
Effects of Different Levels of Physical Damage Combined with Fungal Induction on Agarwood Formation
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Isolation, Purification, and Identification of Endophytic Fungal from Agarwood
2.3. Plant Samples and Fungal Induction of Agarwood Formation
2.4. Histochemical Observation of Slices
2.5. Scanning Electron Microscopic Observation of Tissue Inclusions
2.6. Non-Structural Carbohydrate Content Detection
2.7. Induced Color Change Range Measurement
2.8. Extraction of Alcohol-Soluble Essential Oils of Agarwood
2.9. Component Analysis of Essential Oils
2.10. Statistics Analysis
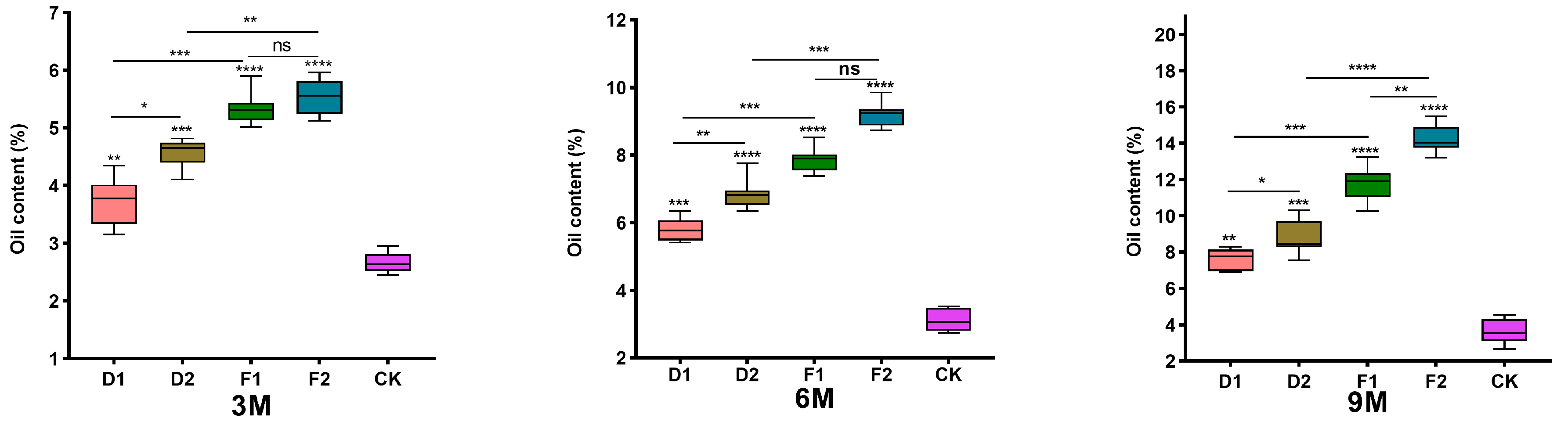
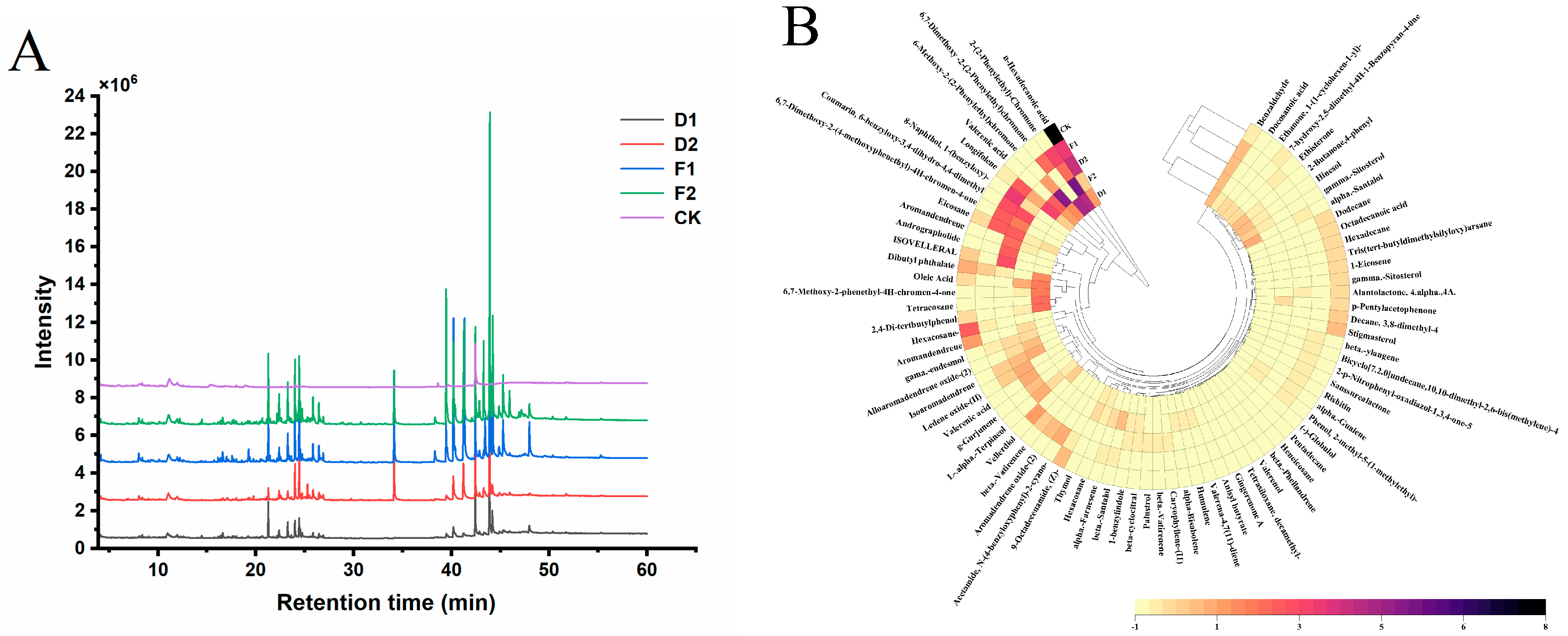
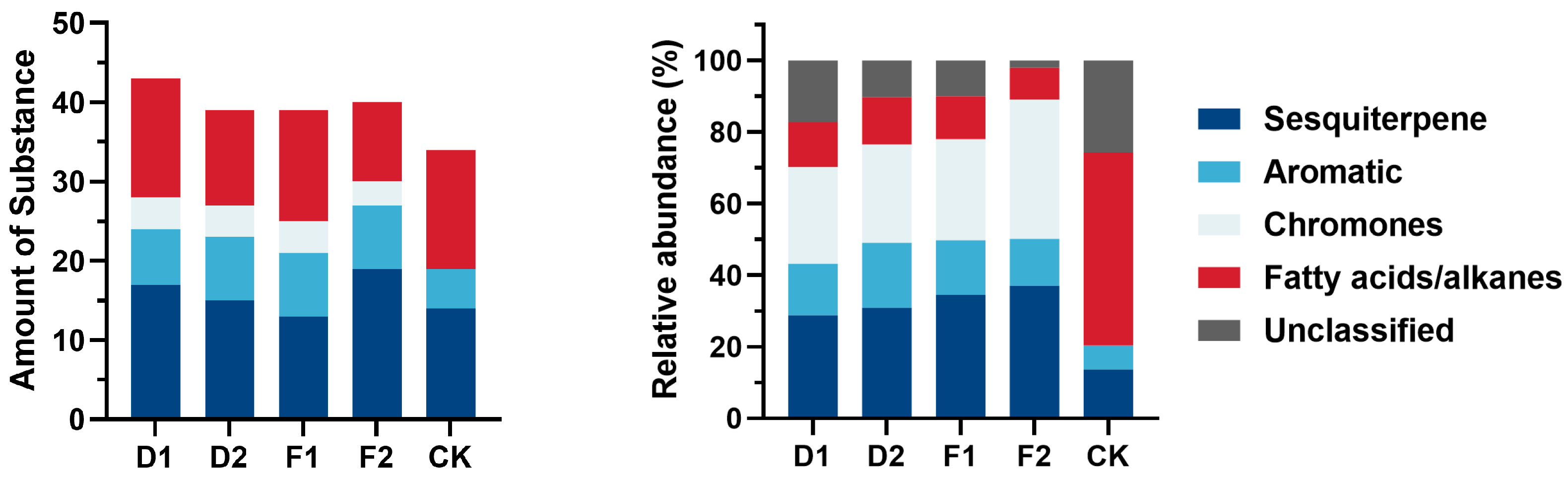
3. Results
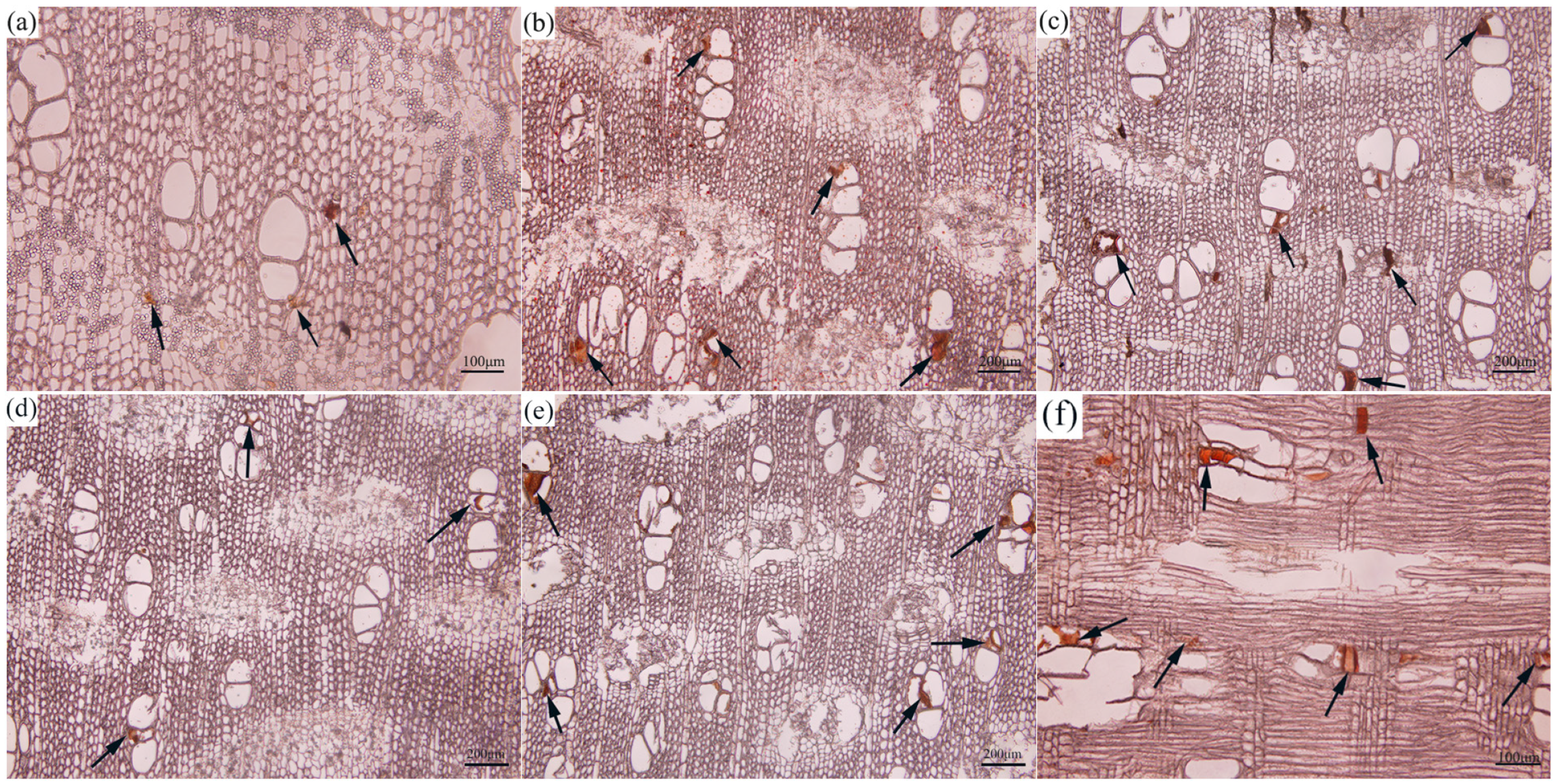
3.1. Histological Formation Process of Agarwood Resin in the Agarwood Layers
3.1.1. Histological Formation Process of Agarwood Resin
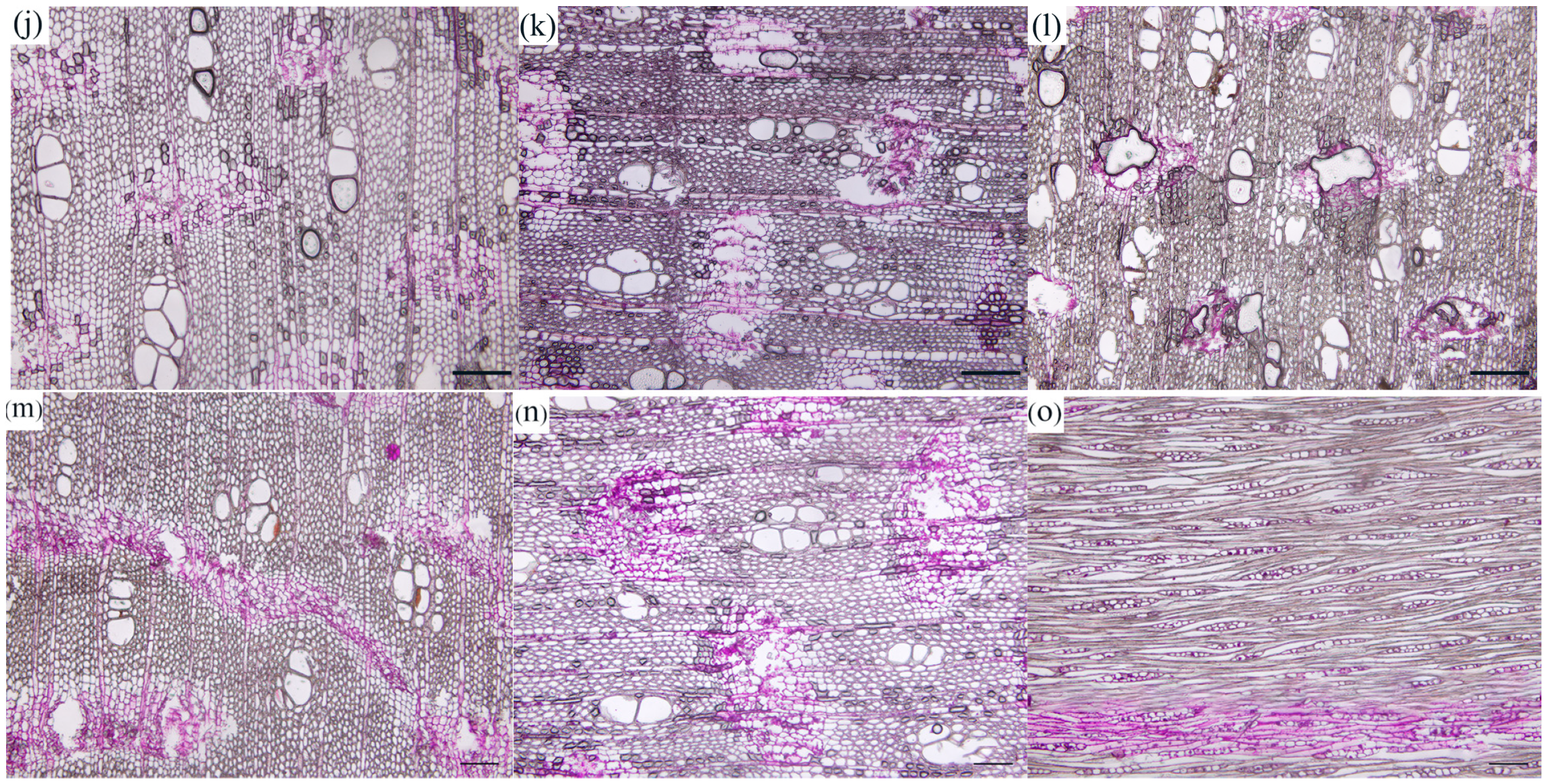
3.1.2. Changes in Starch Grains during the Formation of Agarwood
3.1.3. Changes in Starch Grains during the Formation of Agarwood
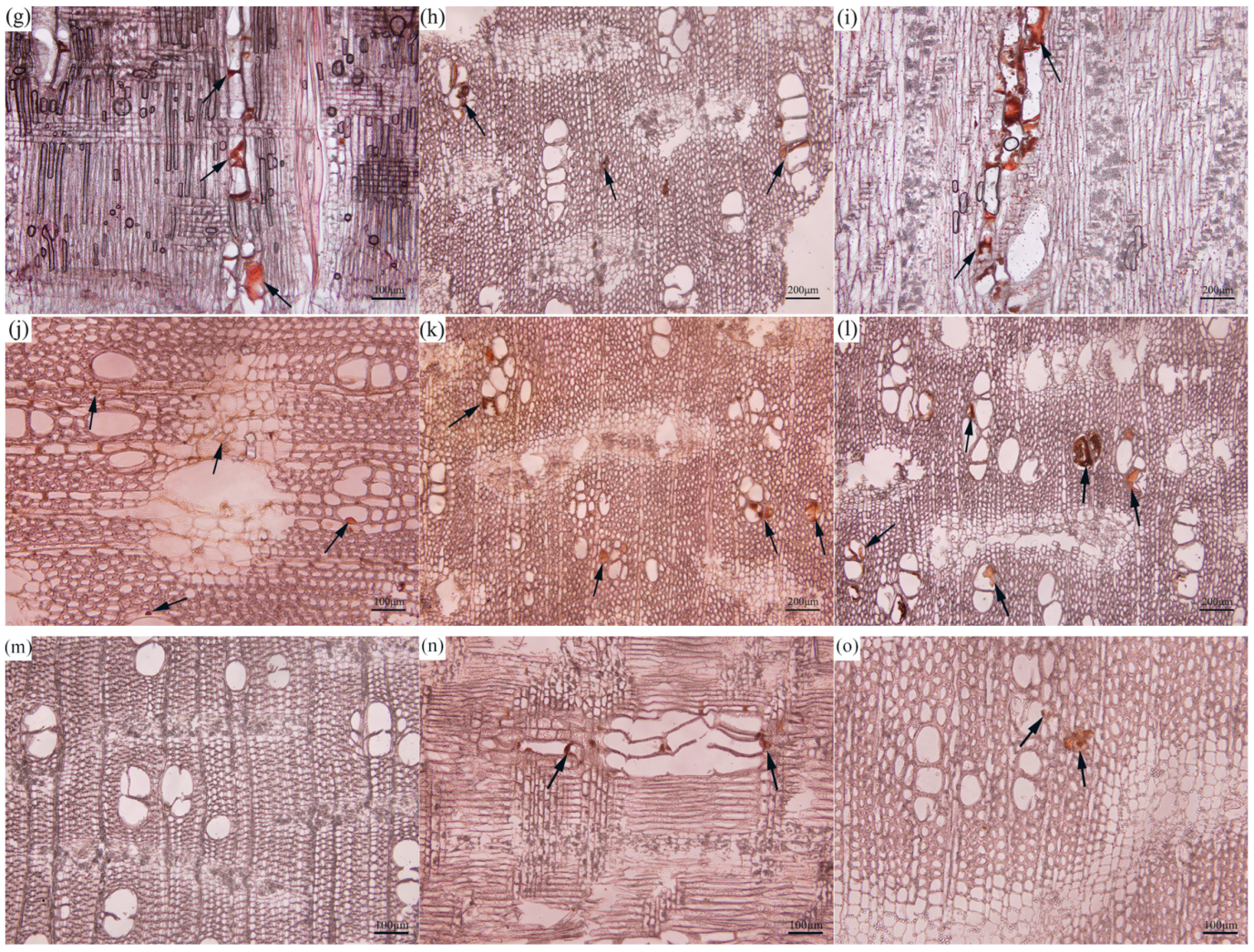
3.2. Structural Characteristics of the Inclusions in the Xylem of Agarwood
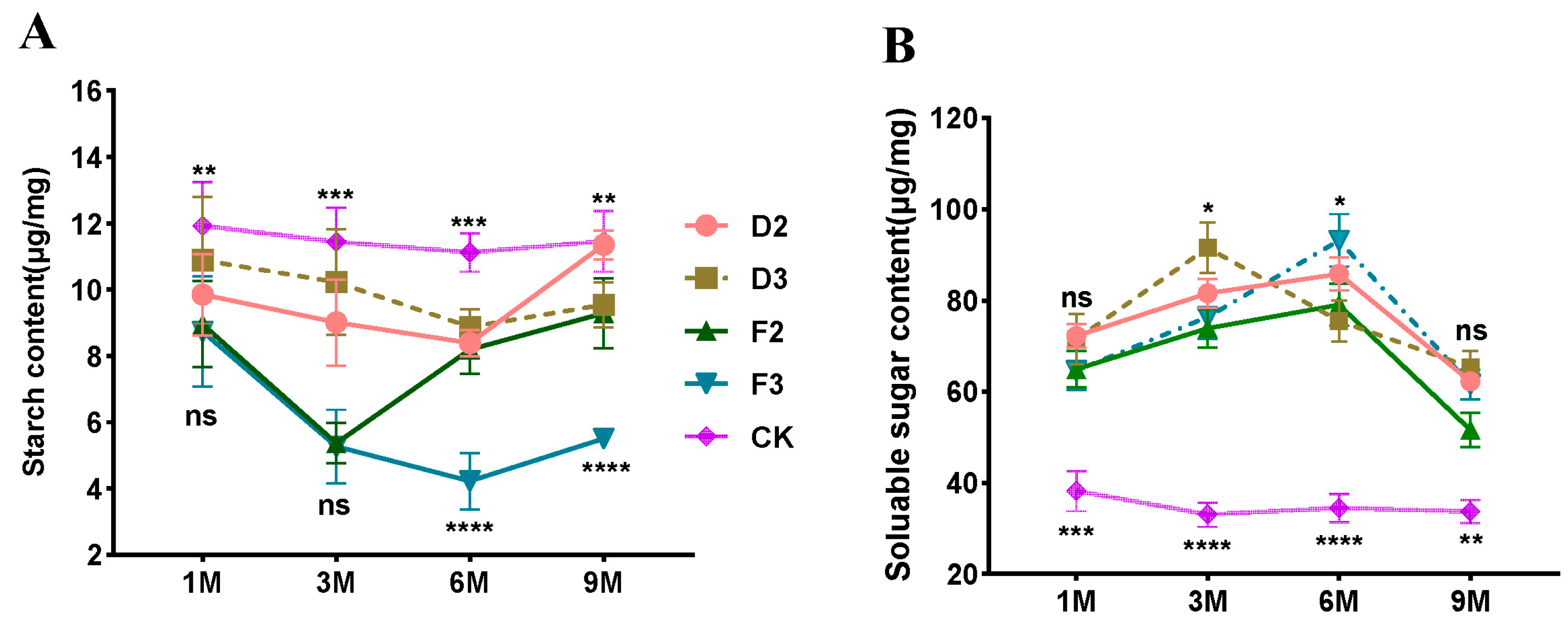
3.3. Non-Structural Carbohydrate Content
3.4. Determination of the Discoloration Range of Agarwood
3.5. Analysis of the Chemical Composition of Agarwood
3.5.1. Alcohol-Soluble Extract Content Analysis
3.5.2. Chemical Composition GC-MS Identification of Alcohol-Soluble Extracts
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound Name | Molecular Formula | Nine Months after Treatment | ||||
| D1 | D2 | F1 | F2 | CK | |||
| 1 | Benzaldehyde | C7H6O | 2.47 | 3.05 | 3.02 | 3.63 | 0.67 |
| 2 | Docosanoic acid | C22H44O2 | 1.01 | -- | 0.56 | 0.18 | -- |
| 3 | Dodecane | C12H26 | -- | -- | -- | -- | 1.14 |
| 4 | beta.-ylangene | C15H24 | 0.37 | -- | 1.24 | -- | -- |
| 5 | (-)-Globulol | C14H22O | -- | -- | -- | -- | -- |
| 6 | 2-Butanone,4-phenyl | C10H12O2 | 2.43 | -- | -- | 1.12 | -- |
| 7 | Valerenol | C15H24O | -- | 0.25 | -- | -- | -- |
| 8 | 1-Eicosene | C20H40 | -- | -- | -- | -- | 2.06 |
| 9 | Hexadecane | C16H34 | -- | -- | 0.21 | -- | 1.53 |
| 10 | Caryophyllene-(I1) | C15H24 | -- | 0.68 | -- | 0.31 | -- |
| 11 | L-.alpha.-Terpineol | C10H18O | 0.67 | 3.80 | -- | 2.14 | -- |
| 12 | 2,4-Di-tertbutylphenol | C14H22O | 5.24 | -- | -- | 0.86 | 1.26 |
| 13 | beta.-Phellandrene | C10H16 | 0.11 | -- | -- | -- | -- |
| 14 | Eicosane | C20H42 | 1.12 | 6.69 | 7.90 | 0.57 | 1.45 |
| 15 | Decane, 3,8-dimethyl- | C12H26 | -- | -- | -- | -- | 2.96 |
| 16 | Isoaromadendrene | C15H24O | -- | 2.89 | 2.42 | 4.64 | -- |
| 17 | alpha.-Guaiene | C15H24 | -- | -- | 0.97 | 1.12 | -- |
| 18 | n-Hexadecanoic acid | C16H32O2 | 3.57 | 10.07 | 9.85 | 2.80 | 30.33 |
| 19 | Dibutyl phthalate | C16H22O4 | 4.25 | 1.18 | 2.27 | -- | 4.38 |
| 20 | Ledene oxide-(II) | C15H24O | -- | 3.31 | 2.17 | -- | -- |
| 21 | Pentadecane | C15H32 | -- | -- | -- | -- | -- |
| 22 | Acetamide, N-(4-benzyloxyphenyl)-2-cyano- | C11H12N2O2 | -- | -- | 3.64 | -- | -- |
| 23 | beta.-Vatirenene | C15H22 | -- | -- | 2.52 | -- | -- |
| 24 | Oleic Acid | C18H34O2 | 3.15 | -- | -- | 2.31 | 1.57 |
| 25 | Saussurealactone | C15H22O2 | -- | -- | 0.76 | -- | -- |
| 26 | Hinesol | C15H26O | 2.47 | -- | -- | 0.50 | -- |
| 27 | beta-cyclocitral | C10H16O | -- | 0.74 | -- | 1.03 | -- |
| 28 | Aromadendrene oxide-(2) | C15H24O | -- | -- | 3.18 | -- | -- |
| 29 | ISOVELLERAL | C16H26O3 | -- | 7.89 | -- | -- | 2.79 |
| 30 | Andrographolide | C20H30O5 | -- | 6.68 | -- | -- | -- |
| 31 | 8-Naphthol, 1-(benzyloxy)- | C17H14O2 | -- | 1.61 | 9.58 | -- | -- |
| 32 | Longifolene | C15H24 | 1.68 | 3.57 | 7.38 | 12.22 | 0.29 |
| 33 | g-Gurjunene | C15H24O | -- | 3.16 | -- | -- | -- |
| 34 | beta.-Santalol | C15H24O | 0.48 | -- | 0.90 | 3.47 | -- |
| 35 | Valerena-4,7(11)-diene | C15H24 | -- | -- | -- | 0.56 | -- |
| 36 | Phenol, 2-methyl-5-(1-methylethyl)- | C10H14O | -- | -- | 0.98 | 0.57 | -- |
| 37 | Ethanone, 1-(1-cyclohexen-1-yl)- | C8H12O | 1.37 | -- | -- | -- | -- |
| 38 | Aromandendrene | C15H24 | -- | -- | -- | 1.68 | 5.69 |
| 39 | beta.-Vatirenene | C15H22 | -- | 0.94 | -- | -- | -- |
| 40 | Alloaromadendrene oxide-(2) | C15H240 | 0.39 | 1.89 | -- | 3.48 | -- |
| 41 | Bicyclo [7.2.0]undecane,10,10-dimethyl-2,6-bis(methylene)- | C15H24 | -- | -- | 1.36 | -- | -- |
| 42 | Valerenic acid | C15H22O2 | -- | 3.48 | -- | -- | -- |
| 43 | Hexacosane | C26H54 | 0.68 | -- | -- | 1.23 | -- |
| 44 | alpha.-Santalol | C15H24O | 2.77 | -- | 1.30 | -- | -- |
| 45 | alpha.-Farnesene | C15H24 | -- | -- | 0.71 | 2.36 | -- |
| 46 | 9-Octadecenamide, (Z)- | C18H35NO | 0.27 | -- | 1.27 | -- | 3.21 |
| 47 | Palustrol | C15H26O2 | -- | 0.87 | -- | -- | -- |
| 48 | Vellerdiol | C15H24O2 | 1.97 | -- | 4.57 | -- | -- |
| 49 | Valerenic acid | C15H22O2 | 3.38 | -- | -- | 9.65 | -- |
| 50 | Heneicosane | C21H44 | -- | -- | -- | -- | -- |
| 51 | Aromandendrene | C15H24 | -- | 6.47 | -- | 0.5 | -- |
| 52 | Humulene | C15H24O | -- | -- | -- | 0.55 | -- |
| 53 | gamma.-Sitosterol | C29H50O | -- | 0.59 | -- | -- | 2.04 |
| 54 | alpha-Bisabolene | C15H24 | -- | -- | -- | 0.79 | -- |
| 55 | Thymol | C10H14O | 0.89 | 0.40 | 0.57 | 2.00 | -- |
| 56 | Hexacosane- | C26H54 | -- | -- | 1.34 | -- | 9.72 |
| 57 | Stigmasterol | C29H48O | -- | -- | -- | -- | 3.42 |
| 58 | Octadecanoic acid | C18H36O2 | -- | -- | -- | -- | 1.12 |
| 59 | Rishitin | C14H22O2 | -- | -- | 0.74 | -- | -- |
| 60 | Coumarin, 6-benzyloxy-3,4-dihydro-4,4-dimethyl- | C18H18O3 | -- | 7.73 | 8.51 | -- | -- |
| 61 | Oleic Acid | C18H34O2 | 1.62 | -- | -- | 2.31 | -- |
| 62 | Alantolactone, 4.alpha.,4A. | C15H20O2 | -- | 0.81 | -- | -- | 1.38 |
| 63 | gama.-eudesmol | C10H18O | -- | 1.91 | 1.32 | 2.24 | -- |
| 64 | Tetracosane | C24H50 | 5.26 | -- | 0.24 | -- | -- |
| 65 | p-Pentylacetophenone | C13H18O | -- | -- | -- | 1.45 | 1.28 |
| 66 | 2-(2-Phenylethyl)-Chromone | C17H14O2 | 8.85 | 6.48 | 9.24 | 19.26 | -- |
| 67 | 6,7-Dimethoxy -2-(2-Phenylethyl)chromone | C19H18O4 | 9.72 | -- | 6.57 | -- | -- |
| 68 | 6-Methoxy-2-(2-Phenylethyl)chromone | C18H16O3 | 3.86 | 4.50 | -- | 18.27 | -- |
| 69 | 6,7-Methoxy-2-phenethyl-4H-chromen-4-one | C19H18O4 | 4.56 | -- | -- | 1.38 | |
| 70 | 6,7-Dimethoxy-2-(4-methoxyphenethyl)-4H-chromen-4-one | C20H20O5 | -- | 5.68 | 7.34 | -- | -- |
| 71 | Ethisterone | C21H28O2 | 1.53 | -- | 1.34 | 0.21 | -- |
| 72 | Anisyl butyrate | C17H14O2 | 0.35 | -- | -- | -- | -- |
| 73 | 1-benzylindole | C15H13N | -- | 1.28 | -- | 0.68 | -- |
| 74 | Tetrasiloxane, decamethyl- | C10H30O3Si4 | -- | -- | 0.38 | -- | -- |
| 75 | 7-hydroxy-2,5-dimethyl-4H-1-Benzopyran-4-0ne | C11H10O3 | 1.34 | -- | -- | 0.24 | 0.68 |
| 76 | Gingerenone A | C21H24O5 | -- | -- | 0.31 | -- | -- |
| 77 | gamma.-Sitosterol | C29H50O | 1.89 | -- | -- | 1.21 | -- |
| 78 | 2-p-Nitrophenyl-oxadiazol-1,3,4-one-5 | C8H5N3O4 | -- | -- | 1.34 | -- | -- |
| 79 | Tris(tert-butyldimethylsilyloxy)arsane | C18H45AsO3Si3 | 0.32 | -- | -- | -- | 1.34 |
References
- Elias, M.F.; Ibrahim, H.; Mahamod, W.R.W. A review on the malaysian Aquilaria species in karas plantation and agarwood production. Int. J. Acad. Res. Bus. Soc. Sci. 2017, 7, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Azren, P.D.; Lee, S.Y.; Emang, D.; Mohamed, R. History and perspectives of induction technology for agarwood production from cultivated Aquilaria in Asia: A review. J. For. Res. 2019, 30, 1–11. [Google Scholar] [CrossRef]
- Tan, C.S.; Isa, N.M.; Ismail, I.; Zainal, Z. Agarwood induction: Current developments and future perspectives. Front. Plant Sci. 2019, 10, 122. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, S.R.; Pokhrel, K.; Baral, S.D. Economic value of agarwood and its prospects of cultivation. Int. J. Appl. Sci. Biotechnol. 2021, 9, 23–31. [Google Scholar] [CrossRef]
- Ma, S.; Fu, Y.; Li, Y.; Wei, P.; Liu, Z. The formation and quality evaluation of agarwood induced by the fungi in Aquilaria sinensis. Ind. Crops Prod. 2021, 173, 114129. [Google Scholar] [CrossRef]
- Yin, Y.; Jiao, L.; Dong, M.; Jiang, X.; Zhang, S. Wood Resources, Identification, and Utilization of Agarwood in China. In Agarwood: Science behind the Fragrance; Mohamed, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 21–38. [Google Scholar]
- Shivanand, P.; Arbie, N.F.; Krishnamoorthy, S.; Ahmad, N. Agarwood—The Fragrant Molecules of a Wounded Tree. Molecules 2022, 27, 3386. [Google Scholar] [CrossRef] [PubMed]
- Rasool, S.; Mohamed, R. Understanding agarwood formation and its challenges. In Agarwood: Science behind the Fragrance; Mohamed, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 39–56. [Google Scholar]
- Naziz, P.S.; Das, R.; Sen, S. The scent of stress: Evidence from the unique fragrance of agarwood. Front. Plant Sci. 2019, 10, 840. [Google Scholar] [CrossRef]
- Pern, Y.C.; Lee, S.Y.; Ng, W.L.; Mohamed, R. Cross-amplification of microsatellite markers across agarwood-producing species of the Aquilarieae tribe (Thymelaeaceae). 3 Biotech. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Mitsilegas, V.; Fasoli, E.; Giuffrida, F.; Fitzmaurice, M. Environmental Crime at the International Level: Criminalisation of Illegal Trade in Wildlife under the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES). In The Legal Regulation of Environmental Crime; Brill Nijhoff: Leiden, The Netherlands, 2022; pp. 7–54. [Google Scholar]
- Thompson, I.D.; Lim, T.; Turjaman, M. Expensive, Exploited and Endangered. In A Review of the Agarwood-Producing Genera Aquilaria and Gyrinops: CITES considerations, Trade Patterns, Conservation, and Management; ITTO Technical Series No. 51; International Tropical Timber Organization (ITTO): Yokohama, Japan, 2022; pp. 14–25. [Google Scholar]
- Dobhal, S.; Bahuguna, D.; Binjola, R.; Bhatt, G.; Kumar, R.; Joshi, A.; Upadhyay, K.; Chauhan, N. Potential and scope of Agarwood (Aquilaria malaccensis lamk.) cultivation in India. Pantnagar. J. Res. 2023, 21, 2. [Google Scholar]
- Adam, A.Z.; Lee, S.Y.; Mohamed, R. Pharmacological properties of agarwood tea derived from Aquilaria (Thymelaeaceae) leaves: An emerging contemporary herbal drink. J. Herb. Med. 2017, 10, 37–44. [Google Scholar] [CrossRef]
- El Enshasy, H.A.; Hanapi, S.Z.; Malek, R.A.; Abdelgalil, S.A.; Leng, O.M. Endophytic fungi: The desired biostimulants for essential oil production. In Endophytic Fungal Research. Fungal Biology; Singh, B., Ed.; Springer: Cham, Switzerland, 2019; pp. 211–232. [Google Scholar]
- Kaleri, A.H.; Song, X.-Q.; Dai, H.F.; Kaleri, A.A.; Muhammad, N.M.; Mehmood, A.; Kaleri, G.S.; Kaleri, A.R. 1. An important variety of organic gemstones-Agarwood. Patent Abstr. Bibliogr. 2020, 9, 1424–1435. [Google Scholar] [CrossRef]
- Hoque, M.; Mondal, M.; Sagor, G.; Hasan, M.; Hannan, A. Inclusive Scenario of Natural Products Obtained from Agar Plant (Aquilaria sp.) In Bangladesh-A Review. J. Biosci. 2021, 29, 151–173. [Google Scholar] [CrossRef]
- Ding, X.; Mei, W.; Lin, Q.; Wang, H.; Wang, J.; Peng, S.; Li, H.; Zhu, J.; Li, W.; Wang, P. Genome sequence of the agarwood tree Aquilaria sinensis (Lour.) Spreng: The first chromosome-level draft genome in the Thymelaeceae family. GigaScience 2020, 9, giaa013. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, H.; Yang, Y.; Zhang, Z.; Wei, J.; Meng, H.; Chen, W.; Feng, J.; Gan, B.; Chen, X. Whole-tree agarwood-inducing technique: An efficient novel technique for producing high-quality agarwood in cultivated Aquilaria sinensis trees. Molecules 2013, 18, 3086–3106. [Google Scholar] [CrossRef]
- Ye, W.; Zhang, W.; Liu, T.; Zhu, M.; Li, S.; Li, H.; Huang, Z.; Gao, X. iTRAQ-based quantitative proteomic analysis of chemically induced Aquilaria sinensis provides insights into agarwood formation mechanism. Proteomics 2018, 18, 1800023. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qiao, M.; Fu, Y.; Wei, P.; Li, Y.; Liu, Z. Tissue structure changes of Aquilaria sinensis xylem after fungus induction. Forests 2022, 13, 43. [Google Scholar] [CrossRef]
- Van Thanh, L.; Van Do, T.; Son, N.H.; Sato, T.; Kozan, O. Impacts of biological, chemical and mechanical treatments on sesquiterpene content in stems of planted Aquilaria crassna trees. Agrofor. Syst. 2015, 89, 973–981. [Google Scholar] [CrossRef]
- Chhipa, H.; Chowdhary, K.; Kaushik, N. Artificial production of agarwood oil in Aquilaria sp. by fungi: A review. Phytochem. Rev. 2017, 16, 835–860. [Google Scholar] [CrossRef]
- Yang, Y.; Feng, J.; Zhu, J.; Liu, Y.; Chen, D.; Wei, J. Quality of agarwood produced by whole tree agarwood-inducing technique (Agar-WIT) from China and Southeast Asian countries. Chin. Pharm. J. 2019, 54, 1988–1994. [Google Scholar]
- Chen, X.; Liu, Y.; Yang, Y.; Feng, J.; Liu, P.; Sui, C.; Wei, J. Trunk surface agarwood-inducing technique with Rigidoporus vinctus: An efficient novel method for agarwood production. PLoS ONE 2018, 13, e0198111. [Google Scholar] [CrossRef]
- Liu, Y. Research on Quality Characteristics of Agarwood Produced by Whole-Tree Agarwood-Inducing Technique. Ph.D. Thesis, Peking Union Medical College, Beijing, China, 2018. [Google Scholar]
- Chen, X.; Sui, C.; Liu, Y.; Yang, Y.; Liu, P.; Zhang, Z.; Wei, J. Agarwood formation induced by fermentation liquid of Lasiodiplodia theobromae, the dominating fungus in wounded wood of Aquilaria sinensis. Curr. Microbiol. 2017, 74, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Subasinghe, S.; Hitihamu, H.; Fernando, K. Use of two fungal species to induce agarwood resin formation in Gyrinops walla. J. For. Res. 2019, 30, 721–726. [Google Scholar] [CrossRef]
- Faizal, A.; Azar, A.W.P.; Turjaman, M.; Esyanti, R.R. Fusarium solani induces the formation of agarwood in Gyrinops versteegii (Gilg.) Domke branches. Symbiosis 2020, 81, 15–23. [Google Scholar] [CrossRef]
- Mohamed, R.; Jong, P.L.; Kamziah, A.K. Fungal inoculation induces agarwood in young Aquilaria malaccensis trees in the nursery. J. For. Res. 2014, 25, 201–204. [Google Scholar] [CrossRef]
- Zhang, Z.; Han, X.M.; Wei, J.H.; Xue, J.; Yang, Y.; Liang, L.; Li, X.J.; Guo, Q.M.; Xu, Y.H.; Gao, Z.H. Compositions and antifungal activities of essential oils from agarwood of Aquilaria sinensis (Lour.) Gilg induced by Lasiodiplodia theobromae (Pat.) Griffon. & Maubl. J. Braz. Chem. Soc. 2014, 25, 20–26. [Google Scholar]
- Tibpromma, S.; Zhang, L.; Karunarathna, S.C.; Du, T.Y.; Phukhamsakda, C.; Rachakunta, M.; Suwannarach, N.; Xu, J.; Mortimer, P.E.; Wang, Y.H. Volatile constituents of endophytic fungi isolated from Aquilaria sinensis with descriptions of two new species of Nemania. Life 2021, 11, 363. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, J. Endophytic fungi of agarwood and their chemical compounds. Fungal Biotec 2022, 2, 16–35. [Google Scholar]
- Zhang, Z.; Zhao, M.; Ran, J.; Gao, M.; Li, N.X.; Ma, Y.M.; Sun, Y.; Li, Y. Fusarium oxysporum infection-induced formation of agarwood (FOIFA): A rapid and efficient method for inducing the production of high quality agarwood. PLoS ONE 2022, 17, e0277136. [Google Scholar] [CrossRef]
- Khalil, A.M.A.; Hassan, S.E.-D.; Alsharif, S.M.; Eid, A.M.; Ewais, E.E.-D.; Azab, E.; Gobouri, A.A.; Elkelish, A.; Fouda, A. Isolation and characterization of fungal endophytes isolated from medicinal plant Ephedra pachyclada as plant growth-promoting. Biomolecules 2021, 11, 140. [Google Scholar] [CrossRef] [PubMed]
- Woudenberg, J.; Groenewald, J.; Binder, M.; Crous, P. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef]
- Kidd, S.; Halliday, C.; Ellis, D. Aspergillus Micheli Ex Link. In Descriptions of Medical Fungi; Kidd, S., Halliday, C., Ellis, D., Eds.; Springer: Adelaide, Australia, 2022; pp. 20–34. [Google Scholar]
- Jedidi, I.; Soldevilla, C.; Lahouar, A.; Marín, P.; González-Jaén, M.T.; Said, S. Mycoflora isolation and molecular characterization of Aspergillus and Fusarium species in Tunisian cereals. Saudi J. Biol. Sci. 2018, 25, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Song, X.; Song, J.; Huang, G.; Wang, X.; Liang, K.; Yang, G.; Zhang, Q. A Biophysical Method for Inducing Incense Formation in Earth Aquilaria Trees and Its Application. C.N. Patent CN201910608768, 31 August 2021. [Google Scholar]
- Zhang, P.; Li, X.; Cui, Z.; Xu, D. Morphological, physiological, biochemical and molecular analyses reveal wounding-induced agarwood formation mechanism in two types of Aquilaria sinensis (Lour.) Spreng. Ind. Crops Prod. 2022, 178, 114603. [Google Scholar] [CrossRef]
- Castillo-Argaez, R.; Vazquez, A.; Konkol, J.L.; Vargas, A.I.; Ploetz, R.C.; Etxeberria, E.; Schaffer, B. Sap flow, xylem anatomy and photosynthetic variables of three Persea species in response to laurel wilt. Tree Physiol. 2021, 41, 1004–1018. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Li, X.; Xu, D.; Yang, Z.; Zhang, N.; Liu, X.; Hong, Z. Physiological changes during heartwood formation induced by plant growth regulators in Dalbergia odorifera (Leguminosae). IAWA J. 2021, 42, 217–234. [Google Scholar] [CrossRef]
- Lin, L.; Guo, D.; Huang, J.; Zhang, X.; Zhang, L.; Wei, C. Molecular structure and enzymatic hydrolysis properties of starches from high-amylose maize inbred lines and their hybrids. Food Hydrocolloid. 2016, 58, 246–254. [Google Scholar] [CrossRef]
- LY/T 2904-2017; Standardization Administration of the People’s Republic of China. Chinese Standard of Forestry, Agarwood; Standards press of China: Beijing, China, 2017.
- Mahajan, M.; Kuiry, R.; Pal, P.K. Understanding the consequence of environmental stress for accumulation of secondary metabolites in medicinal and aromatic plants. J. Appl. Res. Med. Aromat. Plants 2020, 18, 100255. [Google Scholar] [CrossRef]
- Thakur, M.; Bhattacharya, S.; Khosla, P.; Puri, S.K. Improving production of plant secondary metabolites through biotic and abiotic elicitation. J. Appl. Res. Med. Aromat. Plants 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Zhan, X.; Chen, Z.; Chen, R.; Shen, C. Environmental and Genetic Factors Involved in Plant Protection-Associated Secondary Metabolite Biosynthesis Pathways. Front. Plant Sci. 2022, 13, 877304. [Google Scholar] [CrossRef]
- Ahmed, E.; Arshad, M.N.; Khan, M.Z.; Amjad, M.; Sadaf, H.M.; Riaz, I.; Sabir, S.; Ahmad, N.; Sabaoon. Secondary metabolites and their multidimensional prospective in plant life. J. Pharmacogn. Phytochem. 2017, 6, 205–214. [Google Scholar]
- Ashraf, M.A.; Iqbal, M.; Rasheed, R.; Hussain, I.; Riaz, M.; Arif, M.S. Environmental Stress and Secondary Metabolites in Plants: An Overview. In Plant Metabolites and Regulation under Environmental Stress; Parvaiz, A., Ahanger, M.A., Singh, V.P., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 153–167. [Google Scholar]
- Verma, N.; Shukla, S.K. Impact of various factors responsible for fluctuation in plant secondary metabolites. J. Appl. Res. Med. Aromat. Plants 2015, 2, 105–113. [Google Scholar] [CrossRef]
- Araya-Cloutier, C.; den Besten, H.M.W.; Aisyah, S.; Gruppen, H.; Vincken, J.P. The position of prenylation of isoflavonoids and stilbenoids from legumes (Fabaceae) modulates the antimicrobial activity against Gram positive pathogens. Food Chem. 2017, 226, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Moola, A.K.; Diana, R.K.B. Elicitation as a means for enhanced plant secondary metabolites through hairy root system. Adv. Sci. Res. 2019, 10, 6–12. [Google Scholar]
- Anderson, C.T.; Kieber, J.J. Dynamic construction, perception, and remodeling of plant cell walls. Annu. Rev. Plant Biol. 2020, 71, 39–69. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; He, M.; Hou, Q.; Zou, L.; Yang, Y.; Wei, Y.; Chen, X. Cell wall associated immunity in plants. Stress Biol. 2021, 1, 3. [Google Scholar] [CrossRef] [PubMed]
- Ninkuu, V.; Yan, J.; Fu, Z.; Yang, T.; Ziemah, J.; Ullrich, M.S.; Kuhnert, N.; Zeng, H. Lignin and Its Pathway-Associated Phytoalexins Modulate Plant Defense against Fungi. J. Fungi 2023, 9, 52. [Google Scholar] [CrossRef] [PubMed]
- Perincherry, L.; Urbaniak, M.; Pawłowicz, I.; Kotowska, K.; Waśkiewicz, A.; Stępień, Ł. Dynamics of Fusarium Mycotoxins and Lytic Enzymes during Pea Plants’ Infection. Int. J. Mol. Sci. 2021, 22, 9888. [Google Scholar] [CrossRef]
- Chaudhary, P.; Agri, U.; Chaudhary, A.; Kumar, A.; Kumar, G. Endophytes and their potential in biotic stress management and crop production. Front. Microbiol. 2022, 13, 933017. [Google Scholar] [CrossRef] [PubMed]
- Miliute, I.; Buzaite, O.; Baniulis, D.; Stanys, V. Bacterial endophytes in agricultural crops and their role in stress tolerance: A review. Zemdirbyste 2015, 102, 465–478. [Google Scholar] [CrossRef]
- Ngadiran, S.; Baba, S.; Nor, N.F.A.; Yahayu, M.; Muhamad, M.F.; Kushairi, A.K.A.; Zaidel, D.N.A.; Muhamad, I.I. The induction techniques of resinous agarwood formation: A review. Bioresour. Technol. Rep. 2023, 21, 101337. [Google Scholar] [CrossRef]
- Du, T.Y.; Dao, C.J.; Mapook, A.; Stephenson, S.L.; Elgorban, A.M.; Al-Rejaie, S.; Suwannarach, N.; Karunarathna, S.C.; Tibpromma, S. Diversity and biosynthetic activities of agarwood associated fungi. Diversity 2022, 14, 211. [Google Scholar] [CrossRef]
- Liu, T.; Liu, Y.; Fu, Y.; Qiao, M.; Wei, P.; Liu, Z.; Li, Y. Structural, defense enzyme activity and chemical composition changes in the xylem of Aquilaria sinensis during fungus induction. Ind. Crops Prod. 2024, 208, 117804. [Google Scholar] [CrossRef]
- Cui, Z.; Li, X.; Xu, D.; Yang, Z. Changes in non-structural carbohydrates, wood properties and essential oil during chemically-induced heartwood formation in Dalbergia odorifera. Front. Plant Sci. 2020, 11, 1161. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Zhang, X.; Yang, Y.; Sui, C.; Xu, Y.; Wei, J. Interxylary phloem and xylem rays are the structural foundation of agarwood resin formation in the stems of Aquilaria sinensis. Trees 2019, 33, 533–542. [Google Scholar] [CrossRef]
- Hartmann, H.; Trumbore, S. Understanding the roles of nonstructural carbohydrates in forest trees–from what we can measure to what we want to know. New Phytol. 2016, 211, 386–403. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, S.N.; Sahney, M.; Tripathi, A.; Pandey, P.; Jatav, H.S.; Minkina, T.; Rajput, V.D. Elucidating the anatomical features, adaptive and ecological significance of Kopsia fruticosa Roxb.(Apocynaceae). Horticulturae 2023, 9, 387. [Google Scholar] [CrossRef]
- Wang, Y.; Hussain, M.; Jiang, Z.; Wang, Z.; Gao, J.; Ye, F.; Mao, R.; Li, H. Aquilaria species (Thymelaeaceae) distribution, volatile and non-volatile phytochemicals, pharmacological uses, agarwood grading system, and induction methods. Molecules 2021, 26, 7708. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Wei, J.H.; Gao, Z.H.; Zhang, Z.; Lyu, J.C. A review of quality assessment and grading for agarwood. Chin. Herb. Med. 2017, 9, 22–30. [Google Scholar] [CrossRef]
- Mohamed, R.; Lee, S.Y. Keeping up appearances: Agarwood grades and quality. In Agarwood: Science behind the Fragrance; Mohamed, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 149–167. [Google Scholar]
- Wangiyana, I.G.A.S. Medicinal Usage of Agarwood Resin in Form of Essential Oil: A Review. J. Silva Samalas 2019, 2, 86–90. [Google Scholar]
- Justin, S.; Lihan, S.; Elvis-Sulang, M.; Chiew, T. Formulated microbial consortium as inoculant for agarwood induction. J. Trop. For. Sci. 2020, 32, 161–169. [Google Scholar] [CrossRef]
- Turjaman, M.; Hidayat, A.; Santoso, E. Development of agarwood induction technology using endophytic fungi. In Agarwood: Science Behind the Fragrance; Mohamed, R., Ed.; Springer: Singapore, 2016; pp. 57–71. [Google Scholar]
- Ramli, A.N.M.; Yusof, S.; Bhuyar, P.; Aminan, A.W.; Tajuddin, S.N. Fungi mediated agarwood (A. malaccensis) production and their pharmaceutical applications: A systematic review. Int. J. Plant Based Pharm. 2022, 2, 261–270. [Google Scholar]
- Faizal, A.; Hermawaty, D.; Junita, E.; Rahmawati, A.; Azar, A.W.P.; Makajanma, M.M.; Turjaman, M. Evaluation of biotic and abiotic stressors to artificially induce agarwood production in Gyrinops versteegii (Gilg.) Domke seedlings. Symbiosis 2022, 86, 229–239. [Google Scholar] [CrossRef]
- Ismail, N.; Rahiman, M.H.F.; Taib, M.N.; Ibrahim, M.; Zareen, S.; Tajuddin, S.N. A review on agarwood and its quality determination. In Proceedings of the 2015 IEEE 6th Control and System Graduate Research Colloquium (ICSGRC), Shah Alam, Malaysia, 10–11 August 2015; pp. 103–108. [Google Scholar]
- Chen, S.T.; Rao, Y.K. An overview of agarwood, phytochemical constituents, pharmacological activities, and analyses. Tradit. Med. 2022, 3, 1–71. [Google Scholar] [CrossRef]
- Li, W.; Chen, H.Q.; Wang, H.; Mei, W.L.; Dai, H.F. Natural products in agarwood and Aquilaria plants: Chemistry, biological activities and biosynthesis. Nat. Prod. Rep. 2021, 38, 528–565. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; He, Q.Q.; Chen, X.Q.; Feng, J.; Wie, J.H.; Liu, Y.Y. Chemical and Bioactivity Diversity of 2-(2-Phenylethyl) chromones in Agarwood: A Review. Chem. Biodivers. 2022, 19, e202200490. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yu, Z.; Wang, C.; Wu, C.; Guo, P.; Wei, J. Chemical constituents and pharmacological activity of agarwood and Aquilaria plants. Molecules 2018, 23, 342. [Google Scholar] [CrossRef]
- Kalra, R.; Kaushik, N. A review of chemistry, quality and analysis of infected agarwood tree (Aquilaria sp.). Phytochem. Rev. 2017, 16, 1045–1079. [Google Scholar] [CrossRef]










| Sample Code | Treatment Method | Collection Time/Month | Number of Trees |
|---|---|---|---|
| D1 | drilling and agar-fungal A | 1, 3, 6, 9 | N = 3 |
| D2 | drilling and agar-fungal B | 1, 3, 6, 9 | N = 3 |
| F1 | burning and agar-fungal A | 1, 3, 6, 9 | N = 3 |
| F2 | burning and agar-fungal B | 1, 3, 6, 9 | N = 3 |
| CK | drilling | 1, 3, 6, 9 | N = 3 |
| Treatment | Discoloration Range | |||||
|---|---|---|---|---|---|---|
| Discoloration Length between Holes (cm) | Discoloration Width (cm) | |||||
| Third Months | Sixth Months | Ninth Months | Third Months | Sixth Months | Ninth Months | |
| D1 | 1.07 ± 0.21 c | 2.30 ± 0.10 c | 3.30 ± 5.2 c | 1.13 ± 2.3 b | 2.13 ± 0.06 c | 2.17 ± 0.06 c |
| D2 | 1.43 ± 0.31 c | 2.27 ± 0.15 c | 3.33 ± 6.0 c | 2.03 ± 6.5 a | 2.03 ± 0.06 c | 2.07 ± 0.06 c |
| F1 | 2.60 ± 0.26 b | 3.50 ± 0.26 b | 4.60 ± 6.3 b | 1.43 ± 1.3 b | 3.13 ± 0.15 b | 3.37 ± 0.12 b |
| F2 | 4.53 ± 0.29 a | 5.70 ± 0.20 a | 10.70 ± 0.46 a | 2.07 ± 0.06 a | 4.43 ± 0.21 a | 5.43 ± 0.61 a |
| CK | 0.27 ± 0.06 e | 0.53 ± 0.06 d | 0.63 ± 0.12 d | 0.17 ± 0.06 c | 0.47 ± 0.06 d | 0.47 ± 0.12 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, W.; Song, X.; Zhou, Z.; Liu, G.; Zhang, Q.; Pang, S. Effects of Different Levels of Physical Damage Combined with Fungal Induction on Agarwood Formation. Forests 2024, 15, 168. https://doi.org/10.3390/f15010168
Zhao W, Song X, Zhou Z, Liu G, Zhang Q, Pang S. Effects of Different Levels of Physical Damage Combined with Fungal Induction on Agarwood Formation. Forests. 2024; 15(1):168. https://doi.org/10.3390/f15010168
Chicago/Turabian StyleZhao, Weiwei, Xiaochen Song, Zaizhi Zhou, Gaofeng Liu, Qingqing Zhang, and Shengjiang Pang. 2024. "Effects of Different Levels of Physical Damage Combined with Fungal Induction on Agarwood Formation" Forests 15, no. 1: 168. https://doi.org/10.3390/f15010168
APA StyleZhao, W., Song, X., Zhou, Z., Liu, G., Zhang, Q., & Pang, S. (2024). Effects of Different Levels of Physical Damage Combined with Fungal Induction on Agarwood Formation. Forests, 15(1), 168. https://doi.org/10.3390/f15010168