
Specificity and Sensitivity of a Rapid LAMP Assay for Early Detection of Emerald Ash Borer (Agrilus planipennis) in Europe
 , , , ,
, , , ,  and
and
Abstract
1. Introduction
2. Methods
2.1. Collection and Extraction of Specimens
2.2. PCR Amplification and Sequencing of Samples
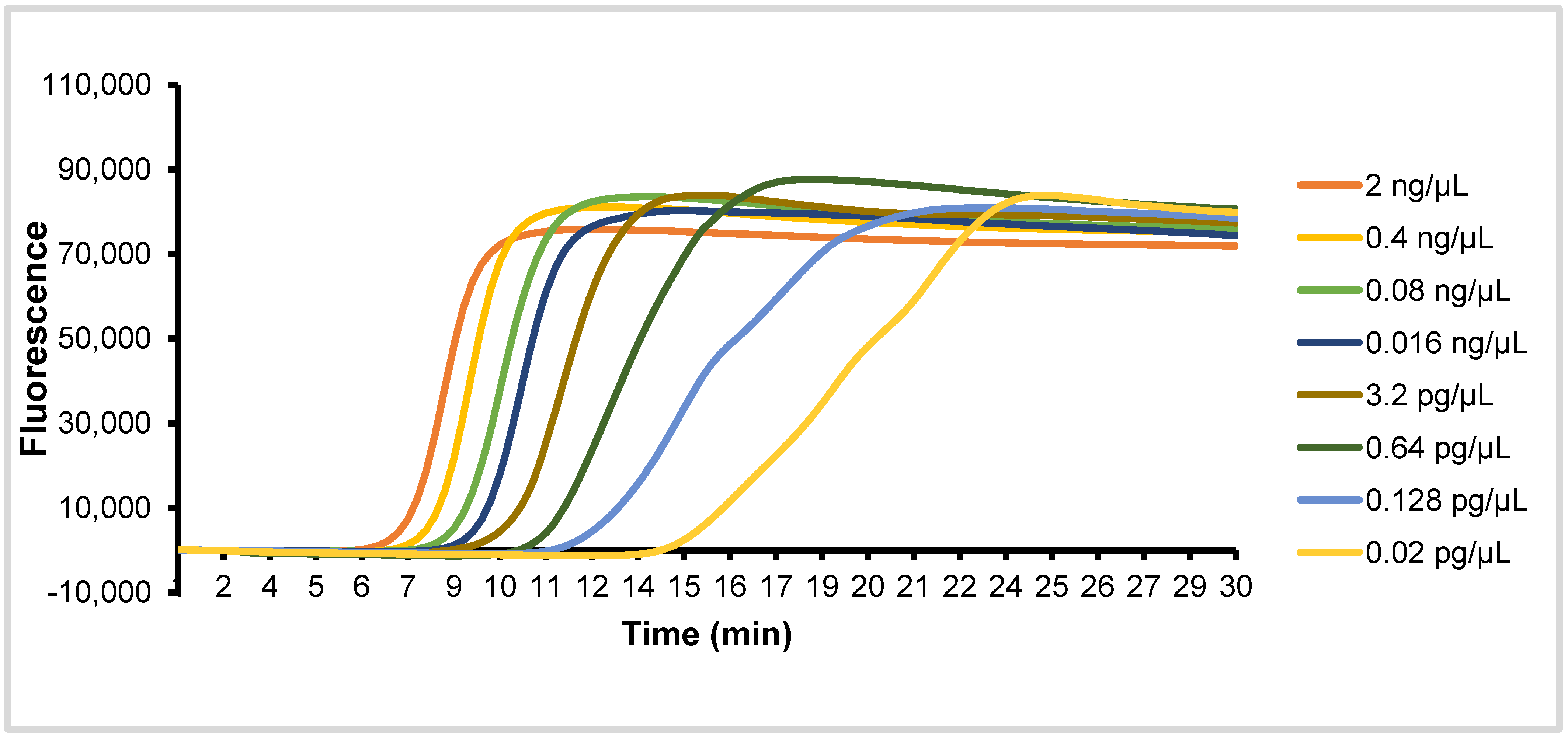
2.3. Specificity and Sensitivity Testing of EAB Assay
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gandhi, K.J.K.; Herms, D.A. Direct and Indirect Effects of Alien Insect Herbivores on Ecological Processes and Interactions in Forests of Eastern North America. Biol. Invasions 2010, 12, 389–405. [Google Scholar] [CrossRef]
- Aukema, J.E.; Leung, B.; Kovacs, K.; Chivers, C.; Britton, K.O.; Englin, J.; Frankel, S.J.; Haight, R.G.; Holmes, T.P.; Liebhold, A.M.; et al. Economic Impacts of Non-Native Forest Insects in the Continental United States. PLoS ONE 2011, 6, e24587. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, C.J.A.; Leroy, B.; Bellard, C.; Roiz, D.; Albert, C.; Fournier, A.; Barbet-Massin, M.; Salles, J.-M.; Simard, F.; Courchamp, F. Massive yet Grossly Underestimated Global Costs of Invasive Insects. Nat. Commun. 2016, 7, 12986. [Google Scholar] [CrossRef] [PubMed]
- Santini, A.; Ghelardini, L.; De Pace, C.; Desprez-Loustau, M.L.; Capretti, P.; Chandelier, A.; Cech, T.; Chira, D.; Diamandis, S.; Gaitniekis, T.; et al. Biogeographical Patterns and Determinants of Invasion by Forest Pathogens in Europe. New Phytol. 2013, 197, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Epanchin-Niell, R.; McAusland, C.; Liebhold, A.; Mwebaze, P.; Springborn, M.R. Biological Invasions and International Trade: Managing a Moving Target. Rev. Environ. Econ. Policy 2021, 15, 180–190. [Google Scholar] [CrossRef]
- Venette, R.C.; Hutchison, W.D. Invasive Insect Species: Global Challenges, Strategies & Opportunities. Front. Insect Sci. 2021, 1, 650520. [Google Scholar]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M.; et al. No Saturation in the Accumulation of Alien Species Worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef]
- Tobin, P.C.; Kean, J.M.; Suckling, D.M.; McCullough, D.G.; Herms, D.A.; Stringer, L.D. Determinants of Successful Arthropod Eradication Programs. Biol. Invasions 2014, 16, 401–414. [Google Scholar] [CrossRef]
- NISC 2016–2018 NISC Management Plan; National Invasive Species Council: Washington, DC, USA, 2016.
- Martinez, B.; Reaser, J.K.; Dehgan, A.; Zamft, B.; Baisch, D.; McCormick, C.; Giordano, A.J.; Aicher, R.; Selbe, S. Technology Innovation: Advancing Capacities for the Early Detection of and Rapid Response to Invasive Species. Biol. Invasions 2020, 22, 75–100. [Google Scholar] [CrossRef]
- Sallé, A.; Nageleisen, L.-M.; Lieutier, F. Bark and Wood Boring Insects Involved in Oak Declines in Europe: Current Knowledge and Future Prospects in a Context of Climate Change. For. Ecol. Manag. 2014, 328, 79–93. [Google Scholar] [CrossRef]
- Haack, R.A.; Petrice, T. Historical Population Increases and Related Inciting Factors of Agrilus Anxius, Agrilus Bilineatus, and Agrilus Granulatus Liragus (Coleoptera: Buprestidae) in the Lake States (Michigan, Minnesota, and Wisconsin). Great Lakes Entomol. 2019, 52, 7. [Google Scholar]
- Muilenburg, V.L.; Herms, D.A. A Review of Bronze Birch Borer (Coleoptera: Buprestidae) Life History, Ecology, and Management. Environ. Entomol. 2012, 41, 1372–1385. [Google Scholar] [CrossRef]
- Coleman, T.W.; Seybold, S.J. Previously Unrecorded Damage to Oak, Quercus Spp., in Southern California by the Goldspotted Oak Borer, Agrilus Coxalis Waterhouse (Coleoptera: Buprestidae). Pan-Pac. Entomol. 2008, 84, 288–300. [Google Scholar] [CrossRef]
- Cappaert, D.; McCullough, D.G.; Poland, T.M.; Siegert, N.W. Emerald Ash Borer in North America: A Research and Regulatory Challenge. Am. Entomol. 2005, 51, 152–165. [Google Scholar] [CrossRef]
- Kovacs, K.F.; Haight, R.G.; McCullough, D.G.; Mercader, R.J.; Siegert, N.W.; Liebhold, A.M. Cost of Potential Emerald Ash Borer Damage in U.S. Communities, 2009–2019. Ecol. Econ. 2010, 69, 569–578. [Google Scholar] [CrossRef]
- Klooster, W.S.; Herms, D.A.; Knight, K.S.; Herms, C.P.; McCullough, D.G.; Smith, A.; Gandhi, K.J.K.; Cardina, J. Ash (Fraxinus Spp.) Mortality, Regeneration, and Seed Bank Dynamics in Mixed Hardwood Forests Following Invasion by Emerald Ash Borer (Agrilus Planipennis). Biol. Invasions 2014, 16, 859–873. [Google Scholar] [CrossRef]
- Baranchikov, Y.; Mozolevskaya, E.; Yurchenko, G.; Kenis, M. Occurrence of the Emerald Ash Borer, Agrilus Planipennis in Russia and Its Potential Impact on European Forestry. EPPO Bull. 2008, 38, 233–238. [Google Scholar] [CrossRef]
- Orlova-Bienkowskaja, M.J.; Drogvalenko, A.N.; Zabaluev, I.A.; Sazhnev, A.S.; Peregudova, E.Y.; Mazurov, S.G.; Komarov, E.V.; Struchaev, V.V.; Martynov, V.V.; Nikulina, T.V. Current Range of Agrilus Planipennis Fairmaire, an Alien Pest of Ash Trees, in European Russia and Ukraine. Ann. For. Sci. 2020, 77, 29. [Google Scholar] [CrossRef]
- Herms, D.A.; McCullough, D.G. Emerald Ash Borer Invasion of North America: History, Biology, Ecology, Impacts, and Management. Annu. Rev. Entomol. 2014, 59, 13–30. [Google Scholar] [CrossRef]
- Hultberg, T.; Sandström, J.; Felton, A.; Öhman, K.; Rönnberg, J.; Witzell, J.; Cleary, M. Ash Dieback Risks an Extinction Cascade. Biol. Conserv. 2020, 244, 108516. [Google Scholar] [CrossRef]
- Cipollini, D.; Rigsby, C.M.; Peterson, D.L. Feeding and Development of Emerald Ash Borer (Coleoptera: Buprestidae) on Cultivated Olive, Olea Europaea. J. Econ. Entomol. 2017, 110, 1935–1937. [Google Scholar] [CrossRef] [PubMed]
- Peterson, D.L.; Cipollini, D. Larval Performance of a Major Forest Pest on Novel Hosts and the Effect of Stressors. Environ. Entomol. 2020, 49, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Epanchin-Niell, R.S.; Haight, R.G.; Berec, L.; Kean, J.M.; Liebhold, A.M. Optimal Surveillance and Eradication of Invasive Species in Heterogeneous Landscapes. Ecol. Lett. 2012, 15, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Siegert, N.W.; McCullough, D.G.; Liebhold, A.M.; Telewski, F.W. Resurrected from the Ashes: A Historical Reconstruction of Emerald Ash Borer Dynamics through Dendrochronological Analysis. In Proceedings of the Emerald Ash Borer and Asian Longhorhed Beetle Research and Development Review Meeting; Cincinnatti, OH, USA, 29 October–2 November 2006, FHTET 2007-04; Mastro, V., Lance, D., Reardon, R., Parra, G., Eds.; US Forest Service, Forest Health Technology Enterprise Team: Morgantown, WV, USA, 2007; pp. 18–19. [Google Scholar]
- Epanchin-Niell, R.S.; Hastings, A. Controlling Established Invaders: Integrating Economics and Spread Dynamics to Determine Optimal Management. Ecol. Lett. 2010, 13, 528–541. [Google Scholar] [CrossRef]
- Kelnarova, I.; Jendek, E.; Grebennikov, V.V.; Bocak, L. First Molecular Phylogeny of Agrilus (Coleoptera: Buprestidae), the Largest Genus on Earth, with DNA Barcode Database for Forestry Pest Diagnostics. Bull. Entomol. Res. 2019, 109, 200–211. [Google Scholar] [CrossRef]
- Luchi, N.; Ioos, R.; Santini, A. Fast and Reliable Molecular Methods to Detect Fungal Pathogens in Woody Plants. Appl. Microbiol. Biotechnol. 2020, 104, 2453–2468. [Google Scholar] [CrossRef]
- Darling, J.A.; Blum, M.J. DNA-Based Methods for Monitoring Invasive Species: A Review and Prospectus. Biol. Invasions 2007, 9, 751–765. [Google Scholar] [CrossRef]
- Jerde, C.L.; Mahon, A.R.; Chadderton, W.L.; Lodge, D.M. “Sight-Unseen” Detection of Rare Aquatic Species Using Environmental DNA. Conserv. Lett. 2011, 4, 150–157. [Google Scholar] [CrossRef]
- Valentin, R.E.; Fonseca, D.M.; Nielsen, A.L.; Leskey, T.C.; Lockwood, J.L. Early Detection of Invasive Exotic Insect Infestations Using EDNA from Crop Surfaces. Front. Ecol. Environ. 2018, 16, 265–270. [Google Scholar] [CrossRef]
- Valentin, R.; Kyle, K.; Allen, M.; Welbourne, D.; Lockwood, J. The state, transport, and fate of aboveground terrestrial arthropod eDNA. Environ. DNA 2021, 3, 1081–1092. [Google Scholar] [CrossRef]
- Allen, M.C.; Nielsen, A.L.; Peterson, D.L.; Lockwood, J.L. Terrestrial EDNA Survey Outperforms Conventional Approach for Detecting an Invasive Pest Insect within an Agricultural Ecosystem. Environ. DNA 2021, 3, 1102–1112. [Google Scholar] [CrossRef]
- Peterson, D.L.; Allen, M.C.; Vastano, A.; Lockwood, J.L. Evaluation of Sample Collection and Storage Protocols for Surface EDNA Surveys of an Invasive Terrestrial Insect. Environ. DNA 2022, 4, 1201–1211. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-Mediated Isothermal Amplification of DNA. Nucleic Acids Res. 2000, 28, e63. [Google Scholar] [CrossRef]
- Kyei-Poku, G.; Gauthier, D.; Quan, G. Development of a Loop-Mediated Isothermal Amplification Assay as an Early-Warning Tool for Detecting Emerald Ash Borer (Coleoptera: Buprestidae) Incursions. J. Econ. Entomol. 2020, 113, 2480–2494. [Google Scholar] [CrossRef]
- Francese, J.A.; Rietz, M.L.; Mastro, V.C. Optimization of Multifunnel Traps for Emerald Ash Borer (Coleoptera: Buprestidae): Influence of Size, Trap Coating, and Color. J. Econ. Entomol. 2013, 106, 2415–2423. [Google Scholar] [CrossRef]
- Rassati, D.; Marini, L.; Marchioro, M.; Rapuzzi, P.; Magnani, G.; Poloni, R.; Di Giovanni, F.; Mayo, P.; Sweeney, J. Developing Trapping Protocols for Wood-Boring Beetles Associated with Broadleaf Trees. J. Pest Sci. 2019, 92, 267–279. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Büsse, S.; von Grumbkow, P.; Hummel, S.; Shah, D.N.; Tachamo Shah, R.D.; Li, J.; Zhang, X.; Yoshizawa, K.; Wedmann, S.; Hörnschemeyer, T. Phylogeographic Analysis Elucidates the Influence of the Ice Ages on the Disjunct Distribution of Relict Dragonflies in Asia. PLoS ONE 2012, 7, e38132. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Aglietti, C.; Luchi, N.; Pepori, A.L.; Bartolini, P.; Pecori, F.; Raio, A.; Capretti, P.; Santini, A. Real-Time Loop-Mediated Isothermal Amplification: An Early-Warning Tool for Quarantine Plant Pathogen Detection. AMB Express 2019, 9, 50. [Google Scholar] [CrossRef]
- Goldberg, C.S.; Turner, C.R.; Deiner, K.; Klymus, K.E.; Thomsen, P.F.; Murphy, M.A.; Spear, S.F.; McKee, A.; Oyler-McCance, S.J.; Cornman, R.S.; et al. Critical Considerations for the Application of Environmental DNA Methods to Detect Aquatic Species. Methods Ecol. Evol. 2016, 7, 1299–1307. [Google Scholar] [CrossRef]
- Rizzo, D.; Taddei, A.; Da Lio, D.; Bruscoli, T.; Cappellini, G.; Bartolini, L.; Salemi, C.; Luchi, N.; Pennacchio, F.; Rossi, E. Molecular Identification of Anoplophora Glabripennis (Coleoptera: Cerambycidae) from Frass by Loop-Mediated Isothermal Amplification. J. Econ. Entomol. 2020, 113, 2911–2919. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, D.; Luchi, N.; Da Lio, D.; Bartolini, L.; Nugnes, F.; Cappellini, G.; Bruscoli, T.; Salemi, C.; Griffo, R.V.; Garonna, A.P.; et al. Development of a Loop-Mediated Isothermal Amplification (LAMP) Assay for the Identification of the Invasive Wood Borer Aromia Bungii (Coleoptera: Cerambycidae) from Frass. 3 Biotech 2021, 11, 85. [Google Scholar] [CrossRef] [PubMed]
- Foo, P.C.; Nurul Najian, A.B.; Muhamad, N.A.; Ahamad, M.; Mohamed, M.; Yean Yean, C.; Lim, B.H. Loop-Mediated Isothermal Amplification (LAMP) Reaction as Viable PCR Substitute for Diagnostic Applications: A Comparative Analysis Study of LAMP, Conventional PCR, Nested PCR (NPCR) and Real-Time PCR (QPCR) Based on Entamoeba Histolytica DNA Derived from Faecal Sample. BMC Biotechnol. 2020, 20, 34. [Google Scholar] [CrossRef]
- Gunasegar, S.; Neela, V.K. Evaluation of Diagnostic Accuracy of Loop-Mediated Isothermal Amplification Method (LAMP) Compared with Polymerase Chain Reaction (PCR) for Leptospira Spp. in Clinical Samples: A Systematic Review and Meta-Analysis. Diagn. Microbiol. Infect. Dis. 2021, 100, 115369. [Google Scholar] [CrossRef]
- World Health Organization. The Use of Loop-Mediated Isothermal Amplification (TB-LAMP) for the Diagnosis of Pulmonary Tuberculosis: Policy Guidance; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Nixon, G.; Garson, J.A.; Grant, P.; Nastouli, E.; Foy, C.A.; Huggett, J.F. Comparative Study of Sensitivity, Linearity, and Resistance to Inhibition of Digital and Nondigital Polymerase Chain Reaction and Loop Mediated Isothermal Amplification Assays for Quantification of Human Cytomegalovirus. Anal. Chem. 2014, 86, 4387–4394. [Google Scholar] [CrossRef]
- Francois, P.; Tangomo, M.; Hibbs, J.; Bonetti, E.-J.; Boehme, C.C.; Notomi, T.; Perkins, M.D.; Schrenzel, J. Robustness of a Loop-Mediated Isothermal Amplification Reaction for Diagnostic Applications. FEMS Immunol. Med. Microbiol. 2011, 62, 41–48. [Google Scholar] [CrossRef]
- Siegert, N.W.; McCullough, D.G.; Liebhold, A.M.; Telewski, F.W. Dendrochronological Reconstruction of the Epicentre and Early Spread of Emerald Ash Borer in North America. Divers. Distrib. 2014, 20, 847–858. [Google Scholar] [CrossRef]


{kind=link}
| Insect Species | Label Name | Collection Date (mm-yr) | Insect Locality | Longitude Latitude | Extraction Kit | PCR Primers (A–F *) | EAB LAMP Detection |
|---|---|---|---|---|---|---|---|
| Buprestidae | |||||||
| 1. Agrilus angustulus | AA | 21 May | Vierzon, France | 2.0327° 47.2611° | Invitrogen | LCO1490-HCO2198 (B) | - |
| 2. Agrilus anxius | BBB | 14,15 June | CT, USA; ON, Canada | Qiagen | LCO1490-HCO2199 (A, D, E) | - | |
| 3. Agrilus ater | AAT | 2020 | Vierzon, France | 2.0327° 47.2611° | Qiagen | 28 s up–28 s low (F) | - |
| 4. Agrilus convexicollis | AC | 21 June | Orléans campus, France | 1.9383° 47.8456° | Qiagen | 16a–16b (F) | - |
| 5. Agrilus curtulus | Acu | 20 August | France | Qiagen | 28 s up–28 s low (F) | - | |
| 6. Agrilus graminis | AG | 20 August | Orléans campus, France | 1.9383° 47.8456° | Qiagen | LCO1490-HCO2199 (D) | - |
| 7. Agrilus hastulifer | AH | 21 June | Orléans campus, France | 1.9383° 47.8456° | Qiagen | 28 s up–28 s low (F) | - |
| 8. Agrilus laticornis | AL | 21 June | Orléans campus, France | 1.9383° 47.8456° | Qiagen | 28 s up–28 s low (F) | - |
| 9. Agrilus obscuricollis | AO | 21 June | Orléans campus, France | 1.9383° 47.8456° | Qiagen | 28 s up–28 s low (F) | - |
| 10. Agrilus olivicolor | AD | 21 June | Orléans campus, France | 1.9383° 47.8456° | Qiagen | LCO1490-HCO2198 (B) | - |
| 11. Agrilus planipennis | EAB | NY, RI, VT, USA | Qiagen | LCO1490-HCO2198 (A) EAB_COIF EAB_COIR (C) | + | ||
| 12. Agrilus roscidus | AR | 2020 | France | Qiagen | LCO1490-HCO2199 (E) | - | |
| 13. Agrilus sulcicollis | AS | 21 May | Vierzon, France | 2.0327° 47.2611° | Invitrogen | LCO1490-HCO2198 (A) | - |
| 14. Agrilus viridis | AV | 21 June | Orléans campus | 1.9383° 47.8456° | Qiagen | 28 s up–28 s low (F) | - |
| 15. Anthaxia nitidula | AN | 21 May | Marcenat | 3.3619° 46.2461° | Qiagen | LCO1490-HCO2199 (E) | - |
| 16. Chrysobothris affinis | CA | 20 June | Vierzon, France | 2.1544° 47.2899° | Qiagen | LCO1490-HCO2198 (E)28 s up–28 s low (F) | - |
| 17. Coraebus undatus | CU | 21 June | Orléans campus, France | 1.9383° 47.8456° | Qiagen | LCO1490-HCO2198 (B)28 s up–28 s low (F) | - |
| 18. Lamprodila mirifica | LM | Summer 2021 | Friuli Venezia Giulia Italy | 13.120926° 45.791784° | Qiagen | 28 s up-28 s low (F) | - |
| Curculionidae: Scolytinae | |||||||
| 19. Anisandrus dispar | A dis | Summer 2021 | Friuli Venezia Giulia, Italy | 13.120926° 45.791784° | Qiagen | LCO1490-HCO2198 (C) | - |
| 20. Xyleborinus saxesenii | XS | Summer 2021 | Friuli Venezia Giulia Italy | 13.120926° 45.791784° | Qiagen | LCO1490-HCO2199 (C) | - |
| Cerambycidae | |||||||
| 21. Aegomorphus clavipes | Aeg | Summer 2021 | Friuli Venezia Giulia, Italy | 13.120926° 45.791784° | Qiagen | 28 s up–28 s low (F) | - |
| 22. Exocentrus punctipennis | EP | Summer 2021 | Friuli Venezia Giulia, Italy | 13.120926° 45.791784° | Qiagen | LCO1490-HCO2199 (C) | - |
| 23. Leiopus nebulosus | LN | Summer 2021 | Friuli Venezia Giulia Italy | 13.120926° 45.791784° | Qiagen | LCO1490-HCO2199 (C) | - |
| 24. Saperda punctata | SP | Summer 2021 | Friuli Venezia Giulia Italy | 13.120926° 45.791784° | Qiagen | LCO1490-HCO2199 (C) | - |
| 25. Trichoferus pallidus | TN | 21 June | Orléans campus, France | 1.9383° 47.8456° | Qiagen | LCO1490-HCO2199 (E)28 s up–28 s low (F) | - |
| PCR Primer | Sequence (5′–3′) | Gene Type | Reference |
|---|---|---|---|
| EAB_COIF | AGG AAT AGT AGG AAC AGC CCT TAG A | COI | [36] |
| EAB_COIR | TAT TTC ATC TAA GGT AGG CAT CTG G | COI | |
| LCO1490 | GGT CAA CAA ATC ATA AAG ATA TTG G | COI | [39] |
| HCO2198 | TAA ACT TCA GGG TGA CCA AAA AAT CA | COI | |
| 28S_D1D2.3.A_UP | GGA ATC CGC TAA GGA GTG TGT AA | 28S | [40] |
| 28S_D1D2.3.A_LOW | AGG GCC TCG CTG GAG TAT TT | 28S | |
| RRNL MTDNA (16A) | CGC CTG TTT AAC AAA AAC AT | mtDNA | [41] |
| RRNL MTDNA (16B) | CCG GTC TGA ACT CAG ATC ATG T | mtDNA | |
| LAMP PRIMERS | |||
| EAB1_F3 | CTC CCT CCC TCT TTA ACA TTA C | COI | [36] |
| EAB1_B3 | GAT CAG ACT AGT AGA GGT GT | COI | |
| EAB1_FIP | ATA TTA GCC GCT AAT GGT GGG AAT AGT CGA AAG AGG AGC AG | COI | |
| EAB1_BIP | GGC TCT GTT GAC TTA GCA AAG GTT ATT CCT ATT GCT CGC | COI | |
| EAB1_LF | ATA TAC TGT CCA ACC AGT CC | COI | |
| EAB1_LB | CTG GAA TCT CCT CAA TTC TAG G | COI |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peterson, D.L.; Kyle, K.; Sallé, A.; Pecori, F.; Migliorini, D.; Santini, A.; Luchi, N.; Cleary, M. Specificity and Sensitivity of a Rapid LAMP Assay for Early Detection of Emerald Ash Borer (Agrilus planipennis) in Europe. Forests 2023, 14, 436. https://doi.org/10.3390/f14020436
Peterson DL, Kyle K, Sallé A, Pecori F, Migliorini D, Santini A, Luchi N, Cleary M. Specificity and Sensitivity of a Rapid LAMP Assay for Early Detection of Emerald Ash Borer (Agrilus planipennis) in Europe. Forests. 2023; 14(2):436. https://doi.org/10.3390/f14020436
Chicago/Turabian StylePeterson, Donnie L., Kathleen Kyle, Aurélien Sallé, Francesco Pecori, Duccio Migliorini, Alberto Santini, Nicola Luchi, and Michelle Cleary. 2023. "Specificity and Sensitivity of a Rapid LAMP Assay for Early Detection of Emerald Ash Borer (Agrilus planipennis) in Europe" Forests 14, no. 2: 436. https://doi.org/10.3390/f14020436
APA StylePeterson, D. L., Kyle, K., Sallé, A., Pecori, F., Migliorini, D., Santini, A., Luchi, N., & Cleary, M. (2023). Specificity and Sensitivity of a Rapid LAMP Assay for Early Detection of Emerald Ash Borer (Agrilus planipennis) in Europe. Forests, 14(2), 436. https://doi.org/10.3390/f14020436