
The Effects of Frost Cracks and Large Poplar Borer Damage on Stem Rot in Hybrid Aspen (Populus tremula L. × Populus tremuloides Michx.) Clones




Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Data Analysis
3. Results
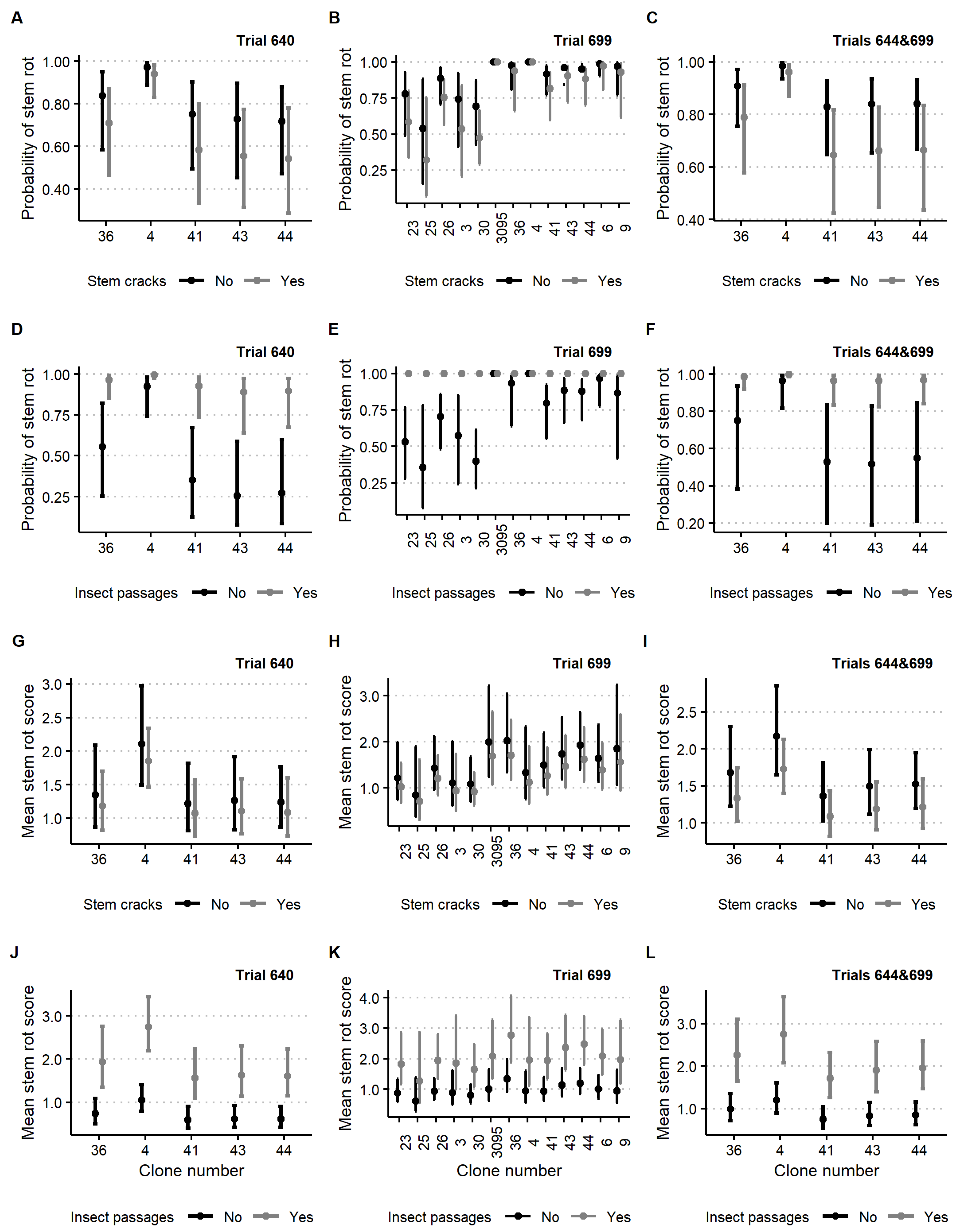
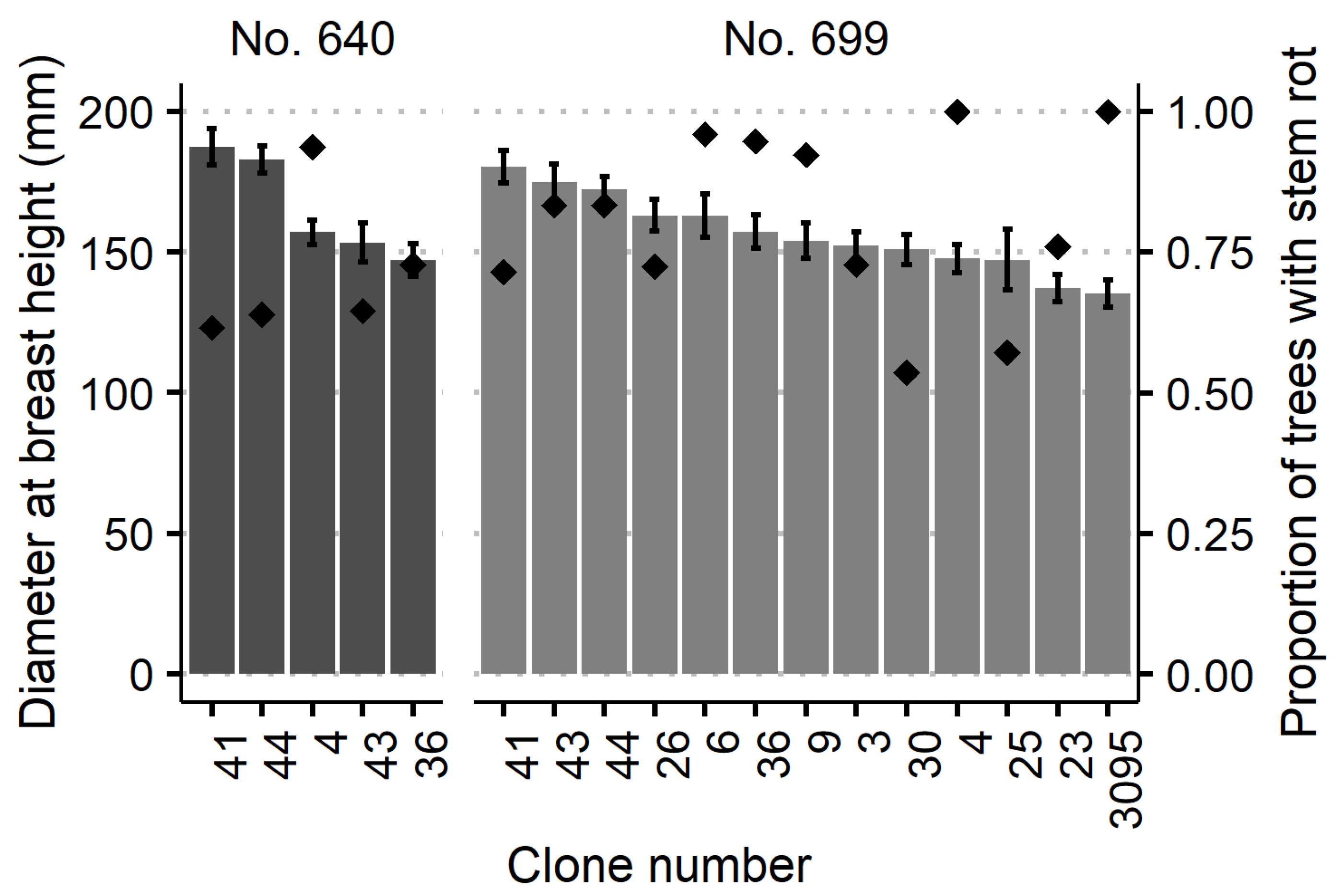
3.1. Stem Rot Impacting Factors
3.2. Genetic Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zeps, M. Apšu Hibrīdu (Populus tremuloides Michx. × Populus tremula L.) Audzēšanas Potenciāls Latvijā. [Potential of Hybrid Aspen (Populus tremuloides Michx. × Populus tremula L.) Production in Latvia]; Latvia University of Life Sciences and Technologies: Jelgava, Latvia, 2017. [Google Scholar]
- Lee, D.; Beuker, E.; Viherä-Aarnio, A.; Hynynen, J. Site index models with density effect for hybrid aspen (Populus tremula L. × P. tremuloides Michx.) plantations in southern Finland. For. Ecol. Manag. 2021, 480, 118669. [Google Scholar] [CrossRef]
- Tullus, A.; Lukason, O.; Vares, A.; Padari, A.; Lutter, R.; Tullus, T.; Karoles, K.; Tullus, H. Economics of Hybrid Aspen (Populus tremula L. × P. tremuloides Michx.) and Silver Birch (Betula pendula Roth.) Plantations on Abandoned Agricultural Lands in Estonia. Balt. For. 2012, 18, 288–298. [Google Scholar]
- Smilga, J.; Zeps, M.; Sisenis, L.; Kalnins, J.; Adamovics, A.; Donis, J. Profitability of hybrid aspen breeding in Latvia. Agron. Res. 2015, 13, 430–435. [Google Scholar]
- Miezīte, O. Meža Aizsardzība. Meža Fitopatoloģija [Forest Protection. Forest Phytopathology], 2nd ed.; Studentu Biedrība «Šalkone»: Jelgava, Latvia, 2017; ISBN 9789934859960. [Google Scholar]
- Christersson, L. Future research on hybrid aspen and hybrid poplar cultivation in Sweden. Biomass Bioenergy 1996, 11, 109–113. [Google Scholar] [CrossRef]
- Nicolai, V. The bark of trees: Thermal properties, microclimate and fauna. Oecologia 1986, 69, 148–160. [Google Scholar] [CrossRef]
- Tullus, A.; Rytter, L.; Tullus, T.; Weih, M.; Tullus, H. Short-rotation forestry with hybrid aspen (Populus tremula L. × P. tremuloides Michx.) in Northern Europe. Scand. J. For. Res. 2012, 27, 10–29. [Google Scholar] [CrossRef]
- Jones, A.C.; Ostry, M.E.; Service, U.F.; Central, N.; Station, F.E.; Ave, F.; Paul, S. Estimating White Trunk Rot in Aspen Stands. North. J. Appl. For. 1998, 15, 33–36. [Google Scholar] [CrossRef]
- Jactel, H.; Vodde, F. Prevalence of Biotic and Abiotic Hazards in European Forests; European Forest Institute: Joensuu, Finland, 2011. [Google Scholar]
- Balla, A.; Silini, A.; Cherif-Silini, H.; Bouket, A.C.; Moser, W.K.; Nowakowska, J.A.; Oszako, T.; Benia, F.; Belbahri, L. The threat of pests and pathogens and the potential for biological control in forest ecosystems. Forests 2021, 12, 1579. [Google Scholar] [CrossRef]
- Silva, L.N.; Freer-Smith, P.; Madsen, P. Production, restoration, mitigation: A new generation of plantations. New For. 2019, 50, 153–168. [Google Scholar] [CrossRef]
- Šēnhofa, S.; Zeps, M.; Gailis, A.; Kāpostiņš, R.; Jansons, Ā. Development of stem cracks in young hybrid aspen plantations. For. Stud. 2016, 65, 16–23. [Google Scholar] [CrossRef]
- Zeps, M.; Senhofa, S.; Zadina, M.; Neimane, U.; Jansons, A. Stem damages caused by heart rot and large poplar borer on hybrid and European aspen. For. Stud. 2017, 66, 21–26. [Google Scholar] [CrossRef][Green Version]
- Christersson, L. Wood production potential in poplar plantations in Sweden. Biomass Bioenergy 2010, 34, 1289–1299. [Google Scholar] [CrossRef]
- Shortle, W.C.; Dudzik, K.R. Wood Decay in Living and Dead Trees: A Pictorial Overview; US Department of Agriculture, Forest Service, Northeasern Research Station: Newtown Square, PN, USA, 2012. [Google Scholar]
- Falconer, D.S.; Mackay, T.F. Introduction to Quantitative Genetics, 4th ed.; Longman Group Ltd.: London, UK, 1996. [Google Scholar]
- R Core Development Team. A Language and Environment for Statistical Computing. R Found. R Found. Stat. Comput. 2020, 2. Available online: https://www.R-project.org (accessed on 2 March 2022).
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. Emmeans: Estimated marginal means, aka least-squares means. R Packag. Vers. 2022, 1, 3. [Google Scholar]
- Hadfield, J.D. MCMC methods for multi-response generalized linear mixed models: The MCMCglmm R package. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar] [CrossRef]
- Hagman, M. Kokemuksia hybridihaavasta [Experience with hybrid aspen]. Sorbifolia 1997, 28, 51–59. [Google Scholar]
- Jackson, J.A.; Jackson, B.J.S. Ecological Relationships between Fungi and Woodpecker Cavity Sites. Condor 2004, 106, 37–49. [Google Scholar] [CrossRef]
- Domański, S. Fungi that destroyed a Populus tremula stand in Lagow Lubuski. Eur. J. For. Pathol. 1983, 13, 166–173. [Google Scholar] [CrossRef]
- Woodward, S.; Stenlid, J.; Riikka, K.; Hüttermann, A. Heterobasidion Annosum: Biology, Ecology, Impact and Control, 1st ed.; Oxford University Press: Wallingford, UK, 1998; ISBN 0851992757. [Google Scholar]
- Brown, J.T.S.; Merill, W. Germination of Fomes igniarius var. populinus basidiospores on Aspen pith. Aspen Bibliogr. 1971, 61, 5549. [Google Scholar]
- Holmer, L.; Nitare, L.; Stenlid, J. Population structure and decay pattern of Phellinus tremulae in Populus tremula as determined by somatic incompatibility. Can. J. Bot. 1994, 72, 1391–1396. [Google Scholar] [CrossRef]
- Etheridge, D.E. Factors affecting branch infection in aspen. Can. J. Bot. 1961, 39, 799–816. [Google Scholar] [CrossRef]
- Debyle, N.V.; Winokur, R.P. Aspen: Ecology and Management in the Western United States; USDA Forest Service: Fort Collins, CO, USA, 1985; Volume 119. [Google Scholar]
- Hajek, A.E.; van Frankenhuyzen, K. Use of Entomopathogens Against Forest Pests. In Microbial Control of Insect and Mite Pests: From Theory to Practice; Academic Press: Amsterdam, The Netherlands, 2017; pp. 313–330. ISBN 9780128035665. [Google Scholar]
- Linnakoski, R.; Forbes, K.M. Pathogens—The hidden face of forest invasions by wood-boring insect pests. Front. Plant Sci. 2019, 10, 90. [Google Scholar] [CrossRef] [PubMed]
- Kasanen, R.; Hantula, J.; Kurkela, T.; Vuorinen, M.; Komulainen, A.; Haapala, J.; Penttinen, H.; Beuker, E. Resistance in hybrid aspen to pathogens. In Forest Pathology Research in the Nordic and Baltic Countries 2005, Proceedings of the SNS-Meeting in Forest Pathology at Skogbrukets Kursinstitutt, Gjøvik, Norway, 28–31 August 2005; Aktuelt Fra Skogforskningen: As, Norway, 2006; pp. 20–22. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Trial | 640 | 699 | ||
|---|---|---|---|---|
| Age (years) | 13 | 12 | ||
| Number of replications | 6 | 4 | ||
| Plot design | 25-tree block-plots | 16-tree block-plots | ||
| Number of analyzed clones | 5 | 13 | ||
| Number of trees | 457 | 677 | ||
| Number of harvested/remaining trees | 202 | 255 | 265 | 412 |
| Number of harvested trees with stem rot | 149 | 207 | ||
| Number of harvested trees with insect passages | 95 | 129 | ||
| Mean height ± standard deviation of harvested/remaining trees (m) | 20.1 ± 2.3 | 20.2 ± 9.0 | 16.2 ± 8.1 | 15.5 ± 3.0 |
| Mean diameter at breast height ± standard deviation of harvested/remaining trees (mm) | 159.0 ± 30.7 | 159.0 ± 33.7 | 153.5 ± 29.1 | 155.5 ± 24.7 |
| Proportion of harvested/remaining trees with stem cracks (%) | 70.0 | 70.5 | 62.5 | 62.9 |
| Trait | Broad-Sense Heritability H2 ± Standard Error | Type-B Genotypic Correlation rG ± Standard Error |
|---|---|---|
| Height | 0.00 ± 0.11 | −0.03 ± 0.55 |
| Diameter at breast height | 0.17 ± 0.02 | 0.57 ± 0.69 |
| Stem rot | 0.21 ± 0.18 | 0.86 ± 0.47 |
| Stem cracks | 0.04 ± 0.06 | 0.02 ± 0.86 |
| Insect passages | 0.00 ± 0.00 | −0.02 ± 0.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čakšs, R.; Zeltiņš, P.; Čakša, L.; Zeps, M.; Jansons, Ā. The Effects of Frost Cracks and Large Poplar Borer Damage on Stem Rot in Hybrid Aspen (Populus tremula L. × Populus tremuloides Michx.) Clones. Forests 2022, 13, 593. https://doi.org/10.3390/f13040593
Čakšs R, Zeltiņš P, Čakša L, Zeps M, Jansons Ā. The Effects of Frost Cracks and Large Poplar Borer Damage on Stem Rot in Hybrid Aspen (Populus tremula L. × Populus tremuloides Michx.) Clones. Forests. 2022; 13(4):593. https://doi.org/10.3390/f13040593
Chicago/Turabian StyleČakšs, Roberts, Pauls Zeltiņš, Linda Čakša, Mārtiņš Zeps, and Āris Jansons. 2022. "The Effects of Frost Cracks and Large Poplar Borer Damage on Stem Rot in Hybrid Aspen (Populus tremula L. × Populus tremuloides Michx.) Clones" Forests 13, no. 4: 593. https://doi.org/10.3390/f13040593
APA StyleČakšs, R., Zeltiņš, P., Čakša, L., Zeps, M., & Jansons, Ā. (2022). The Effects of Frost Cracks and Large Poplar Borer Damage on Stem Rot in Hybrid Aspen (Populus tremula L. × Populus tremuloides Michx.) Clones. Forests, 13(4), 593. https://doi.org/10.3390/f13040593