
Climate Change-Driven Cumulative Mountain Pine Beetle-Caused Whitebark Pine Mortality in the Greater Yellowstone Ecosystem
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
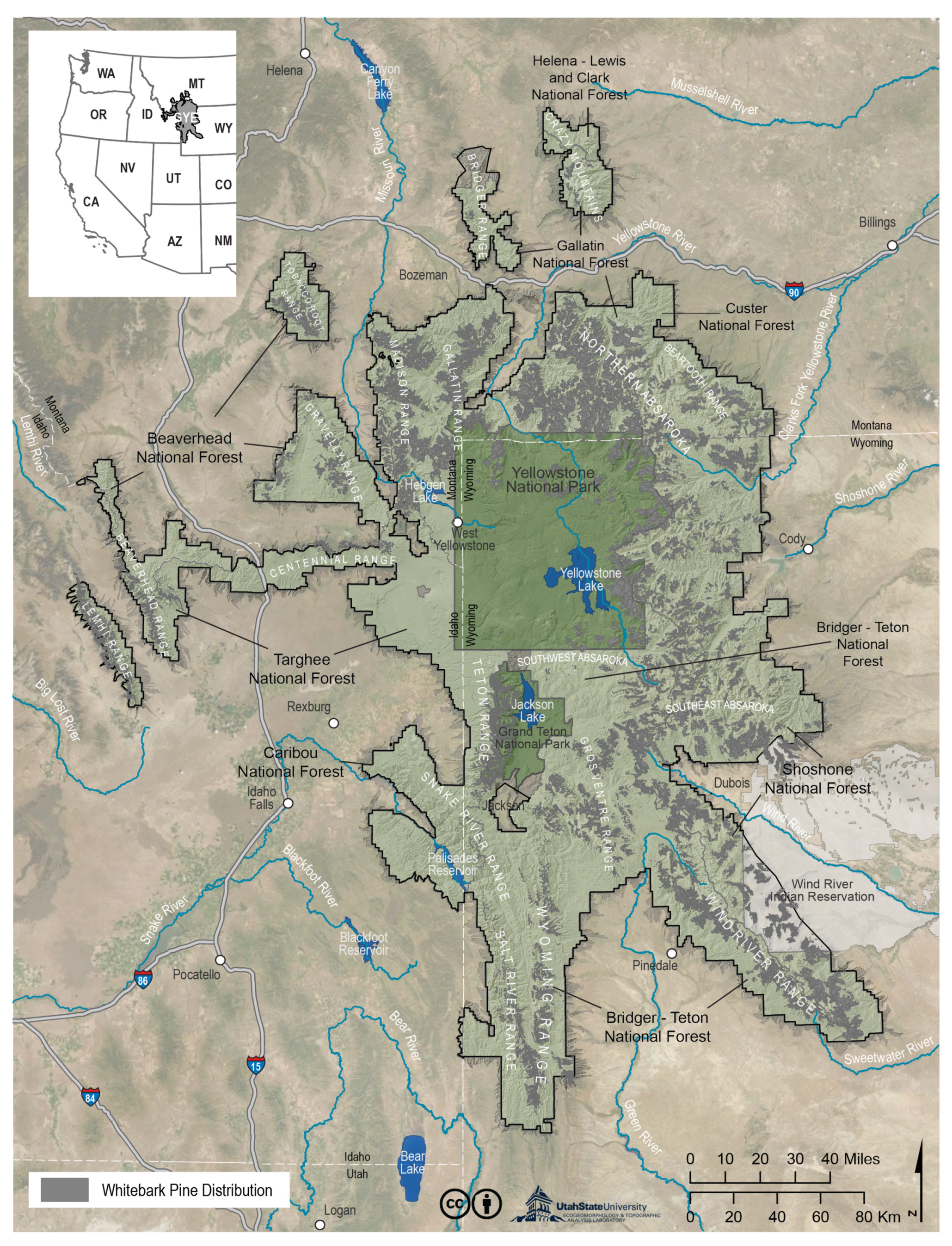
2.1. Study Area
2.2. Landscape Assessment System and Mountain Pine Beetle-Caused Mortality Rating System
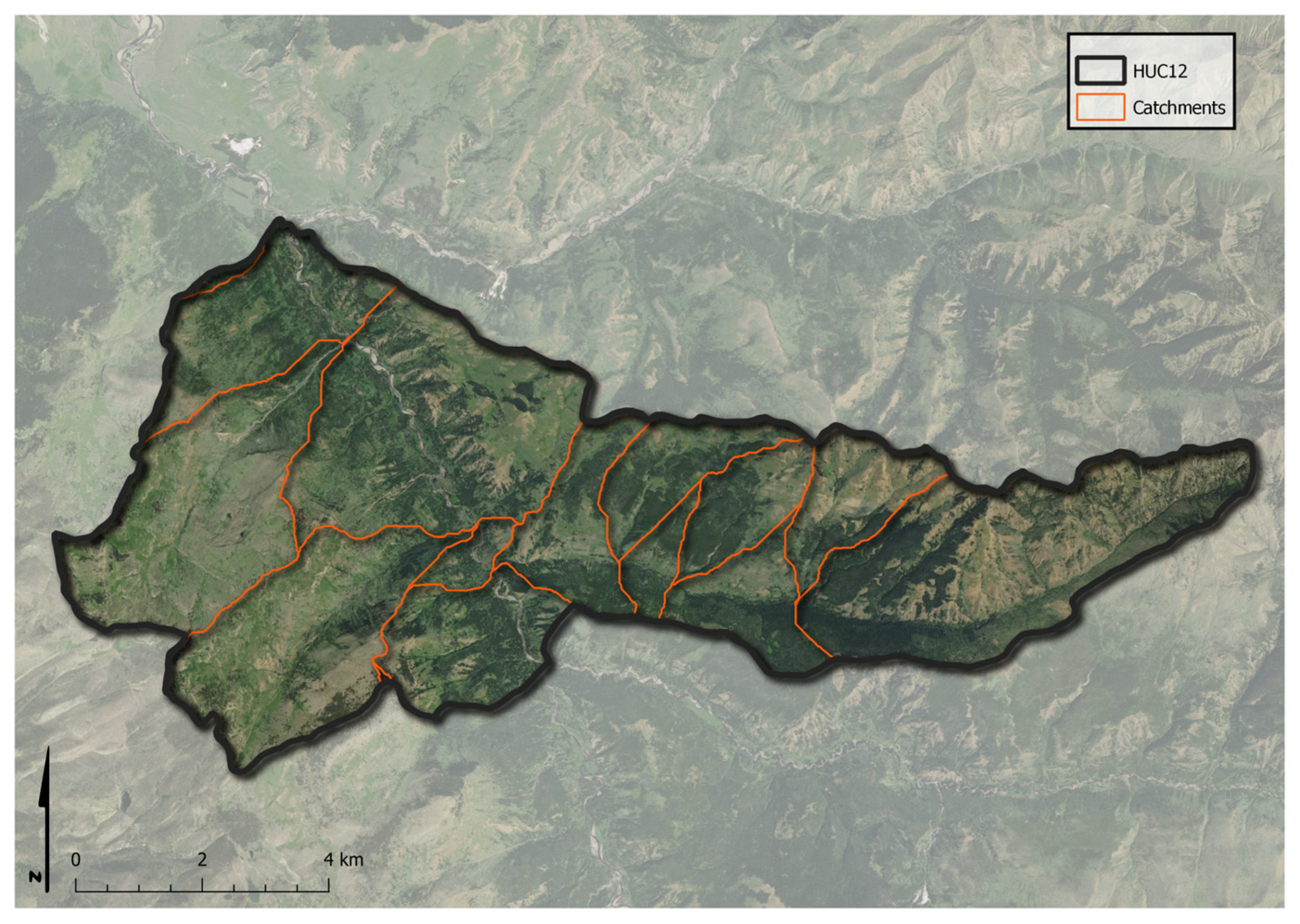
2.3. Spatial Scale of Landscape Assessment System
2.4. Landscape Assessment System Data Collection and Processing
2.4.1. Flightline Development
2.4.2. Over-Flights and Oblique Aerial Photo Capture
2.4.3. Image Processing
2.4.4. Photo Point Generation
2.4.5. Catchment-Level Photo Generation
2.4.6. Mortality Assessment and Mapping
2.5. Precision Assessment
2.6. Accuracy Assessment
3. Results
3.1. Oblique Aerial Photos
3.2. Point Level Mortality Data
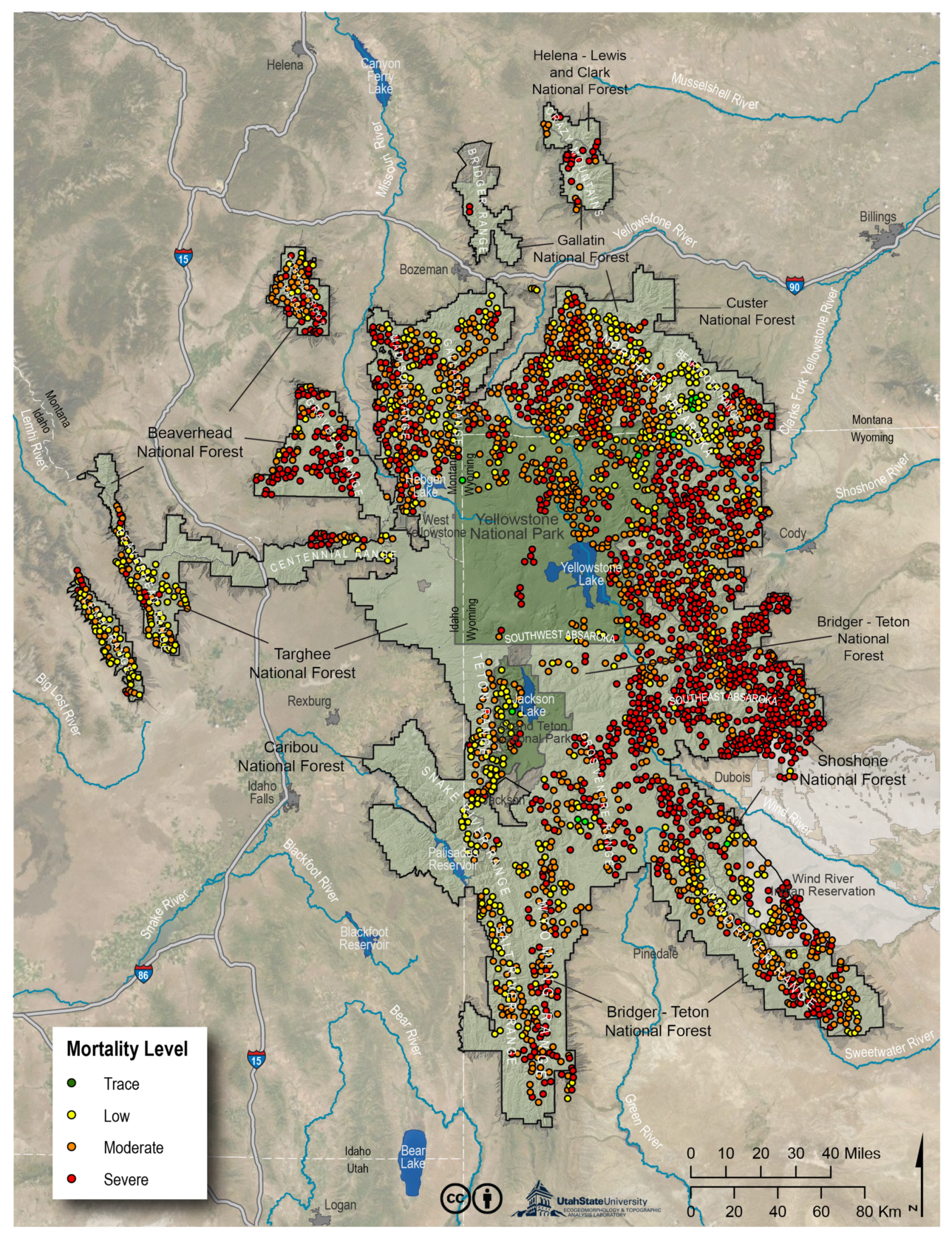
3.3. Catchment-Level Mortality Map
3.4. Precision Assessment
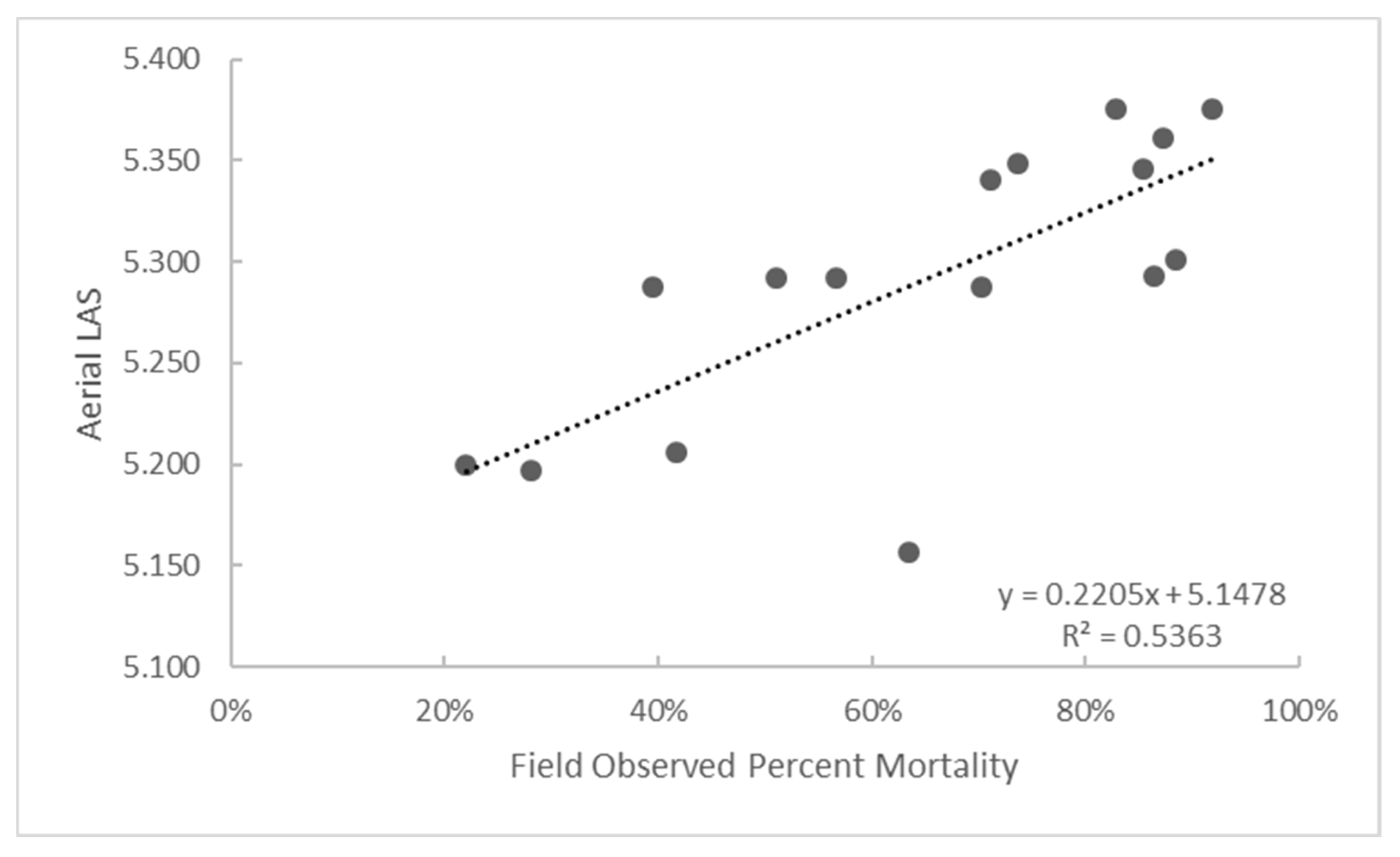
3.5. Accuracy Assessment
4. Discussion
4.1. Accuracy Assessment Limitations
4.2. Examining Cumulative Mortality over Time
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tomback, D.F.; Arno, S.F.; Keane, R.E. (Eds.) Whitebark Pine Communities: Ecology and Restoration, 1st ed.; Island Press: Washington, DC, USA, 2001; ISBN 978-1-55963-718-3. [Google Scholar]
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliott, K.; Ford, C.R.; Foster, D.R.; Kloeppel, B.D.; Knoepp, J.D.; Lovett, G.M.; et al. Loss of Foundation Species: Consequences for the Structure and Dynamics of Forested Ecosystems. Front. Ecol. Environ. 2005, 3, 479–486. [Google Scholar] [CrossRef]
- Resler, L.M.; Tomback, D.F. Blister Rust Prevalence in Krummholz Whitebark Pine: Implications for Treeline Dynamics, Northern Rocky Mountains, Montana, U.S.A. Arct. Antarct. Alp. Res. 2008, 40, 161–170. [Google Scholar] [CrossRef]
- Mattson, D.J. Causes and Consequences of Dietary Differences among Yellowstone Grizzly Bears (Ursus Arctos); University of Idaho: Moscow, ID, USA, 2000. [Google Scholar]
- Pease, C.M.; Mattson, D.J. Demography of the Yellowstone Grizzly Bears. Ecology 1999, 80, 957–975. [Google Scholar] [CrossRef]
- Tomback, D.F.; Linhart, Y.B. The Evolution of Bird-Dispersed Pines. Evol. Ecol. 1990, 4, 185–219. [Google Scholar] [CrossRef]
- Tomback, D.F.; Sund, S.K.; Hoffmann, L.A. Post-Fire Regeneration of Pinus albicaulis: Height–Age Relationships, Age Structure, and Microsite Characteristics. Can. J. For. Res. 1993, 23, 113–119. [Google Scholar] [CrossRef]
- Callaway, R.M. Competition and Facilitation on Elevation Gradients in Subalpine Forests of the Northern Rocky Mountains, USA. Oikos 1998, 82, 561–573. [Google Scholar] [CrossRef]
- Arno, S.F. Timberline: Mountain and Arctic Forest Frontiers, 1st ed.; Mountaineers Books: Seattle, WA, USA, 1984; ISBN 978-0-89886-085-6. [Google Scholar]
- Farnes, P. Snotel and Snow Course Data: Describing the Hydrology of Whitebark Pine Ecosystems (1990). In Proceedings of the Symposium on Whitebark Pine Ecosystems: Ecology and Management of a High Mountain Resource, Bozeman, MT, USA, 29–31 March 1989; pp. 302–304. [Google Scholar]
- Hann, W.J. Landscape and Ecosystem-Level Management in Whitebark Pine Ecosystems; U.S. Department of Agriculture Forest Service: Washington, DC, USA, 1990.
- Logan, J.A.; Macfarlane, W.W.; Willcox, L. Whitebark Pine Vulnerability to Climate-Driven Mountain Pine Beetle Disturbance in the Greater Yellowstone Ecosystem. Ecol. Appl. 2010, 20, 895–902. [Google Scholar] [CrossRef]
- Macfarlane, W.W.; Logan, J.A.; Kern, W.R. An Innovative Aerial Assessment of Greater Yellowstone Ecosystem Mountain Pine Beetle-Caused Whitebark Pine Mortality. Ecol. Appl. 2013, 23, 421–437. [Google Scholar] [CrossRef]
- Logan, J.A.; Macfarlane, W.W.; Willcox, L. Effective Monitoring as a Basis for Adaptive Management: A Case History of Mountain Pine Beetle in Greater Yellowstone Ecosystem Whitebark Pine. iForest Biogeosciences For. 2009, 2, 19. [Google Scholar] [CrossRef]
- Shanahan, E.; Irvine, K.M.; Thoma, D.; Wilmoth, S.; Ray, A.; Legg, K.; Shovic, H. Whitebark Pine Mortality Related to White Pine Blister Rust, Mountain Pine Beetle Outbreak, and Water Availability. Ecosphere 2016, 7, e01610. [Google Scholar] [CrossRef]
- Endangered and Threatened Wildlife and Plants; Threatened Species Status with Section 4(d) Rule for Whitebark Pine (Pinus Albicaulis). Available online: https://www.federalregister.gov/documents/2022/12/15/2022-27087/endangered-and-threatened-wildlife-and-plants-threatened-species-status-with-section-4d-rule-for (accessed on 21 November 2023).
- Safranyik, L. Effects of Climate and Weather on Mountain Pine Beetle Populations; Canadian Forest Service Publications: Sault Ste. Marie, ON, Canada, 1978; pp. 77–84. [Google Scholar]
- Amman, G.D. Population Changes of the Mountain Pine Beetle in Relation to Elevation. Environ. Entomol. 1973, 2, 541–548. [Google Scholar] [CrossRef]
- Jewett, J.T.; Lawrence, R.L.; Marshall, L.A.; Gessler, P.E.; Powell, S.L.; Savage, S.L. Spatiotemporal Relationships between Climate and Whitebark Pine Mortality in the Greater Yellowstone Ecosystem. For. Sci. 2011, 57, 320–335. [Google Scholar]
- Logan, J.A.; Amman, G.D. A Distribution Model for Egg Development in Mountain Pine Beetle. Can. Entomol. 1986, 118, 361–372. [Google Scholar] [CrossRef]
- Bentz, B.J.; Logan, J.A.; Amman, G.D. Temperature Dependent Development of the Mountain Pine Beetle (Coleoptera: Scolytidae) and Simulation of Its Phenology. Can. Entomol. 1991, 123, 1083–1094. [Google Scholar] [CrossRef]
- Régnière, J.; Cooke, B.J.; Bergeron, V. BioSIM: A Computer-Based Decision Support Tool for Seasonal Planning of Pest Management Activities; User’s Manual; Natural Resources Canada: Ottawa, ON, Canada, 1995; Volume 116, ISBN 978-0-662-24018-1.
- Logan, J.A.; Régnière, J.; Powell, J.A. Assessing the Impacts of Global Warming on Forest Pest Dynamics. Front. Ecol. Environ. 2003, 1, 130–137. [Google Scholar] [CrossRef]
- Logan, J.A.; Bentz, B.J. Model Analysis of Mountain Pine Beetle (Coleoptera: Scolytidae) Seasonality. Environ. Entomol. 1999, 28, 924–934. [Google Scholar] [CrossRef]
- Bentz, B.J.; Régnière, J.; Fettig, C.J.; Hansen, E.M.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.G.; Negrón, J.F.; Seybold, S.J. Climate Change and Bark Beetles of the Western United States and Canada: Direct and Indirect Effects. Bioscience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Perkins, D.L.; Swetnam, T.W. A Dendroecological Assessment of Whitebark Pine in the Sawtooth–Salmon River Region, Idaho. Can. J. For. Res. 1996, 26, 2123–2133. [Google Scholar] [CrossRef]
- Houghton, J.T. Climate Change: IPCC Scientific Assessment. Published for the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 1990; p. 365. [Google Scholar]
- Logan, J.A.; Powell, J.A. Ghost Forests, Global Warming, and the Mountain Pine Beetle (Coleoptera: Scolytidae). Am. Entomol. 2001, 47, 160–173. [Google Scholar] [CrossRef]
- Logan, J.; Powell, J. Ecological Consequences of Climate Change Altered Forest Insect Disturbance Regimes. In Climate Warming in Western North America: Evidence and Environmental Effects; Wagner, F.H., Ed.; University of Utah Press: Salt Lake City, UT, USA, 2009; pp. 98–109. [Google Scholar]
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and Earlier Spring Increase Western U.S. Forest Wildfire Activity. Science 2006, 313, 940–943. [Google Scholar] [CrossRef]
- Hostetler, S.; Whitlock, C.; Shuman, B.; Liefert, D.; Drimal, C.; Bischke, S. Greater Yellowstone Climate Assessment: Past, Present, and Future Climate Change in Greater Yellowstone Watersheds; Montana State University, Institute on Ecosystems: Bozeman, MT, USA, 2021; 260p. [Google Scholar] [CrossRef]
- Gibson, K. Mountain Pine Beetle Conditions in Whitebark Pine Stands in the Greater Yellowstone Ecosystem, 2006; Forest Health Protection Numbered Report 06-03; USDA Forest Service, Forest Health Protection Missoula Field Office: Missoula, MT, USA, 2006; pp. 1–7.
- Shanahan, E.K. SHORT: An Uncertain Future: The Persistence of Whitebark Pine in the Greater Yellowstone Ecosystem (U.S. National Park Service). Available online: https://www.nps.gov/articles/persistence-of-whitebark-pine-in-the-greater-yellowstone-ecosystem.htm (accessed on 2 May 2023).
- Powell, J.A.; Bentz, B.J. Connecting Phenological Predictions with Population Growth Rates for Mountain Pine Beetle, an Outbreak Insect. Landsc. Ecol. 2009, 24, 657–672. [Google Scholar] [CrossRef]
- Logan, J.A. Seeing (Less) Red: Bark Beetles and Global Warming. Ask an Ecologist (Blog); July 29, 2011; Ecotone: News and Views on Ecological Science. Available online: https://www.esa.org/esablog/2011/07/29/seeing-less-red-bark-beetles-and-global-warming/ (accessed on 1 October 2023).
- Dooley, E.M.; Six, D.L. Severe White Pine Blister Rust Infection in Whitebark Pine Alters Mountain Pine Beetle (Coleoptera: Curculionidae) Attack Density, Emergence Rate, and Body Size. Environ. Entomol. 2015, 44, 1384–1394. [Google Scholar] [CrossRef] [PubMed]
- Summary of Preliminary Step-Trend Analysis from the Interagency Whitebark Pine Long-term Monitoring Program—2004–2013|U.S. Geological Survey. Available online: https://www.usgs.gov/publications/summary-preliminary-step-trend-analysis-interagency-whitebark-pine-long-termmonitoring (accessed on 21 November 2023).
- Marston, R.A.; Anderson, J.E. Watersheds and Vegetation of the Greater Yellowstone Ecosystem. Conserv. Biol. 1991, 5, 338–346. [Google Scholar] [CrossRef]
- Romme, W.H.; Boyce, M.S.; Gresswell, R.; Merrill, E.H.; Minshall, G.W.; Whitlock, C.; Turner, M.G. Twenty Years After the 1988 Yellowstone Fires: Lessons About Disturbance and Ecosystems. Ecosystems 2011, 14, 1196–1215. [Google Scholar] [CrossRef]
- Bockino, N. and Macfarlane, W. Whitebark pine distribution and condition assessment for the Greater Yellowstone (Map). Cooperation with USDA Forest Service—Forest Health and Protection and Grand Teton National Park. In Whitebark Pine Strategy for the Greater Yellowstone Area; Greater Yellowstone Coordinating Committee: Bozeman, MT, USA, 2010; 41p. [Google Scholar]
- Macfarlane, W.W.; Wheaton, J.M.; Bouwes, N.; Jensen, M.L.; Gilbert, J.T.; Hough-Snee, N.; Shivik, J.A. Modeling the Capacity of Riverscapes to Support Beaver Dams. Geomorphology 2017, 277, 72–99. [Google Scholar] [CrossRef]
- Macfarlane, W.W.; McGinty, C.M.; Laub, B.G.; Gifford, S.J. High-Resolution Riparian Vegetation Mapping to Prioritize Conservation and Restoration in an Impaired Desert River. Restor. Ecol. 2017, 25, 333–341. [Google Scholar] [CrossRef]
- Ireland, K.B.; Hansen, A.J.; Keane, R.E.; Legg, K.; Gump, R.L. Putting Climate Adaptation on the Map: Developing Spatial Management Strategies for Whitebark Pine in the Greater Yellowstone Ecosystem. Environ. Manag. 2018, 61, 981–1001. [Google Scholar] [CrossRef]
- Congalton, R.G. Accuracy Assessment and Validation of Remotely Sensed and Other Spatial Information. Int. J. Wildland Fire 2001, 10, 321. [Google Scholar] [CrossRef]
- Turiano, T. Select Peaks of Greater Yellowstone: A Mountaineering History and Guide; Indomitus Books: Jackson, WY, USA, 2003; ISBN 978-0-9745619-0-5. [Google Scholar]
- Locke, W.W. Present Climate and Glaciation of Western Montana, USA. Arct. Alp. Res. 1989, 21, 234–244. [Google Scholar] [CrossRef]
- Landis, J.R.; Koch, G.G. An Application of Hierarchical Kappa-Type Statistics in the Assessment of Majority Agreement among Multiple Observers. Biometrics 1977, 33, 363–374. [Google Scholar] [CrossRef]
- Kunegel-Lion, M.; Lewis, M.A. Factors Governing Outbreak Dynamics in a Forest Intensively Managed for Mountain Pine Beetle. Sci. Rep. 2020, 10, 7601. [Google Scholar] [CrossRef]
- Buotte, P.C.; Hicke, J.A.; Preisler, H.K.; Abatzoglou, J.T.; Raffa, K.F.; Logan, J.A. Climate Influences on Whitebark Pine Mortality from Mountain Pine Beetle in the Greater Yellowstone Ecosystem. Ecol. Appl. 2016, 26, 2507–2524. [Google Scholar] [CrossRef]
- Dooley, E.M.; Six, D.L.; Powell, J.A. A Comparison of Mountain Pine Beetle (Coleoptera: Curculionidae, Scolytinae) Productivity and Survival in Lodgepole and Whitebark Pine after a Region-Wide Cold Weather Event. For. Sci. 2015, 61, 235–246. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Numeric Rating | Mortality Level | Description |
|---|---|---|
| 0–0.75 | Trace | Green forest with trace levels of mortality. “Trace” mortality refers to a catchment that contains an occasional gray or red tree and there is no evidence of mortality expanding to neighboring trees. |
| 1–1.75 | Low | Green forest with occasional spots of gray or red trees across the catchment. The increasing frequency of current year red spots is assessed with a 1.25, 1.5, and 1.75 rating. Red or gray spots do not show evidence of multi-year mountain pine beetle-caused mortality. |
| 2–2.75 | Moderate | Primarily green forest with multiple spots of red and/or gray trees across the catchment. Spots show evidence of two or more years of subsequent mortality. The increasing magnitude of these spots is assessed with a 2.25, 2.5 and 2.75 rating. |
| 3–4 | Severe | Primarily red forest where spots of red and gray trees have coalesced across the catchment. A catchment may display a varying degree of coalescing spots ranging from initial coalescence, category 3, to increasing coalescence that is assessed with a 3.25, 3.5, 3.75 and 4 rating where essentially the entire whitebark pine overstory is red. |
| 5.1–5.4 | Old attack | Old attack mortality where a catchment may contain a varying degree of dead (gray) trees and remaining green forest after a mountain pine beetle attack. This variation is captured with a 5.0–5.4 rating based on the amount of dead and gray whitebark pine overstory. These mortality intensity values are equivalent to those found in the 0–4 categories (i.e., a 5.35 rating is equal to a 3.5 at the end of the outbreak cycle). Rating of 5.x pertains to an old attack (gray trees) as opposed to a 1–4 that indicates active attack (red trees). |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Macfarlane, W.W.; Howell, B.; Logan, J.A.; Smith, A.L.; Rasmussen, C.C.; Spangler, R.E. Climate Change-Driven Cumulative Mountain Pine Beetle-Caused Whitebark Pine Mortality in the Greater Yellowstone Ecosystem. Forests 2023, 14, 2361. https://doi.org/10.3390/f14122361
Macfarlane WW, Howell B, Logan JA, Smith AL, Rasmussen CC, Spangler RE. Climate Change-Driven Cumulative Mountain Pine Beetle-Caused Whitebark Pine Mortality in the Greater Yellowstone Ecosystem. Forests. 2023; 14(12):2361. https://doi.org/10.3390/f14122361
Chicago/Turabian StyleMacfarlane, William W., Brian Howell, Jesse A. Logan, Ally L. Smith, Cashe C. Rasmussen, and Robert E. Spangler. 2023. "Climate Change-Driven Cumulative Mountain Pine Beetle-Caused Whitebark Pine Mortality in the Greater Yellowstone Ecosystem" Forests 14, no. 12: 2361. https://doi.org/10.3390/f14122361
APA StyleMacfarlane, W. W., Howell, B., Logan, J. A., Smith, A. L., Rasmussen, C. C., & Spangler, R. E. (2023). Climate Change-Driven Cumulative Mountain Pine Beetle-Caused Whitebark Pine Mortality in the Greater Yellowstone Ecosystem. Forests, 14(12), 2361. https://doi.org/10.3390/f14122361