
MaxEnt Modeling to Estimate the Impact of Climate Factors on Distribution of Pinus densiflora

Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Climatic Data
2.3. Model Simulation
2.4. Geospatial Analysis
3. Results
3.1. Evaluation of MaxEnt Model Prediction Accuracy
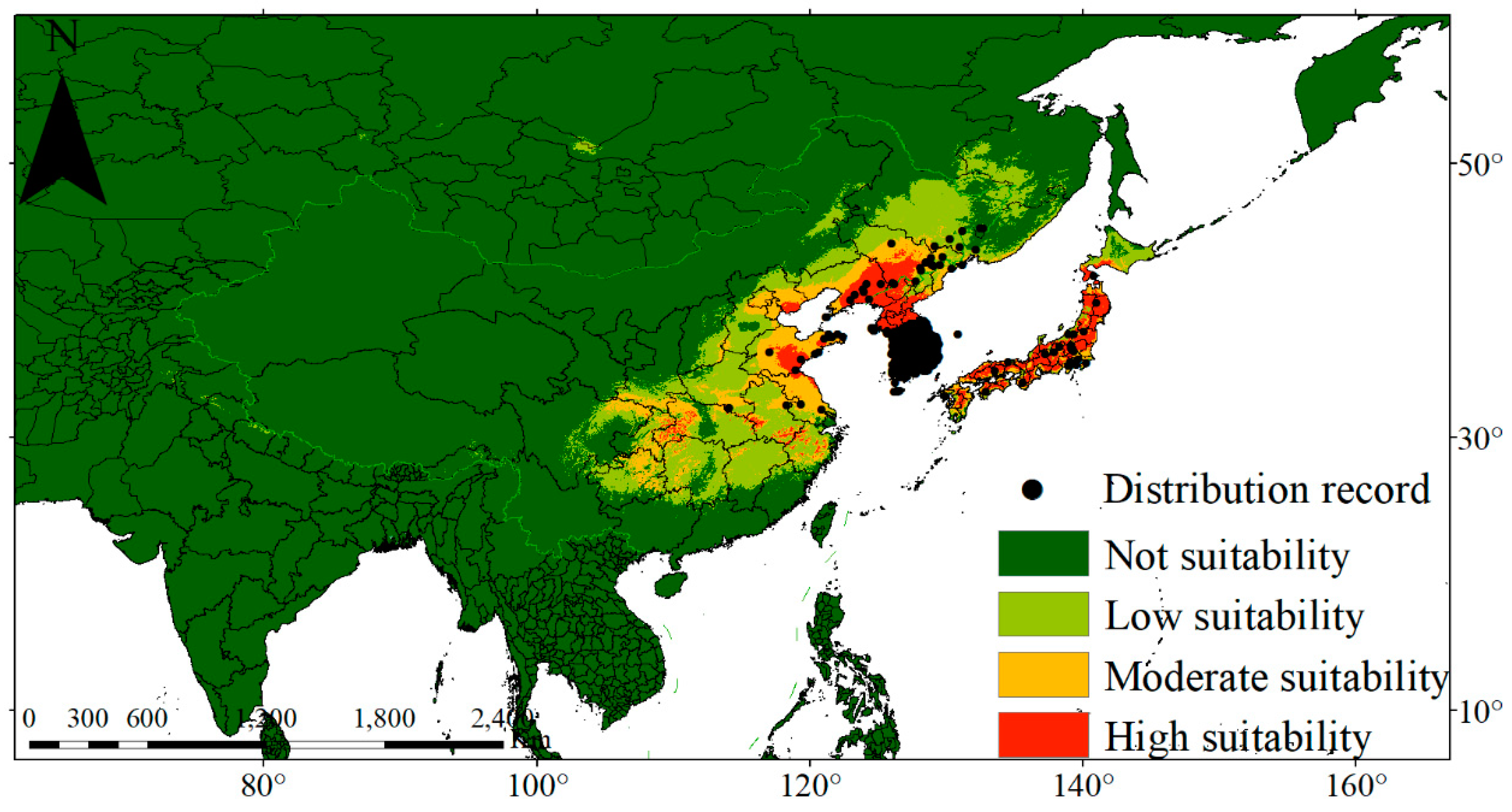
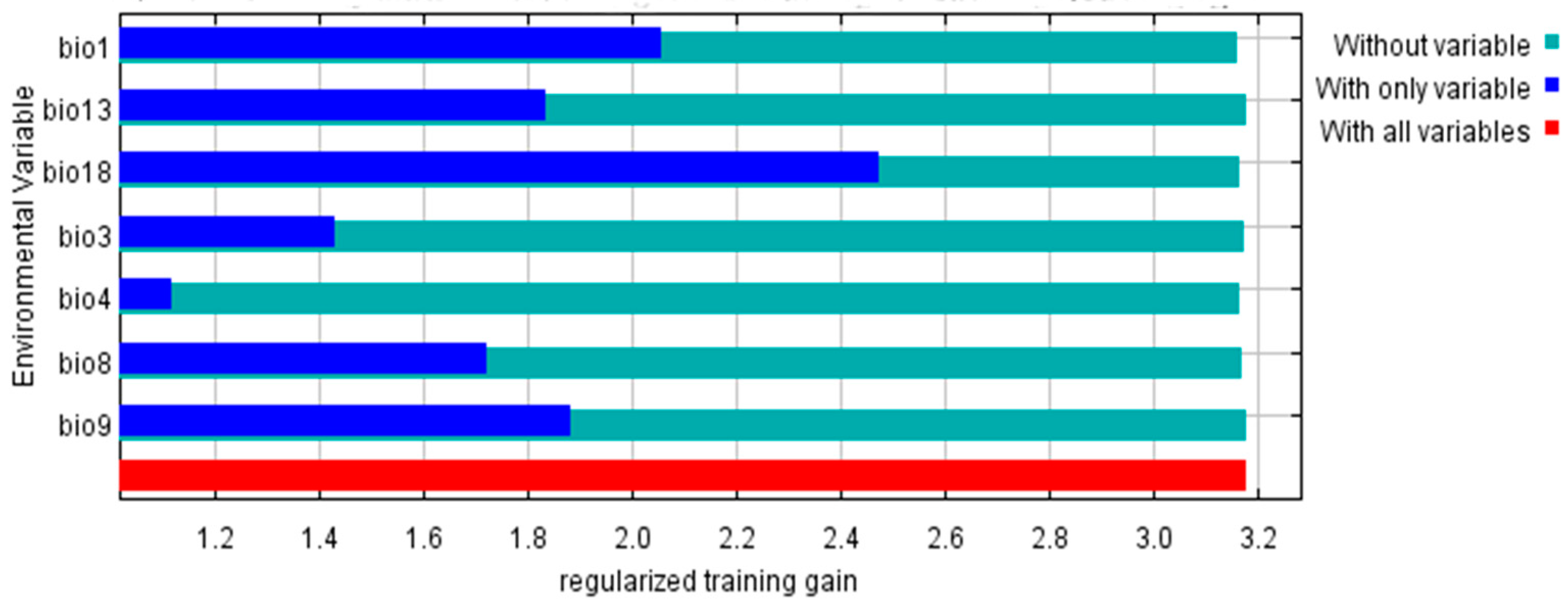
3.2. Climate-Dominant Factors and the Potential Distribution Areas of P. densiflora
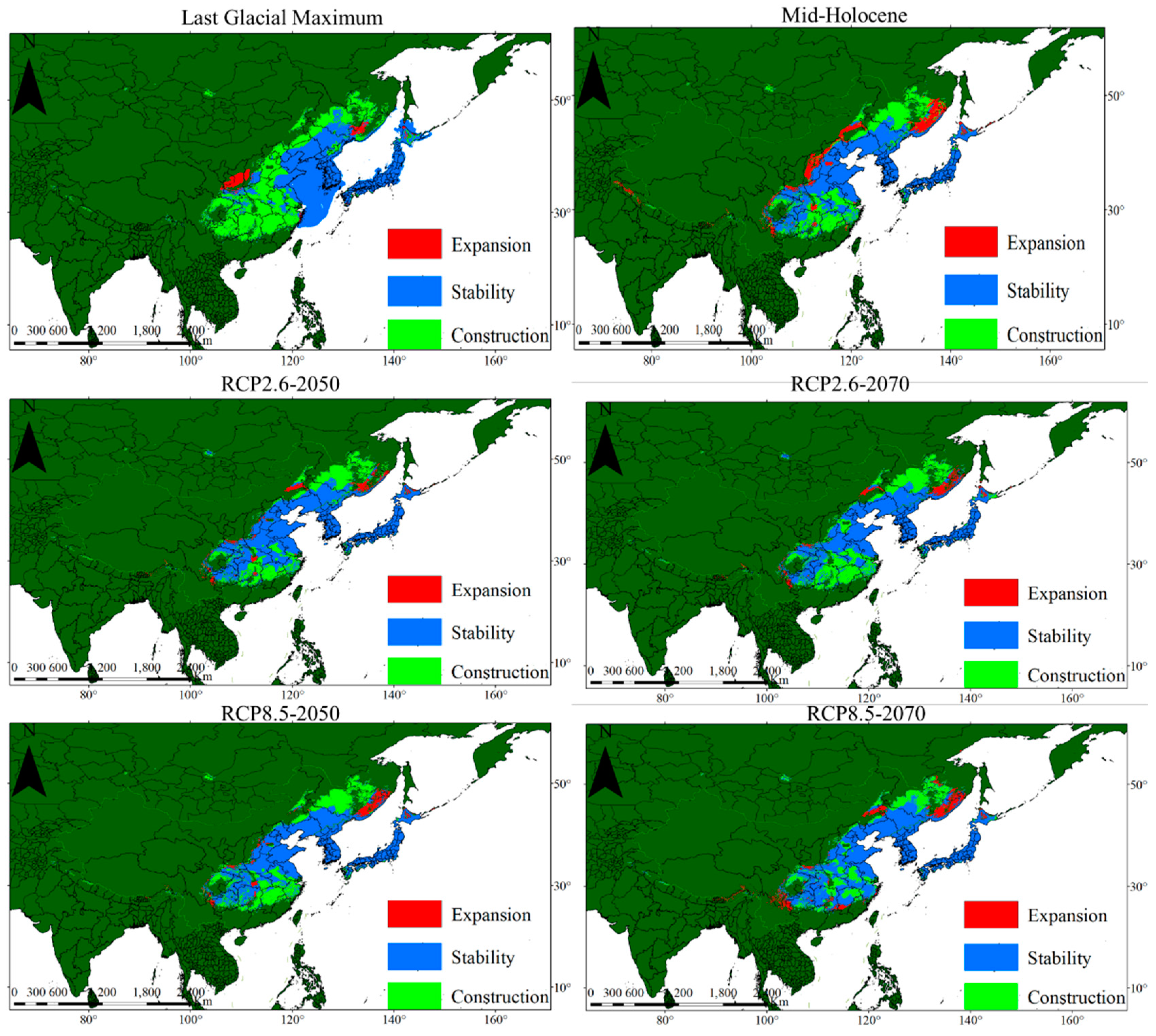
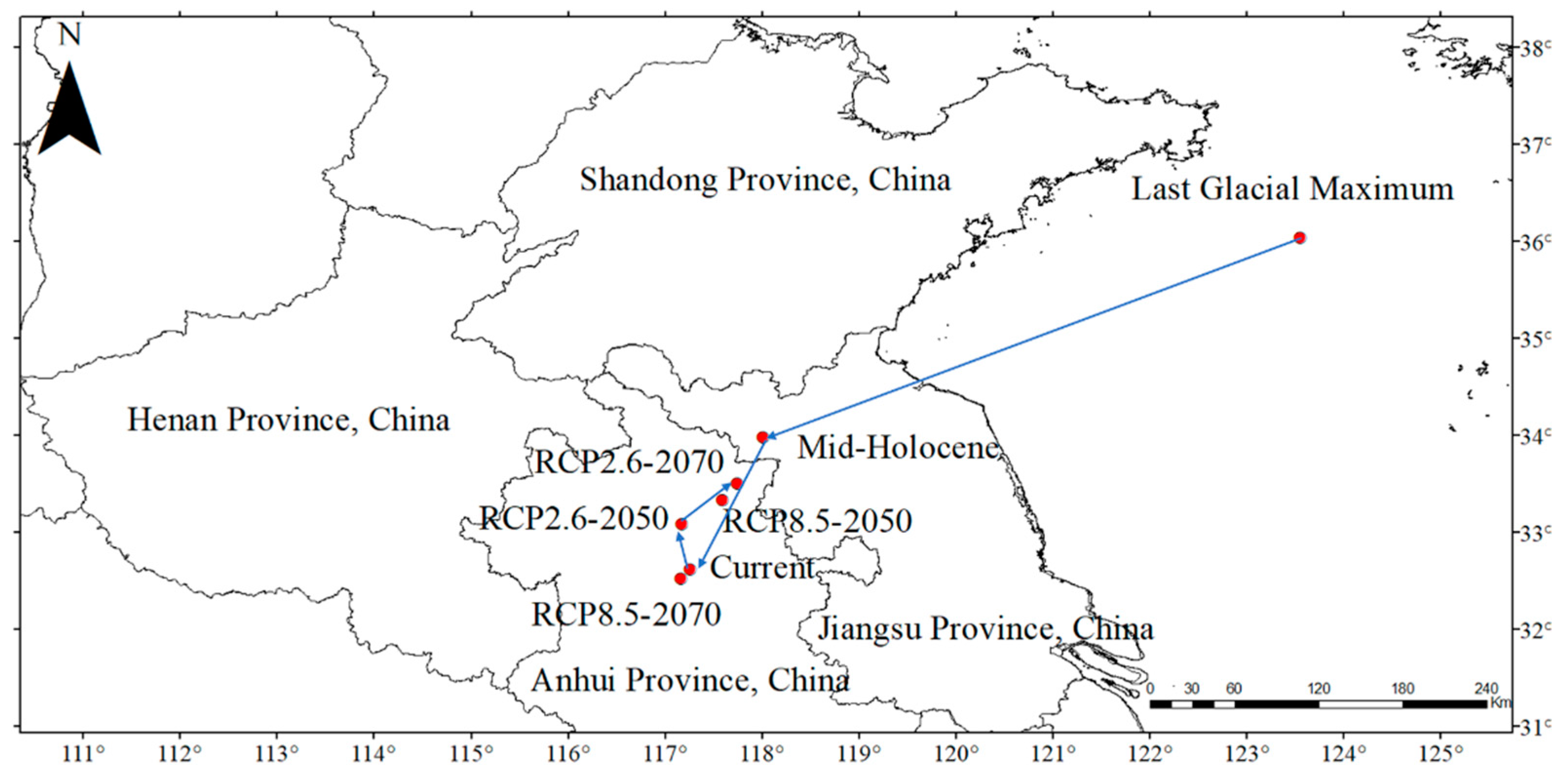
3.3. Potential Distribution and Changes of P. densiflora
4. Discussion
4.1. Evaluation of the MaxEnt Model
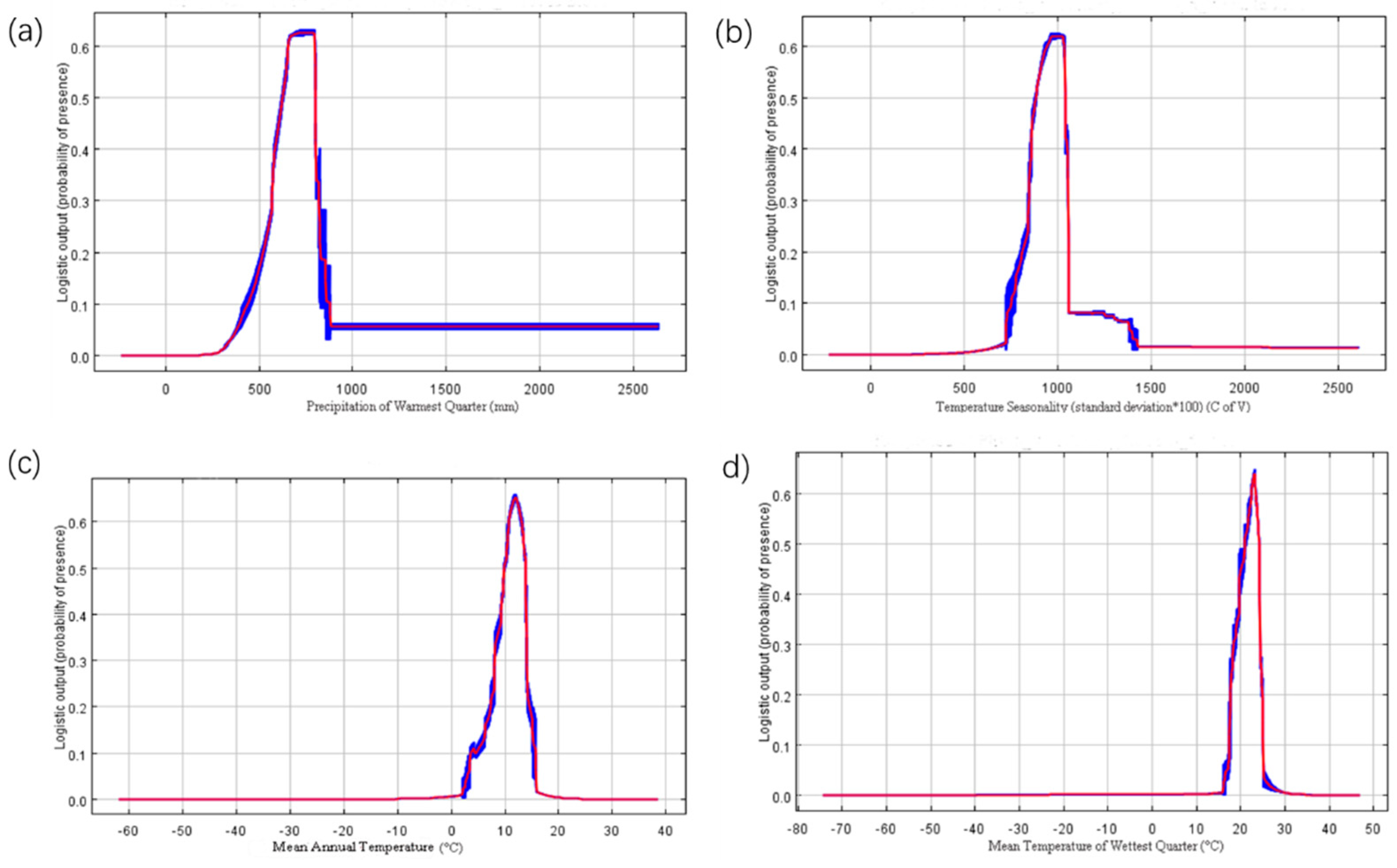
4.2. Habitat Suitability Response to Environmental Variables
4.3. Geographical Distribution and Change of P. densiflora
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hewitt, G.M. Genetic consequences of climatic oscillations in the Quaternary. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2004, 359, 183–195; discussion 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, J.; Yu, G.; Harrison, S.P.; Prentice, I.C. Palaeovegetation in China during the late Quaternary: Biome reconstructions based on a global scheme of plant functional types. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 289, 44–61. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Ni, J. Quantitative palaeovegetation reconstruction at lager scale based on the pollen records. J. Plant Ecol. 2008, 32, 1201–1212. [Google Scholar]
- Wu, J.; Lv, J.; Ai, L. The impacts of climate change on the biodiversity: Vulnerability and adaptation. Ecol. Environ. Sci. 2009, 18, 693–703. [Google Scholar]
- Intergovernmental Panel on Climate Change (IPCC). Climate Change 2013: The Physical Science Basis Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Bertrand, R.; Lenoir, J.; Piedallu, C.; Riofrio-Dillon, G.; de Ruffray, P.; Vidal, C.; Pierrat, J.C.; Gegout, J.C. Changes in plant community composition lag behind climate warming in lowland forests. Nature 2011, 479, 517–520. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavorel, S.; Araújo, M.; Sykes, M.; Prentice, I. Climate change threats to plant diversity in Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar] [CrossRef] [Green Version]
- Heller, N.E.; Zavaleta, E.S. Biodiversity management in the face of climate change: A review of 22 years of recommendations. Biol. Conserv. 2009, 142, 14–32. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [Green Version]
- Diez, J.M.; D’Antonio, C.M.; Dukes, J.S.; Grosholz, E.D.; Olden, J.D.; Sorte, C.J.B.; Blumenthal, D.M.; Bradley, B.A.; Early, R.; Ibáñez, I.; et al. Will extreme climatic events facilitate biological invasions? Front. Ecol. Environ. 2012, 10, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Grimm, N.B.; Chapin, F.S.; Bierwagen, B.; Gonzalez, P.; Groffman, P.M.; Luo, Y.; Melton, F.; Nadelhoffer, K.; Pairis, A.; Raymond, P.A.; et al. The impacts of climate change on ecosystem structure and function. Front. Ecol. Environ. 2013, 11, 474–482. [Google Scholar] [CrossRef] [Green Version]
- Qu, H.; Wang, C.-J.; Zhang, Z.-X. Planning priority conservation areas under climate change for six plant species with extremely small populations in China. Nat. Conserv. 2018, 25, 89–106. [Google Scholar] [CrossRef] [Green Version]
- John, W.W.; Stephen, T.J.; John, E.K. Projected distributions of novel and disappearing climates by 2100 AD. Proc. Natl. Acad. Sci. USA 2007, 104, 5738–5742. [Google Scholar]
- John, A.W.; Diana, S.; Dennis, J.; Christine, A.H.; Mark, A.S. Niches, models, and climate change: Assessing the assumptions and uncertainties. Proc. Natl. Acad. Sci. USA 2009, 106, 19729–19736. [Google Scholar]
- Bezeng, B.S.; Tesfamichael, S.G.; Dayananda, B. Predicting the effect of climate change on a range-restricted lizard in southeastern Australia. Curr. Zool. 2018, 64, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Çoban, H.O.; Örücü, Ö.K.; Arslan, E.S. MaxEnt Modeling for Predicting the Current and Future Potential Geographical Distribution of Quercus libani Olivier. Sustainability 2020, 12, 2671. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.-Q.; Kushwaha, S.P.S.; Saran, S.; Xu, J.; Roy, P.S. Maxent modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda L. in Lesser Himalayan foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Li, J.; Chang, H.; Liu, T.; Zhang, C. The potential geographical distribution of Haloxylon across Central Asia under climate change in the 21st century. Agric. For. Meteorol. 2019, 275, 243–254. [Google Scholar] [CrossRef]
- Jane, E.; Catherine, H.G.; Robert, P.A.; Miroslav, D.K.; Simon, F.; Antoine, G.; Robert, J.H.; Falk, H.; John, R.L.; Anthony, L.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar]
- Petitpierre, B.; Kueffer, C.; Broennimann, O.; Randin, C.; Daehler, C.; Guisan, A. Climatic niche shifts are rare among terrestrial plant invaders. Science 2012, 335, 1344–1348. [Google Scholar] [CrossRef] [Green Version]
- Sadia, E.A.; Greg, M.; Kenton, O.H.; Richard, H.; Lara, S.; Stephen, E.; Lucas, N.J. Scientists and software—Surveying the species distribution modelling community. Divers. Distrib. 2015, 21, 258–267. [Google Scholar]
- Barbosa, F.G.; Schneck, F. Characteristics of the top-cited papers in species distribution predictive models. Ecol. Model. 2015, 313, 77–83. [Google Scholar] [CrossRef]
- Zhang, K.; Yao, L.; Meng, J.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Townsend Peterson, A. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2006, 34, 102–117. [Google Scholar] [CrossRef]
- Stockwell, D. The GARP modelling system: Problems and solutions to automated spatial prediction. Int. J. Geogr. Inf. Sci. 1999, 13, 143–158. [Google Scholar] [CrossRef]
- Elith, J.; Kearney, M.; Phillips, S. The art of modelling range-shifting species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Li, G.; Xu, G.; Guo, K.; Du, S. Geographical boundary and climatic analysis of Pinus tabulaeformis in China: Insights on its afforestation. Ecol. Eng. 2016, 86, 75–84. [Google Scholar] [CrossRef]
- Hu, Z.J.; Zhang, Y.L.; Yu, H.B. Simulation of Stipa purpurea distribution pattern on Tibetan Plateau based on MaxEnt model and GIS. Chin. J. Appl. Ecol. 2015, 26, 505–511. [Google Scholar]
- Lee, S.J.; Han, S.; Yoon, T.K.; Han, S.H.; Jung, Y.; Yun, S.J.; Son, Y. Growth and Physiological Characteristics of Pinus densiflora Seedlings in Response to Open-field Experimental Warming using the Infrared Lamp. J. Korean For. Soc. 2013, 102, 522–529. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.S.; Chun, Y.M.; Lee, H.; Pi, J.H.; Lim, C.H. Establishment, Regeneration, and Succession of Korean Red Pine (Pinus densiflora S. et Z.) Forest in Korea. In Conifers; IntechOpen: London, UK, 2018. [Google Scholar]
- Weng, Y.H.; Liu, K.J.; Chen, Y.B.; Li, Y.; Wang, J.; Meng, Q.F. Variation in cone and seed traits in a clonal seed orchard of red pine (Pinus koraiensis Sieb. et Zucc.). Scand. J. For. Res. 2020, 35, 1–9. [Google Scholar] [CrossRef]
- Liang, D.; Ding, C.; Zhao, G.; Leng, W.; Zhang, M.; Zhao, X.; Qu, G. Variation and selection analysis of Pinus koraiensis clones in northeast China. J. For. Res. 2017, 29, 611–622. [Google Scholar] [CrossRef]
- Park, J.-M.; Kwon, S.-H.; Lee, H.-J.; Na, S.-J.; El-Kassaby, Y.A.; Kang, K.-S. Integrating fecundity variation and genetic relatedness in estimating the gene diversity of seed crops: Pinus koraiensis seed orchard as an example. Can. J. For. Res. 2017, 47, 366–370. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Otto-Bliesner, B.L.; Marshall, S.J.; Overpeck, J.T.; Miller, G.H.; Hu, A. Simulating Arctic climate warmth and icefield retreat in the last interglaciation. Science 2006, 311, 1751–1753. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Yi, Y.-J.; Cheng, X.; Yang, Z.-F.; Zhang, S.-H. Maxent modeling for predicting the potential distribution of endangered medicinal plant (H. riparia Lour) in Yunnan, China. Ecol. Eng. 2016, 92, 260–269. [Google Scholar] [CrossRef]
- Li, J.; Fan, G.; He, Y. Predicting the current and future distribution of three Coptis herbs in China under climate change conditions, using the MaxEnt model and chemical analysis. Sci. Total Environ. 2020, 698, 134141. [Google Scholar] [CrossRef]
- Brown, J.L.; Anderson, B. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Franklin, J. Species distribution models in conservation biogeography: Developments and challenges. Divers. Distrib. 2013, 19, 1217–1223. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Wang, Y. Application of ROC curve analysis in evaluating the performance of alien species’ potential distribution models. Biodivers. Sci. 2007, 15, 365–372. [Google Scholar]
- Wang, R.; Yang, H.; Luo, W.; Wang, M.; Lu, X.; Huang, T.; Zhao, J.; Li, Q. Predicting the potential distribution of the Asian citrus psyllid, Diaphorina citri (Kuwayama), in China using the MaxEnt model. PeerJ 2019, 7, e7323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaiah, G.; Amon, M.; Munyaradzi, D.S.; Mhosisi, M.; Christopher, C. Precipitation of the warmest quarter and temperature of the warmest month are key to understanding the effect of climate change on plant species diversity in Southern African savannah. Afr. J. Ecol. 2014, 52, 209–216. [Google Scholar]
- Wang, J.R.; Hawkins, C.D.B.; Letchford, T. Photosynthesis, water and nitrogen use effciencies of four paper birch (Betula papyrifera) populations grown under different soil moisture and nutrient regimes. For. Ecol. Manag. 1998, 112, 233–244. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, H.; Su, J.; Tao, J.; Zhao, D. Research Progress on the Effect of Abiotic Stress on the Growth and Development of Paeonia Plant. Mol. Plant Breed. 2018, 16, 5072–5079. [Google Scholar]
- Hu, J.; Liu, Y. Unveiling the conservation biogeography of a data-deficient endangered bird species under climate change. PLoS ONE 2014, 9, e84529. [Google Scholar] [CrossRef]
- Qiu, Y.X.; Fu, C.X.; Comes, H.P. Plant molecular phylogeography in China and adjacent regions: Tracing the genetic imprints of Quaternary climate and environmental change in the world’s most diverse temperate flora. Mol. Phylogenet. Evol. 2011, 59, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Provan, J.; Bennett, K.D. Phylogeographic insights into cryptic glacial refugia. Trends Ecol. Evol. 2008, 23, 564–571. [Google Scholar] [CrossRef]
- Nogués-Bravo, D. Predicting the past distribution of species climatic niches. Glob. Ecol. Biogeogr. 2009, 18, 521–531. [Google Scholar] [CrossRef]
- Thomas, E.; van Zonneveld, M.; Loo, J.; Hodgkin, T.; Galluzzi, G.; van Etten, J. Present spatial diversity patterns of Theobroma cacao L. in the neotropics reflect genetic differentiation in pleistocene refugia followed by human-influenced dispersal. PLoS ONE 2012, 7, e47676. [Google Scholar] [CrossRef] [Green Version]
- Leng, W.; He, H.S.; Bu, R.; Dai, L.; Hu, Y.; Wang, X. Predicting the distributions of suitable habitat for three larch species under climate warming in Northeastern China. For. Ecol. Manag. 2008, 254, 420–428. [Google Scholar] [CrossRef]
- Flower, A.; Murdock, T.Q.; Taylor, S.W.; Zwiers, F.W. Using an ensemble of downscaled climate model projections to assess impacts of climate change on the potential distribution of spruce and Douglas-fir forests in British Columbia. Environ. Sci. Policy 2013, 26, 63–74. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, X.P.; Zhang, Y.F.; Hu, R.; Pan, Y.X.; Chen, N. Responses of plant growth of different life forms to rainfall amount changes in an arid desert area. Chin. J. Ecol. 2015, 34, 1847–1853. [Google Scholar]
- Chen, Y.; Xie, H.; Luo, H.; Yang, B.; Xiong, D. Impacts of climate change on the distribution of Cymbidium kanran and the simulation of distribution pattern. Chin. J. Appl. Ecol. 2019, 30, 3419–3425. [Google Scholar]
- Welter, S.; Brunner, K.; Hofstraat, J.W.; De Cola, L. Electroluminescent device with reversible switching between red and green emission. Nature 2003, 421, 54–57. [Google Scholar] [CrossRef]
- Lenoir, J.; Gegout, J.C.; Marquet, P.A.; de Ruffray, P.; Brisse, H. A significant upward shift in plant species optimum elevation during the 20th century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef]
- Li, J.; Xu, Z.; Shi, M.; Chen, J. Prediction of potential geographical distribution patterns of Salix tetrasperma Roxb. in Asia under different climate scenarios. Acta Ecol. Sin. 2019, 39, 3224–3234. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Description | Whether to Use for Modeling |
|---|---|---|
| Bio1 | Mean Annual Temperature (°C) | Yes |
| Bio2 | Mean Diurnal Range (Mean of monthly (max temp-min temp)) (°C) | No |
| Bio3 | Isothermally (Bio2/Bio7) (×100) | Yes |
| Bio4 | Temperature Seasonality (standard deviation ×100) (C of V) | Yes |
| Bio5 | Max Temperature of Warmest Month (°C) | No |
| Bio6 | Min Temperature of Coldest Month (°C) | No |
| Bio7 | Temperature Annual Range (Bio5–Bio6) (°C) | No |
| Bio8 | Mean Temperature of Wettest Quarter (°C) | Yes |
| Bio9 | Mean Temperature of Driest Quarter (°C) | Yes |
| Bio10 | Mean Temperature of Warmest Quarter (°C) | No |
| Bio11 | Mean Temperature of Coldest Quarter (°C) | No |
| Bio12 | Annual Precipitation (mm) | No |
| Bio13 | Precipitation of Wettest Month (mm) | Yes |
| Bio14 | Precipitation of Driest Month (mm) | No |
| Bio15 | Precipitation Seasonality (C of V) | No |
| Bio16 | Precipitation of Wettest Quarter (mm) | No |
| Bio17 | Precipitation of Driest Quarter (mm) | No |
| Bio18 | Precipitation of Warmest Quarter (mm) | Yes |
| Bio19 | Precipitation of Coldest Quarter (mm) | No |
| Abbreviation | Description | Contribution (%) |
|---|---|---|
| Bio1 | Mean Annual Temperature (°C) | 6.6 |
| Bio3 | Isothermality (Bio2/Bio7) (×100) | 5.1 |
| Bio4 | Temperature Seasonality (standard deviation ×100) (C of V) | 13.9 |
| Bio8 | Mean Temperature of the Wettest Quarter (°C) | 5.6 |
| Bio9 | Mean Temperature of the Driest Quarter (°C) | 0.2 |
| Bio13 | Precipitation of the Wettest Month (mm) | 0.9 |
| Bio18 | Precipitation of the Warmest Quarter (mm) | 67.7 |
| Period | Low Suitability Area/104 km2 | Moderate Suitability Area/104 km2 | High Suitability Area/104 km2 | Total Suitable Area/104 km2 |
|---|---|---|---|---|
| LGM | 128.78 | 59.41 | 80.6 | 268.79 |
| Mid-Holocene | 171.32 | 76.27 | 72.29 | 319.88 |
| Current | 232.75 | 74.69 | 54.62 | 362.06 |
| PCR2.6-2050 | 142.21 | 58.18 | 52.36 | 252.75 |
| PCR2.6-2070 | 119.92 | 49.86 | 61.27 | 231.05 |
| PCR 8.5-2050 | 131.11 | 58.66 | 53.79 | 243.56 |
| PCR 8.5-2070 | 127.52 | 46 | 56 | 229.52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, X.; Li, J.; Wu, S. MaxEnt Modeling to Estimate the Impact of Climate Factors on Distribution of Pinus densiflora. Forests 2022, 13, 402. https://doi.org/10.3390/f13030402
Duan X, Li J, Wu S. MaxEnt Modeling to Estimate the Impact of Climate Factors on Distribution of Pinus densiflora. Forests. 2022; 13(3):402. https://doi.org/10.3390/f13030402
Chicago/Turabian StyleDuan, Xiangguang, Junqing Li, and Shuhong Wu. 2022. "MaxEnt Modeling to Estimate the Impact of Climate Factors on Distribution of Pinus densiflora" Forests 13, no. 3: 402. https://doi.org/10.3390/f13030402
APA StyleDuan, X., Li, J., & Wu, S. (2022). MaxEnt Modeling to Estimate the Impact of Climate Factors on Distribution of Pinus densiflora. Forests, 13(3), 402. https://doi.org/10.3390/f13030402