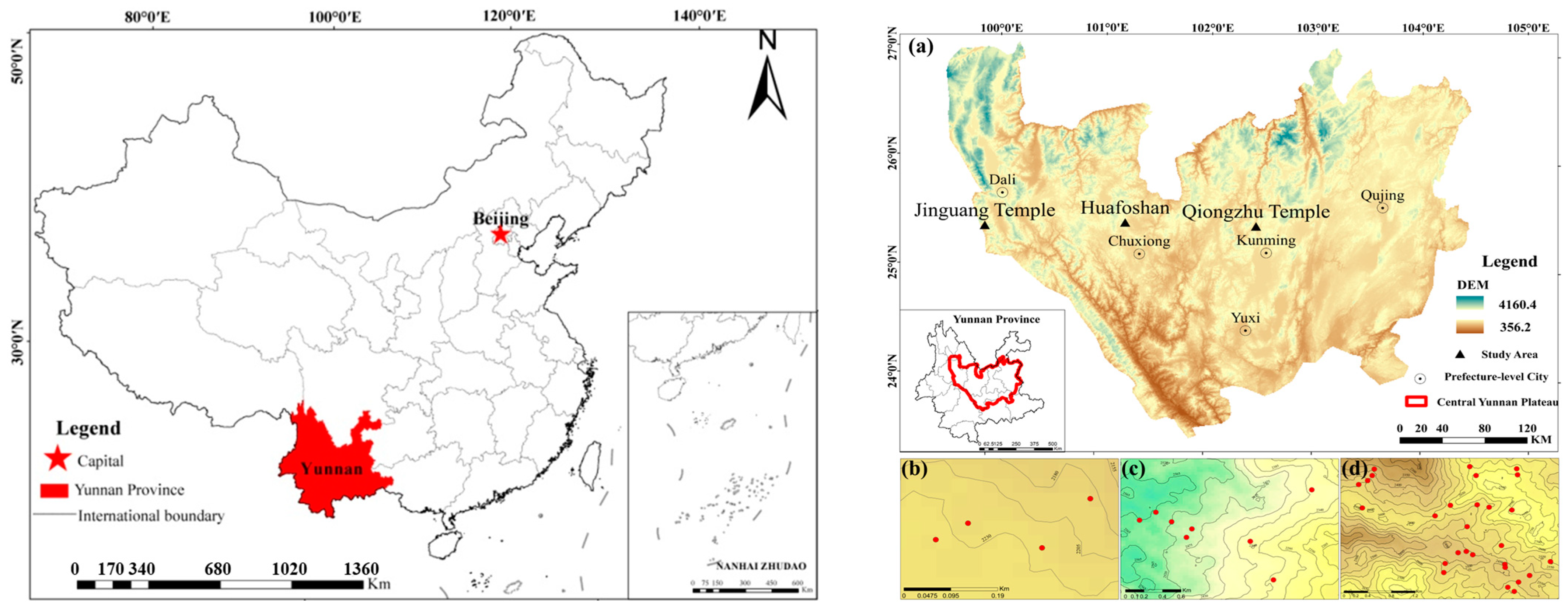
2. Materials and Methods
3. Results
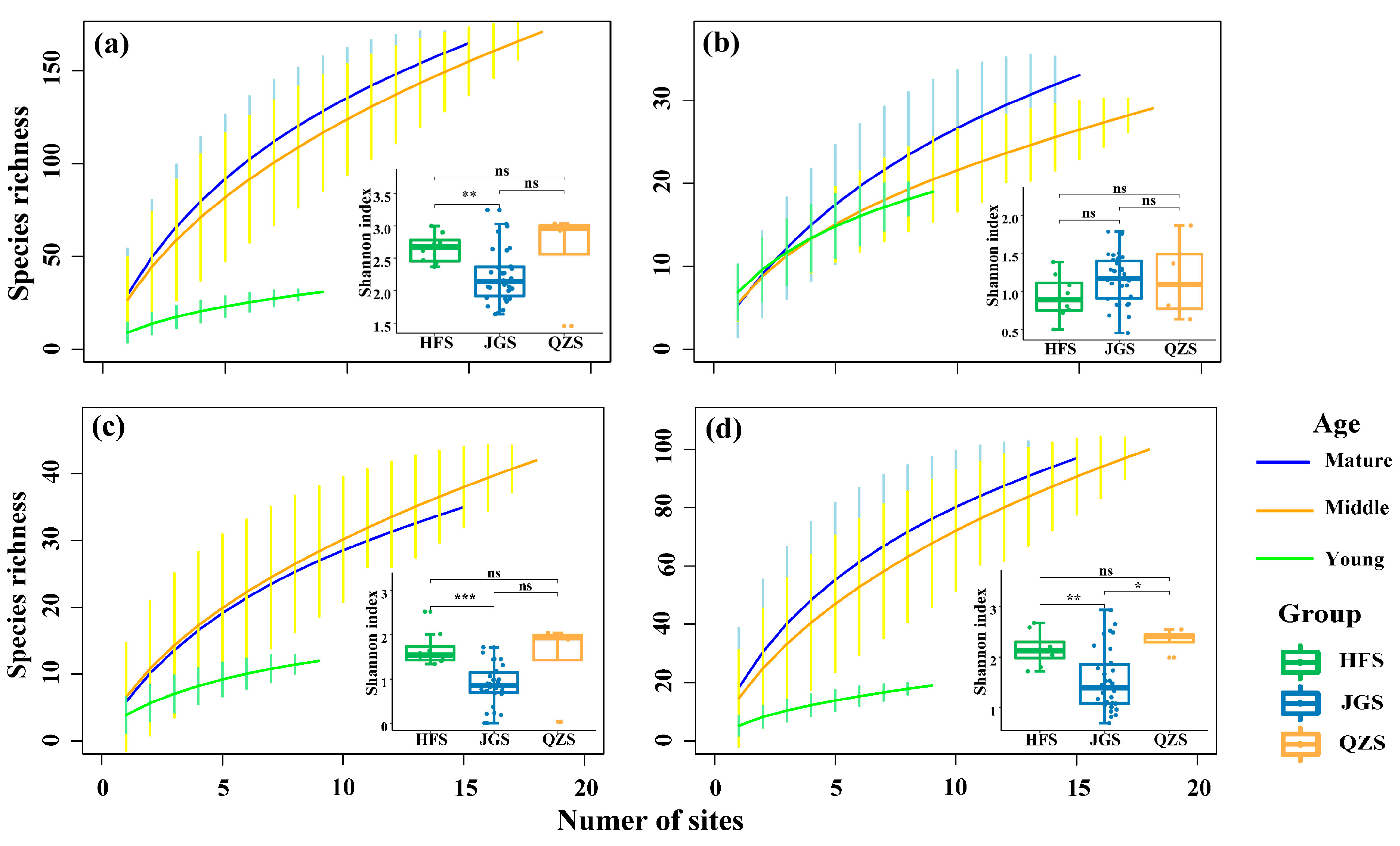
3.1. Plant Species Richness of Castanopsis Orthacantha Forests Across Sites and Age Classes
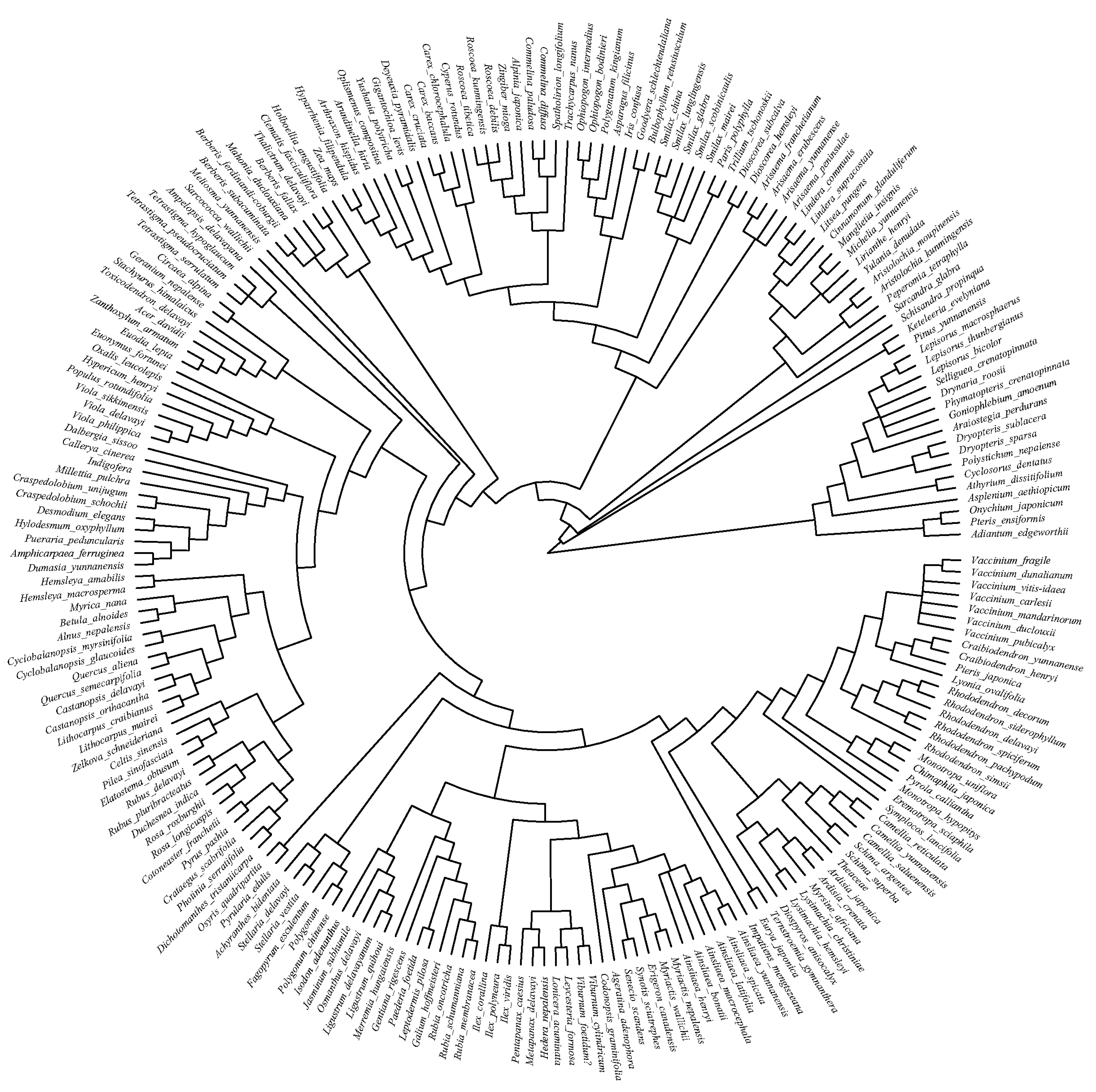
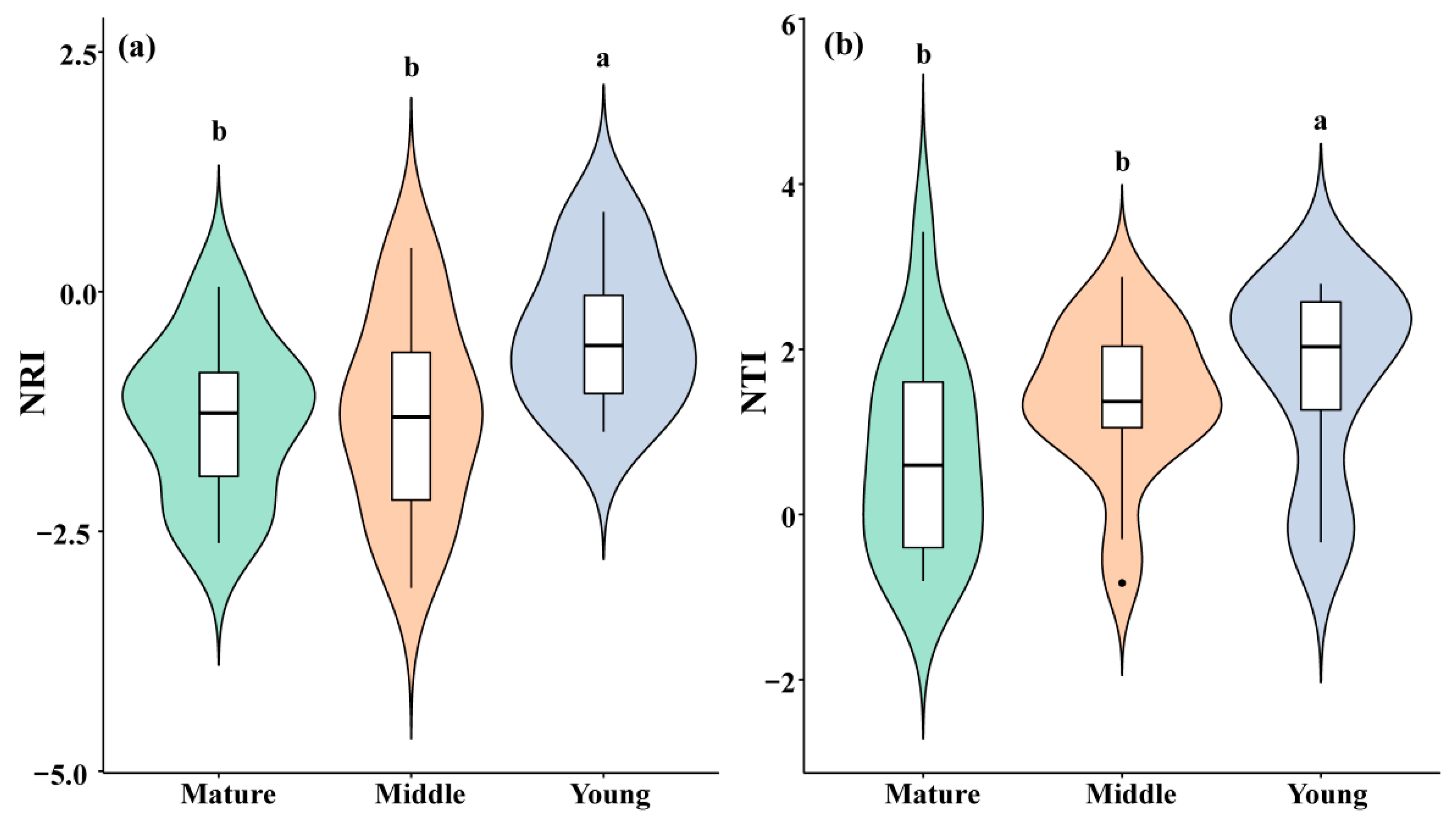
3.2. Phylogenetic Structure of Communities of Different Age Classes
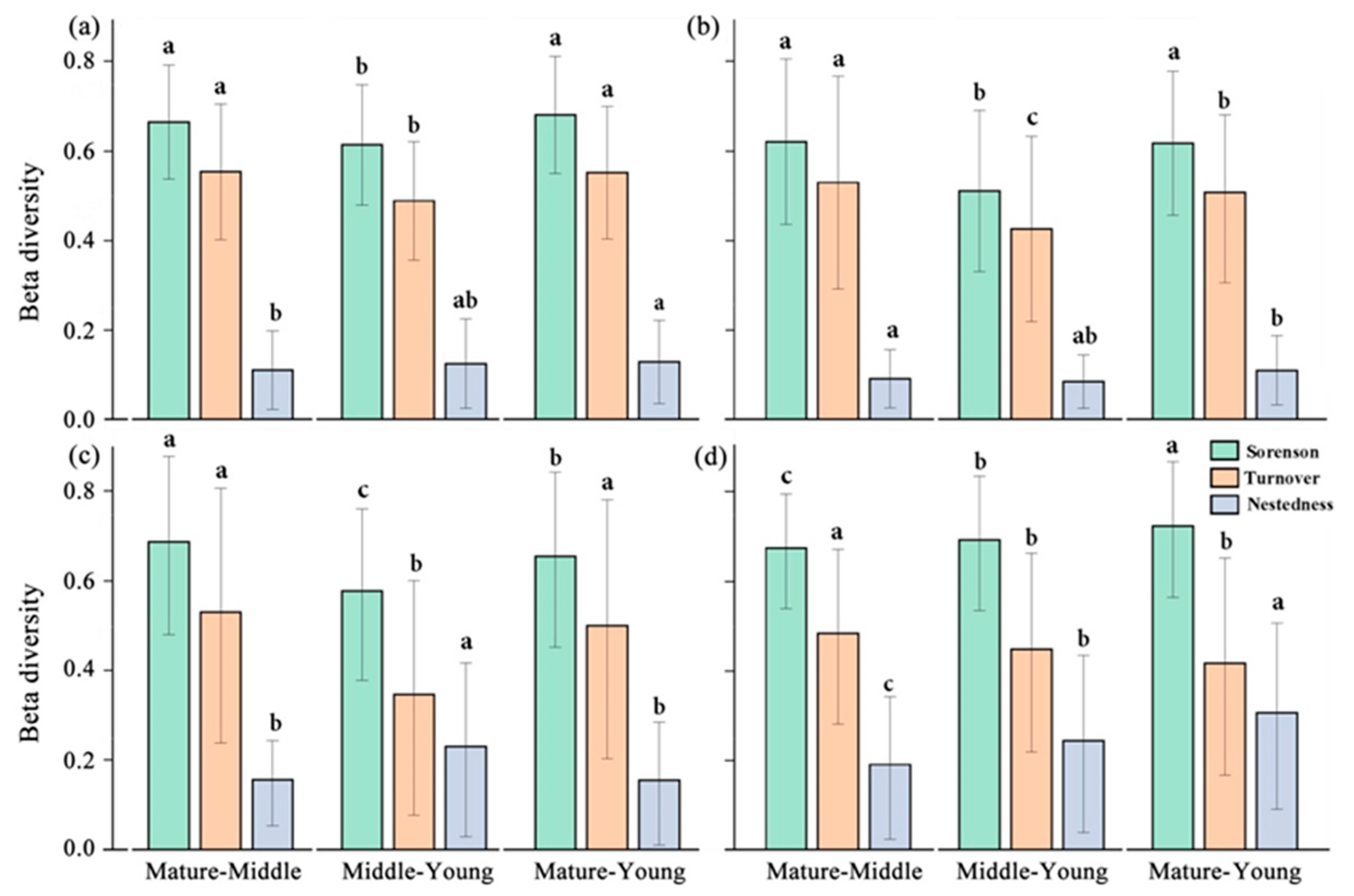
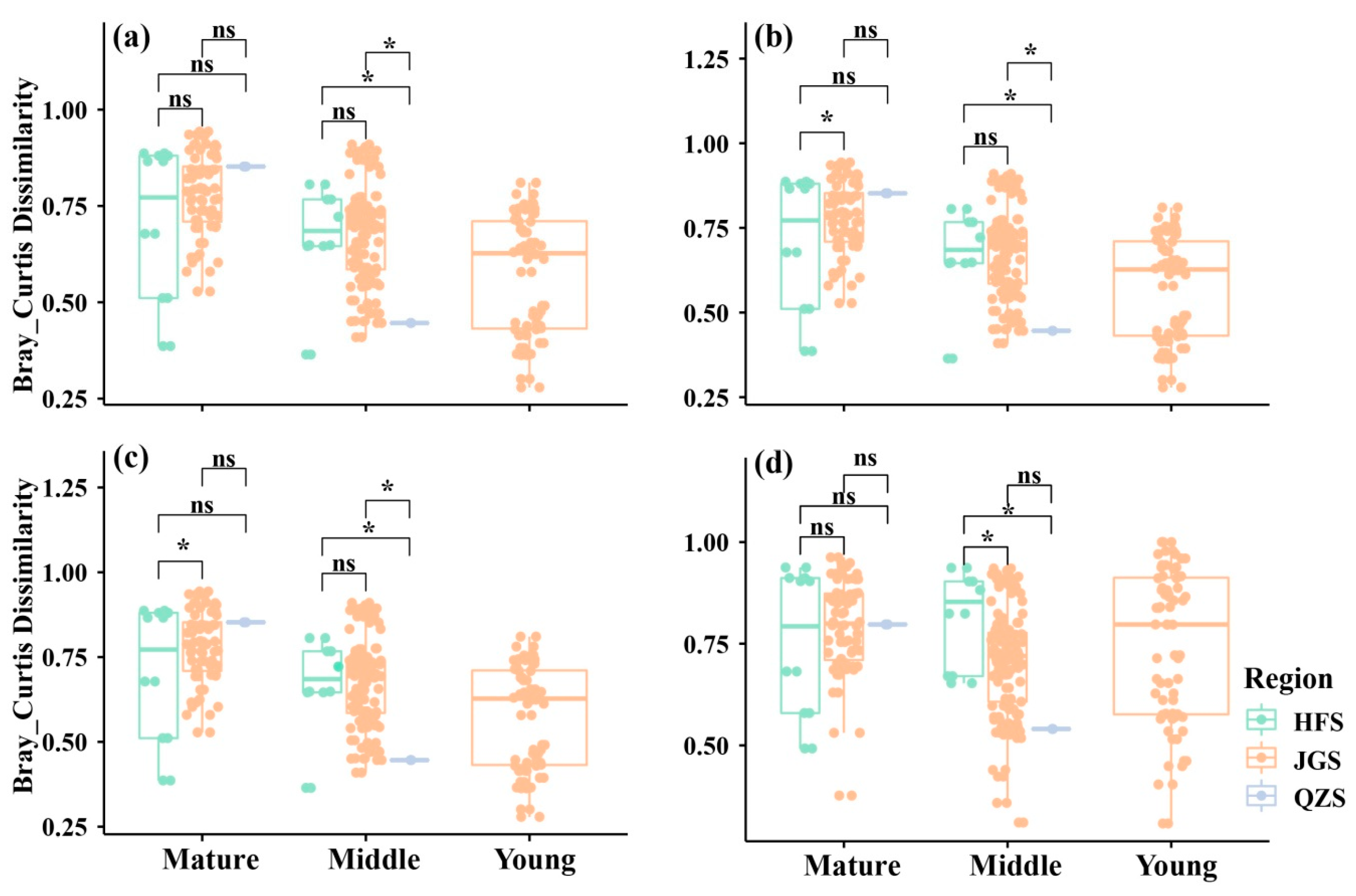
3.3. Temporal and Spatial Patterns of Community Beta Diversity and Its Components
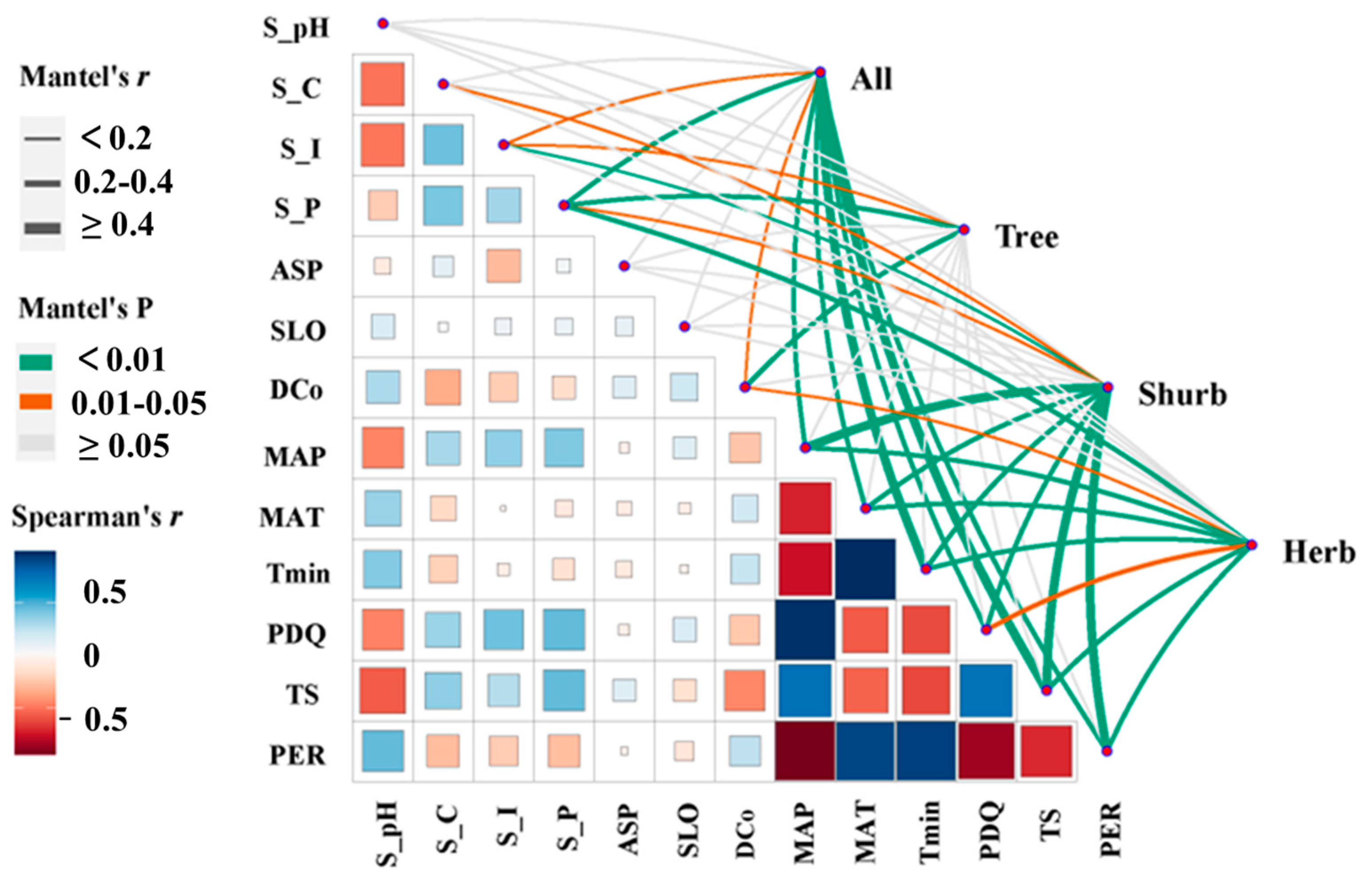
3.4. Driving Factors of Forest Beta Diversity Patterns at Tree, Shrub, and Herb Layers
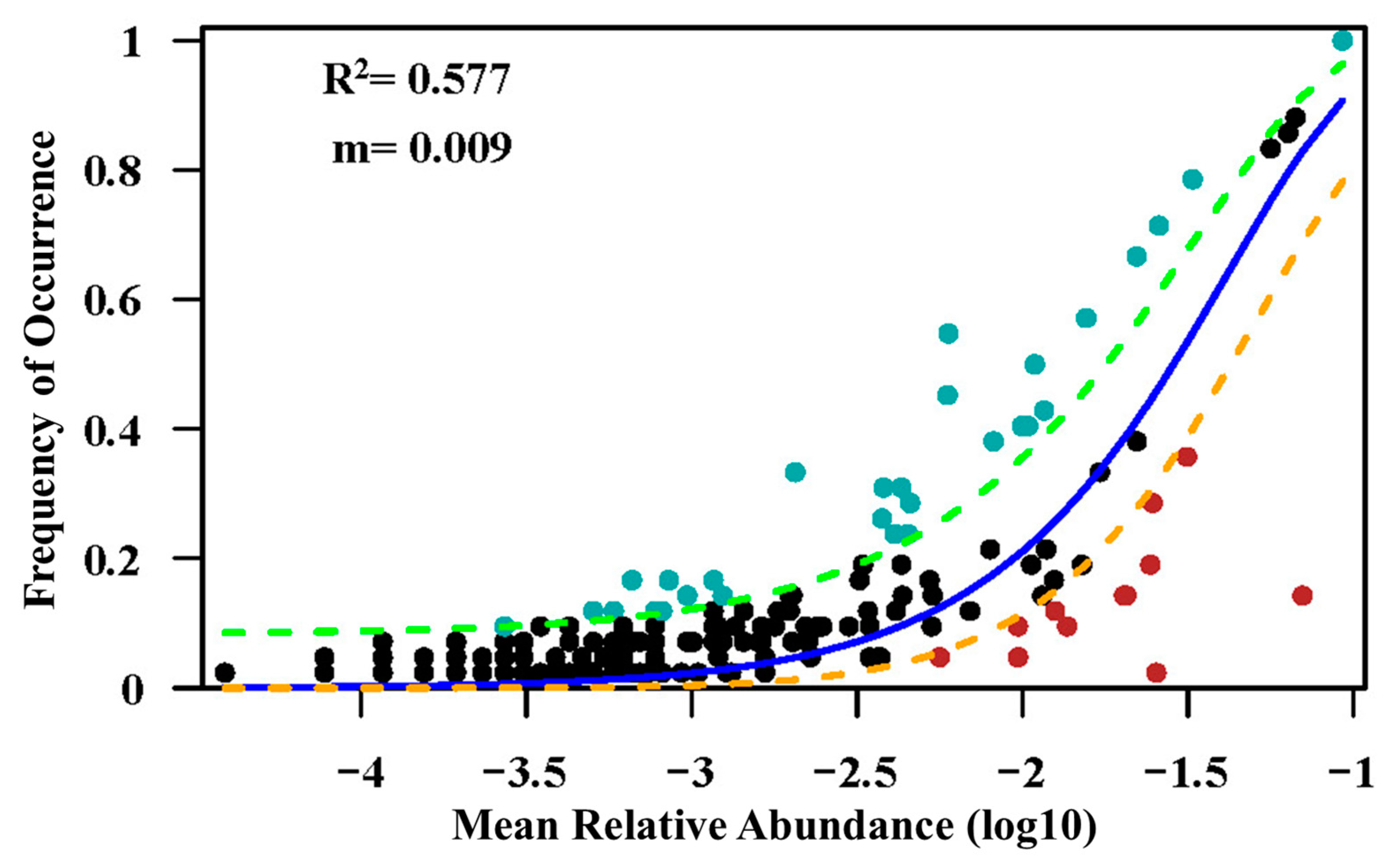
3.5. Determination of Forest Community Assembly at Local and Regional Scales
4. Discussion
4.1. Differences in Beta Diversity at Different Spatial Scales and Their Drivers
4.2. Main Drivers of Different Community Assembly Stages
4.3. Recommendations on the Protection of Zonal Forest Vegetation
4.4. Research Limitations and Perspectives for Future Exploration
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Variables | Beta Diversity | |||
|---|---|---|---|---|
| Community | Tree Layer | Shrub Layer | Herb Layer | |
| S_PH | 0.027 | −0.041 | 0.087 | −0.003 |
| S_C | 0.126 | 0.064 | 0.142 ** | 0.063 |
| S_I | 0.148 * | 0.170 * | 0.169 *** | 0.044 |
| S_P | 0.278 *** | 0.235 ** | 0.174 ** | 0.234 ** |
| ASP | 0.028 | 0.034 | 0.028 | 0.017 |
| SLO | −0.029 | 0.027 | −0.078 | −0.078 |
| DCo | 0.151 * | 0.315 *** | 0.015 | 0.166 * |
| AP | 0.069 | 0.092 | 0.415 *** | 0.246 *** |
| MAT | 0.349 *** | 0.062 | 0.339 *** | 0.271 *** |
| Tmin | 0.400 *** | 0.072 | 0.372 *** | 0.291 *** |
| PDQ | 0.313 *** | 0.098 | 0.375 *** | 0.225 ** |
| TS | 0.456 *** | 0.149 | 0.443 *** | 0.365 *** |
| PER | 0.396 *** | 0.074 | 0.434 *** | 0.283 *** |
References
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef]
- Tang, Z.; Wang, Z.; Fang, J. Historical hypothesis in explaining spatial patterns of species richness. Biodivers. Sci. 2009, 17, 635–643. [Google Scholar]
- Socolar, J.B.; Gilroy, J.J.; Kunin, W.E.; Edwards, D.P. How Should Beta-Diversity Inform Biodiversity Conservation? Trends Ecol. Evol. 2016, 31, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.H. Evolution and Measurement of Species Diversity. TAXON 1972, 21, 213–251. [Google Scholar] [CrossRef]
- Whittaker, R.H. Vegetation of the Siskiyou Mountains, Oregon and California. Ecol. Monogr. 1960, 30, 280–338. [Google Scholar] [CrossRef]
- Condit, R.; Pitman, N.; Leigh, E.G.; Chave, J.; Terborgh, J.; Foster, R.B.; Núñez, P.; Aguilar, S.; Valencia, R.; Villa, G.; et al. Beta-diversity in tropical forest trees. Science 2002, 295, 666–669. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Comita, L.S.; Chase, J.M.; Sanders, N.J.; Swenson, N.G.; Crist, T.O.; Stegen, J.C.; Vellend, M.; Boyle, B.; Anderson, M.J.; et al. Disentangling the Drivers of β Diversity Along Latitudinal and Elevational Gradients. Science 2011, 333, 1755–1758. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J.; Crist, T.O.; Chase, J.M.; Vellend, M.; Inouye, B.D.; Freestone, A.L.; Sanders, N.J.; Cornell, H.V.; Comita, L.S.; Davies, K.F.; et al. Navigating the multiple meanings of β diversity: A roadmap for the practicing ecologist. Ecol. Lett. 2011, 14, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Carroll, C.; Roberts, D.R.; Michalak, J.L.; Lawler, J.J.; Nielsen, S.E.; Stralberg, D.; Hamann, A.; Mcrae, B.H.; Wang, T.L. Scale-dependent complementarity of climatic velocity and environmental diversity for identifying priority areas for conservation under climate change. Global Change Biol. 2017, 23, 4508–4520. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Xiao, N.W.; Luo, Z.L.; Liu, D.M.; Zhao, Z.P.; Guan, X.; Zang, C.X.; Li, J.S.; Shen, Z.H. Identifying conservation priority areas for gymnosperm species under climate changes in China. Biol. Conserv. 2021, 253, 108914. [Google Scholar] [CrossRef]
- Weiher, E.; Keddy, P.A. Assembly Rules, Null Models, and Trait Dispersion—New Questions Front Old Patterns. Oikos 1995, 74, 159–164. [Google Scholar] [CrossRef]
- Fernandez-Going, B.M.; Harrison, S.P.; Anacker, B.L.; Safford, H.D. Climate interacts with soil to produce beta diversity in Californian plant communities. Ecology 2013, 94, 2007–2018. [Google Scholar] [CrossRef] [PubMed]
- Gause, G. The Struggle for Existence; Dover Publications: New York, NY, USA, 2003. [Google Scholar]
- Hubbell, S. The Unified Neutral Theory of Biodiversity and Biogeography; Princeton University Press: Princeton, NJ, USA, 2001. [Google Scholar]
- Soininen, J.; McDonald, R.; Hillebrand, H. The distance decay of similarity in ecological communities. Ecography 2007, 30, 3–12. [Google Scholar] [CrossRef]
- Litvak, M.K.; Hansell, R.I.C. A Community Perspective on the Multidimensional Niche. J. Anim. Ecol. 1990, 59, 931–940. [Google Scholar] [CrossRef]
- Tuomisto, H.; Ruokolainen, K.; Yli-Halla, M. Dispersal, environment, and floristic variation of western Amazonian forests. Science 2003, 299, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Hubbell, S.P. Tree Dispersion, Abundance, and Diversity in a Tropical Dry Forest. Science 1979, 203, 1299–1309. [Google Scholar] [CrossRef]
- Chu, C.; Wang, Y.; Liu, Y.; Jiang, L.; He, F. Advances in species coexistence theory. Biodivers. Sci. 2017, 25, 345–354. [Google Scholar] [CrossRef]
- HilleRisLambers, J.; Adler, P.B.; Harpole, W.S.; Levine, J.M.; Mayfield, M.M. Rethinking Community Assembly through the Lens of Coexistence Theory. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 227–248. [Google Scholar] [CrossRef]
- Chase, J.M.; McGill, B.J.; Thompson, P.L.; Antao, L.H.; Bates, A.E.; Blowes, S.A.; Dornelas, M.; Gonzalez, A.; Magurran, A.E.; Supp, S.R.; et al. Species richness change across spatial scales. Oikos 2019, 128, 1079–1091. [Google Scholar] [CrossRef]
- Ricklefs, R.E. Community Diversity—Relative Roles of Local and Regional Processes. Science 1987, 235, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Harrison, S.; Ross, S.J.; Lawton, J.H. Beta-Diversity on Geographic Gradients in Britain. J. Anim. Ecol. 1992, 61, 151–158. [Google Scholar] [CrossRef]
- Niu, K.; Liu, Y.; Shen, Z.; He, F.; Fang, J. Community assembly: The relative importance of neutral theory and niche theory. Biodivers. Sci. 2009, 17, 579–593. [Google Scholar]
- Myers, J.A.; Chase, J.M.; Jiménez, I.; Jorgensen, P.M.; Araujo-Murakami, A.; Paniagua-Zambrana, N.; Seidel, R. Beta-diversity in temperate and tropical forests reflects dissimilar mechanisms of community assembly. Ecol. Lett. 2013, 16, 151–157. [Google Scholar] [CrossRef]
- Jones, M.M.; Tuomisto, H.; Clark, D.B.; Olivas, P. Effects of mesoscale environmental heterogeneity and dispersal limitation on floristic variation in rain forest ferns. J. Ecol. 2006, 94, 181–195. [Google Scholar] [CrossRef]
- Page, N.V.; Shanker, K. Environment and dispersal influence changes in species composition at different scales in woody plants of the Western Ghats, India. J. Veg. Sci. 2018, 29, 74–83. [Google Scholar] [CrossRef]
- Fang, W.J.; Cai, Q.; Zhao, Q.; Ji, C.J.; Zhu, J.L.; Tang, Z.Y.; Fang, J.Y. Species richness patterns and the determinants of larch forests in China. Plant Divers. 2022, 44, 436–444. [Google Scholar] [CrossRef]
- Villa, P.M.; Martins, S.V.; Diniz, É.S.; Neto, S.N.D.; Neri, A.V.; Pinto, H., Jr.; Nunes, J.A.; Bueno, M.L.; Ali, A. Taxonomic and functional beta diversity of woody communities along Amazon forest succession: The relative importance of stand age, soil properties and spatial factor. For. Ecol. Manag. 2021, 482, 118885. [Google Scholar] [CrossRef]
- Leprieur, F.; Tedesco, P.A.; Hugueny, B.; Beauchard, O.; Dürr, H.H.; Brosse, S.; Oberdorff, T. Partitioning global patterns of freshwater fish beta diversity reveals contrasting signatures of past climate changes. Ecol. Lett. 2011, 14, 325–334. [Google Scholar] [CrossRef]
- Ponisio, L.C.; M’Gonigle, L.K.; Kremen, C. On-farm habitat restoration counters biotic homogenization in intensively managed agriculture. Glob. Change Biol. 2016, 22, 704–715. [Google Scholar] [CrossRef]
- Kneitel, J.M.; Miller, T.E. Dispersal rates affect species composition in metacommunities of Sarracenia purpurea inquilines. Am. Nat. 2003, 162, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Kraft, N.J.B.; Ackerly, D.D. Assembly of Plant Communities. In Ecology and the Environment; Monson, R.K., Ed.; Springer: New York, NY, USA, 2014; pp. 67–88. [Google Scholar]
- Graham, C.H.; Fine, P.V.A. Phylogenetic beta diversity: Linking ecological and evolutionary processes across space in time. Ecol. Lett. 2008, 11, 1265–1277. [Google Scholar] [CrossRef]
- Cássia-Silva, C.; Freitas, C.G.; Alves, D.M.C.C.; Bacon, C.D.; Collevatti, R.G. Niche conservatism drives a global discrepancy in palm species richness between seasonally dry and moist habitats. Glob. Ecol. Biogeogr. 2019, 28, 814–825. [Google Scholar] [CrossRef]
- Purschke, O.; Schmid, B.C.; Sykes, M.T.; Poschlod, P.; Michalski, S.G.; Durka, W.; Kühn, I.; Winter, M.; Prentice, H.C. Contrasting changes in taxonomic, phylogenetic and functional diversity during a long-term succession: Insights into assembly processes. J. Ecol. 2013, 101, 857–866. [Google Scholar] [CrossRef]
- Liu, B.; Chen, H.Y.H.; Yang, J. Understory Community Assembly Following Wildfire in Boreal Forests: Shift from Stochasticity to Competitive Exclusion and Environmental Filtering. Front. Plant Sci. 2018, 9, 1854. [Google Scholar] [CrossRef]
- Grime, J.P. Trait convergence and trait divergence in herbaceous plant communities: Mechanisms and consequences. J. Veg. Sci. 2006, 17, 255–260. [Google Scholar] [CrossRef]
- Verdú, M.; Rey, P.J.; Alcántara, J.M.; Siles, G.; Valiente-Banuet, A. Phylogenetic signatures of facilitation and competition in successional communities. J. Ecol. 2009, 97, 1171–1180. [Google Scholar] [CrossRef]
- Norden, N.; Mesquita, R.C.G.; Bentos, T.V.; Chazdon, R.L.; Williamson, G.B. Contrasting community compensatory trends in alternative successional pathways in central Amazonia. Oikos 2011, 120, 143–151. [Google Scholar] [CrossRef]
- McGill, B.J.; Dornelas, M.; Gotelli, N.J.; Magurran, A.E. Fifteen forms of biodiversity trend in the Anthropocene. Trends Ecol. Evol. 2015, 30, 104–113. [Google Scholar] [CrossRef]
- Hill, S.L.L.; Harfoot, M.; Purvis, A.; Purves, D.W.; Collen, B.; Newbold, T.; Burgess, N.D.; Mace, G.M. Reconciling Biodiversity Indicators to Guide Understanding and Action. Conserv. Lett. 2016, 9, 405–412. [Google Scholar] [CrossRef]
- Bråthen, K.A.; Gonzalez, V.T.; Yoccoz, N.G. Gatekeepers to the effects of climate warming? Niche construction restricts plant community changes along a temperature gradient. Perspect. Plant Ecol. Evol. Syst. 2018, 30, 71–81. [Google Scholar] [CrossRef]
- Crockett, E.T.H.; Vellend, M.; Bennett, E.M. Tree biodiversity in northern forests shows temporal stability over 35 years at different scales, levels and dimensions. J. Ecol. 2022, 110, 2388–2403. [Google Scholar] [CrossRef]
- Levins, R. Some Demographic and Genetic Consequences of Environmental Heterogeneity for Biological Control1. Bull. Entomol. Soc. Am. 1969, 15, 237–240. [Google Scholar] [CrossRef]
- Leibold, M.A.; Mikkelson, G.M. Coherence, species turnover, and boundary clumping: Elements of meta-community structure. Oikos 2002, 97, 237–250. [Google Scholar] [CrossRef]
- Wilson, D.S. Complex Interactions in Metacommunities, with Implications for Biodiversity and Higher Levels of Selection. Ecology 1992, 73, 1984–2000. [Google Scholar] [CrossRef]
- Holyoak, M.; Leibold, M.A.; Holt, R.D. Metacommunities: Spatial Dynamics and Ecological Communities; University of Chicago Press: Chicago, IL, USA, 2005. [Google Scholar]
- Ricklefs, R.E. Disintegration of the Ecological Community. Am. Nat. 2008, 172, 741–750. [Google Scholar] [CrossRef]
- Saade, C.; Kefi, S.; Gougat-Barbera, C.; Rosenbaum, B.; Fronhofer, E.A. Spatial autocorrelation of local patch extinctions drives recovery dynamics in metacommunities. Proc. R. Soc. B-Biol. Sci. 2022, 289, 20220543. [Google Scholar] [CrossRef] [PubMed]
- Nekola, J.C.; White, P.S. The distance decay of similarity in biogeography and ecology. J. Biogeogr. 1999, 26, 867–878. [Google Scholar] [CrossRef]
- Peres-Neto, P.R.; Legendre, P.; Dray, S.; Borcard, D. Variation partitioning of species data matrices: Estimation and comparison of fractions. Ecology 2006, 87, 2614–2625. [Google Scholar] [CrossRef] [PubMed]
- Gross, T.; Allhoff, K.T.; Blasius, B.; Brose, U.; Drossel, B.; Fahimipour, A.K.; Guill, C.; Yeakel, J.D.; Zeng, F.Q. Modern models of trophic meta-communities. Philos. Trans. R. Soc. B-Biol. Sci. 2020, 375, 20190455. [Google Scholar] [CrossRef]
- Cañedo-Argüelles, M.; Boersma, K.S.; Bogan, M.T.; Olden, J.D.; Phillipsen, I.; Schriever, T.A.; Lytle, D.A. Dispersal strength determines meta-community structure in a dendritic riverine network. J. Biogeogr. 2015, 42, 778–790. [Google Scholar] [CrossRef]
- Tonkin, J.D.; Altermatt, F.; Finn, D.S.; Heino, J.; Olden, J.D.; Pauls, S.U.; Lytle, D.A. The role of dispersal in river network metacommunities: Patterns, processes, and pathways. Freshw. Biol. 2018, 63, 141–163. [Google Scholar] [CrossRef]
- Wu, Z.; Zhu, Y.; Jiang, H. Vegetation of Yunnan; Science Press: Beijing, China, 1987. [Google Scholar]
- Song, Q.; Chen, X.; Wang, X. Studies on Evergreen Broad-leaved Forests of China: A Retrospect and Prospect. J. East. China Norm. Univ. Nat. Sci. 2005, 2005, 1–8. [Google Scholar]
- Wang, R.; Bai, Y.; Alatalo, J.M.; Guo, G.; Yang, Z.; Yang, Z.; Yang, W. Impacts of urbanization at city cluster scale on ecosystem services along an urban-rural gradient: A case study of Central Yunnan City Cluster, China. Environ. Sci. Pollut. Res. 2022, 29, 88852–88865. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Peng, J. Vegetation of Kunming; Yunnan Science and Technology Press: Kunming, China, 1998. [Google Scholar]
- Tang, C.Q.; Chiou, C.R.; Lin, C.T.; Lin, J.R.; Hsieh, C.F.; Tang, J.W.; Su, W.H.; Hou, X.L. Plant diversity patterns in subtropical evergreen broad-leaved forests of Yunnan and Taiwan. Ecol. Res. 2013, 28, 81–92. [Google Scholar] [CrossRef]
- Tang, C.Q.; He, L.Y.; Su, W.H.; Zhang, G.F.; Wang, H.C.; Peng, M.C.; Wu, Z.L.; Wang, C.Y. Regeneration, recovery and succession of a Pinus yunnanensis community five years after a mega-fire in central Yunnan, China. For. Ecol. Manag. 2013, 294, 188–196. [Google Scholar] [CrossRef]
- Shen, Y.; Liu, W.; Li, Y.; Cui, J. Community ecology study on Karst semi-humid evergreen broad-leaved forest at the central part of Yunnan. Guihaia 2005, 25, 321–326. [Google Scholar]
- Wang, Z.; Duan, C.; Yang, J. Plant biodiversity and community structure of semi-humid evergreen broadleaved forests at different secondary succession stages. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2006, 17, 1583–1587. [Google Scholar]
- Han, J.; Shen, Z.H.; Li, Y.Y.; Luo, C.F.; Xu, Q.; Yang, K.; Zhang, Z.M. Beta Diversity Patterns of Post-fire Forests in Central Yunnan Plateau, Southwest China: Disturbances Intensify the Priority Effect in the Community Assembly. Front. Plant Sci. 2018, 9, 1000. [Google Scholar] [CrossRef]
- Luo, C.F.; Liu, Y.Q.; Shen, Z.H.; Yang, K.; Wang, X.P.; Jiang, Y.X. Modifying regeneration strategies classification to enhance the understanding of dominant species growth in fire-prone forest in Southwest China. For. Ecosyst. 2022, 9, 100009. [Google Scholar] [CrossRef]
- Xiao, M.; He, H.; Wang, Y.; Xiong, J.; He, Z. Genetic Properties and Taxonomy of Forest Soil in Central Yunnan Plateau. Mt. Res. 2019, 37, 359–370. [Google Scholar]
- Fang, J.; Shen, Z.; Tang, Z.; Wang, Z. The Protocol for the Survey Plan for Plant Species Diversity of China’s Mountains. Chin. Biodivers. 2004, 12, 5–9. [Google Scholar]
- Wang, H. A Field Guide to Wild Plants of Central Yunnan; Science Press: Beijing, China, 2020. [Google Scholar]
- iPlant. Available online: https://www.iplant.cn/ (accessed on 30 June 2023).
- Lu, R. Methods of Soil and Agro-Chemical Analysis; China Agricultural Science and Technology Press: Beijing, China, 2000. [Google Scholar]
- Zhang, G.; Gong, Z. Soil Survey Laboratory Methods; Science Press: Beijing, China, 2012. [Google Scholar]
- Ying, L.X.; Shen, Z.H.; Yang, M.Z.; Piao, S.L. Wildfire Detection Probability of MODIS Fire Products under the Constraint of Environmental Factors: A Study Based on Confirmed Ground Wildfire Records. Remote Sens. 2019, 11, 3031. [Google Scholar] [CrossRef]
- Holdridge, L.; Grenke, W.; Hatheway, W.; Liang, T.; Tosi, J. Forest Environments in Tropical Life Zones: A Pilot Study; Pergamon Press: New York, NY, USA, 1971. [Google Scholar]
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 326–349. [Google Scholar] [CrossRef]
- Zhang, J.T. Quantitative Ecology; Science Press: Beijing, China, 2018. [Google Scholar]
- Oksanen, J.; Simpson, G.L.; Blanche, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. R Package Version 2.6-2. 2022. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 21 January 2025).
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Baselga, A.; Orme, C.D.L. betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Colwell, R.K.; Mao, C.X.; Chang, J. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 2004, 85, 2717–2727. [Google Scholar] [CrossRef]
- Tuomisto, H.; Ruokolainen, K. Analyzing or explaining beta diversity? Understanding the targets of different methods of analysis. Ecology 2006, 87, 2697–2708. [Google Scholar] [CrossRef]
- Hijmans, R.J. Geosphere: Spherical Trigonometry. R Package Version 1.5-18. 2023. Available online: https://cran.r-project.org/web/packages/geosphere/index.html (accessed on 21 January 2025).
- Kassambara, A. Ggpubr: ‘Ggplot2’ Based Publication Ready Plots. R Package Version 0.6.0. 2023. Available online: https://cran.r-project.org/web/packages/ggpubr/index.html (accessed on 21 January 2025).
- Zhang, J.; Liu, B.; Liu, S.; Feng, Z.; Jiang, K. Plantlist: Looking Up the Status of Plant Scientific Names Based on the Plant List Database. R Package Version 0.8.0. 2023. Available online: https://github.com/helixcn/plantlist/ (accessed on 21 January 2025).
- Jin, Y.; Qian, H.U. PhyloMaker: An R package that can generate large phylogenetic trees for plants and animals. Plant Divers. 2023, 45, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Haston, E.; Richardson, J.E.; Stevens, P.F.; Chase, M.W.; Harris, D.J. The Linear Angiosperm Phylogeny Group (LAPG) III: A linear sequence of the families in APG III. Bot. J. Linn. Soc. 2009, 161, 128–131. [Google Scholar] [CrossRef]
- Zanne, A.E.; Tank, D.C.; Cornwell, W.K.; Eastman, J.M.; Smith, S.A.; FitzJohn, R.G.; McGlinn, D.J.; O’Meara, B.C.; Moles, A.T.; Reich, P.B.; et al. Three keys to the radiation of angiosperms into freezing environments. Nature 2014, 506, 89. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.A.; Brown, J.W. Constructing a broadly inclusive seed plant phylogeny. Am. J. Bot. 2018, 105, 302–314. [Google Scholar] [CrossRef]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef]
- Cooper, N.; Rodríguez, J.; Purvis, A. A common tendency for phylogenetic overdispersion in mammalian assemblages. Proc. R. Soc. B-Biol. Sci. 2008, 275, 2031–2037. [Google Scholar] [CrossRef]
- Cooper, N.; Purvis, A. What factors shape rates of phenotypic evolution? A comparative study of cranial morphology of four mammalian clades. J. Evol. Biol. 2009, 22, 1024–1035. [Google Scholar] [CrossRef] [PubMed]
- Proches, S.; Wilson, J.R.U.; Cowling, R.M. How much evolutionary history in a 10 × 10 m plot? Proc. R. Soc. B-Biol. Sci. 2006, 273, 1143–1148. [Google Scholar] [CrossRef] [PubMed]
- Webb, C.O.; Ackerly, D.D.; Kembel, S.W. Phylocom: Software for the analysis of phylogenetic community structure and trait evolution. Bioinformatics 2008, 24, 2098–2100. [Google Scholar] [CrossRef]
- Kassambara, A. Rstatix: Pipe-Friendly Framework for Basic Statistical Tests. R Package Version 0.7.2. 2023. Available online: https://rpkgs.datanovia.com/rstatix/ (accessed on 21 January 2025).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer: New York, NY, USA, 2016. [Google Scholar] [CrossRef]
- Freestone, A.L.; Inouye, B.D. Dispersal limitation and environmental heterogeneity shape scale-dependent diversity patterns in plant communities. Ecology 2006, 87, 2425–2432. [Google Scholar] [CrossRef]
- Blundo, C.; González-Espinosa, M.; Malizia, L.R. Relative contribution of niche and neutral processes on tree species turnover across scales in seasonal forests of NW Argentina. Plant Ecol. 2016, 217, 359–368. [Google Scholar] [CrossRef]
- Jones, M.M.; Gibson, N.; Yates, C.; Ferrier, S.; Mokany, K.; Williams, K.J.; Manion, G.; Svenning, J.C. Underestimated effects of climate on plant species turnover in the Southwest Australian Floristic Region. J. Biogeogr. 2016, 43, 289–300. [Google Scholar] [CrossRef]
- Tang, Z.Y.; Fang, J.Y.; Chi, X.L.; Yang, Y.H.; Ma, W.H.; Mohhamot, A.; Guo, Z.D.; Liu, Y.N.; Gaston, K.J. Geography, environment, and spatial turnover of species in China’s grasslands. Ecography 2012, 35, 1103–1109. [Google Scholar] [CrossRef]
- Goldberg, E.E.; Lande, R. Species’ borders and dispersal barriers. Am. Nat. 2007, 170, 297–304. [Google Scholar] [CrossRef]
- Wang, G.H.; Zhou, G.S.; Yang, L.M.; Li, Z.Q. Distribution, species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slopes of Qilianshan Mountains, Gansu, China. Plant Ecol. 2003, 165, 169–181. [Google Scholar] [CrossRef]
- Jansson, R.; Dynesius, M. The fate of clades in a world of recurrent climatic change: Milankovitch oscillations and evolution. Annu. Rev. Ecol. Syst. 2002, 33, 741–777. [Google Scholar] [CrossRef]
- Dobrovolski, R.; Melo, A.S.; Cassemiro, F.A.S.; Diniz, J.A.F. Climatic history and dispersal ability explain the relative importance of turnover and nestedness components of beta diversity. Global Ecol. Biogeogr. 2012, 21, 191–197. [Google Scholar] [CrossRef]
- Guèze, M.; Paneque-Gálvez, J.; Luz, A.C.; Pino, J.; Orta-Martínez, M.; Reyes-García, V.; Macía, M.J. Determinants of tree species turnover in a southern Amazonian rain forest. J. Veg. Sci. 2013, 24, 284–295. [Google Scholar] [CrossRef]
- Qian, H.; Shimono, A. Effects of geographic distance and climatic dissimilarity on species turnover in alpine meadow communities across a broad spatial extent on the Tibetan Plateau. Plant Ecol. 2012, 213, 1357–1364. [Google Scholar] [CrossRef]
- Rosindell, J.; Hubbell, S.P.; Etienne, R.S. The Unified Neutral Theory of Biodiversity and Biogeography at age ten. Trends Ecol. Evol. 2011, 26, 340–348. [Google Scholar] [CrossRef]
- Aiello-Lammens, M.E.; Slingsby, J.A.; Merow, C.; Mollmann, H.K.; Euston-Brown, D.; Jones, C.S.; Silander, J.A. Processes of community assembly in an environmentally heterogeneous, high biodiversity region. Ecography 2017, 40, 561–576. [Google Scholar] [CrossRef]
- Oliveira, A.T.; Fontes, M.A.L. Patterns of floristic differentiation among Atlantic forests in southeastern Brazil and the influence of climate. Biotropica 2000, 32, 793–810. [Google Scholar] [CrossRef]
- Cottenie, K. Integrating environmental and spatial processes in ecological community dynamics. Ecol. Lett. 2005, 8, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.P.; Su, X.Y.; Shrestha, N.; Xu, X.T.; Wang, S.Y.; Li, Y.Q.; Wang, Q.G.; Sandanov, D.; Wang, Z.H. Effects of contemporary environment and Quaternary climate change on drylands plant diversity differ between growth forms. Ecography 2019, 42, 334–345. [Google Scholar] [CrossRef]
- Arneth, A.; Shin, Y.J.; Leadley, P.; Rondinini, C.; Bukvareva, E.; Kolb, M.; Midgley, G.F.; Oberdorff, T.; Palomo, I.; Saito, O. Post-2020 biodiversity targets need to embrace climate change. Proc. Natl. Acad. Sci. USA 2020, 117, 30882–30891. [Google Scholar] [CrossRef]
- John, R.; Dalling, J.W.; Harms, K.E.; Yavitt, J.B.; Stallard, R.F.; Mirabello, M.; Hubbell, S.P.; Valencia, R.; Navarrete, H.; Vallejo, M.; et al. Soil nutrients influence spatial distributions of tropical tree species. Proc. Natl. Acad. Sci. USA 2007, 104, 864–869. [Google Scholar] [CrossRef] [PubMed]
- Valencia, R.; Foster, R.B.; Villa, G.; Condit, R.; Svenning, J.C.; Hernández, C.; Romoleroux, K.; Losos, E.; Magård, E.; Balslev, H. Tree species distributions and local habitat variation in the Amazon: Large forest plot in eastern Ecuador. J. Ecol. 2004, 92, 214–229. [Google Scholar] [CrossRef]
- Baldeck, C.A.; Harms, K.E.; Yavitt, J.B.; John, R.; Turner, B.L.; Valencia, R.; Navarrete, H.; Davies, S.J.; Chuyong, G.B.; Kenfack, D.; et al. Soil resources and topography shape local tree community structure in tropical forests. Proc. R. Soc. B-Biol. Sci. 2013, 280, 20122532. [Google Scholar] [CrossRef]
- Legendre, P. Spatial Autocorrelation—Trouble or New Paradigm. Ecology 1993, 74, 1659–1673. [Google Scholar] [CrossRef]
- Wagner, H.H. Spatial covariance in plant communities: Integrating ordination, geostatistics, and variance testing. Ecology 2003, 84, 1045–1057. [Google Scholar] [CrossRef]
- Abbas, S.; Nichol, J.E.; Zhang, J.L.; Fischer, G.A. The accumulation of species and recovery of species composition along a 70 year succession in a tropical secondary forest. Ecol. Indic. 2019, 106, 105524. [Google Scholar] [CrossRef]
- Zhu, K. Understanding forest dynamics by integrating age and environmental change. New Phytol. 2020, 228, 1728–1733. [Google Scholar] [CrossRef] [PubMed]
- Bongers, F.J.; Schmid, B.; Bruelheide, H.; Bongers, F.; Li, S.; von Oheimb, G.; Li, Y.; Cheng, A.P.; Ma, K.P.; Liu, X.J. Functional diversity effects on productivity increase with age in a forest biodiversity experiment. Nat. Ecol. Evol. 2021, 5, 1594. [Google Scholar] [CrossRef]
- Cavender-Bares, J.; Ackerly, D.D.; Baum, D.A.; Bazzaz, F.A. Phylogenetic overdispersion in Floridian oak communities. Am. Nat. 2004, 163, 823–843. [Google Scholar] [CrossRef]
- Swenson, N.G.; Enquist, B.J.; Thompson, J.; Zimmerman, J.K. The influence of spatial and size scale on phylogenetic relatedness in tropical forest communities. Ecology 2007, 88, 1770–1780. [Google Scholar] [CrossRef]
- Letcher, S.G. Phylogenetic structure of angiosperm communities during tropical forest succession. Proc. R. Soc. B-Biol. Sci. 2010, 277, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Hou, M.; Li, X.; Wang, J.; Liu, S.; Zhao, X. Phylogenetic development and functional structures during successional stages of conifer and broad-leaved mixed forest communities in Changbai Mountains, China. Acta Ecol. Sin. 2017, 37, 7503–7513. [Google Scholar]
- Xu, Y.; Shen, Z.H.; Ying, L.X.; Wang, Z.H.; Huang, J.H.; Zang, R.G.; Jiang, Y.X. Hotspot analyses indicate significant conservation gaps for evergreen broadleaved woody plants in China. Sci. Rep. 2017, 7, 1859. [Google Scholar] [CrossRef]
- Savilaakso, S.; Häkkilä, M.; Johansson, A.; Uusitalo, A.; Sandgren, T.; Mönkkönen, M.; Puttonen, P. What are the effects of even-aged and uneven-aged forest management on boreal forest biodiversity in Fennoscandia and European Russia? A systematic review protocol. Environ. Evid. 2019, 8, 17. [Google Scholar] [CrossRef]
| Factor | Variable | Unit | Description of Method or Algorithm | |
|---|---|---|---|---|
| ENV | Climate | MAP | mm | Mean annual precipitation of 1986–2015 |
| MAT | °C | Mean annual temperature of 1986–2015 | ||
| PET | °C | Potential evapotranspiration: 58.93 × ABT *; ABT = ΣTi/12; Ti, monthly mean temperature over 0 °C (assigned to 30 °C when Ti > 30 °C [73]) | ||
| PDQ | mm | Precipitation in the driest quarter, i.e., winter | ||
| PER | °C/mm | Moisture index = PET/MAP | ||
| TS | - | Temperature seasonality: SD (monthly mean temperature) × 100 | ||
| Topography | ASP | - | Cos(aspect), aspect ranges from 0 (north) to 180 (south) | |
| SLO | ° | Slope angle | ||
| Soil | S_PH | - | Soil pH | |
| S_C | g/kg | Soil organic carbon content | ||
| S_N | g/kg | Soil total nitrogen | ||
| S_I | cmol/kg | Soil cation exchange capacity | ||
| S_P | g/kg | Soil total phosphorus | ||
| Dominance of C. orthacantha | DCo | % | The proportion of the total basal area of C. orthacantha Castanopsis orthacantha Franch. in the total basal area of the tree layer in each plot | |
| Age | AGE | Year | Three age classes: old (>60 yr), mediate (40–60 yr), young (25–40 yr) | |
| Geographic distance | GEO | Km | Euclidean distance between two plots, calculated using geographical coordinates | |
| Variables | All Species | Tree Layer | Shrub Layer | Herb Layer | ||||
|---|---|---|---|---|---|---|---|---|
| Sørensen | Bray | Sørensen | Bray | Sørensen | Bray | Sørensen | Bray | |
| GEO | 0.617 *** | 0.544 *** | 0.532 *** | 0.404 *** | 0.485 *** | 0.562 *** | 0.443 *** | 0.429 *** |
| ENV | 0.592 *** | 0.557 *** | 0.462 *** | 0.385 ** | 0.550 *** | 0.590 *** | 0.363 *** | 0.409 *** |
| Climate | 0.576 *** | 0.515 *** | 0.439 *** | 0.288 ** | 0.491 *** | 0.533 *** | 0.405 *** | 0.414 *** |
| Soil | 0.278 ** | 0.303 *** | 0.223 ** | 0.263 ** | 0.312 ** | 0.299 ** | 0.113 | 0.211 ** |
| TOPO | 0.066 | 0.092 | 0.090 | 0.125 * | 0.039 | −0.009 | 0.040 | 0.059 |
| AGE | 0.054 | 0.122 * | 0.060 | 0.215 ** | 0.029 | 0.009 | 0.072 | 0.074 * |
| GEO|ENV | 0.355 *** | 0.263 *** | 0.331 *** | 0.207 ** | 0.172 * | 0.263 *** | 0.285 *** | 0.220 ** |
| ENV|GEO | 0.289 ** | 0.298 * | 0.153 * | 0.159 * | 0.339 ** | 0.337 * | 0.137 * | 0.172 * |
| Climate|GEO | 0.227 ** | 0.196 * | 0.075 | −0.023 | 0.222 ** | 0.205 * | 0.124 * | 0.157 * |
| Soil|GEO | 0.205 ** | 0.239 ** | 0.143 | 0.205 ** | 0.254 ** | 0.223 ** | 0.081 | 0.144 * |
| TOPO|GEO | 0.050 | 0.082 | 0.079 | 0.118 | 0.020 | 0.040 | 0.023 | 0.045 |
| AGE|GEO | −0.008 | 0.042 | 0.009 | 0.193 ** | −0.020 | −0.055 | 0.032 | 0.036 |
| AGE|Climate | 0.096 | 0.132 * | 0.088 | 0.250 *** | 0.061 | 0.040 | 0.092 | 0.098 |
| Variables | Inter-Sites | Intra-Site | ||
|---|---|---|---|---|
| Sørensen | Bray | Sørensen | Bray | |
| GEO | 0.579 *** | 0.375 *** | 0.165 * | 0.294 *** |
| ENV | 0.531 *** | 0.205 * | 0.244 *** | 0.308 *** |
| Climate | 0.508 *** | 0.153 * | 0.22 *** | 0.261 *** |
| Soil | 0.068 | 0.100 | 0.215 *** | 0.230 *** |
| TOPO | 0.087 | 0.167 * | 0.186 ** | 0.255 *** |
| AGE | 0.115 * | 0.127 * | 0.019 | 0.184 ** |
| GEO|ENV | 0.393 *** | 0.355 *** | 0.141 * | 0.287 *** |
| ENV|GEO | 0.315 *** | 0.180 ** | 0.196 ** | 0.234 *** |
| Climate|GEO | 0.301 *** | −0.131 | 0.172 ** | 0.243 *** |
| Soil|GEO | 0.002 | 0.064 | 0.170 ** | 0.221 *** |
| TOPO|GEO | 0.051 | −0.107 | 0.159 * | 0.238 *** |
| AGE|GEO | −0.049 | 0.051 | −0.024 | 0.127 * |
| AGE|Climate | 0.184 ** | 0.144 * | 0.019 | 0.202 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Zhang, Q.; Yang, T.; Tian, X.; Zhang, Y.; Shen, Z. Roles of Spatial Distance, Habitat Difference, and Community Age on Plant Diversity Patterns of Fragmented Castanopsis orthacantha Franch. Forests in Central Yunnan, Southwest China. Forests 2025, 16, 245. https://doi.org/10.3390/f16020245
Wang X, Zhang Q, Yang T, Tian X, Zhang Y, Shen Z. Roles of Spatial Distance, Habitat Difference, and Community Age on Plant Diversity Patterns of Fragmented Castanopsis orthacantha Franch. Forests in Central Yunnan, Southwest China. Forests. 2025; 16(2):245. https://doi.org/10.3390/f16020245
Chicago/Turabian StyleWang, Xinpei, Qiuyu Zhang, Tao Yang, Xi Tian, Ying Zhang, and Zehao Shen. 2025. "Roles of Spatial Distance, Habitat Difference, and Community Age on Plant Diversity Patterns of Fragmented Castanopsis orthacantha Franch. Forests in Central Yunnan, Southwest China" Forests 16, no. 2: 245. https://doi.org/10.3390/f16020245
APA StyleWang, X., Zhang, Q., Yang, T., Tian, X., Zhang, Y., & Shen, Z. (2025). Roles of Spatial Distance, Habitat Difference, and Community Age on Plant Diversity Patterns of Fragmented Castanopsis orthacantha Franch. Forests in Central Yunnan, Southwest China. Forests, 16(2), 245. https://doi.org/10.3390/f16020245