
Significant Shifts in Predominant Plant Dispersal Modes in Pine Forests of the Southern Urals (Russia): Responses to Technogenic Pollution and Ground Fires

Abstract
1. Introduction
2. Materials and Methods
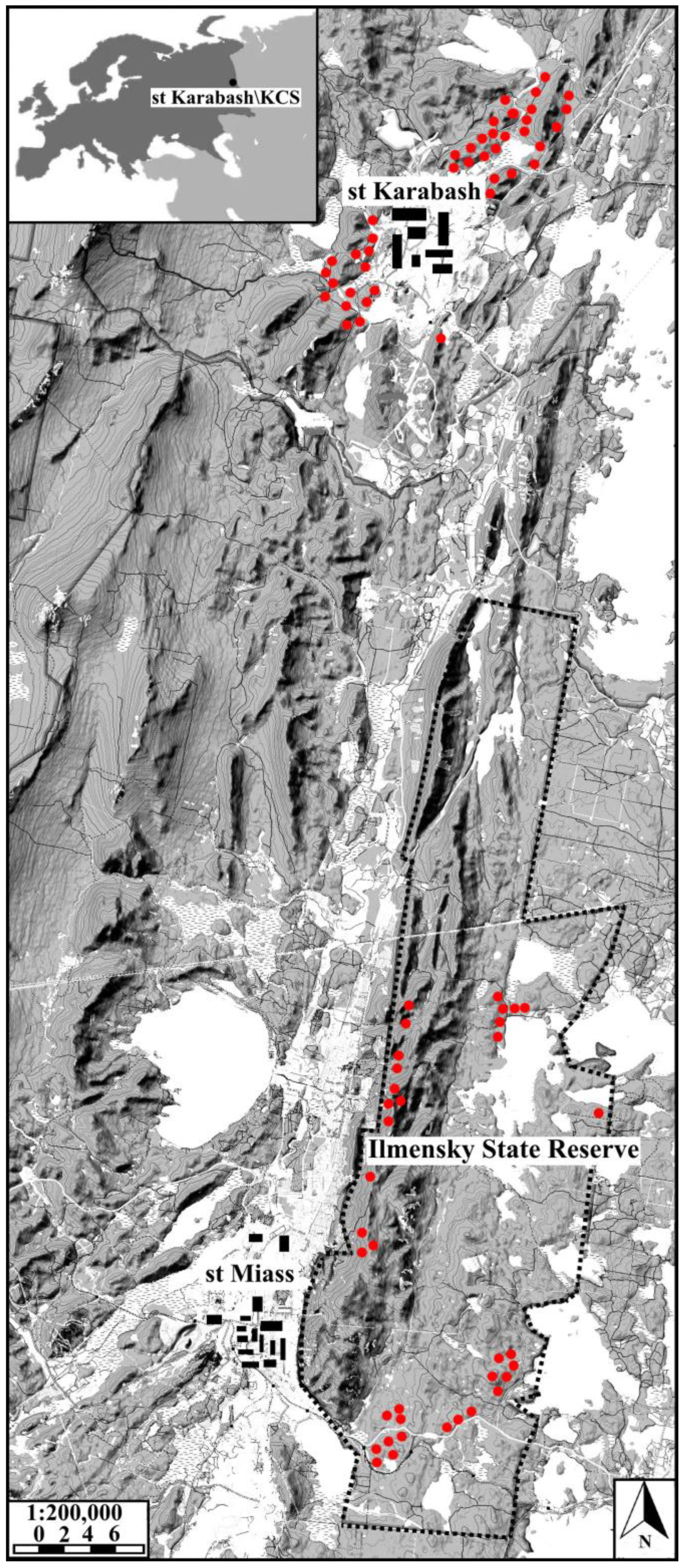
2.1. Study Area
2.2. Sample Plots
2.3. Characteristics of Plant Species and Plant Communities
2.4. Determination of Technogenic Impact
2.5. Data Analysis
3. Results
3.1. The Ratio of Results Obtained Using Traditional and New Classifications
3.2. Selection of a Combination of Predictors Explaining the Variability of Herb–Shrub Layer Diversity Estimates
3.3. Direction and Strength of Influence of N years no fire and Log(PI) on Diversity Parameters
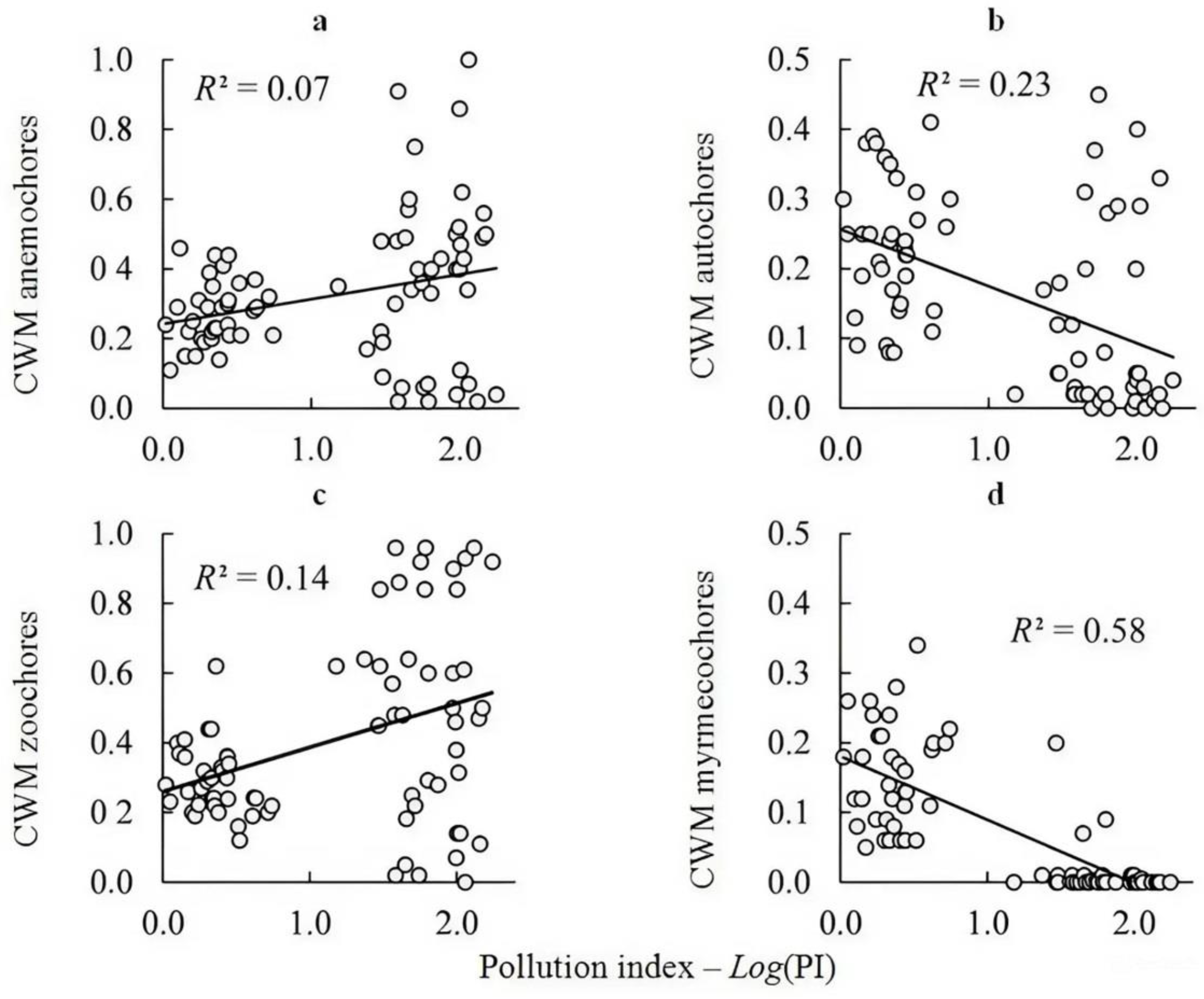
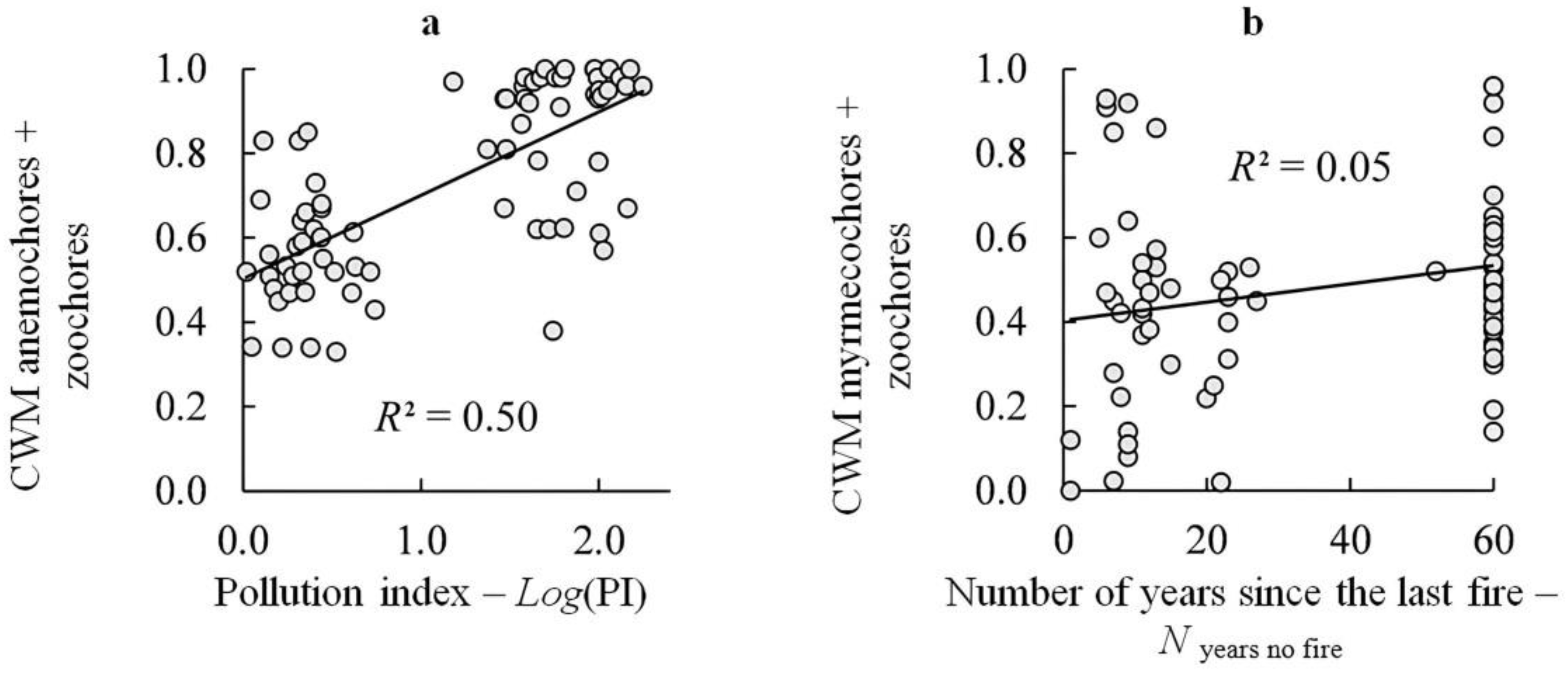
3.4. The Ratio of Species Groups with Different Diaspore Dispersal Distances in Connection with Pollution
- -
- Autochores and myrmecochores—seeds disperse not far, from tens of centimeters to tens of meters;
- -
- Anemochores and zoochores—seeds disperse far, from tens to hundreds of meters, possibly further.
3.5. The Ratio of Species Groups with Different Diaspore Dispersal Agents in Relation to the Time Since the Last Fire Disturbances
- -
- Seeds are dispersed without animals—anemochores and autochores;
- -
- Seeds are dispersed by invertebrates and vertebrates—myrmecochores and zoochores.
4. Discussion
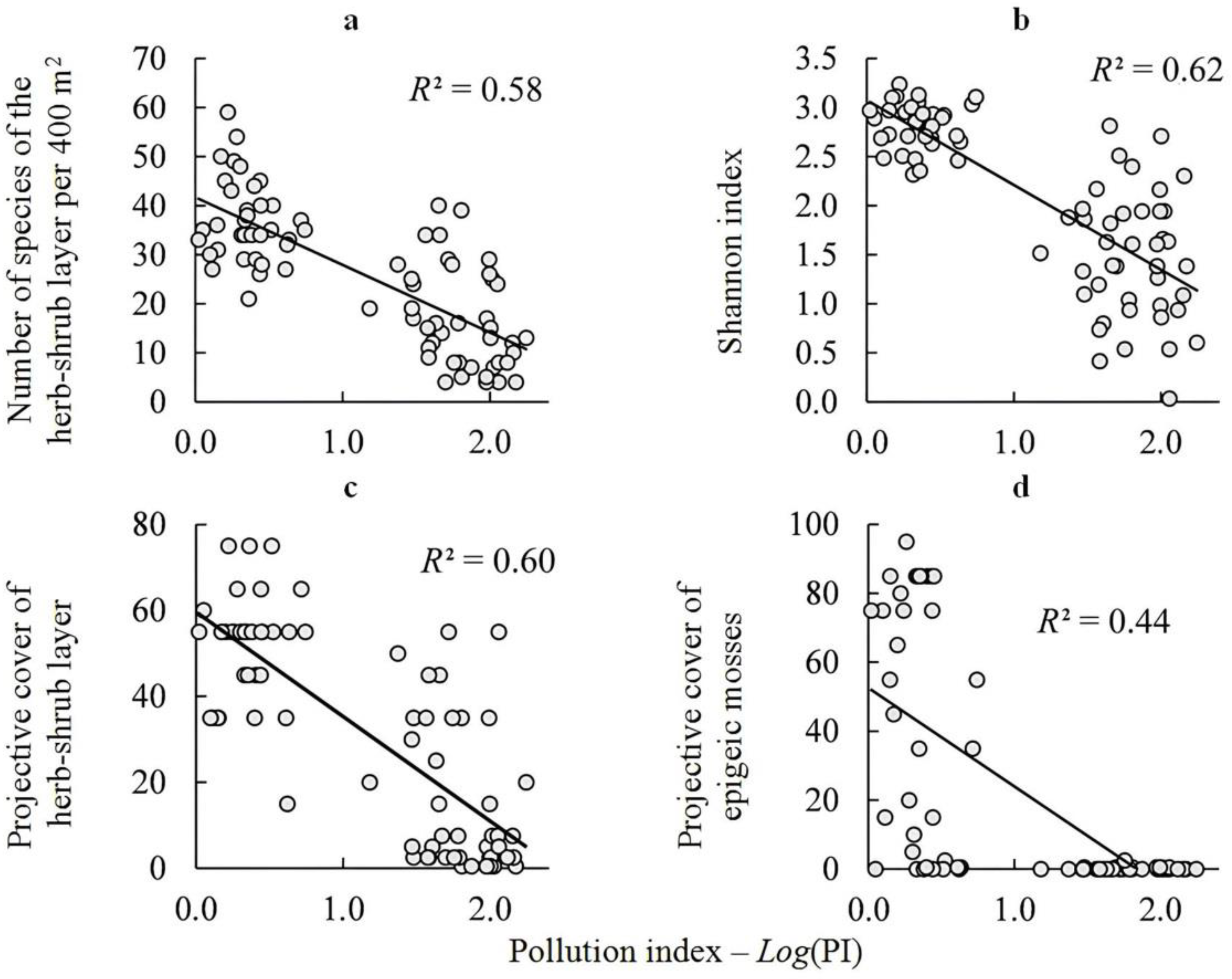
4.1. Impact of Pollution
4.2. Impact of Fires
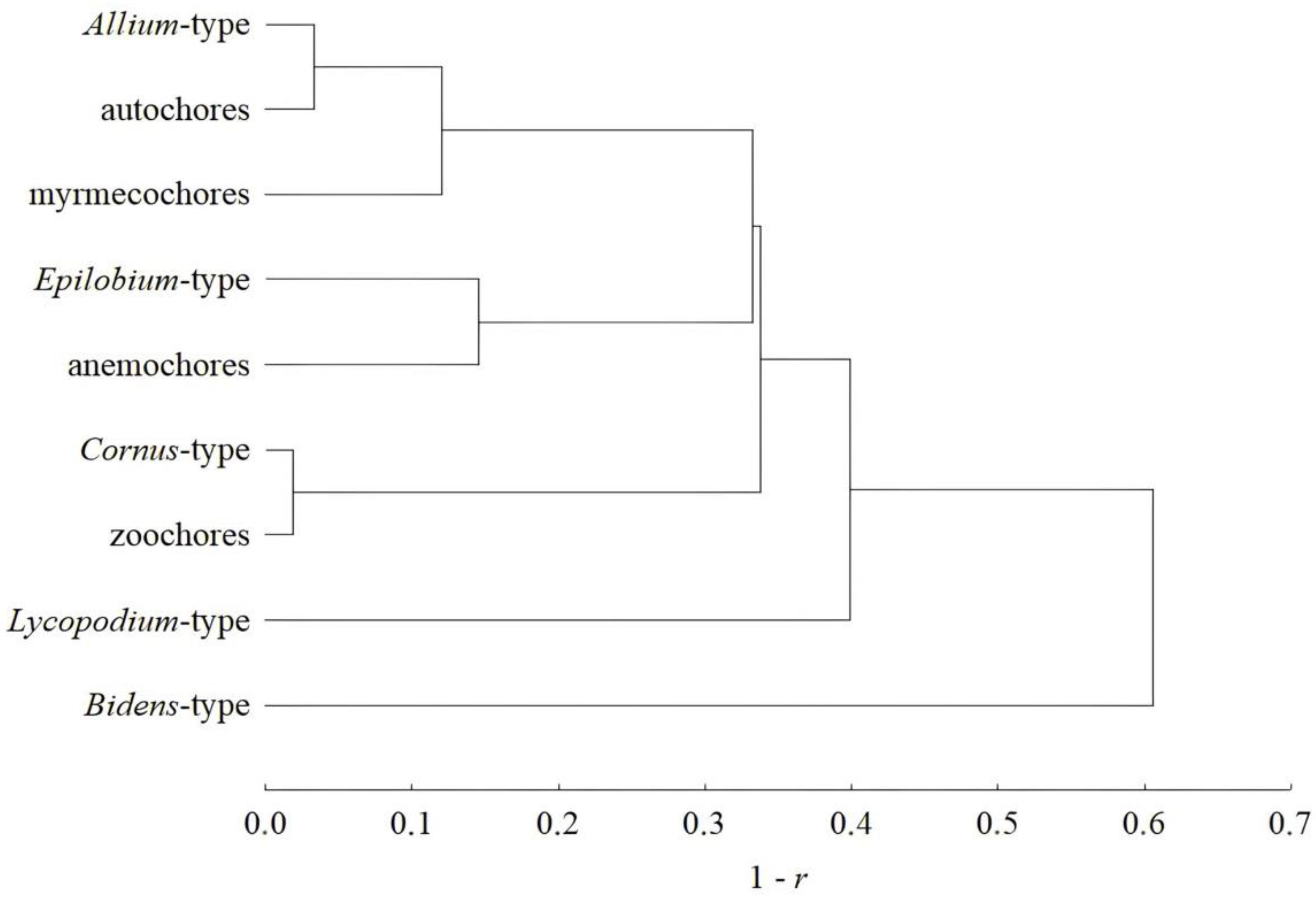
4.3. The Correspondence Between “Dispersal Syndromes” and “Dispersal Strategies”
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Willson, M.F.; Traveset, A. The ecology of seed dispersal. In Seeds: The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CABI Publishing: Wallingford, UK, 2000; Chapter 4; pp. 85–110. [Google Scholar] [CrossRef]
- Huanca Nuñez, N.; Chazdon, R.L.; Russo, S.E. Seed rain–successional feedbacks in wet tropical forests. Ecology 2021, 102, e03362. [Google Scholar] [CrossRef] [PubMed]
- Wendt, A.L.; Chazdon, R.L.; Ramirez, O.V. Successional trajectories of seed dispersal mode and seed size of canopy tree species in wet tropical forests. Front. For. Glob. Chang. 2022, 5, 946541. [Google Scholar] [CrossRef]
- Treep, J.; de Jager, M.; Bartumeus, F.; Soons, M.B. Seed dispersal as a search strategy: Dynamic and fragmented landscapes select for multi-scale movement strategies in plants. Mov. Ecol. 2021, 9, 4. [Google Scholar] [CrossRef] [PubMed]
- Vittoz, P.; Engler, R. Seed dispersal distances: A typology based on dispersal modes and plant traits. Bot. Helv. 2007, 117, 109–124. [Google Scholar] [CrossRef]
- Lososová, Z.; Axmanová, I.; Chytrý, M.; Midolo, G.; Abdulhak, S.; Karger, D.N.; Renaud, J.; Van Es, J.; Vittoz, P.; Thuiller, W. Seed dispersal distance classes and dispersal modes for the European flora. Glob. Ecol. Biogeogr. 2023, 32, 1485–1494. [Google Scholar] [CrossRef]
- McClanahan, T.R. The effect of a seed source on primary succession in a forest ecosystem. Vegetatio 1986, 65, 175–178. [Google Scholar] [CrossRef]
- Duncan, R.S.; Chapman, C.A. Seed dispersal and potential forest succession in abandoned agriculture in tropical Africa. Ecol. Appl. 1999, 9, 998–1008. [Google Scholar] [CrossRef]
- Dovčiak, M.; Frelich, L.E.; Reich, P.B. Pathways in old-field succession to white pine: Seed rain, shade, and climate effects. Ecol. Monogr. 2005, 75, 363–378. [Google Scholar] [CrossRef]
- Reid, J.L.; Holl, K.D. Arrival ≠ Survival. Restor. Ecol. 2013, 21, 153–155. [Google Scholar] [CrossRef]
- Yang, X.; Yan, C.; Zhao, Q.; Holyoak, M.; Fortuna, M.A.; Bascompte, J.; Jansen, P.A.; Zhang, Z. Ecological succession drives the structural change of seed-rodent interaction networks in fragmented forests. For. Ecol. Manag. 2018, 419–420, 42–50. [Google Scholar] [CrossRef]
- Dent, D.H.; Estrada-Villegas, S. Uniting niche differentiation and dispersal limitation predicts tropical forest succession. Trends Ecol. Evol. 2021, 36, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Coomes, D.A.; Grubb, P.J. Colonization, tolerance, competition and seed-size variation within functional groups. Trends Ecol. Evol. 2003, 18, 283–291. [Google Scholar] [CrossRef]
- Palma, A.C.; Goosem, M.; Fensham, R.J.; Goosem, S.; Preece, N.D.; Stevenson, P.R.; Laurance, S.G.W. Dispersal and recruitment limitations in secondary forests. J. Veg. Sci. 2021, 32, e12975. [Google Scholar] [CrossRef]
- Benedicto-Royuela, J.; Costa, J.M.; Heleno, R.; Silva, J.S.; Freitas, H.; Lopes, P.; Mendes, S.B.; Timóteo, S. What is the value of biotic seed dispersal in post-fire forest regeneration? Conserv. Lett. 2023, 17, e12990. [Google Scholar] [CrossRef]
- Dehaudt, B.; Bruce, T.; Deblauwe, V.; Ferraz, A.; Gardner, B.; Bibila, T.G.B.; LeBreton, M.; Mempong, G.; Njabo, K.; Nkengbeza, S.N.; et al. Divergent seed dispersal outcomes: Interactions between seed, disperser, and forest traits. Ecology 2024, 105, e4409. [Google Scholar] [CrossRef]
- Kozlov, M.V.; Zvereva, E.L.; Zverev, V.E. Impacts of Point Polluters on Terrestrial Biota; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; London, UK; New York, NY, USA, 2009. [Google Scholar]
- Chibilev, A.A.; Veselkin, D.V.; Kuyantseva, N.B.; Chashchina, O.E.; Dubinin, A.E. Dynamics of forest fires and climate in Ilmen nature reserve, 1948–2013. Dokl. Earth Sci. 2016, 468, 619–622. [Google Scholar] [CrossRef]
- Veselkin, D.; Pustovalova, L.; Kuyantseva, N.; Mumber, A. Trends in forest fire occurrence in the Ilmensky nature reserve, Southern Urals, Russia, between 1948 and 2014. Forests 2022, 13, 528. [Google Scholar] [CrossRef]
- Zvereva, E.L.; Toivonen, E.; Kozlov, M.V. Changes in species richness of vascular plants under the impact of air pollution: A global perspective. Glob. Ecol. Biogeogr. 2007, 17, 305–319. [Google Scholar] [CrossRef]
- Chernenkova, T.V.; Kabirov, R.R.; Basova, E.V. Regeneration successions of northern taiga spruce forests under reduction of aerotechnogenic impact. Contemp. Probl. Ecol. 2011, 4, 742–757. [Google Scholar] [CrossRef]
- Chernenkova, T.V. Biodiversity of forest vegetation under industrial pollution. Russ. J. Ecol. 2014, 45, 1–10. [Google Scholar] [CrossRef]
- Trubina, M.R.; Vorobeichik, E.L.; Khantemirova, E.V.; Bergman, I.E.; Kaigorodova, S.Y. Dynamics of forest vegetation after the reduction of industrial emissions: Fast recovery or continued degradation? Dokl. Biol. Sci. 2014, 458, 302–305. [Google Scholar] [CrossRef] [PubMed]
- Richter, C.; Rejmánek, M.; Miller, J.E.D.; Welch, K.R.; Weeks, J.; Safford, H. The species diversity × fire severity relationship is hump-shaped in semiarid yellow pine and mixed conifer forests. Ecosphere 2019, 10, e02882. [Google Scholar] [CrossRef]
- Strand, E.K.; Satterberg, K.L.; Hudak, A.T.; Byrne, J.; Khalyani, A.H.; Smith, A.M.S. Does burn severity affect plant community diversity and composition in mixed conifer forests of the United States Intermountain West one decade post fire? Fire Ecol. 2019, 15, 25. [Google Scholar] [CrossRef]
- Geraskina, A.P.; Tebenkova, D.N.; Ershov, D.V.; Ruchinskaya, E.V.; Sibirtseva, N.V.; Lukina, N.V. Wildfires as a factor of the loss of biodiversity and functions of forest ecosystems. For. Sci. Issues 2021, 4, 82, (In Russian with English Abstract). [Google Scholar] [CrossRef]
- Kovaleva, N.M.; Ivanova, G.A. Recovery of ground vegetation at the initial stage of fire succession. Contemp. Probl. Ecol. 2013, 6, 162–169. [Google Scholar] [CrossRef]
- Braslavskaya, T.Y.; Geraskina, A.P.; Aleinikov, A.A.; Sibgatullin, R.Z.; Belyaeva, N.V.; Ukhova, N.L.; Korotkov, V.N.; Shilov, D.S.; Lugovaya, D.L.; Smirnova, O.V. Long-term effect of wildfires on vascular plant and soil invertebrate diversity in primary fir-spruce forests of the Ural Mountains (North Eurasia). Russ. J. Ecosyst. Ecol. 2022, 7, 1–26. [Google Scholar] [CrossRef]
- Eales, J.; Haddaway, N.R.; Bernes, C.; Cooke, S.J.; Jonsson, B.G.; Kouki, J.; Petrokofsky, G.; Taylor, J.J. What is the effect of prescribed burning in temperate and boreal forest on biodiversity, beyond pyrophilous and saproxylic species? A systematic review. Environ. Evid. 2018, 7, 19. [Google Scholar] [CrossRef]
- Cote, J.; Bestion, E.; Jacob, S.; Travis, J.; Legrand, D.; Baguette, M. Evolution of dispersal strategies and dispersal syndromes in fragmented landscapes. Ecography 2017, 40, 56–73. [Google Scholar] [CrossRef]
- Warneke, C.R.; Caughlin, T.T.; Damschen, E.I.; Haddad, N.M.; Levey, D.J.; Brudvig, L.A. Habitat fragmentation alters the distance of abiotic seed dispersal through edge effects and direction of dispersal. Ecology 2022, 103, e03586. [Google Scholar] [CrossRef]
- Trubina, M.R. Vulnerability to copper smelter emissions in species of the herb–dwarf shrub layer: Role of differences in the type of diaspore dispersal. Russ. J. Ecol. 2020, 51, 107–117. [Google Scholar] [CrossRef]
- Howe, H.F.; Smallwood, J. Ecology of seed dispersal. Annu. Rev. Ecol. Syst. 1982, 13, 201–228. [Google Scholar] [CrossRef]
- Céspedes, B.; Torres, I.; Luna, B.; Pérez, B.; Moreno, J.M. Soil seed bank, fire season, and temporal patterns of germination in a seeder-dominated Mediterranean shrubland. Plant Ecol. 2012, 213, 383–393. [Google Scholar] [CrossRef]
- Hartnett, D.C.; Richardson, D.R. Population biology of Bonamia grandiflora (Convolvulaceae): Effects of fire on plant and seed bank dynamics. Am. J. Bot. 1989, 76, 361–369. [Google Scholar] [CrossRef]
- Lamont, B.B.; Runciman, H.V. Fire may stimulate flowering, branching, seed production and seedling establishment in two kangaroo paws (Haemodoraceae). J. Appl. Ecol. 1993, 30, 256–264. [Google Scholar] [CrossRef]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef]
- Usoltsev, V.A.; Vorobeychik, E.L.; Bergman, I.E. Biological Productiviti of Ural Forests under Conditions of Air Pollutions: An Investigation of a System of Regularities; Ural’skiy Gosudarstvennyy Lesotekhnicheskiy Universitet: Yekaterinburg, Russia, 2012. (In Russian) [Google Scholar]
- Mikryukov, V.S.; Dulya, O.V.; Vorobeichik, E.L. Diversity and spatial structure of soil fungi and arbuscular mycorrhizal fungi in forest litter contaminated with copper smelter emissions. Water Air Soil Pollut. 2015, 226, 114. [Google Scholar] [CrossRef]
- Mikryukov, V.S.; Dulya, O.V. Contamination-induced transformation of bacterial and fungal communities in spruce-fir and birch forest litter. Appl. Soil Ecol. 2017, 114, 111–122. [Google Scholar] [CrossRef]
- Veselkin, D.V.; Chashchina, O.E.; Kuyantseva, N.B.; Mumber, A.G. Stable carbon and nitrogen isotopes in woody plants and herbs near the large copper smelting plant. Geochem. Int. 2019, 57, 575–582. [Google Scholar] [CrossRef]
- Veselkin, D.; Kuyantseva, N.; Mumber, A.; Molchanova, D.; Kiseleva, D. δ15N in birch and pine leaves in the vicinity of a large copper smelter indicating a change in the conditions of their soil nutrition. Forests 2022, 13, 1299. [Google Scholar] [CrossRef]
- Plants of the World Online. Available online: https://powo.science.kew.org (accessed on 9 July 2024).
- Ovesnov, S.A.; Efimik, E.G. Biodiversity and Ecology of Higher Plants: Textbook. Study Guide; Perm State University: Perm, Russia, 2009. (In Russian) [Google Scholar]
- Dronin, G.V.; Rakov, N.S. Ecological and florogenetic aspects of coenoflora of pine forest near village Staroe Chirkovo (Ulyanovsk Pre-Volga region). Phytodiversity East. Eur. 2013, 7, 29–62. (In Russian) [Google Scholar]
- Levina, R.E. Means of Dispersal of Fruits and Seeds; Moscow State University: Moscow, Russia, 1957. (In Russian) [Google Scholar]
- Sádlo, J.; Chytrý, M.; Pergl, J.; Pyšek, P. Plant dispersal strategies: A new classification based on themultiple dispersal modes of individual species. Preslia 2018, 90, 1–22. [Google Scholar] [CrossRef]
- Pladias—Database of the Czech Flora and Vegetation. Available online: www.pladias.cz (accessed on 24 July 2024).
- Casanoves, F.; Pla, L.; Di Rienzo, J.A.; Díaz, S. FDiversity: A software package for the integrated analysis of functional diversity. Methods Ecol. Evol. 2011, 2, 233–237. [Google Scholar] [CrossRef]
- Du Laing, G. Analysis and fractionation of trace elements in soils. In Trace Elements in Soils; Hooda, P.S., Ed.; Wiley: Chichester, UK, 2010; Chapter 4; pp. 53–80. [Google Scholar] [CrossRef]
- Vorobeychik, E.L.; Sadykov, O.F.; Farafontov, M.G. Ecological Standardization of Terrestrial Ecosystems Technogenic Pollution (Local Scale); Nauka: Yekaterinburg, Russia, 1994. (In Russian) [Google Scholar]
- Copeland, S.M.; Harrison, S.P.; Latimer, A.M.; Damschen, E.I.; Eskelinen, A.M.; Fernandez-Going, B.; Spasojevic, M.J.; Anacker, B.L.; Thorne, J.H. Ecological effects of extreme drought on Californian herbaceous plant communities. Ecol. Monogr. 2016, 86, 295–311. [Google Scholar] [CrossRef]
- Mikryukov, V.S.; Vorobeichik, E.L.; Mikhailova, I.N. Variation of the epiphytic lichen diversity in a gradient of atmospheric pollution: Do taxonomic, genetic, and functional distances between species add any information? Dokl. Biol. Sci. 2014, 454, 22–25. [Google Scholar] [CrossRef]
- Li, H.; Luo, P.; Yang, H.; Luo, C.; Xie, W.; Jia, H.; Cheng, Y.; Huang, Y. Assessing the effect of roads on mountain plant diversity beyond species richness. Front. Plant Sci. 2022, 13, 985673. [Google Scholar] [CrossRef]
- Belskaya, E.; Gilev, A.; Trubina, M.; Belskii, E. Diversity of ants (Hymenoptera, Formicidae) along a heavy metal pollution gradient: Evidence of a hump-shaped effect. Ecol. Indic. 2019, 106, 105447. [Google Scholar] [CrossRef]
- Kuyantseva, N.B.; Molchanova, D.A.; Mumber, A.G.; Veselkin, D.V. The number of seedlings from the soil seed bank of pine forests with differently dated fires near the Karabash copper smelter. Russ. J. Ecol. 2024, 55, 241–252. [Google Scholar] [CrossRef]
- Zharkova, D.A.; Kuyantseva, N.B.; Mumber, A.G.; Veselkin, D.V. Changes in the diversity of the herb-shrub layer of pine forests caused by severe pollution and fire disturbances in the Southern Urals. Biol. Bull. 2024; in print. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dispersal Syndrome/Strategy | Total Number of Species | Mean Value per Relevé in the Entire Set of 77 Relevés (±SE) | Typical Representatives | |
|---|---|---|---|---|
| Cover | CWM | |||
| Dispersal syndromes according to traditional classification | ||||
| Anemochores | 58 | 17.6 ± 1.8 | 0.32 ± 0.02 | Calamagrostis arundinacea (L.) Roth, Sanguisorba officinalis L., Orthilia secunda (L.) House, Lilium pilosiusculum (Freyn) Miscz. |
| Autochores | 71 | 16.9 ± 2.1 | 0.16 ± 0.01 | Brachypodium pinnatum (L.) Beauv., Stachys officinalis (L.) Trevis., Lathyrus pisiformis L., Trifolium medium L., Seseli libanotis (L.) Koch, Vicia cracca L., Geranium pseudosibiricum J. Mayer. |
| Zoochores | 16 | 18.7 ± 1.6 | 0.41 ± 0.03 | Galium boreale L., Vaccinium myrtillus L., Polygonatum odoratum (Mill.) Druce, Rubus saxatilis L., Vaccinium vitis-idaea L. |
| Myrmecochores | 13 | 8.3 ± 1.3 | 0.08 ± 0.01 | Luzula pilosa (L.) Willd., Carex montana L., Pulmonaria mollis Wulfen ex Hornem. |
| Dispersal strategies under the new classification | ||||
| Allium-type | 91 | 28.9 ± 3.6 | 0.33 ± 0.03 | Adenophora lilifolia (L.) A. DC., Brachypodium pinnatum (L.) Beauv., Galium boreale L., Sanguisorba officinalis L. |
| Bidens-type | 3 | 0.2 ± 0.1 | 0.002 ± 0.001 | Agrimonia pilosa Ledeb., Geum rivale L., Geum urbanum L. |
| Cornus-type | 8 | 16.1 ± 1.3 | 0.39 ± 0.03 | Polygonatum odoratum (Mill.) Druce, Rubus saxatilis L., Vaccinium vitis-idaea L. |
| Epilobium-type | 15 | 9.2 ± 1.2 | 0.19 ± 0.02 | Calamagrostis arundinacea (L.) Roth, Hieracium umbellatum L., Trommsdorffia maculata (L.) Bernh. |
| Lycopodium-type | 19 | 4.8 ± 0.7 | 0.09 ± 0.01 | Orthilia secunda (L.) House, Pyrola rotundifolia L., Chimaphila umbellata (L.) W.P.C. Barton, Pteridium aquilinum (L.) Kuhn. |
| Characteristics | Number of Years Since the Last Fire—N years no fire | Pollution Index—Log(PI) | Predictor Combination | |
|---|---|---|---|---|
| Without Interaction: N years no fire + Log(PI) | with Interaction: N years no fire + Log(PI) + N years no fire × Log(PI) | |||
| State characteristics of the herb–shrub and moss layers | ||||
| Number of species of the herb–shrub layer per 400 m2 | 624.81 | 560.84 | 562.83 | 564.81 |
| Shannon index | 193.57 | 120.47 | 122.39 | 123.99 |
| Projective cover of | ||||
| Herb–shrub layer | 709.98 | 646.13 | 648.13 | 650.12 |
| epigeic mosses | 754.96 | 710.67 | 705.72 | 706.91 |
| Functional diversity: dispersal syndromes according to the traditional classification | ||||
| Projective cover of species groups | ||||
| Anemochores | 648.16 | 624.16 | 623.75 | 623.93 |
| Autochores | 669.34 | 608.86 | 609.06 | 607.81 |
| Zoochores | 624.68 | 540.41 | 541.23 | 542.68 |
| Myrmecochores | 589.15 | 534.79 | 536.35 | 538.29 |
| CWM | ||||
| Anemochores | −30.41 | −31.48 | −33.24 | −31.28 |
| Autochores | −91.60 | −110.74 | −109.01 | −111.90 |
| Zoochores | 9.72 | 0.55 | −3.06 | −1.42 |
| Myrmecochores | −148.42 | −208.07 | −206.09 | −204.13 |
| FAD | 233.57 | 199.86 | 200.10 | 200.63 |
| Rao’s quadratic entropy | −33.15 | −68.71 | −67.87 | −67.72 |
| Functional diversity: dispersal strategies according to the new classification | ||||
| Projective cover of species groups | ||||
| Allium-type | 753.03 | 683.76 | 684.76 | 684.88 |
| Bidens-type | 241.53 | 243.38 | 236.97 | 236.97 |
| Cornus-type | 596.04 | 531.02 | 532.13 | 533.26 |
| Epilobium-type | 573.61 | 568.95 | 566.38 | 548.30 |
| Lycopodium-type | 508.26 | 495.18 | 497.12 | 498.31 |
| CWM | ||||
| Allium-type | 7.54 | −45.84 | −43.88 | −47.16 |
| Bidens-type | −495.24 | −493.16 | −499.06 | −499.06 |
| Cornus-type | 16.56 | 2.27 | −1.94 | −0.31 |
| Epilobium-type | −42.00 | −42.75 | −44.21 | −42.26 |
| Lycopodium-type | −113.66 | −115.00 | −113.28 | −112.69 |
| FAD | 151.23 | 132.24 | 133.87 | 135.55 |
| Rao’s quadratic entropy | −31.48 | −43.90 | −41.91 | −41.89 |
| Characteristics | Number of Years Since the Last Fire—N years no fire | Pollution Index— Log(PI) | R2adj | ||
|---|---|---|---|---|---|
| β ± SE | p | β ± SE | p | ||
| State characteristics of the herb–shrub and moss layers | |||||
| Number of species of the herb–shrub layer per 400 m2 | −0.016 ± 0.076 | 0.8361 | −0.764 ± 0.076 | <0.0001 | 0.57 |
| Shannon index | −0.035 ± 0.072 | 0.6271 | −0.790 ± 0.072 | <0.0001 | 0.61 |
| Projective cover of | |||||
| Herb–shrub layer | −0.020 ± 0.074 | 0.7841 | −0.776 ± 0.074 | <0.0001 | 0.59 |
| epigeic mosses | 0.146 ± 0.086 | 0.0937 | −0.647 ± 0.086 | <0.0001 | 0.45 |
| Functional diversity: dispersal syndromes according to the traditional classification | |||||
| Projective cover of species groups | |||||
| Anemochores | −0.193 ± 0.096 | 0.0475 | −0.571 ± 0.096 | <0.0001 | 0.32 |
| Autochores | −0.015 ± 0.077 | 0.8497 | −0.759 ± 0.077 | <0.0001 | 0.56 |
| Zoochores | 0.078 ± 0.065 | 0.2351 | −0.818 ± 0.065 | <0.0001 | 0.68 |
| Myrmecochores | 0.012 ± 0.080 | 0.8795 | −0.733 ± 0.080 | <0.0001 | 0.53 |
| CWM | |||||
| Anemochores | −0.215 ± 0.110 | 0.0542 | 0.244 ± 0.110 | 0.0297 | 0.10 |
| Autochores | −0.130 ± 0.102 | 0.2070 | −0.491 ± 0.102 | <0.0001 | 0.22 |
| Zoochores | 0.222 ± 0.106 | 0.0391 | 0.403 ± 0.106 | 0.0003 | 0.17 |
| Myrmecochores | −0.001 ± 0.076 | 0.9942 | −0.763 ± 0.076 | <0.0001 | 0.57 |
| FAD | 0.129 ± 0.090 | 0.1579 | −0.609 ± 0.090 | <0.0001 | 0.39 |
| Rao’s quadratic entropy | 0.115 ± 0.090 | 0.2039 | −0.615 ± 0.090 | <0.0001 | 0.39 |
| Functional diversity: dispersal strategies according to the new classification | |||||
| Projective cover of species groups | |||||
| Allium-type | −0.024 ± 0.072 | 0.7410 | −0.795 ± 0.072 | <0.0001 | 0.62 |
| Bidens-type | 0.175 ± 0.112 | 0.1233 | −0.201 ± 0.112 | 0.0773 | 0.06 |
| Cornus-type | 0.089 ± 0.074 | 0.2325 | −0.758 ± 0.074 | <0.0001 | 0.59 |
| Epilobium-type | −0.264 ± 0.106 | 0.0151 | −0.368 ± 0.106 | 0.0009 | 0.16 |
| Lycopodium-type | 0.012 ± 0.106 | 0.9077 | −0.416 ± 0.106 | 0.0002 | 0.15 |
| CWM | |||||
| Allium-type | −0.109 ± 0.081 | 0.1852 | −0.725 ± 0.081 | <0.0001 | 0.50 |
| Bidens-type | 0.172 ± 0.113 | 0.1304 | −0.195 ± 0.113 | 0.0870 | 0.05 |
| Cornus-type | 0.218 ± 0.102 | 0.0357 | 0.496 ± 0.102 | <0.0001 | 0.22 |
| Epilobium-type | −0.198 ± 0.111 | 0.0784 | 0.227 ± 0.111 | 0.0442 | 0.08 |
| Lycopodium-type | 0.036 ± 0.116 | 0.7553 | 0.133 ± 0.116 | 0.2561 | 0.01 |
| FAD | 0.064 ± 0.102 | 0.5308 | −0.474 ± 0.102 | <0.0001 | 0.22 |
| Rao’s quadratic entropy | 0.010 ± 0.108 | 0.9279 | −0.391 ± 0.108 | 0.0005 | 0.13 |
| Characteristics | Number of Years Since the Last Fire–N years no fire | Pollution Index— Log(PI) | R2adj | ||
|---|---|---|---|---|---|
| β ± SE | p | β ± SE | p | ||
| CWM of functional groups with different seed dispersal distances | |||||
| Autochores + myrmecochores | −0.090 ± 0.084 | 0.2872 | −0.704 ± 0.084 | <0.0001 | 0.47 |
| Anemochores + zoochores | 0.064 ± 0.082 | 0.4377 | 0.717 ± 0.082 | <0.0001 | 0.49 |
| CWM of functional groups with different seed dispersal agents | |||||
| Autochores + anemochores | −0.265 ± 0.113 | 0.0219 | −0.068 ± 0.113 | 0.5513 | 0.05 |
| Myrmecochores + zoochores | 0.244 ± 0.113 | 0.0342 | 0.146 ± 0.113 | 0.2007 | 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veselkin, D.; Kuyantseva, N.; Mumber, A.; Zharkova, D. Significant Shifts in Predominant Plant Dispersal Modes in Pine Forests of the Southern Urals (Russia): Responses to Technogenic Pollution and Ground Fires. Forests 2024, 15, 2161. https://doi.org/10.3390/f15122161
Veselkin D, Kuyantseva N, Mumber A, Zharkova D. Significant Shifts in Predominant Plant Dispersal Modes in Pine Forests of the Southern Urals (Russia): Responses to Technogenic Pollution and Ground Fires. Forests. 2024; 15(12):2161. https://doi.org/10.3390/f15122161
Chicago/Turabian StyleVeselkin, Denis, Nadezhda Kuyantseva, Aleksandr Mumber, and Darya Zharkova. 2024. "Significant Shifts in Predominant Plant Dispersal Modes in Pine Forests of the Southern Urals (Russia): Responses to Technogenic Pollution and Ground Fires" Forests 15, no. 12: 2161. https://doi.org/10.3390/f15122161
APA StyleVeselkin, D., Kuyantseva, N., Mumber, A., & Zharkova, D. (2024). Significant Shifts in Predominant Plant Dispersal Modes in Pine Forests of the Southern Urals (Russia): Responses to Technogenic Pollution and Ground Fires. Forests, 15(12), 2161. https://doi.org/10.3390/f15122161