

Conserving Potential and Endangered Species of Pericopsis mooniana Thwaites in Indonesia

 , , ,
, , , 
 , , , ,
, , , ,  , , ,
, , , 
Abstract
1. Introduction
2. Methods
3. Results
3.1. Conservation
3.1.1. Botanical Description of Kayu Kuku (P. mooniana)
- Kingdom: Plantae
- Division: Spermatophytae
- Sub division: Angiospermae
- Class: Magnoliopsida
- Order: Fabales
- Family: Fabaceae
- Genus: Pericopsis
- Species: Pericopsis mooniana Thwaites.
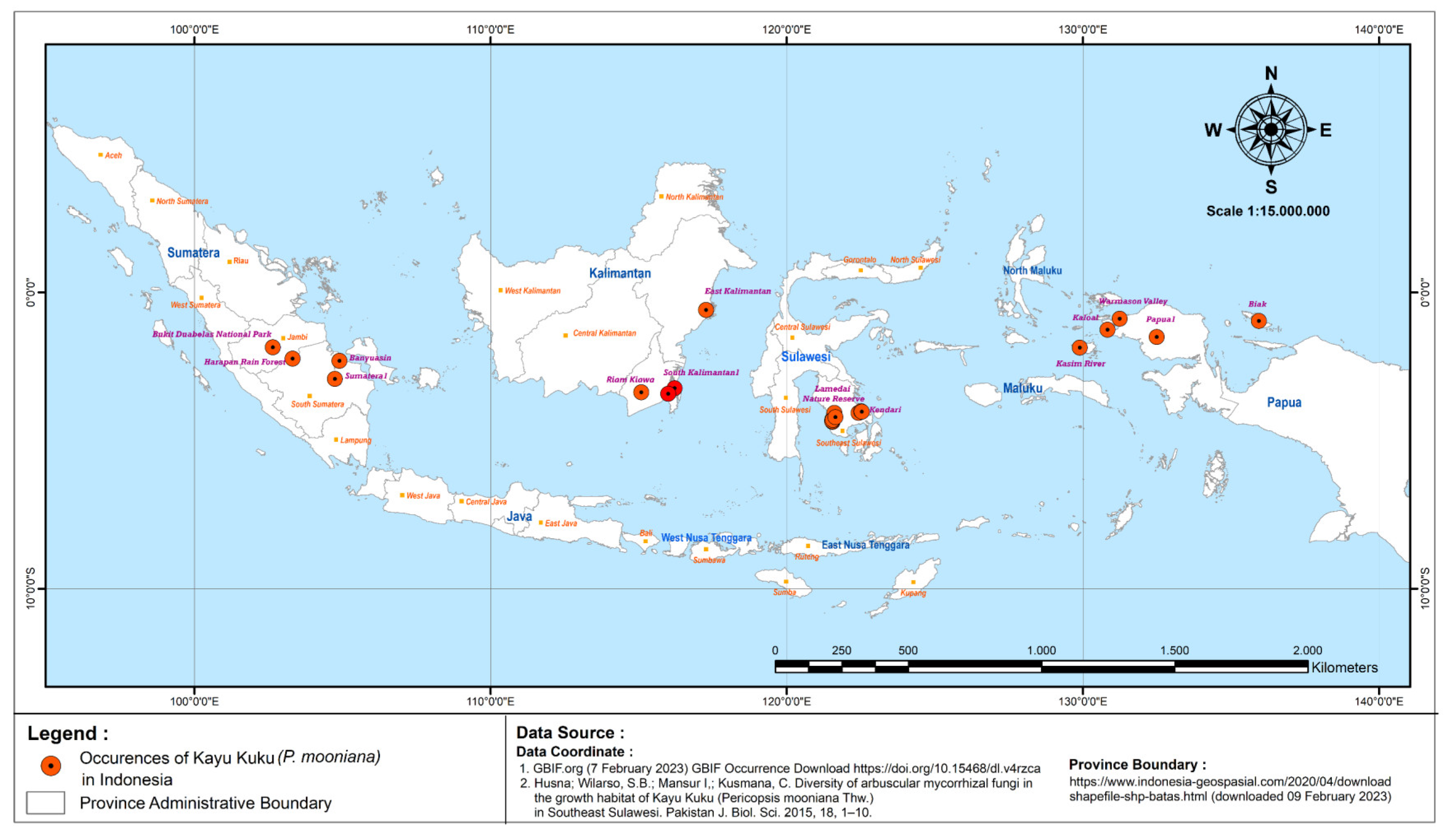
3.1.2. Habitat Characteristics and Distribution
3.1.3. Genetic Resources of P. mooniana
3.2. Cultivation of P. mooniana
3.2.1. Seed Technology
3.2.2. Propagation Techniques
Generative
Vegetative Propagation
3.2.3. Seedling Improvement Technique
3.2.4. Pest and Disease of P. mooniana
3.2.5. Mycorrhizal Fungi Application
3.2.6. Silviculture Techniques
- In the nickel post-mining area, planting holes were 60 × 60 × 60 cm. Plants were planted with a distance between planting holes of 4 m x 4 m. Before planting, the soil is mixed with compost and urea (5 kg and 100 g, respectively), one-fifth of the standard carried out by PT. Vale Indonesia Tbk and mixing sago pulp with soil media was carried out, with a height and diameter increment of 0.44 m and 0.8 cm [2,96]. P. mooniana was planted between reclaimed trees at a spacing of 4 × 5 m and filled with 25 kg of active compost and 0.4 kg of urea + TSP + KCl + dolomite each per planting hole. In the first and second 6 months of maintenance, 0.4 kg each of urea + TSP + KCl was added, and in the third period, pruning was conducted and mulch was placed at the base of the stem circle. This treatment increases height and diameter increment of 0.93 m and 1.06 cm, respectively (personal communication with Mr. Guntur Sambernyowo, Retired PT Vale Indonesia).
- In gold post-mining land, the spacing is 2 × 2 m with a planting hole of 40 × 40 × 40 cm. P. mooniana seedlings with a height of ±30 cm were transferred and planted in the holes with 4 kg of compost. A total of 50 g of NPK fertilizer per plant was added after one day of planting and then treated with watering and weed control. It increased height and diameter averages of 0.2 m and 0.36 cm, respectively [83].
- In the land occupied by Imperata cylindrica, the planting distance was 2.5 × 2.5 m with a planting hole of 30 × 30 × 30 cm and basic fertilizer of 1 kg of compost. The mean increase in height and diameter is 0.89 m and 1.20 cm at 17 years old (observation on site).
- In the green open space of Halu Oleo University, the planting spacing was 3 × 3 m with 30 × 30 × 30 cm planting holes and no further fertilization. The mean increase in height and diameter at 13 years old is 0.84 m and 1.75 cm (observation on site).
3.2.7. Future Opportunities in Improving Seed and Seedling Quality
3.2.8. Harvesting Technique
3.3. Importance of P. mooniana for the Wood Industry
3.3.1. Wood Properties
3.3.2. Wood Anatomy Structure
3.3.3. Wood Uses
3.4. Economics and Regulation
3.4.1. Trading and Its Regulations
3.4.2. Sustainable Wood Supply
3.5. Discussion and Conservation Strategies for P. mooniana
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Soerianegara, I.; Lemmens, R. Sumber Daya Nabati Asia Tenggara: Pohon Penghasil Kayu Perdagangan Yang Utama, 1st ed.; Prosea-Balai Pustaka: Jakarta, Indonesia, 2002; Volume 5. [Google Scholar]
- Husna. Budi Daya Dan Konservasi Kayu Kuku, 1st ed.; Tuheteru, F.D., Elviandri, Y., Eds.; IPB Press: Bogor, Indonesia, 2015. [Google Scholar]
- Alfaizin, D. Potensi Kayu Kuku (Pericopsis mooniana THW) untuk revegetasi lahan kritis. In Prosiding Seminar Biologi from Basic Science to Comprehensive Education; Universitas Islam Negeri Alauddin: Makassar, Indonesia, 2016; pp. 219–225. [Google Scholar]
- Suhartati; Nursyamsi; Alfaizin, D. Mengenal morfologi, tipe buah dan biji pada pohon Kayu Kuku (Pericopsis mooniana THW). Info Tek. Eboni 2015, 12, 87–96. [Google Scholar]
- Affre, A.; Kathe, W.; Raymakers, C. Looking Under the Veneer. In Implementation Manual on EU Timber Trade Control: Focus on CITES-Listed Trees; TRAFFIC Europe: Brussels, Belgium, 2004. [Google Scholar]
- Akbar, A.; Rusmana. Membangkitkan Primadona yang Mulai Langka: Kayu Kuku (Pericopsis mooniana Thw). Bekantan 2013, 1, 4–6. [Google Scholar]
- Hills, R. Pericopsis elata. Available online: https://www.gbif.org/species/176802781 (accessed on 16 August 2022).
- Asian Regional Workshop. Pericopsis mooniana. Available online: https://www.gbif.org/species/176802779 (accessed on 20 August 2022).
- Yuskianti, V.; Salihin. Sebaran Kayu Kuku (Pericopsis mooniana) di tegakan benih teridentifikasi (TBT) L. Gani di Anaiwoi, Sulawesi Tenggara. Wana Benih 2016, 17, 9–15. [Google Scholar]
- Botanic Gardens Conservation International. Pericopsis laxiflora. Available online: http://www.worldfloraonline.org/taxon/wfo-0000212085 (accessed on 21 August 2022).
- Micheneau, C.; Dauby, G.; Bourland, N.; Doucet, J.L.; Hardy, O.J. Development and Characterization of Microsatellite Loci in Pericopsis elata (Fabaceae) Using A Cost-efficient Approach. J. Bot 2011, 98, 268–270. [Google Scholar] [CrossRef]
- Nkulu, S.N.; Meerts, P.; Wa Ilunga, E.I.; Shutcha, M.N.; Bauman, D. Medicinal Vitex species (Lamiaceae) occupy different niches in Haut-Katanga Tropical Dry Woodlands. Plant Ecol. Evol. 2022, 155, 236–247. [Google Scholar] [CrossRef]
- Gunatilleke, N.; Pethiyagoda, R.; Gunatilleke, S. Biodiversity of Sri Lanka. J. Natl. Sci. Found. Sri Lanka 2008, 36, 25–61. [Google Scholar] [CrossRef]
- Ministry of Environment and Forestry(MoEF). The State of Indonesia’s Forests, 1st ed.; Efransjah, S., Murniningtyas, Erwinsyah, Damayanti, E., Eds.; Ministry of Environment and Forestry: Jakarta, Indonesia, 2022. [Google Scholar]
- Setyowati, R.D.N.; Amala, N.A.; Aini, N.N.U. Studi pemilihan tanaman revegetasi untuk keberhasilan reklamasi lahan bekas tambang. Al-Ard J. Tek. Lingkung 2017, 3, 14–20. [Google Scholar] [CrossRef]
- Saputra, R.A.; Riniarti, M. Pertumbuhan dan mutu fisik bibit Kayu Kuku (Pericopsis mooniana) pada media tanam tailing pertambangan emas rakyat dengan pemberian amelioran. ULIN J. Hutan Trop. 2020, 4, 58–69. [Google Scholar] [CrossRef]
- Suhartati; Alfaizin, D. Teknik pembibitan spesies Kayu Kuku (Pericopsis mooniana) untuk reklamasi lahan bekas tambang tanah liat. J. Faloak 2018, 2, 103–114. [Google Scholar] [CrossRef]
- Perala, I.; Wulandari, A.S. Kayu Kuku (Pericopsis mooniana Thw.) seedlings growth response to tailing media added with vermicompost, rhizobium, and arbuscular mycorrhizalf. IOP Conf. Ser. Earth Environ. Sci. 2019, 394. [Google Scholar] [CrossRef]
- Alfaizin, D.; Suhartati; Kurniawan, E. Benih dan perkecambahan kayu kuku( Pericopsis mooniana THW). Info Tek. Eboni 2016, 13, 1–11. [Google Scholar]
- Suhartati; Alfaizin, D.; Kurniawan, E. Budidaya tanaman Kayu Kuku (Pericopsis mooniana Thw). In Budidaya dan pemanfaatan spesies pohon kayu lokal Sulawesi; Nas Media Pustaka: Makassar, Indonesia, 2020; pp. 61–97. [Google Scholar]
- Heyne, K. Tumbuhan Berguna Indonesia; Yayasan Sarana Wana Jaya: Jakarta, Indonesia, 1987. [Google Scholar]
- Wulandari, A.S.; Farzana, A.R. Mutu fisik dan teknik pematahan dormansi benih Kayu Kuku (Pericopsis mooniana (Thw.) Thw.). J. Trop. Silvic. 2020, 11, 199–205. [Google Scholar] [CrossRef]
- Suhartati; Alfaizin, D. Seed germination of Pericopsis mooniana Thw. based on color and scarification techniques. J. Perbenihan Tanam. Hutan 2017, 5, 115–124. [Google Scholar] [CrossRef][Green Version]
- Lemmens, R.H.M.; Soerianegara, I.; Wong, W. Plant Resources of South-East Asia. Timber Tress: Minor Commercial Timbers; Backhuys Publishers: Leiden, The Netherlands, 1994; Volume 5. [Google Scholar]
- Sidiyasa, K.; Soerianegara, I.; Martawijaya, A.; Sudo, S. Plant Resources of South-East Asia. Timber Tree: Major Commercial Timbers; PROSEA: Bogor, Indonesia, 1994; Volume 5. [Google Scholar]
- Yuniarti, N.; Syamsuwida, D. Kayu kuku (Pericopsis mooniana Thw). In Atlas Benih Tanaman Hutan Indonesia, 2nd ed.; Balai Penelitian Teknologi Perbenihan Tanaman Hutan: Bogor, Indonesia, 2011; Volume 5, pp. 32–34. [Google Scholar]
- Hardiyanto, E.B.; Na’iem, M. Present Status of Conservation, Utilization, and Management of Forest Genetic Resources in Indonesia. Available online: https://www.fao.org/3/ac648e/ac648e05.htm#TopOfPage (accessed on 17 July 2022).
- Lestari, D.; Santoso, W. Inventory and habitat study of orchids species in Lamedai Nature Reserve, Kolaka, Southeast Sulawesi. J. Biol. Divers. 2011, 12, 28–33. [Google Scholar] [CrossRef]
- Widyatmoko, D. Strategi dan inovasi konservasi tumbuhan Indonesia untuk pemanfaatan secara berkelanjutan. In Proceedings of the Seminar Nasional Pendidikan Biologi dan Saintek (SNPBS) ke-IV, Surakarta, Indonesia, 27 April 2019; pp. 1–22. [Google Scholar]
- Frankham, R. 2019 Conservation Genetics. In Encyclopedia of Ecology, 2nd ed; Elsevier: Oxford, UK, 2019; Volume 1, pp. 382–390. [Google Scholar]
- Coker, O.M. Importance of genetics in conservation of biodiversity. Niger. J. Wildl. Manag. 2017, 1, 11–18. [Google Scholar]
- Maia, R.T. Introductory Chapter: Genetic Variation-The Source of Biological Diversity. Available online: https://www.intechopen.com/chapters/75504 (accessed on 23 August 2022).
- Li, Y.; Liu, C.; Wang, R.; Luo, S.; Nong, S.; Wang, J.; Chen, X. Applications of molecular markers in conserving endangered. J. Species Biodivers. Sci. 2020, 28, 367–375. [Google Scholar]
- Nurtjahjaningsih, I.; Widyatmoko, A.; Rimbawanto, A. Genetic diversity of kayu kuku (Pericopsis mooniana (Thwaites) Thwaites) population at Lamedai forest Southeast Sulawesi based on RAPD markers. J. Pemuliaan Tanam. Hutan 2019, 13, 25–32. [Google Scholar] [CrossRef]
- Yudohartono, T.P.; Fiani, A.; Yuliah. Genetic diversity of kayu kuku (Pericopsis mooniana Thw) from Southeast Sulawesi provenance observed at seedling stage. J. Pemuliaan Tanam. Hutan 2019, 13, 11–17. [Google Scholar] [CrossRef]
- Yuskianti, V.; Sulistyawati, P. Genetic diversity of Pericopsis mooniana from South Kalimantan based on Random Amplified Polymorphism DNA (RAPD) Markers. IOP Conf. Ser. Earth Environ. Sci. 2021, 914, 012029. [Google Scholar] [CrossRef]
- KPHP Model Sorong. Rencana Pengelolaan Hutan Jangka Panjang KPHP Model Sorong. Available online: http://kph.menlhk.go.id/sinpasdok/public/RPHJP/RPHJP_SORONG_SELATAN.pdf (accessed on 25 August 2022).
- Yuskianti, V. Genetic conservation of a threatened species, Kayu kuku (Pericopsis mooniana) in Indonesia. In IUFRO-INAFOR Joint International Conference 2017 Promoting Sustainable Resources from Plantation for Economic Growth and Community Benefits; FORDIA Ministry of Environment and Forestry Republic of Indonesia: Bogor, Indonesia, 2018; pp. 403–410. [Google Scholar]
- Amandita, F.Y.; Rembold, K.; Vornam, B.; Rahayu, S.; Siregar, I.Z.; Kreft, H.; Finkeldey, R. DNA barcoding of flowering plants in Sumatra, Indonesia. Ecol. Evol. 2019, 9, 1858–1868. [Google Scholar] [CrossRef]
- Suhartati; Alfaizin, D. Cultivation of Pericopsis mooniana Thw Case Study: KHDTK Malili, Luwu Timur Regency. IOP Conf. Ser. Earth Environ. Sci. 2020, 486, 012106. [Google Scholar] [CrossRef]
- Heywood, V.H. An overview of in situ conservation of plant species in the Mediterranean Flora. J. Mediterr 2014, 24, 5–24. [Google Scholar]
- Sulistyawati, P.; Widyatmoko, A. Keragaman genetik populasi Kayu Merah (Pterocarpus indicus Willd) menggunakan penanda Random Amplified Polymorphism DNA. J. Pemuliaan Tanam. Hutan 2017, 11, 67–76. [Google Scholar] [CrossRef]
- Widyatmoko, A.Y.P.B.C.; Aprianto, A. Keragaman genetik Gonystylus bancanus (miq.) kurz berdasarkan penanda RAPD (Random Amplified polymorphic DNA). J. Pemuliaan Tanam. Hutan 2013, 7, 53–71. [Google Scholar] [CrossRef]
- Hakim, L.; Widyatmoko, A.Y.P.B.C. Strategi konservasi ex-situ jenis ulin (Eusideroxylon zwageri), Eboni (Diospyros celebica), dan Cempaka (Michelia spp.). In Status Konservasi Dan Formulasi Strategi Konservasi Jenis-Jenis Pohon Terancam Punah (Ulin, Eboni Dan Michelia), 1st ed.; Bismark, M., Murniati, Eds.; Pusat Penelitian dan Pengembangan Konservasi dan Rehabilitasi Badan Litbang Kehutanan dan ITTO: Bogor, Indonesia, 2011; pp. 163–177. [Google Scholar]
- Byrne, M. A molecular journey in conservation genetics. Pacific. Conserv. Biol. 2018, 24, 235–243. [Google Scholar] [CrossRef]
- Singh, P.; Sharma, H.; Nag, A.; Bhau, B.S.; Sharma, R.K. Development, and characterization of polymorphic microsatellites markers in endangered Aquilaria malaccensis. Conserv. Genet. Resour. 2015, 7, 61–73. [Google Scholar] [CrossRef]
- Lee, C.T.; Lee, S.L.; Tnah, L.H.; Ng, K.K.S.; Ng, C.H.; Cheng, S.; Tani, N. Isolation and characterization of 16 microsatellite markers in Intsia palembanica, a high-value tropical hardwood species. Conserv. Genet. Resour. 2014, 6, 389–391. [Google Scholar] [CrossRef]
- Rimbawanto, A.; Widyatmoko, A. Identifikasi Aquilaria malaccensis dan A. microcarpa menggunakan Penanda RAPD. J. Pemuliaan Tanam. Hutan 2011, 5, 23–30. [Google Scholar] [CrossRef]
- Widyatmoko, A.Y.P.B.C.; Shiraishi, S. Development of SCAR markers for identification of teak clones. AIP Conf. Proc. 2019, 2120, 030001. [Google Scholar] [CrossRef]
- Widyatmoko, A.Y.P.B.C.; Rimbawanto, A.; Suharyanto. Keragaman genetik Araucaria cuminghamii menggunakan penanda RAPD. In Proceedings Seminar Nasional Peningkatan Produktivitas Hutan: Peran Konservasi Sumberdaya Genetik, Pemuliaan dan Silvikultur dalam Mendukung Rehabilitasi Hutan; UGM dan ITTO: Yogyakarta, Indonesia, 2005; pp. 397–408. [Google Scholar]
- Widyatmoko, A.Y.P.B.C.; Afritanti, R.; Taryono; Rimbawanto, A. Keragaman genetik lima populasi Gyrinops verstegii di Lombok menggunakan penanda RAPD. J. Pemuliaan Tanam. Hutan 2009, 3, 1–10. [Google Scholar] [CrossRef]
- Hartati, D.; Rimbawanto, A.; Taryono; Sulistyaningsih, E.; Widyatmoko, A.Y.P.B.C. Pendugaan keragaman genetik di dalam dan antar provenan pulai (Alstonia scholaris (L.) R. Br.) menggunakan penanda RAPD. J. Pemuliaan Tanam. Hutan 2007, 1, 89–98. [Google Scholar]
- Liu, U.; Cossu, T.A.; Davies, R.M.; Forest, F.; Dickie, J.B.; Breman, E. Conserving orthodox seeds of globally threatened plants ex situ in the Millennium Seed Bank Royal Botanic Gardens, Kew, UK: The status of seed collections. Biodivers. Conserv. 2020, 29, 2901–2949. [Google Scholar] [CrossRef]
- Laure, N.M.; Samuel, G.L.; Francois, M.; Emmanuel, E.; Din, N. Phenological Behavior of Pericopsis elata in a Production Forest at Libongo (Southeast Cameroon). Int. J. Sci. Res. Methodol. 2019, 12, 9–26. [Google Scholar]
- Angbonda, D.M.A.; Monthe, F.K.; Bourland, N.; Boyemba, F.; Hardy, O.J. Seed and pollen dispersal and fine-scale spatial genetic structure of a threatened tree species: Pericopsis elata (HARMS) Meeuwen (Fabaceae). Tree Genet. Genomes 2021, 17, 1–14. [Google Scholar] [CrossRef]
- Tandoh, P.; Banful, B.; Gaveh, E.; Amponsah, J. Effects of packaging materials and storage periods on seed quality and longevity dynamics of Pericopsis elata seeds. Environ. Earth Ecol. 2017, 1, 27–38. [Google Scholar] [CrossRef]
- Nkengurutse, J.; Khalid, A.; Mzabri, I.; Kakunze, A.C.; Masharabu, T.; Berrichi, A. Germination optimization study of five indigenous fabaceae tree species from Burundi miombo woodlands. J. Mater. Environ. Sci. 2016, 7, 4391–4403. [Google Scholar]
- Romdyah, N.L.; Riniarti, M.; Asmarahman, C.; Yuwono, S.B. Skarifikasi Awal Dan Penambahan Beberapa Jenis Zat Pengatur Tumbuh Untuk Percepatan Perkecambahan Benih Kayu Kuku (Pericopsis moonianna Thw). EnviroScienteae 2020, 16, 296–308. [Google Scholar] [CrossRef]
- Nursyamsi. Teknik skarifikasi benih Kayu Kuku (Pericopsis Mooniana Thw) untuk mematahkan dormansi melalui kultur jaringan. In Proceedings of the Seminar Nasional from Basic Science to Comprehensive Education, Makassar, Indonesia, 26 August 2016. [Google Scholar]
- Araújo, S.d.S.; Paparella, S.; Dondi, D.; Bentivoglio, A.; Carbonera, D.; Balestrazzi, A. Physical methods for seed invigoration: Advantages and challenges in seed technology. Front. Plant Sci. 2016, 7, 1–12. [Google Scholar] [CrossRef]
- Adetunji, A.E.; Sershen; Varghese, B.; Pammenter, N. Effects of exogenous application of five antioxidants on vigour, viability, oxidative metabolism and germination enzymes in aged cabbage and lettuce seeds. S. Afr. J. Bot. 2021, 137, 85–97. [Google Scholar] [CrossRef]
- Siregar, I.Z.; Muharam, K.F.; Purwanto, Y.A.; Sudrajat, D.J. Seed germination characteristics in different storage time of gmelina arborea treated with ultrafine bubbles priming. Biodiversitas 2020, 21, 4558–4564. [Google Scholar] [CrossRef]
- Prasetyawati, C.A.; Nursyamsi; Alfaizin, D. Effect of seed storage methods on germination growth of Pericopsis mooniana thw. through in-vitro technique. IOP Conf. Ser. Earth Environ. Sci. 2020, 473, 6–11. [Google Scholar] [CrossRef]
- Sukendro, A.; Ayuning, D. Study of vegetative propagation on Pericopsis Mooniana Thw with cutting. J. Silvikultur Trop. 2016, 7, 53–57. [Google Scholar]
- Nursyamsi; Toaha, A.Q. Tahapan sterilisasi dan skarifikasi benih kayu kuku (Pericopsis mooniana THW) untuk mempercepat perkecambahan secara in vitro. J. Info Tek. Eboni 2017, 14, 11–22. [Google Scholar]
- Abeyratne, W.; Bandara, D.; Senanayake, Y.D. In vitro Propagation of Nadun (Pericopsis mooniana). Trop. Agric. Res. 1990, 2, 20–35. [Google Scholar]
- Indravathi, G. In vitro propagation studies of Albizia amara (Roxb.) Boiv. African J. Plant Sci. 2013, 7, 1–8. [Google Scholar] [CrossRef]
- Pereira, S.R.; Laura, V.A.; Souza, A.L.T. Establishment of Fabaceae tree species in a tropical pasture: Influence of seed size and weeding methods. Restor. Ecol. 2013, 21, 67–74. [Google Scholar] [CrossRef]
- Aini, F.N.; Sulistyowati, E. Mewaspadai hama minor pada Kakao: Zeuzera coffae Nietner. War. Pus. Penelit. Kopi dan Kakao Indones. 2016, 28, 24–29. [Google Scholar]
- Fiani, A.; Yuliah; Pamungkas, T. Inventarisasi jenis hama yang menyerang bibit Kayu Kuku (Pericopsis mooniana) umur satu tahun di persemaian. In Proceedings of the Seminar Nasional Pendidikan Biologi Dan Saintek (Snpbs), Surakarta, Indonesia, 27 April 2019; pp. 59–65. [Google Scholar]
- Roychoudhury, N.; Mishra, R.K. Coffee borer, Zeuzera coffeae: Its pest profile and use as tribal food. Van Sangyan 2022, 9, 24–26. [Google Scholar]
- Wiratno; Deciyanto, S. Ciri-ciri dan siklus hidup serangga penggulung daun nilam, Sylepta sp. (Lepidoptera:Pyralidae). Bul. Littro 1991, 1, 15–19. [Google Scholar]
- Tawaraya, K.; Turjaman, M. Use of arbuscular mycorrhizal fungi for reforestation of Degraded Tropical Forests. In Mycorrhizal Fungi: Use in Sustainable Agriculture and Land Restoration; Springer: Berlin/Heidelberg, Germany, 2014; pp. 357–373. [Google Scholar]
- Asmelash, F.; Bekele, T.; Birhane, E. The potential role of arbuscular mycorrhizal fungi in the restoration of degraded lands. Front. Microbiol. 2016, 7, 1–15. [Google Scholar] [CrossRef]
- Husna; Tuheteru, F.D.; Arif, A. Arbuscular mycorrhizal fungi and growth on serpentine solis. In Arbuscular Mycorrhizas and Stress Tolerance of Plants; Springer Nature: Singapore, 2017; pp. 293–303. [Google Scholar]
- Riaz, M.; Kamran, M.; Fang, Y.; Wang, Q.; Cao, H.; Yang, G.; Deng, L.; Wang, Y.; Zhou, Y.; Anastopoulos, I.; et al. Arbuscular mycorrhizal fungi-induced mitigation of heavy metal phytotoxicity in metal contaminated soils: A critical review. J. Hazard. Mater. 2021, 402, 123919. [Google Scholar]
- Faad, H.; Tuheteru, F.D.; Arif, A. Arbuscular mycorrhizal fungi symbiosis and conservation of endangered tropical Legume Trees. In Root Biology, Soil Biology 52; Giri, B., Prasad, R., Varma, A., Eds.; Springer International Publishing: Singapore, 2018; pp. 465–486. [Google Scholar]
- Husna; Tuheteru, F.D.; Arif, A. The potential of arbuscular mycorrhizal fungi to conserve Kalappia celebica, an endangered endemic legume on gold mine tailings in Sulawesi, Indonesia. J. For. Res. 2021, 32, 675–682. [Google Scholar] [CrossRef]
- Husna; Tuheteru, F.D.; Arif, A.; Basrudin; Albasari. Biodiversity of arbuscular mycorrhizal fungi in tropical Indonesia. In An Introduction to Microorganisms, 1st ed.; Wu, Q., Zou, Y., Zhang, F., Shu, B., Eds.; Nova Science Publisher Inc: Hauppauge, NY, USA, 2021; pp. 201–247. [Google Scholar]
- Husna; Wilarso, S.B.; Mansur, I.; Kusmana, C.; Kramadibrata, K. Fungi mikoriza arbuskula pada rizosfer Pericopsis mooniana (thw.) thw. di Sulawesi Tenggara. Ber. Biol. 2014, 13, 263–273. [Google Scholar]
- Husna; Wilarso, S.B.; Mansur, I.; Kusmana, C. Diversity of arbuscular mycorrhizal fungi in the growth habitat of Kayu Kuku (Pericopsis mooniana Thw.) in Southeast Sulawesi. Pakistan. J. Biol. Sci. 2015, 18, 1–10. [Google Scholar]
- Husna; Tuheteru, F.D.; Basri, A.; Arif, A.; Basruddin; Umar, Y. Serapan hara tanaman kayu kuku (Pericopsis moonia Thw) bermikoriza pada interval penyiraman berbeda. J. Hutan Trop. 2020, 15, 88–101. [Google Scholar]
- Husna; Tuheteru, F.D.; Arif, A. Arbuscular mycorrhizal fungi to enhance the growth of tropical endangered species Pterocarpus indicus and Pericopsis mooniana in post gold mine field in Southeast Sulawesi, Indonesia. Biodiversitas 2021, 22, 3844–3853. [Google Scholar]
- Kusuma, G.A.; Husna; Mamma, S. The Response of kayu kuku (Pericopsis mooniana (Thw).Thw.) to mycorrhizae and the level of water availability in gold tailing media. J. Berk. Penelit. Agron. 2022, 10, 68–84. [Google Scholar]
- Tuheteru, F.D.; Husna; Nuranisa; Basrudin; Arif, A.; Albasri; Safitri, I.; Karepesina, S.; Nurdin, W. Effects of arbuscular mycorrhizal fungi on nutrient uptake by Pericopsis mooniana in media post gold mining land. J. Trop. Mycorrhiza 2022, 1, 9–16. [Google Scholar]
- Husna. Pertumbuhan Bibit Kayu Kuku (Pericopsis Mooniana Thw) Melalui Aplikasi Fungi Mikoriza Arbuskula (FMA) Dan Ampas Sagu Pada Media Bekas Tambang Nikel; Halu Oleo University: Kendari, Indonesia, 2010. [Google Scholar]
- Husna; Budi, S.W.; Mansur, I.; Kusmana, C. Growth and nutrient status of kayu kuku [Pericopsis mooniana (Thw.) Thw] with mycorrhiza in soil media of nickel post mining site. Pakistan. J. Biol. Sci. 2016, 19, 158–170. [Google Scholar] [CrossRef]
- Husna; Tuheteru, F.D.; Salim, M.I.; Adam, L.A.; Nirwan; Arif, A.; Basrudin; Albasri. Pemanfaatan pupuk hayati mikoriza lokal Sulawesi Tenggara untuk memacu pertumbuhan Kayu Kuku pada Media tanah pascatambang nikel. In Proceedings Seminar Nasional Mikoriza: Mikoriza Untuk Pembangunan Pertanian dan Kehutanan Berkelanjutan; UHO Edu Press: Kendari, Indonesia, 2021; pp. 101–114. [Google Scholar]
- Husna; Tuheteru, F.D.; Wulandari, J.; Arif, A.; Basrudin; Albasari; Nurdin, W. Respons pertumbuhan semai Kayu Kuku (Pericopsis Mooniana ( Thw .) Thw .) dengan inokulasi fungi mikoriza arbuskula ( FMA ) lokal dan urea pada media tanah serpentine. In Proceedings Seminar Nasional: Mikoriza Untuk Pembangunan Pertanian Dan Kehutanan Berkelanjutan; UHO Edu Press: Kendari, Indonesia, 2021; pp. 129–148. [Google Scholar]
- Husna; Mansur, I.; Budi, S.W.; Tuheteru, F.D.; Arif, A.; Tuheteru, E.J.; Albasri. Effects of arbuscular mycorrhizal fungi and organic material on growth and nutrient uptake by pericopsis mooniana in coal mine. Asian J. Plant Sci. 2019, 18, 101–109. [Google Scholar] [CrossRef]
- Iskandar, F. Peningkatan Kualitas Bibit Kayu Kuku (Pericopsis Mooniana Thwaites) Yang Diberi Fungi Mikoriza Arbuskula Dan Tepung Tulang; Skripsi, Haluoleo University: Kendari, Indonesia, 2010. [Google Scholar]
- Husna; Raharjo, W.P.; Tuheteru, F.D.; Arif, A.; Basrudin; Albasri; Nurdin, W. Perbaikan kualitas bibit kayu kuku (Pericopsis Mooniana ( Thw .) Thw. dengan aplikasi fungi mikoriza arbuskula ( Fma ) lokal Sulawesi Tenggara. In Proceedings Seminar Nasional: Mikoriza Untuk Pembangunan Pertanian Dan Kehutanan Berkelanjutan; UHO Edu Press: Kendari, Indonesia, 2021; pp. 251–264. [Google Scholar]
- Husna; Wilarso, S.; Mansur, I.; Kusmana, C. Respon pertumbuhan bibit kayu kuku (Pericopsis mooniana (Thw.)Thw) terhadap inokulasi fungi mikoriza arbuskula lokal. J. Pemuliaan Tanam. Hutan 2015, 9, 131–148. [Google Scholar] [CrossRef][Green Version]
- Husna. Potensi Fungi Mikoriza Arbuskula (FMA) Lokal Dalam Konservasi Ex-Situ Jenis Terancam Punah Kayu Kuku [Pericopsis mooniana (Thw.) Thw.]. Disertasi, Bogor Agricultural University, Bogor, Indonesia, 2015. [Google Scholar]
- Husna; Tuheteru, F.D.; Wigati, E. Short communication: Growth response and dependency of endangered nedun tree species (Pericopsis mooniana) affected by indigenous arbuscular mycorrhizal fungi inoculation. Nusant. Biosci. 2017, 9, 57–61. [Google Scholar] [CrossRef]
- Husna; Saputra, A.; Tuheteru, F.D.; Arif, A. Keberhasilan hidup tanaman kayu kuku bermikoriza umur tiga tahun pada lahan pascatambang nikel PT Vale Indonesia. In Proceedings Seminar Nasional: Mikoriza untuk Pembangunan Pertanian Dan Kehutanan Berkelanjutan; UHO Edu Press: Kendari, Indonesia, 2021; pp. 237–250. [Google Scholar]
- Wijesundara, T.I.L.; Van Holm, L.H.J.; Kulasooriya, S.A. Rhizobiology and nitrogen fixation of some tree legumes native to Sri Lanka. Biol. Fertil. Soils 2000, 30, 535–543. [Google Scholar] [CrossRef]
- Husna; Hamirul, H.; Arief, L. Uji Efektivitas VA Mikoriza Dan Rhizobium Terhadap Pertumbuhan Semai Kayu Kuku (Pericopsis mooniana Thw.); Departemen Pendidikan dan Kebudayaan: Jakarta, Indonesia, 1994. [Google Scholar]
- Javaid, A. Microbes for legume improvement microbes legum. In Microbes for Legume Improvement; Khan, M.S., Zaidi, A., Musarrat, J., Eds.; Springer: Vienna, Austria, 2010; pp. 409–426. [Google Scholar]
- Prihatini, I.; Widyatmoko, A.Y.P.B.C.; Nurtjahjaningsih, I.L.G.; Yuskianti, V.; Danarto, S.A. DNA barcoding of Pericopsis mooniana from two different populations in Indonesia based on rDNA ITS (Internal Transcribed Spacer). IOP Conf. Ser. Mater. Sci. Eng. 2020, 935, 012024. [Google Scholar] [CrossRef]
- Zanzibar, M.; Nugraheni, Y.M.M.A.; Putri, K.P.; Yuniarti, N.; Sudrajat, D.J.; Yulianti; Syamsuwida, D.; Aminah, A.; Rustam, E.; Sukendro, A. Toona sureni (blume) merr. seeds invigoration using 60Co Gamma-Rays irradiation. Forest. Sci. Technol. 2021, 17, 80–87. [Google Scholar]
- Rocha, M.; Singh, N.; Ahsan, K.; Beiriger, A.; Prince, V.E. Neural crest development: Insights from the zebrafish. Dev. Dyn. 2020, 249, 88–111. [Google Scholar] [CrossRef]
- Elias. Dampak penerapan RIL terhadap simpanan karbon di hutan pegunungan tropis. In Proceedings of the Seminar Hasil Penelitian Teknologi Dan Inovasi Keteknikan Kehutanan Dan Pengolahan Hasil Hutan Dalam Menunjang Industri Pengolahan Hasil Hutan; Badan Penelitian dan Pengembangan dan Inovasi, Kementerian Lingkungan Hidup dan Kehutanan: Bogor, Indonesia, 2015; pp. 3–14. [Google Scholar]
- Ellis, P.W.; Gopalakrishna, T.; Goodman, R.C.; Putz, F.E.; Roopsind, A.; Umunay, P.M.; Zalman, J.; Ellis, E.A.; Mo, K.; Gregoire, T.G.; et al. Reduced-impact logging for climate change mitigation (RIL-C) can halve selective logging emissions from tropical forests. For. Ecol. Manag. 2019, 438, 255–266. [Google Scholar] [CrossRef]
- Suhartana, S.; Yuniawati; Rahmat. Pemanenan kayu ramah lingkungan di hutan tanaman lahan kering di Sumatera, Kalimantan dan Jawa Barat. In Proceedings of the Seminar Nasional MAPEKI 16: Pemanfaatan sumberdaya terbarukan untuk Kesejahteraan Manusia dan Kelestarian Lingkungan; Masyarakat Peneliti Kayu Indonesia: Balikpapan, Indonesia, 2014; pp. 402–406. [Google Scholar]
- Bicknell, J.E.; Struebig, M.J.; Edwards, D.P.; Davies, Z.G. Improved timber harvest techniques maintain biodiversity in tropical forests. Curr. Biol. 2014, 24, 119–120. [Google Scholar] [CrossRef]
- Direktorat Jenderal Pengelolaan Hutan Produksi Standar dan pedoman pelaksanaan penilaian kinerja pengelolaan hutan produksi lestari (PHPL) dan verifikasi legalitas kayu (VLK). Available online: https://silk.menlhk.go.id/app/Upload/hukum/20160801/24fbdaeef1ae1cd12488cb4051740dce.pdf (accessed on 26 October 2022).
- Soenarno; Yuniawati; Dulsalam; Suhartana, S. Determination of standardized natural forest exploitation factor and logging waste of Sub Region Central Kalimantan Province. J. Penelit. Has. Hutan 2021, 39, 155–169. [Google Scholar]
- Soenarno; Endom, W.; Bustomi, S. Residual stand damage due to timber harvesting on hilly tropical forest inCentral Kalimantan. J. Penelit. Has. Hutan 2017, 35, 273–288. [Google Scholar]
- Mousavi, M.R. Comparison of damage to residual stand due to applying two different harvesting methodsin the Hyrcanian forest of Iran: Cut-to-length vs. tree length. Casp. J. Environ. Sci. 2017, 15, 13–27. [Google Scholar]
- Sist, P.; Nolan, T.; Bertault, J.G.; Dykstra, D. Harvesting intensity versus sustainability in Indonesia. For. Ecol.Manage. 1998, 108, 251–260. [Google Scholar] [CrossRef]
- Rodríguez-Calcerrada, J.; P´erez-Ramos, I.M.; Ourcival, J.-M.; Limousin, J.-M.; Joffre, R.; Rambal, S. Is selective thinning an adequate practice for adapting Quercus ilex coppices to climate change? Ann. For. Sci. 2011, 68, 575–585. [Google Scholar] [CrossRef]
- Pakhriazad, H.; Shinohara, T.; Nakama, Y.; Ykutake, K. A Selective Management System (SMS): A case study in the implementation of SMS in managing the dipterocarp forests of Peninsular Malaysia. Kyushu J.For.Res 2004, 57, 39–44. [Google Scholar]
- Yuniawati; Dulsalam; Andini, S. Potential carbon storage and CO2 emissions due to felling in Papua natural forest. J. Penelit. Has. Hutan 2022, 40, 61–73. [Google Scholar]
- Elias. Pengertian dan perkembangan IPTEKS pemanenan kayu; PT. Media Anugrah Perkasa: Jakarta, Indonesia, 2015. [Google Scholar]
- Galante, M.V.; Dutschke, M.; Patenaude, G.; Vickers, B. Climate change mitigation through reduced-impact logging and the hierarchy of production forest management. Forests 2012, 3, 59–74. [Google Scholar] [CrossRef]
- Abdulah, L. Metode Pendugaan Konsumsi Kayu Untuk Penggunaan Rumah Tangga; Bogor Agricultural University: Bogor, Indonesia, 2020. [Google Scholar]
- Abdulah, L.; Suhendang, E.; Purnomo, H.; Matangaran, J.R. The role of urban household wood product consumption on forest management and its impact: A system modelling simulation approach in Bogor City. IOP Conf. Ser. Earth Environ. Sci. 2021, 800, 1–12. [Google Scholar] [CrossRef]
- Abdulah, L.; Suhendang, E.; Purnomo, H.; Matangaran, J.R. Measuring the sustainability of wood consumption at the household level in indonesia: Case study in Bogor, Indonesia. Biodiversitas 2020, 21, 457–464. [Google Scholar] [CrossRef]
- Abdulah, L.; Irawan, A.; Arini, D.I.D.; Suryaningsih, R. The Use of Marketing Monitoring Web System Application to Estimate Cempaka Wood Product Consumption Level in North Sulawesi, Indonesia. IOP Conf. Ser. Earth Environ. Sci. 2021, 891, 1–13. [Google Scholar] [CrossRef]
- Abdulah, L.; Yulianti, M.; Mindawati. How the growth of natural forests meets Indonesia’s demand for wood for construction and household furniture? IOP Conf. Ser. Earth Environ. Sci. 2022, 959, 012070. [Google Scholar] [CrossRef]
- Seng, O. Specific Gravity of Indonesian Woods and Its Significance for Parctical Use; Pusat Penelitian dan Pengembangan Hasil Hutan, Departemen Kehutanan: Jakarta, Indonesia, 1990. [Google Scholar]
- Munandar Budidaya Tanaman Kehutanan Jenis Kuku (Pericopsis mooniana Thwaites). Available online: https://www.mounandar.blogspot.com/2010/06/budidaya-tanaman-kehutananjenis-kuku.html (accessed on 14 July 2015).
- Krisdianto; Dewi, L.M. Jenis Kayu Untuk Mebel, 1st ed.; Pusat Penelitian dan Pengembangan Keteknikan Kehutanan dan Pengolahan Hasil Hutan, Badan Penelitian dan Pengembangan Kehutanan-Kementerian Kehutanan: Bogor, Indonesia, 2012. [Google Scholar]
- Pericopsis elata (Harms) Meeuwen. Available online: https://www.prota4u.org/database/protav8.asp?g=psk&p=Pericopsis%20elata (accessed on 18 September 2022).
- Djarwanto; Damayanti, R.; Balfas, J.; Basri, E.; Jasni, I.M.S.; Andianto; Martono, D.; Pari, G.; Sopandi, A.; Mardiansyah. Pengelompokan Jenis Kayu Perdagangan, 1st ed.; Forda Press: Bogor, Indonesia, 2017; p. 26. [Google Scholar]
- Ishiguri, F.; Wahyudi, I.; Takeuchi, M.; Takashima, Y.; Iizuka, K.; Yokota, S.; Yoshizawa, N. Wood properties of Pericopsis mooniana grown in a plantation in Indonesia. J. Wood Sci. 2011, 57, 241–246. [Google Scholar] [CrossRef]
- Anoop, E.V.; Sindhumathi, C.R.; Jijeesh, C.M.; Jayasree, C.E. Radial variation in wood properties of nedun (Pericopsis mooniana), an introduced species to South India. J. Trop. Agric. 2016, 54, 27–34. [Google Scholar]
- Martawijaya, A.; Kartasujana, I. Ciri Umum, Sifat Dan Kegunaan Jenis-Jenis Kayu Indonesia, 1st ed.; Lembaga Penelitian Hasil Hutan: Bogor, Indonesia, 1977. [Google Scholar]
- Pandit, I.K.N. Karakteristik struktur anatomi Kayu Kuku (Pericopsis mooniana Thwaiters). Ilmu Teknol. Kayu Trop. 2005, 3, 1–5. [Google Scholar]
- Pericopsis elata (Harms) van Meeuwen (Afrormosia, Kokrodua)-CITES II. Available online: https://www.delta-intkey.com/citeswood/en/www/fabpeel.htm (accessed on 18 September 2022).
- Soerianegara, I.; Lemmens, R. Plant Resources of Southeast Asia. No 5(1): Timber Trees: Major Commercial Timbers; Pudoc Scientific Publishers: Wageningen, The Netherlands, 1993. [Google Scholar]
- Murodovna, R.; Sergeevna, K. Wood-As a Decorative Material of the Interior. Int. J. Integr. Educ. 2022, 5, 310–315. [Google Scholar]
- Burley, J.; Evans, J.; Youngquist, J. Encyclopedia of Forest Sciences, 1st ed.; Elsevier Academic Press: Oxford, UK, 2004. [Google Scholar]
- Zanuttini, R.; Negro, F. Wood-based composites: Innovation towards a sustainable future. Forests 2021, 12, 10–14. [Google Scholar] [CrossRef]
- Sabaruddin, B. Nilai Ekonomi Kayu Kuku. Available online: https://www.forestdigest.com/detail/933/nilai-ekonomi-kayu-kuku (accessed on 17 October 2022).
- Heropolo. Lamedae Nature Reserve. Available online: https://www.indonesianforest.or.id/lamedae-nature-reserve/ (accessed on 18 October 2022).
- Hermawan, B. Sustainable agroforestry models for proposed food production in post-mined land sites of South Sumatera. Int. J. Adv. Sci. Eng. Inf. Technol. 2016, 6, 245–251. [Google Scholar] [CrossRef]
- Guidelines for Species Conservation Planning. Available online: https://portals.iucn.org/library/sites/library/files/documents/2017-065.pdf (accessed on 5 September 2022).
- Maxted, N. In Situ, Ex Situ Conservation. Encycl. Biodivers. Second Ed. 2013, 4, 313–332. [Google Scholar]
- Rahman, W.; Brehm, J.M.; Maxted, N.; Phillips, J.; Cotreras-Toledo, A.R.; Faraji, M.; Quijano, M.P. Gap analyses of priority wild relatives of food crop in current ex situ and in situ conservation in Indonesia. Biodivers. Conserv. 2021, 30, 2827–2855. [Google Scholar] [CrossRef]
- Malhotra, N.; Panatu, S.; Singh, B.; Negi, N.; Singh, D.; Singh, M.; Chandora, R. Genetic Resources: Collection, Conservation, Characterization and Maintenance. In Lentils, 1st ed.; Singh, M., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 21–41. [Google Scholar]
- Rajpurohit, D.; Jhang, T. In Situ and Ex Situ conservation of plant genetic resources and traditional knowledge. In Plant Genetic Resources and Traditional Knowledge for Food Security, 1st ed.; Salgotra, R.K., Gupta, B.B., Eds.; Springer Science Business Media: Singapore, 2015; pp. 137–162. [Google Scholar]
- National Research Council and Board on Agriculture. In Situ Conservation of Genetic Resources Managing Global Genetic Resources: Agricultural Crop Issues and Policies; National Academic Press: Washington, DC, USA, 1993; pp. 1–17. [Google Scholar]
- Meyer, S.; Birnbaum, P.; Bruy, D.; Cazé, H.; Garnier, D.; Gâteblé, G.; Lannuzel, G.; McCoy, S.; Tanguy, V.; Veillon, J.M. The New Caledonia Plants RLA: Bringing Botanists Together for the Conservation of the archipelago’s Crown Jewel. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2022; pp. 859–874, in press. [Google Scholar]
- Ajayi, S.S.S. Principles for the management of protected areas. In Wildlife Conservation in Africa; Elsevier Inc.: Amsterdam, The Netherlands, 2019; pp. 85–93. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AMF Species/Treatment | Media | Period (Month) | Effect | Experiment Type | References |
|---|---|---|---|---|---|
| Septoglomus constrictum + two days of watering | Gold tailings media | 4 | Increase shoot P levels and uptake | Greenhouse | [82] |
| G. claroideum and G. coronatum | Gold tailings media | 4 | Increase the height diameter, number of nodules, dry weight of plants, and uptake of N and P in the roots | Greenhouse | [83] |
| G. claroideum + field capacity 100 and 75 % | Gold tailings media | 4 | Increase plant height, diameter, total leaf area, nodulation, and dry weight | Greenhouse | [84] |
| G. claroideum and G. coronatum | Gold tailings media | 4 | Increase the uptake of N, P, Mn, and Fe leaves | Greenhouse | [85] |
| 5 g AMF inoculum Mycofer (mix. Glomus manihotis, Glomus etunicatum, Acaulospora tuberculata, Gigaspora margarita) and addition sago waste 20 g | Soil media of nickel post-mining site | 3 | Increase height, dry weight, root nodules, and absorption of K and Ca and reduce Ni by 32% | Greenhouse | [86] |
| AMF native from rhizosfer P. mooniana | Soil media of nickel post-mining site | 5 | Increase growth and dry weight, absorption of N, P, and K in three parts of the plants, of Ca (in stems and leaves) and of Mg in leaf tissues and root nodules; and reduce Ni content | Greenhouse | [87] |
| C. etunicatum/Ha | Soil media of nickel post-mining site | 5 | Increase the height, diameter, number, and area of leaves, plant dry weight, and Seedlimh quality index | Greenhouse | [88] |
| C. etunicatum/Ha + without treatment of Urea | Soil media of nickel post-mining site | 5 | Increase the number of leaves, nodulation, and dry weight of shoots and roots | Greenhouse | [89] |
| AMF local from rhizosfer P. mooniana | Overburden coal media | Increase in the growth and dry weight of plants and also tend to increase the C, N, P, K, Ca, and Mg accumulation | Greenhouse | [90] | |
| Mycofer IPB (Glomus etunicatum, G. manihotis, Acaulospora tuberculata, and Gigaspora rosea) | Ultisol | 3 | Increase growth and dry weight | Greenhouse | [91] |
| Glomus sp. (HA) | Ultisol | 3 | Increase height, diameter, number of leaves, dry weight, root nodules, root, and leaf length | Greenhouse | [75] |
| Acaulospora sp. 1 (HA) | Ultisol | 3 | Increase growth in height, diameter, number, and area of leaves | Greenhouse | [92] |
| Local AMF from P. mooniana rhizosphere | Inceptisol | 5 | Increase in height, diameter, number of leaves and root nodules, total dry weight, total chlorophyll, P, K, Ca, and Mg uptake | Greenhouse | [93] |
| AMF from P. mooniana rhizosphere from Lamedai NR (non-serpentine) and PT. Vale Indonesia (serpentine) | Nickel post-mining land | 3 | Increase survival rate, growth, dry weight, and accumulation of N, P, and K | Field | [94] |
| 24 | Increase height and diameter | [95] | |||
| 36 | Increase growth in height, diameter, number, length, and width of leaves and dry weight of wet and dry leaves | [96] | |||
| G. coronatum | Gold post-mining land | 4 | Increase the height, diameter, and dry weight of the leaves as well as the uptake of N, P, K, Mn, and Fe | Field | [83] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kinho, J.; Suhartati; Husna; Tuheteru, F.D.; Arini, D.I.D.; Lawasi, M.A.; Ura’, R.; Prayudyaningsih, R.; Yulianti; Subarudi; et al. Conserving Potential and Endangered Species of Pericopsis mooniana Thwaites in Indonesia. Forests 2023, 14, 437. https://doi.org/10.3390/f14020437
Kinho J, Suhartati, Husna, Tuheteru FD, Arini DID, Lawasi MA, Ura’ R, Prayudyaningsih R, Yulianti, Subarudi, et al. Conserving Potential and Endangered Species of Pericopsis mooniana Thwaites in Indonesia. Forests. 2023; 14(2):437. https://doi.org/10.3390/f14020437
Chicago/Turabian StyleKinho, Julianus, Suhartati, Husna, Faisal Danu Tuheteru, Diah Irawati Dwi Arini, Moh. Andika Lawasi, Resti Ura’, Retno Prayudyaningsih, Yulianti, Subarudi, and et al. 2023. "Conserving Potential and Endangered Species of Pericopsis mooniana Thwaites in Indonesia" Forests 14, no. 2: 437. https://doi.org/10.3390/f14020437
APA StyleKinho, J., Suhartati, Husna, Tuheteru, F. D., Arini, D. I. D., Lawasi, M. A., Ura’, R., Prayudyaningsih, R., Yulianti, Subarudi, Abdulah, L., Susanti, R., Waluyo, T. K., Suhartana, S., Andianto, Wardani, M., Kalima, T., Tambaru, E., Isnan, W., ... Soenarno. (2023). Conserving Potential and Endangered Species of Pericopsis mooniana Thwaites in Indonesia. Forests, 14(2), 437. https://doi.org/10.3390/f14020437