
Phenomenon and Mechanisms of Sonneratia apetala Introduction and Spread Promoting Excessive Growth of Derris trifoliata

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
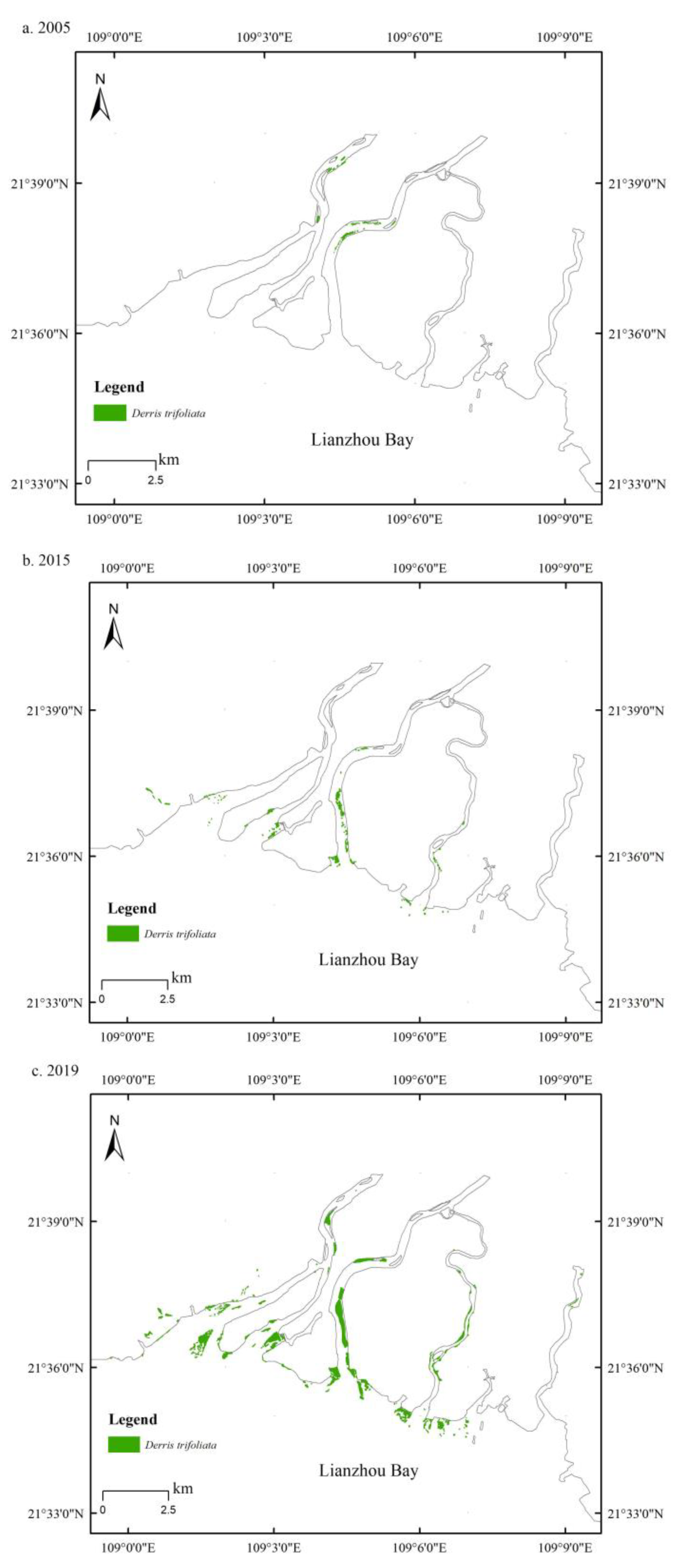
2.1. Distribution Survey of S. apetala and D. trifoliata
2.2. Comparison of Seed Quantity of D. trifoliata on Different Plants
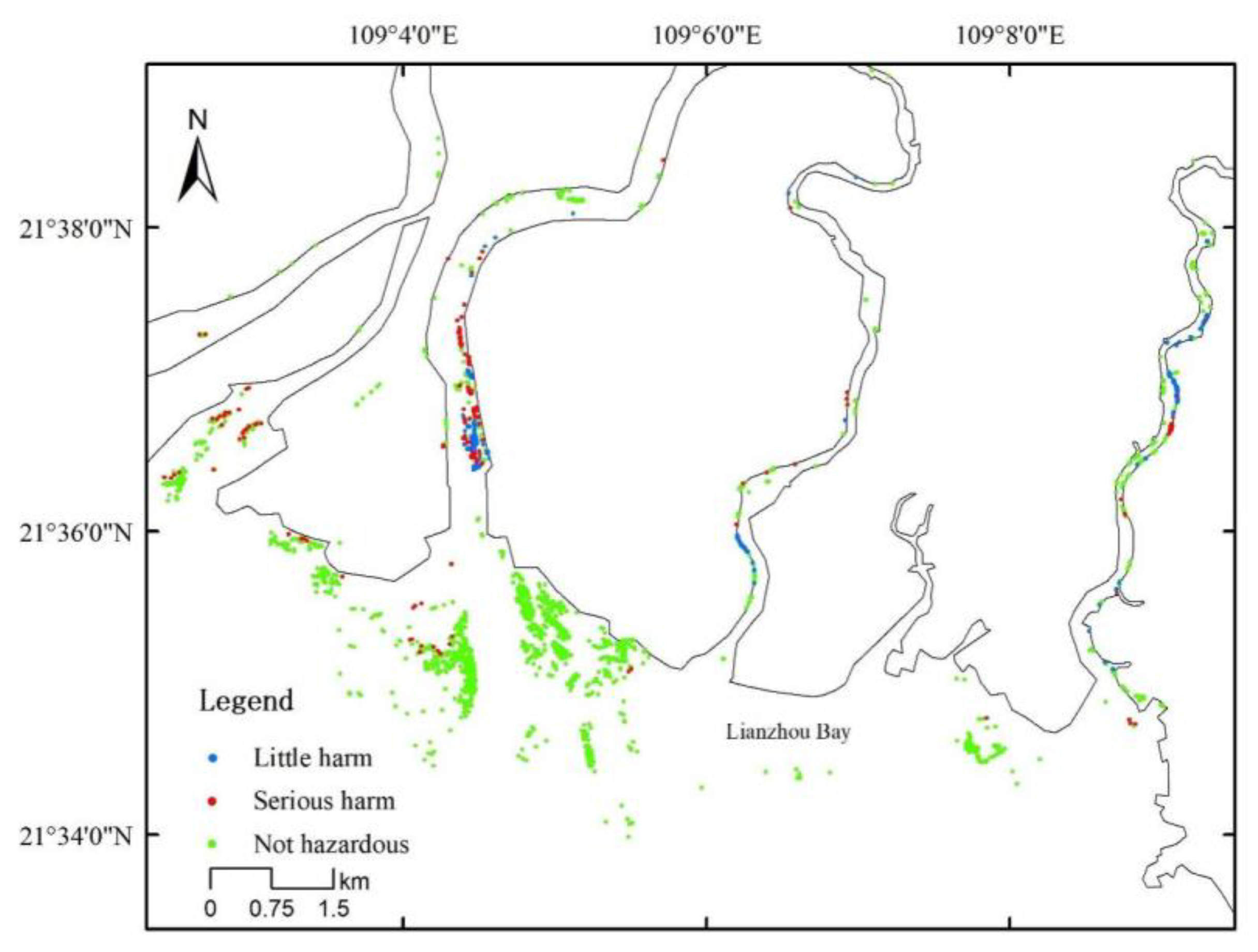
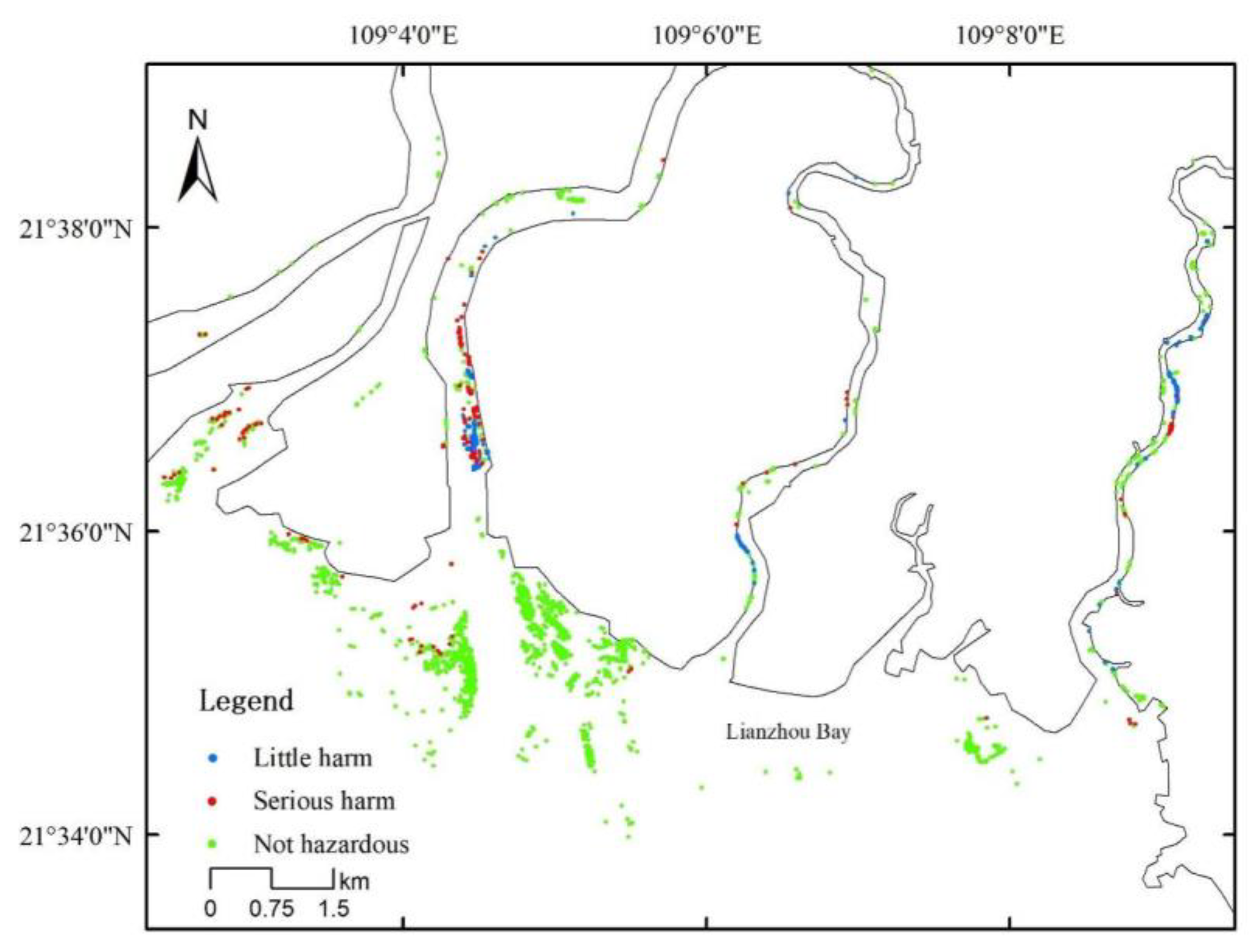
2.3. Survey of D. trifoliata Damage on S. apetala
2.4. MaxEnt Model
2.5. Data Analysis
3. Results and Analysis
3.1. Overgrowth of D. trifoliata and Spread of S. apetala after Introduction
3.2. Long-Term Seed Production of D. trifoliata on S. apetala
3.3. Active Propagation of S. apetala and Its Rapid Growth Facilitate the Expansion of D. trifoliata
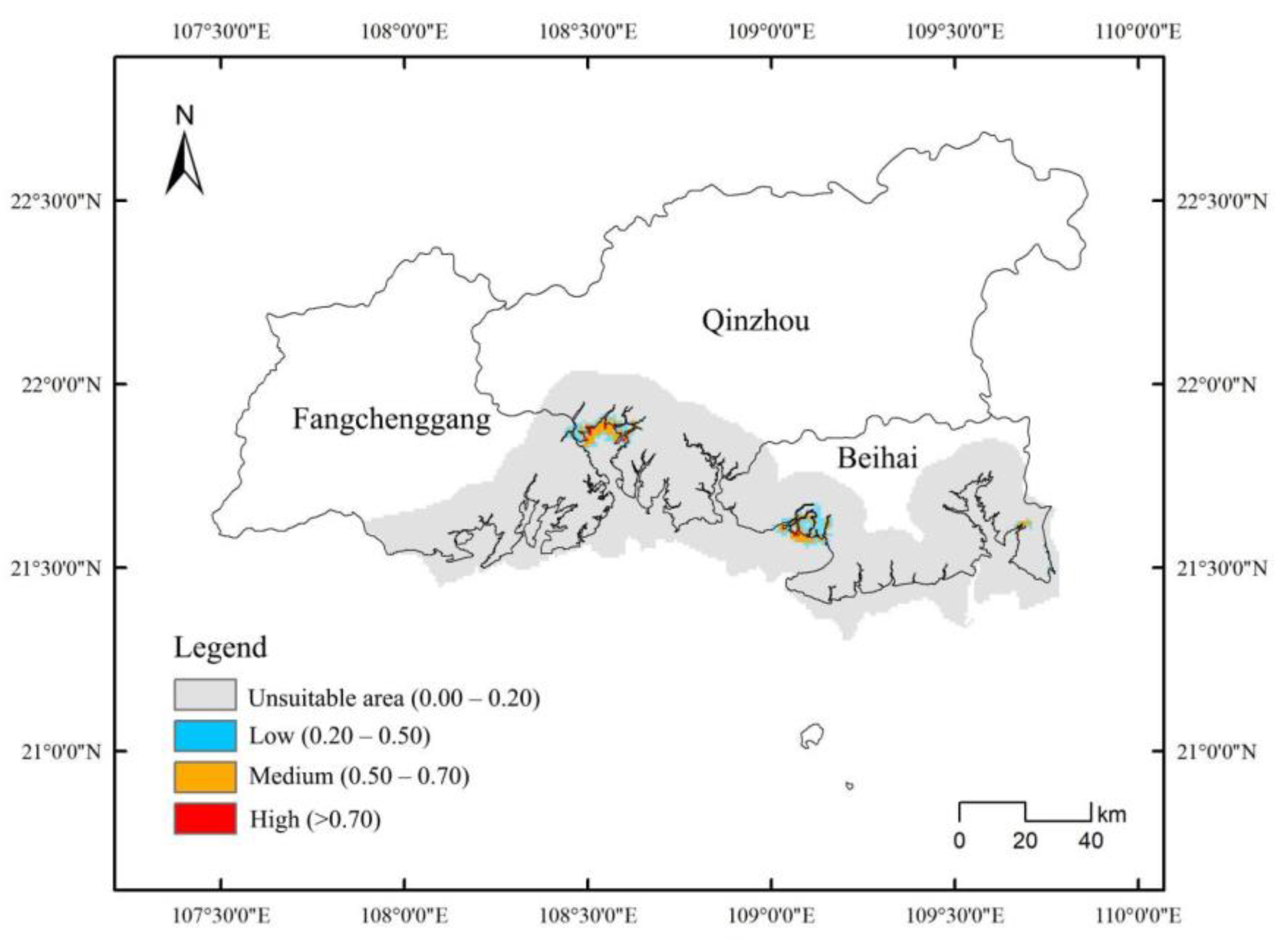
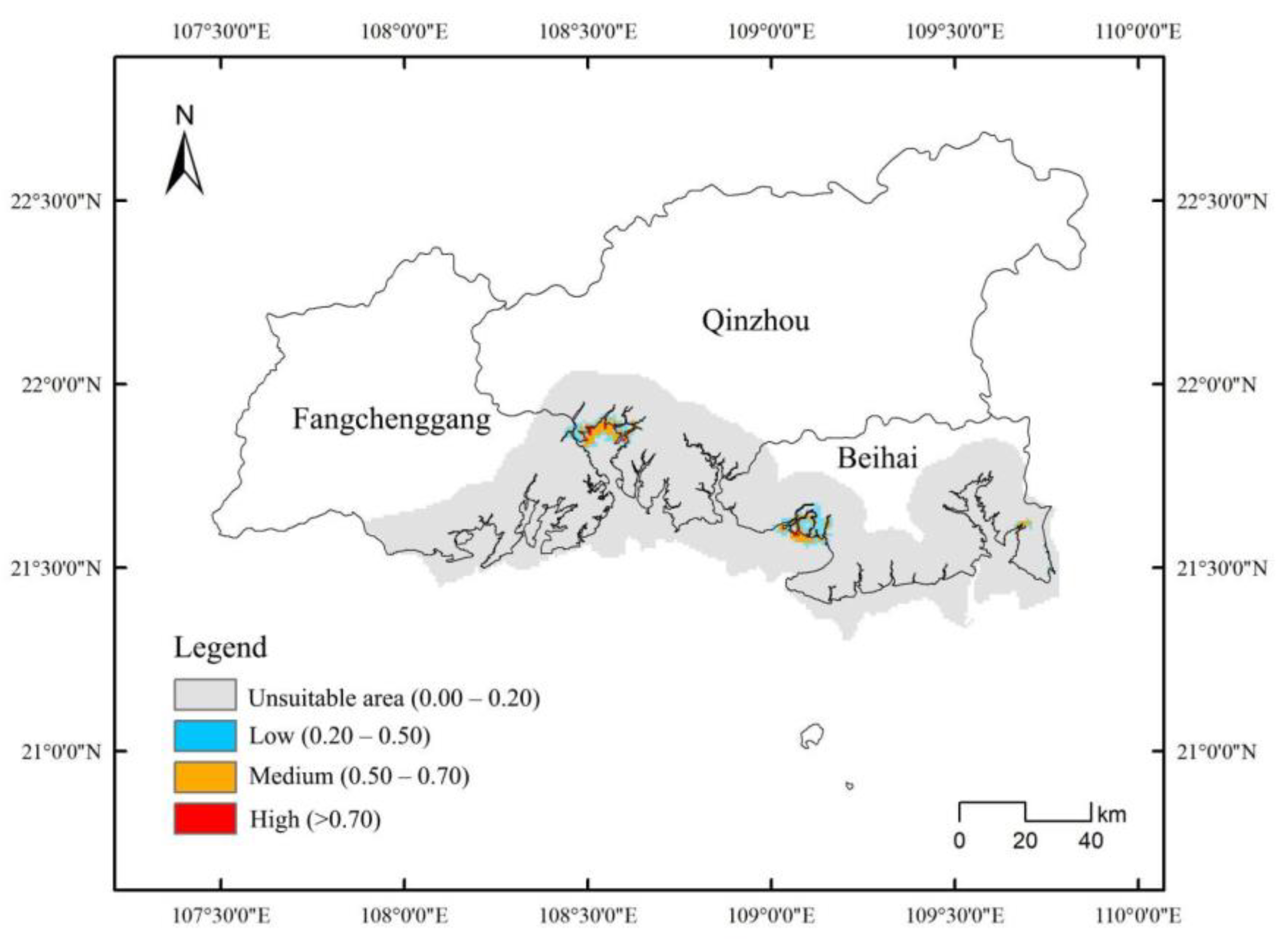
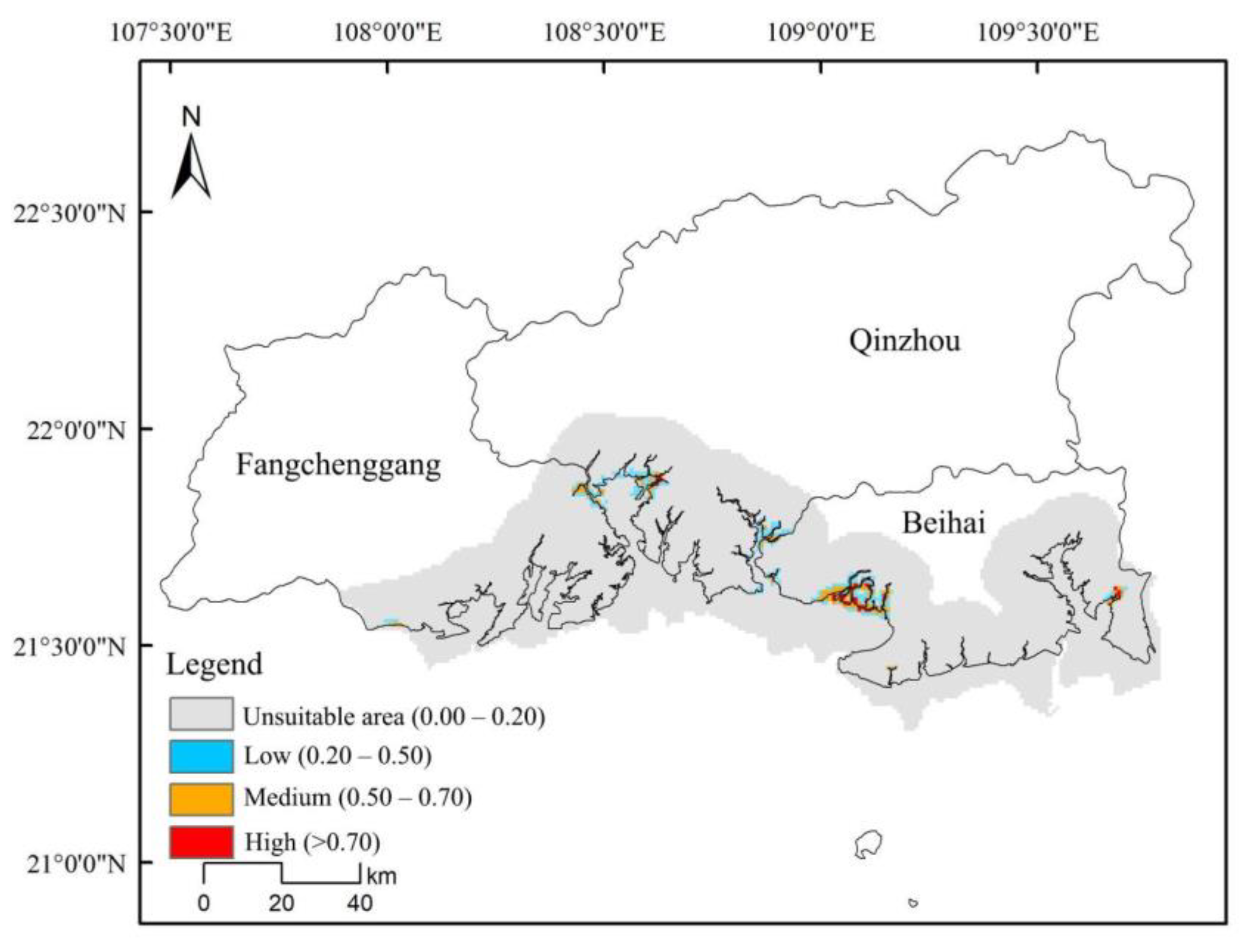
3.4. High Overlap in the Suitable Areas for D. trifoliata and S. apetala
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biswas, S.R.; Biswas, P.L.; Limon, S.H.; Yan, E.R.; Xu, M.S.; Khan, M.S.I. Plant invasion in mangrove forests worldwide. For. Ecol. Manag. 2018, 429, 480–492. [Google Scholar] [CrossRef]
- Chen, Q.; Ma, K.M. Overview and trends in research on biological invasion in mangrove ecosystems. Acta Pharmacol. Sin. 2015, 39, 283–299. [Google Scholar] [CrossRef]
- Pan, L.H.; Shi, X.F.; Zeng, C.; Chen, Y.S. Plant types in mangroves of Guangxi. Guangxi Sci. 2018, 25, 352–362. [Google Scholar] [CrossRef]
- Wu, D.Q. Research on the spread status of Sonneratia apetala in the Zhangjiangkou national mangrove nature Reserve. For. Prot. Sci. Technol. 2016, 33–35. [Google Scholar] [CrossRef]
- Zhao, C.; Qin, C.Z.; Wang, Z.; Mao, D.; Wang, Y.; Jia, M. Decision surface optimization in mapping exotic mangrove species (Sonneratia apetala) across latitudinal coastal areas of China. ISPRS J. Photogramm. 2022, 193, 269–283. [Google Scholar] [CrossRef]
- Knauf, A.E.; Litton, C.M.; Cole, R.J.; Sparks, J.P.; Giardina, C.P.; Gerow, K.G.; Quiones-Santiago, M. Nutrient use strategy and not competition determines native and invasive species response to changes in soil nutrient availability. Restor. Ecol. 2021, 29, e13374. [Google Scholar] [CrossRef]
- Chen, Y.J.; Liao, B.W.; Zheng, S.F.; Li, M.; Song, X.Y. Dynamic Changes and Species Diversity of the Mangrove Community with Excoecaria agallocha, Kandelia candel, and Avicennia marina. Chin. J. Appl. Ecol. 2004, 15, 924–928. [Google Scholar]
- Huang, Z.J.; Li, Z.; Wang, M.H.; Zhu, D.H.; Yang, Q.; Yu, S.X. Coastal nutrient enrichments facilitated reproductive output in exotic mangrove species over two decades. Front. For. Glob. Chang. 2023, 6, 1–13. [Google Scholar] [CrossRef]
- Zhang, J.; Lin, Q.; Peng, Y.; Pan, L.; Chen, Y.; Zhang, Y.; Chen, L. Distributions of the non-native mangrove Sonneratia apetala in China: Based on google earth imagery and field survey. Wetlands 2022, 42, 35. [Google Scholar] [CrossRef]
- Hu, Z.W.; Wu, J.J.; Wang, J.Z.; Zhang, Y.H.; Zhou, H.C.; Gao, C.J.; Wang, J.J.; Wu, G.F. How exotic Sonneratia species affect the spatiotemporal dynamics of mangroves in Shenzhen Bay, China: A remote sensing perspective. Ecol. Indic. 2023, 153, 110479. [Google Scholar] [CrossRef]
- Chen, G.G.; Li, Y.Y.; Cai, L.Q.; Xu, J.J.; Lin, P.R.; Chen, W.C.; Huang, S.L. Influence of the exotic mangrove species Excoecaria agallocha on the morphological characteristics and biomass of the indigenous species Avicennia marina. Oceanol. Limnol. Sin. 2017, 41, 26–33. [Google Scholar]
- Cao, H.; Liu, S.H.; Liu, S.Y. Ecological health assessment and protection strategies of mangrove ecosystem in Huachang Bay, Hainan. S. China For. Investig. Plan. 2020, 39, 16–19+28. [Google Scholar] [CrossRef]
- Li, L.F.; Liu, W.I.; Ai, J.W.; Cai, S.J.; Dong, J.W. Predicting Mangrove Distributions in the Beibu Gulf, Guangxi, China, Using the MaxEnt Model: Determining Tree Species Selection. Forests 2023, 14, 149. [Google Scholar] [CrossRef]
- Li, L.F.; Lin, J. Distribution and control suggestions of Excoecaria agallocha in Dongzhaigang national nature Reserve, Hainan. Trop. For. 2019, 47, 36–38. [Google Scholar] [CrossRef]
- Huang, X.Y.; Zhong, C.; Chen, S.Y.; Liu, Y.; Liang, S.C. Harm and management of Fishvine (Kandelia candel) on Mangrove Plants. Wetl. Sci. Manag. 2015, 11, 26–29. [Google Scholar]
- Cobos, M.E.; Peterson, A.T.; Barve, N.; Osorio-Olvera, L. kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef]
- Chao, B.X.; Hu, W.J.; Chen, B.; Zhang, D.; Chen, G.C.; Yu, W.W.; Ma, Z.Y.; Lei, G.C.; Wang, Y.Y. Potential suitable habitat of mangroves and conservation gap analysis in Guangdong Province with MaxEnt Modeling. Chin. J. Ecol. 2020, 39, 3785–3794. [Google Scholar] [CrossRef]
- Wang, W.T.; Gao, S.Y.; Wang, S.F. Predictive Study on Potential Invasion Areas of Four Toxic Weeds in Gansu Grasslands. Acta Ecol. Sin. 2019, 39, 5301–5307. [Google Scholar] [CrossRef]
- Peng, Y.G.; Xu, Z.C.; Liu, M.C. Introduction and Ecological Impact of the Exotic Mangrove Plant Sonneratia apetala. Acta Ecol. Sin. 2012, 32, 2259–2270. [Google Scholar] [CrossRef]
- Wu, Z.H.; Cai, J.X.; Ye, Q.H. Analysis of the introduction and promotion effect of Sonneratia apetala. Guangdong For. Sci. Technol. 2000, 16, 6–10. [Google Scholar]
- Hong, P.; Wen, Y.; Xiong, Y.; Diao, L.; Gu, X.; Feng, H.; Yang, C.; Chen, L. Latitudinal gradients and climatic controls on reproduction and dispersal of the non-native mangrove Sonneratia apetala in China. Estuar. Coast. Shelf Sci. 2021, 248, 106749. [Google Scholar] [CrossRef]
- Powell, K.I.; Chase, J.M.; Knight, T.M. Invasive plants have scale-dependent effects on diversity by altering species-area relationships. Science 2013, 339, 316–318. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, S.; Kumar, D.; Singh, R.K.; Gupta, A.K.; Premkumar, K.; Kewat, A.K. Investigating the phenology and interactions of competitive plant species co-occurring with invasive Lantana camara in Indian Himalayan Region. Sci. Rep. 2024, 14, 400. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.F.; Yang, Q.S.; Feng, J.X.; Yang, Z.H.; Yu, C.X.; Zhang, J.; Ling, J.; Dong, J.D. Introduction of exotic species Sonneratia apetala alters diazotrophic community and stimulates nitrogen fixation in mangrove sediments. Ecol. Indic. 2022, 142, 109179. [Google Scholar] [CrossRef]
- Ren, H.; Lu, H.F.; Shen, W.J.; Huang, C.; Guo, Q.F.; Li, Z.A.; Jian, S.G. Sonneratia apetala Buch. Ham in the mangrove ecosystems of China: An invasive species or restoration species? Ecol. Eng. 2009, 35, 1243–1248. [Google Scholar] [CrossRef]
- Eysink, G.G.J.; Hatamura, E.; Schaffer-Novelli, Y. First occurrence in mangroves of South America of the exotic species Sonneratia apetala Buch.-Ham. from the Indo-Malayan region. Biota Neotrop. 2023, 23, e20231575. [Google Scholar] [CrossRef]
- Lu, W.Z.; Yang, S.C.; Chen, L.Z.; Wang, W.Q.; Du, X.N.; Wang, C.M.; Ma, Y.; Lin, G.X.; Lin, G.H. Changes in carbon pool and stand structure of a native subtropical mangrove forest after inter-planting with exotic species Sonneratia apetala. PLoS ONE 2014, 9, e91238. [Google Scholar] [CrossRef]
- Xin, K.; Zhou, Q.; Arndt, S.K.; Yang, X. Invasive Capacity of the Mangrove Sonneratia apetala in Hainan Island, China. J. Trop. For. Sci. 2013, 25, 70–78. [Google Scholar]
- Huang, X.F.; Feng, J.X.; Yang, Q.S.; Chen, L.X.; Zhang, J.; Yang, B.; Tang, X.Y.; Yu, C.X.; Ling, J.; Dong, J.D. High site elevation enhanced nitrogen fixation and the stability of diazotrophic community in planted Sonneratia apetala mangrove sediments. Appl. Soil Ecol. 2023, 191, 105059. [Google Scholar] [CrossRef]
- Zhang, Y.; Xin, K.; Liao, B.W.; Sheng, N.; Ai, X.H. The characteristics of pods and seeds of liana species Derris trifoliata and their relationship with environmental factors in Guangdong, China. Ecol. Indic. 2021, 129, 107930. [Google Scholar] [CrossRef]
- Xu, Y.W.; Liao, B.W.; Jiang, Z.M.; Xin, K.; Xiong, Y.M.; Zhang, Y. Examining the Differences between Invasive Sonneratia apetala and Native Kandelia obovata for Mangrove Restoration: Soil Organic Carbon, Nitrogen, and Phosphorus Content and Pools. J. Coast. Res. 2021, 37, 708–715. [Google Scholar] [CrossRef]
- Teng, Z.Z.; Lin, X.B. Sediment nitrates reduction processes affected by non-native Sonneratia apetala plantation in South China. Sci. Total Environ. 2024, 906, 167523. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.X.; Feng, J.X.; Liu, K.; Wang, G.; Zhu, Y.H.; Cheng, H.; Guan, D.S. Changes of ecosystem carbon stock following the plantation of exotic mangrove Sonneratia apetala in Qi’ao Island, China. Sci. Total Environ. 2020, 717, 137142. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liang, F.P.; Yang, Y.; Li, J.W.; Zhang, J.W.; Zhang, S.J.; Bai, H.; Liu, Q.; Zhong, C.R.; Li, L. Allelopathic effects of leachates from two alien mangrove species, Sonneratia apetala and laguncularia racemosa on seed germination, seedling growth and antioxidative activity of native mangrove species sonneratia caseolaris. Allelopath. J. 2018, 44, 119–129. [Google Scholar] [CrossRef]
- Jiang, Z.M.; Guan, W.; Xiong, Y.M.; Li, M.; Chen, Y.J.; Liao, B.W. Interactive Effects of Intertidal Elevation and Light Level on Early Growth of Five Mangrove Species under Sonneratia apetala Buch. Hamplantation Canopy: Turning Monocultures to Mixed Forests. Forests 2019, 10, 83. [Google Scholar] [CrossRef]
- Zhu, D.H.; Hui, D.F.; Huang, Z.J.; Qiao, X.T.; Tong, S.; Wang, M.Q.; Yang, Q.; Yu, S.X. Comparative impact of light and neighbor effect on the growth of introduced species Sonneratia apetala and native mangrove species in China: Implications for restoration. Restor. Ecol. 2022, 30, e13522. [Google Scholar] [CrossRef]
- Chen, B.B.; Liu, T.T.; Yang, Z.Y.; Yang, S.X.; Chen, J.H. PacBio Full-Length Transcriptome Sequencing Reveals the Mechanism of Salt Stress Response in Sonneratia apetala. Plants 2023, 12, 3849. [Google Scholar] [CrossRef]
- Nasrin, S.; Hossain, M.; Rahman, M.M. Adaptive responses to salinity: Nutrient resorption efficiency of Sonneratia apetala (Buch.-Ham.) along the salinity gradient in the Sundarbans of Bangladesh. Wetl. Ecol. Manag. 2019, 27, 343–351. [Google Scholar] [CrossRef]
- Rahman, M.S.; Sass-Klaassen, U.; Zuidema, P.A.; Chowdhury, M.Q.; Beeckman, H. Salinity drives growth dynamics of the mangrove tree Sonneratia apetala Buch. -Ham. in the Sundarbans, Bangladesh. Dendrochronologia 2020, 62, 125711. [Google Scholar] [CrossRef]
- Simlai, A.; Gangwar, A.; Sarthaki, G.; Roy, A. Antimicrobial and Antioxidative Activities in the Stem Extracts of Derris trifoliata, a Mangrove Shrub. J. Pharm. Res. Int. 2017, 17, 1–10. [Google Scholar] [CrossRef]
- Nguyen, H.T.T.; Hardy, G.E.S.; Le, T.V.; Nguyen, H.Q.; Le, D.H.; Nguyen, T.V.; Dell, B. Mangrove Dieback and Leaf Disease in Sonneratia apetala and Sonneratia caseolaris in Vietnam. Forests 2021, 12, 1273. [Google Scholar] [CrossRef]
- Wu, B.; Geng, S.L.; Shu, B. Genetic variation and the conservation of isolated populations of Derris trifoliata (Leguminosae), a mangrove-associated vine, in southern China. Biochem. Syst. Ecol. 2012, 40, 118–125. [Google Scholar] [CrossRef]
- Li, R.L.; Chai, M.W.; Li, R.Y.; Xu, H.L.; He, B.; Qiu, G.Y. Influence of introduced Sonneratia apetala on nutrients and heavy metals in intertidal sediments, South China. Environ. Sci. Pollut. Res. 2017, 24, 2914–2927. [Google Scholar] [CrossRef]
- Yang, X.L.; Hu, C.Y.; Wang, B.; Lin, H.; Xu, Y.P.; Guo, H.; Liu, G.Z.; Ye, J.Q.; Gao, D.Z. Sediment nitrogen mineralization and immobilization affected by non-native Sonneratia apetala plantation in an intertidal wetland of South China. Environ. Pollut. 2022, 305, 119289. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data | Year | Data Source | Data Download Website or Reference |
|---|---|---|---|
| Bioclimatic factors | 1970–2000 | The World Climate Database | www.worldclim.org (accessed on accessed on 25 June 2022) |
| Terrain data | 2022 | The ETOP01 terrain elevation and ocean seabed terrain data released by the United States Geophysical Center | https://www.ngdc.noaa.gov/mgg/global/global.html (accessed on 14 July 2022) |
| Sea surface temperature data | 2020 | National Environmental Information Center of the Oceanic and Atmospheric Administration of the United States (1981–2020 SST data) | https://ftp.emc.ncep.noaa.gov/cmb/sst/oisst_v2/ (accessed on 14 July 2022) |
| Salinity data | 2020 | The marine salinity products of the Institute of Atmospheric Physics, Chinese Academy of Sciences. | http://159.226.119.60/cheng/ (accessed on 2 February 2022) |
| Land-use data | 2020 | The ESRI 10-m cover data (2020) in the GEE | https://livingatlas.arcgis.com/landcover/ (accessed on 24 June 2022) |
| Data Type | Variable | Description | Unit |
|---|---|---|---|
| Bioclimatic | Bio2 | Mean diurnal range [mean of monthly (max. temp–min. temp)] | °C × 10 |
| Bio3 | Isothermality (BIO2/BIO7) (×100) | % | |
| Bio5 | Maximum temperature of warmest month | °C × 10 | |
| Bio6 | Minimum temperature of the coldest month | °C × 10 | |
| Bio10 | Mean temperature of the warmest quarter | °C × 10 | |
| Bio15 | Precipitation seasonality (coefficient of variation) | % | |
| Bio18 | Precipitation in the warmest quarter | mm | |
| Bio19 | Precipitation in the coldest quarter | mm | |
| Terrain | Elevation | Topographic elevation | m |
| WTI | Wetland index | -- | |
| Ocean salinity | C_sss | Mean sea surface salinity in the coldest season | ‰ |
| W_sss | Mean sea surface salinity in the warmest season | ‰ | |
| Sea surface temperature | C_sst | Mean sea surface temperature in the coldest season | °C |
| W_sst | Mean SST in the warmest season | °C | |
| Substrate type | Substrate | Substrate type | -- |
| Land-use data | Land-use | Land use type | -- |
| Host Tree | Vegetation Type | Average Height and Crown Diameter of Adult Plants (m) | Coverage Area | Coverage Time (a) | Seed Output (ind./m2) | Maximum Base Diameter (mm) | Damage Degrees |
|---|---|---|---|---|---|---|---|
| Cyperus malaccensis | herbaceous | 1.60 c, ≤1 | Small | ≤1 | 0 c | 12.3 | Mild |
| Phragmites australis | herbaceous | 1.77 c, ≤1 | Small | ≤1 | 1.60 c | 5.3 | Mild |
| Acanthus ilicifolius | shrub | 1.11 c, ≤1 | Moderate | ≤3 | 1.50 c | 11.1 | Moderate |
| Aegiceras corniculatum | shrub | 1.80 c, 2–3 | Big | ≤3 | 10.50 c | 15.2 | Serious |
| Kandelia obovata | tree | 1.93 c, 2–3 | Moderate | ≤5 | 21.55 bc | 15.9 | Moderate |
| Excoecaria agallocha | tree | 3.20 b, 2–3 | Moderate | ≤5 | 53.17 b | 16.8 | Moderate |
| Sonneratia apetala | tree | 7.86 a, 4–5 | Big | ≥5 | 97.43 a | 28.4 | Serious |
| Sonneratia apetala | Number of S. apetala | Damage Percentage | |||||
|---|---|---|---|---|---|---|---|
| All | Mild Damage | Moderate Damage | Damage | Mild Damage | Moderate Damage | Damage | |
| Adult | 534 | 234 | 88 | 322 | 43.82% | 27.33% | 60.30% |
| Seedling | 97 | 14 | 0 | 14 | 14.43% | 0.00% | 14.43% |
| Plant Species | Optimal Suitable Area (hm2) | Medium Suitable Area (hm2) |
|---|---|---|
| D. trifoliata | 3000 | 11,900 |
| S. apetala | 1800 | 14,000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Li, L.; Xue, Y.; Liang, Q.; Tao, Y.; Wu, H.; Jiang, W. Phenomenon and Mechanisms of Sonneratia apetala Introduction and Spread Promoting Excessive Growth of Derris trifoliata. Forests 2024, 15, 525. https://doi.org/10.3390/f15030525
Liu W, Li L, Xue Y, Liang Q, Tao Y, Wu H, Jiang W. Phenomenon and Mechanisms of Sonneratia apetala Introduction and Spread Promoting Excessive Growth of Derris trifoliata. Forests. 2024; 15(3):525. https://doi.org/10.3390/f15030525
Chicago/Turabian StyleLiu, Wenai, Lifeng Li, Yunhong Xue, Qiuxia Liang, Yancheng Tao, Huiying Wu, and Weiguo Jiang. 2024. "Phenomenon and Mechanisms of Sonneratia apetala Introduction and Spread Promoting Excessive Growth of Derris trifoliata" Forests 15, no. 3: 525. https://doi.org/10.3390/f15030525
APA StyleLiu, W., Li, L., Xue, Y., Liang, Q., Tao, Y., Wu, H., & Jiang, W. (2024). Phenomenon and Mechanisms of Sonneratia apetala Introduction and Spread Promoting Excessive Growth of Derris trifoliata. Forests, 15(3), 525. https://doi.org/10.3390/f15030525