
Fungal and Bacterial Communities in Tuber melanosporum Plantations from Northern Spain

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Sampling
2.2. Data Analysis
2.2.1. Descriptive Analysis
2.2.2. Cluster Analysis
3. Results
3.1. Descriptive Analysis
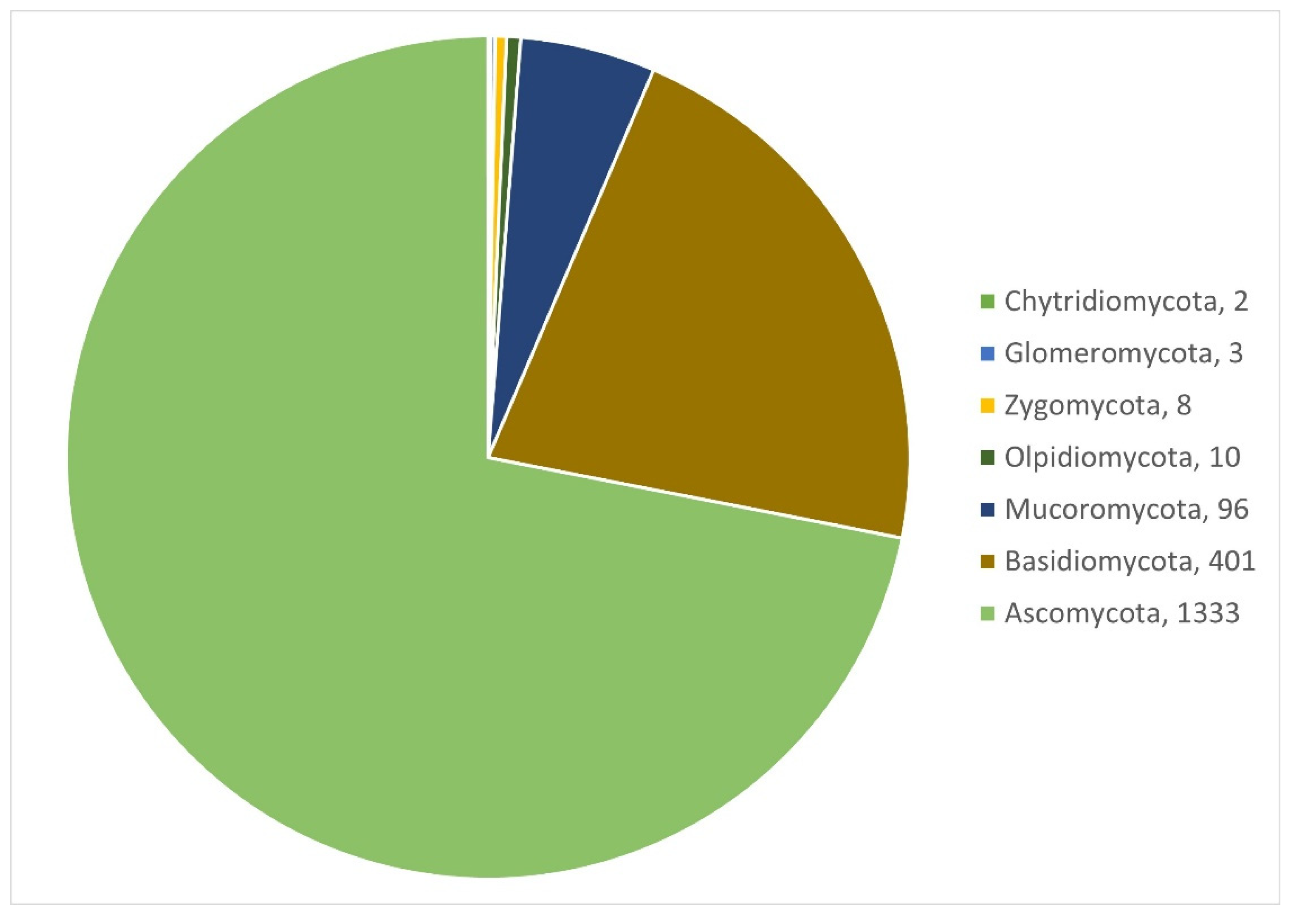
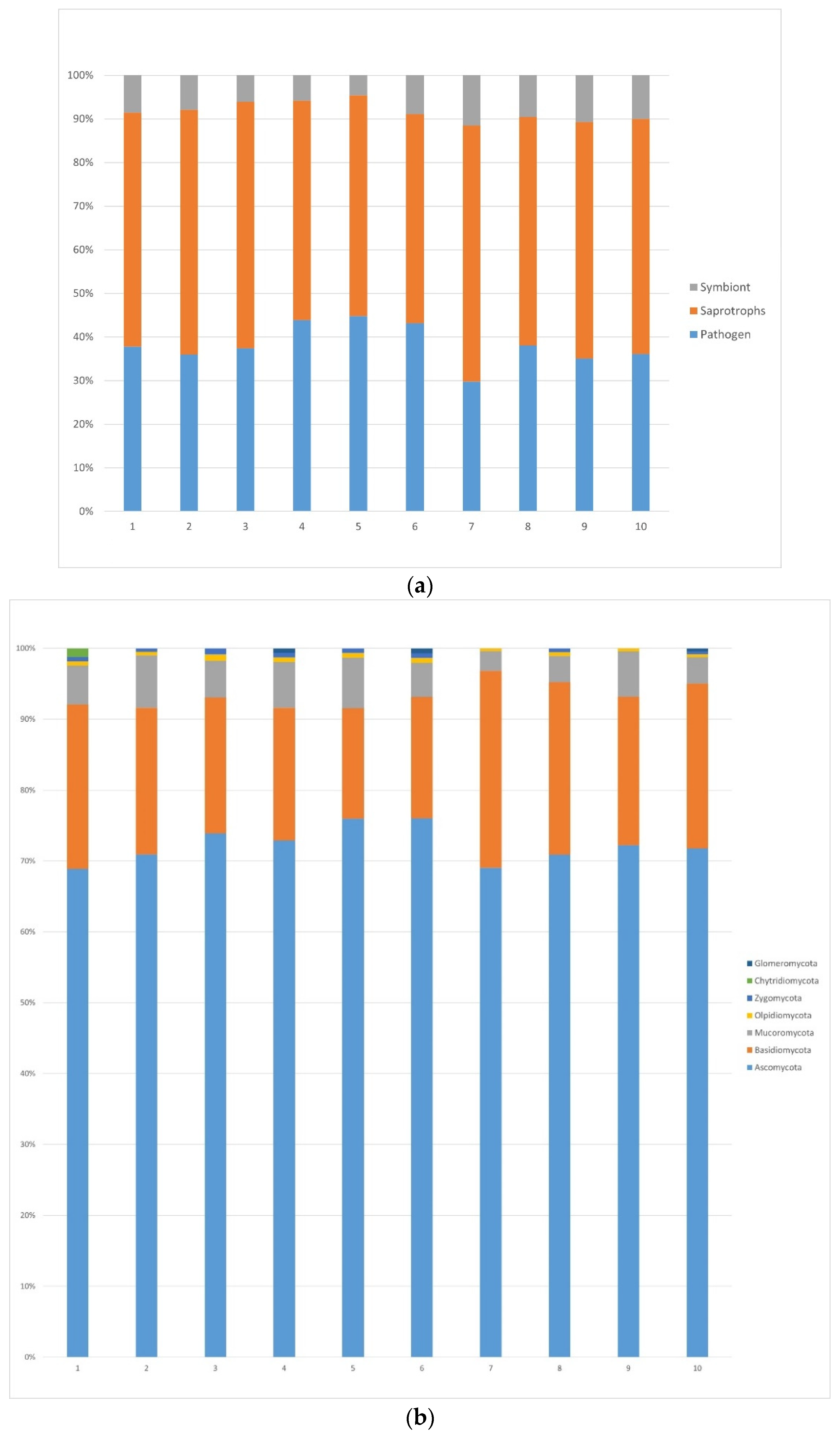
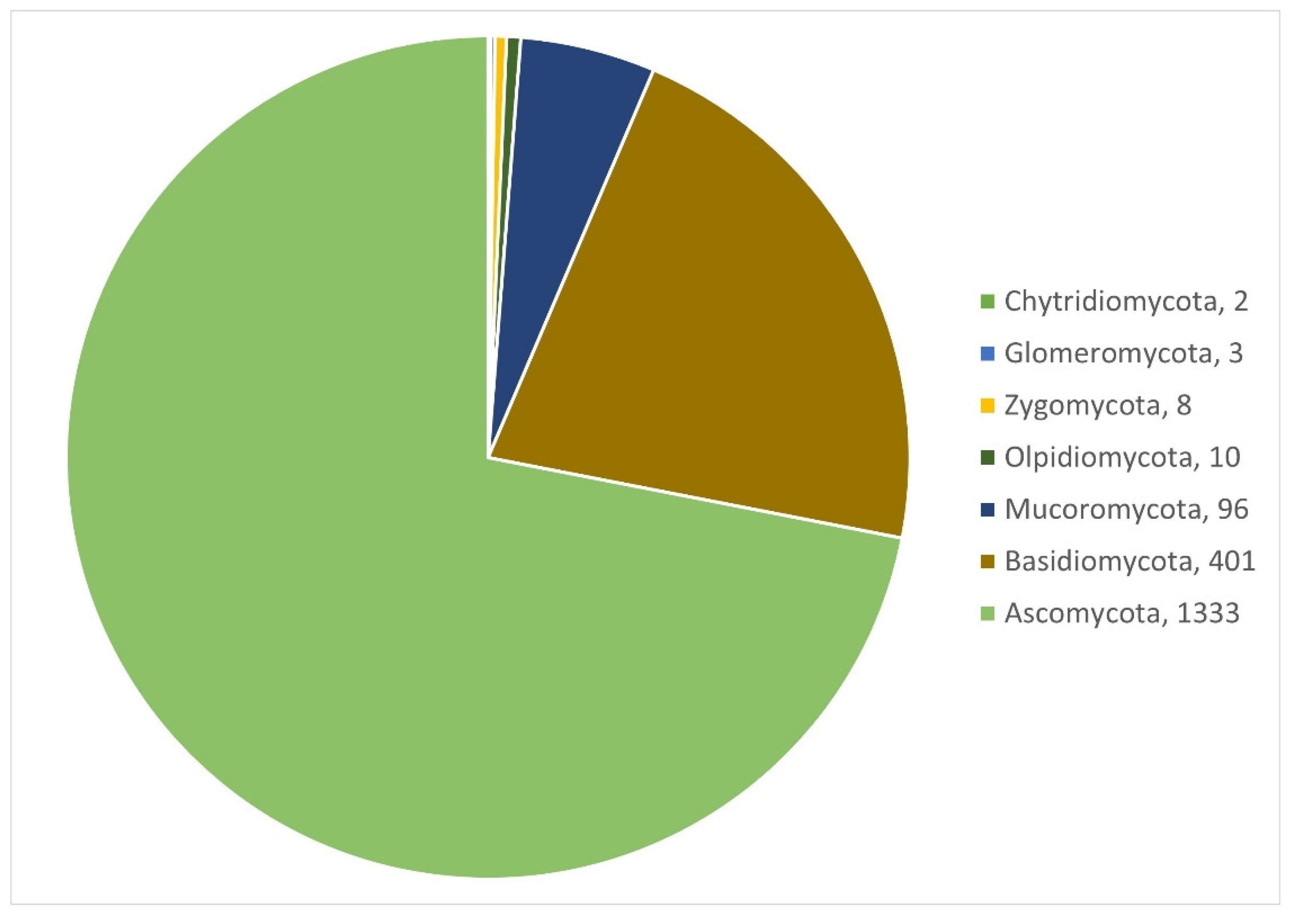
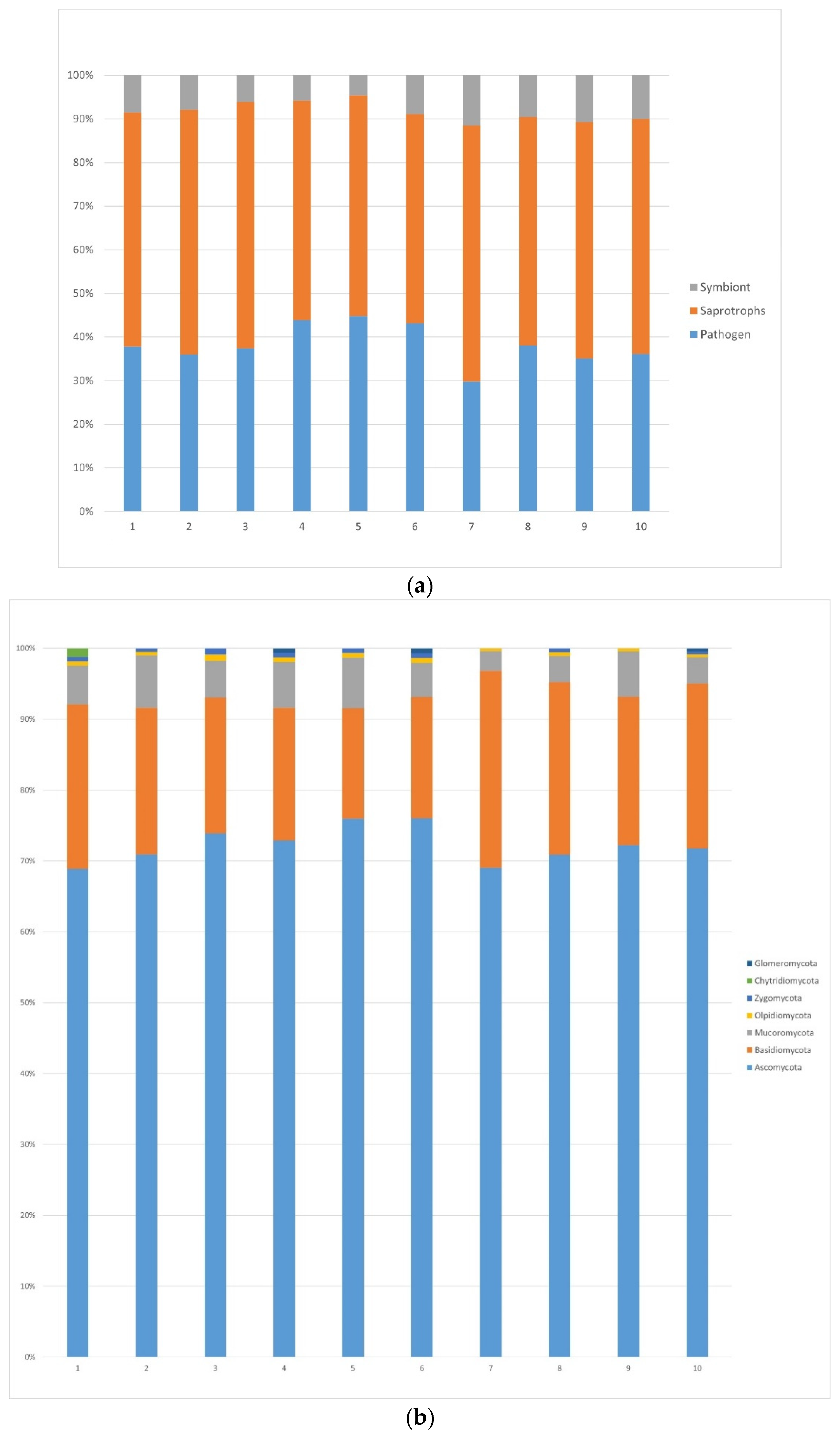
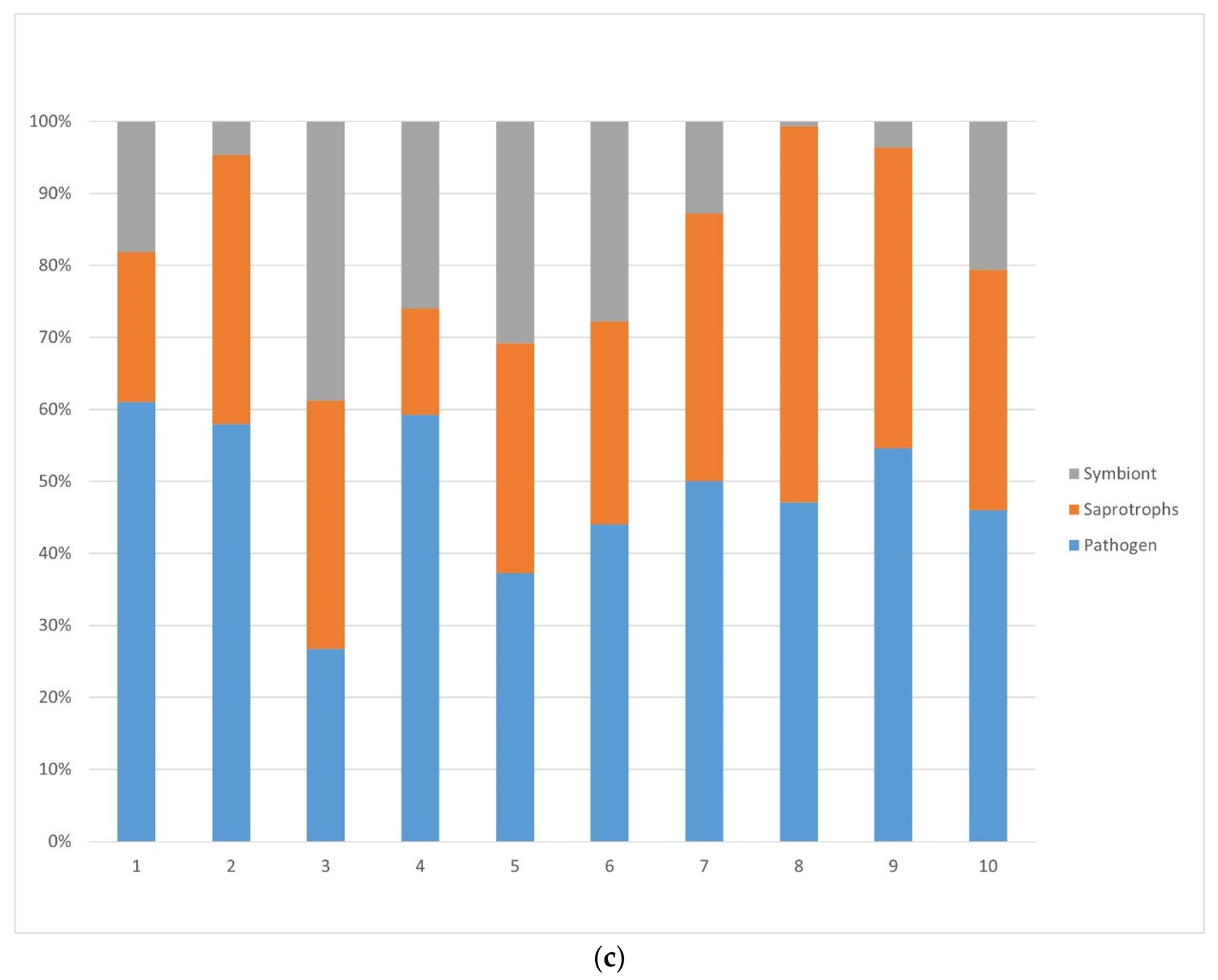
3.1.1. Fungi
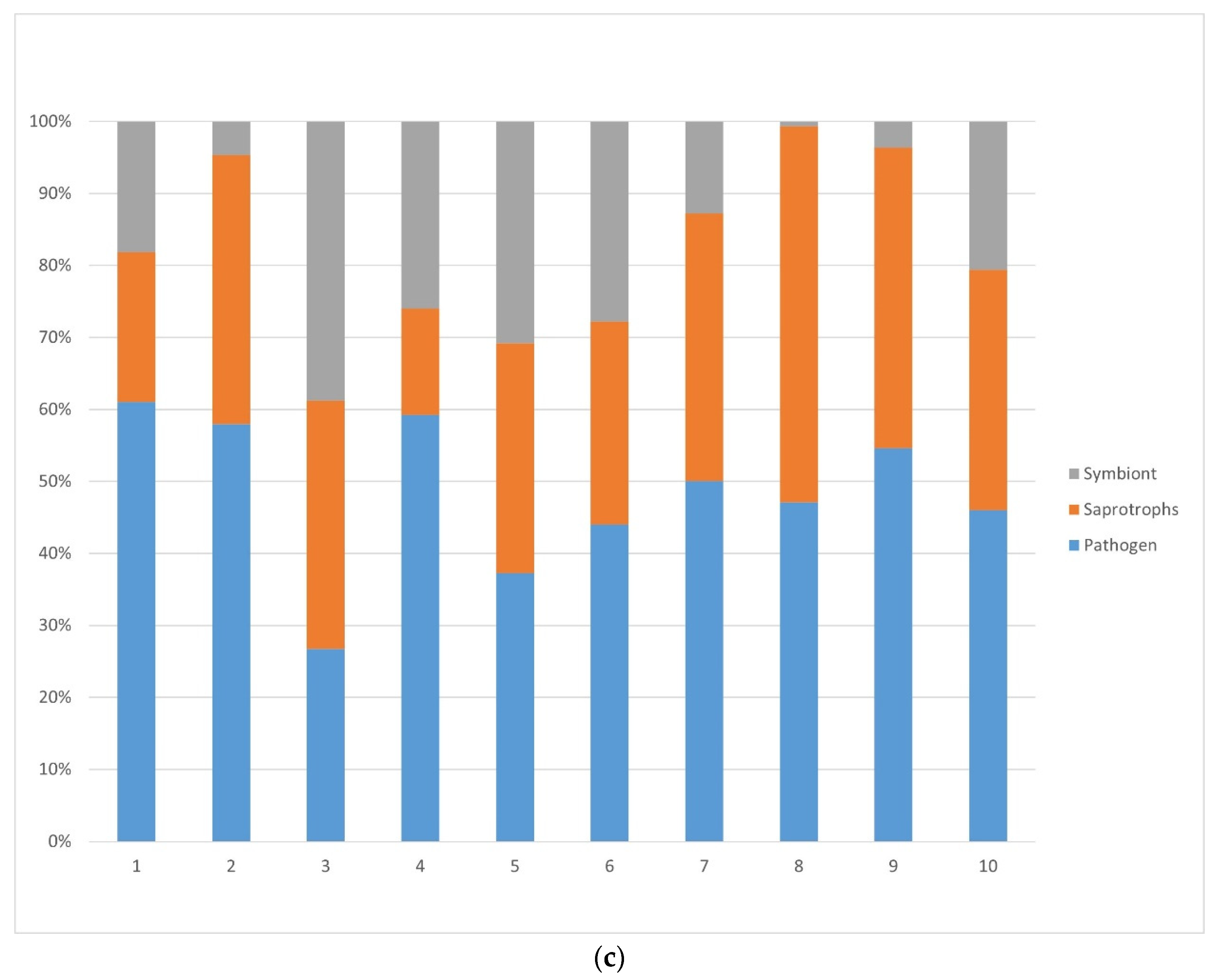
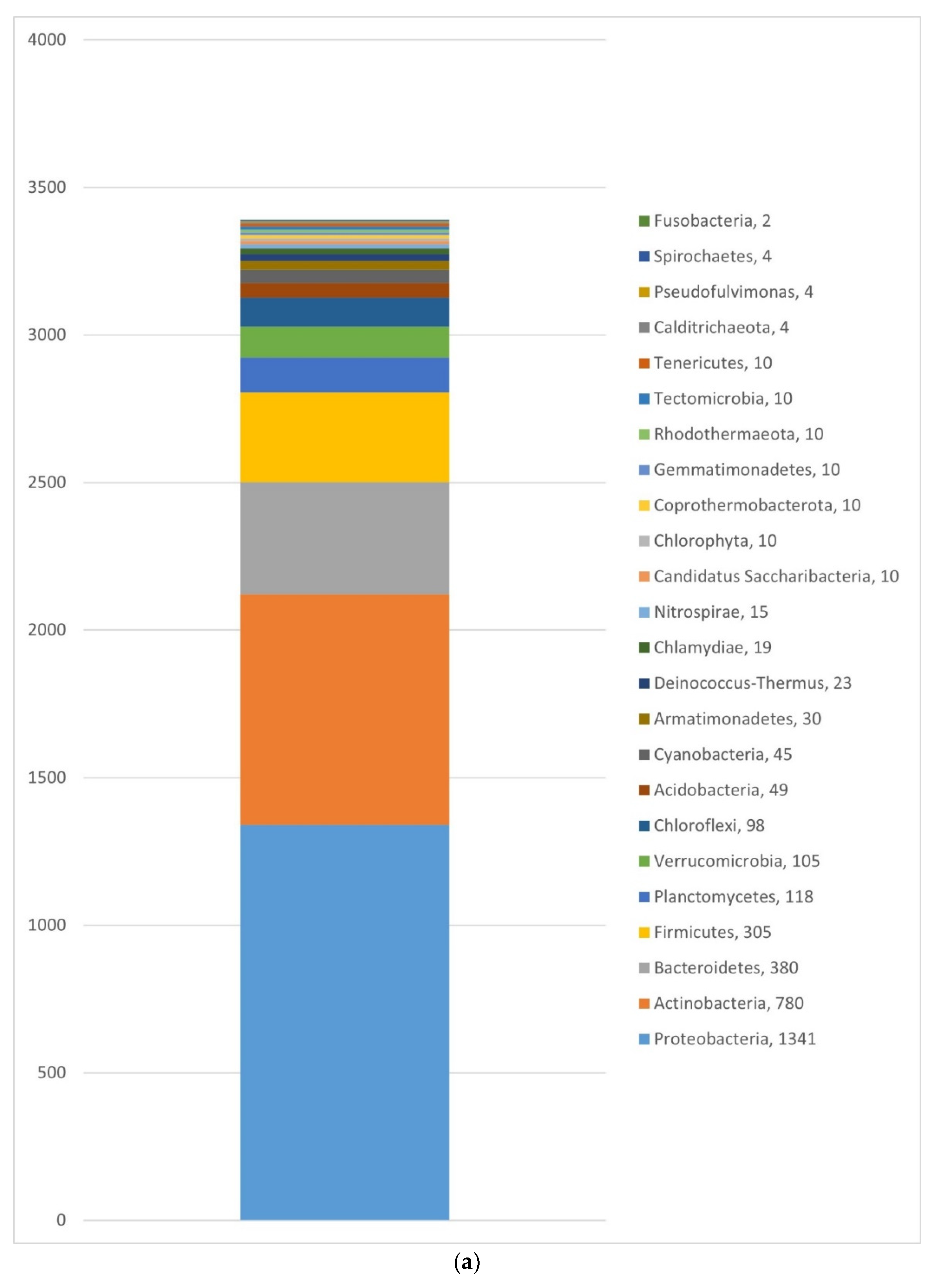
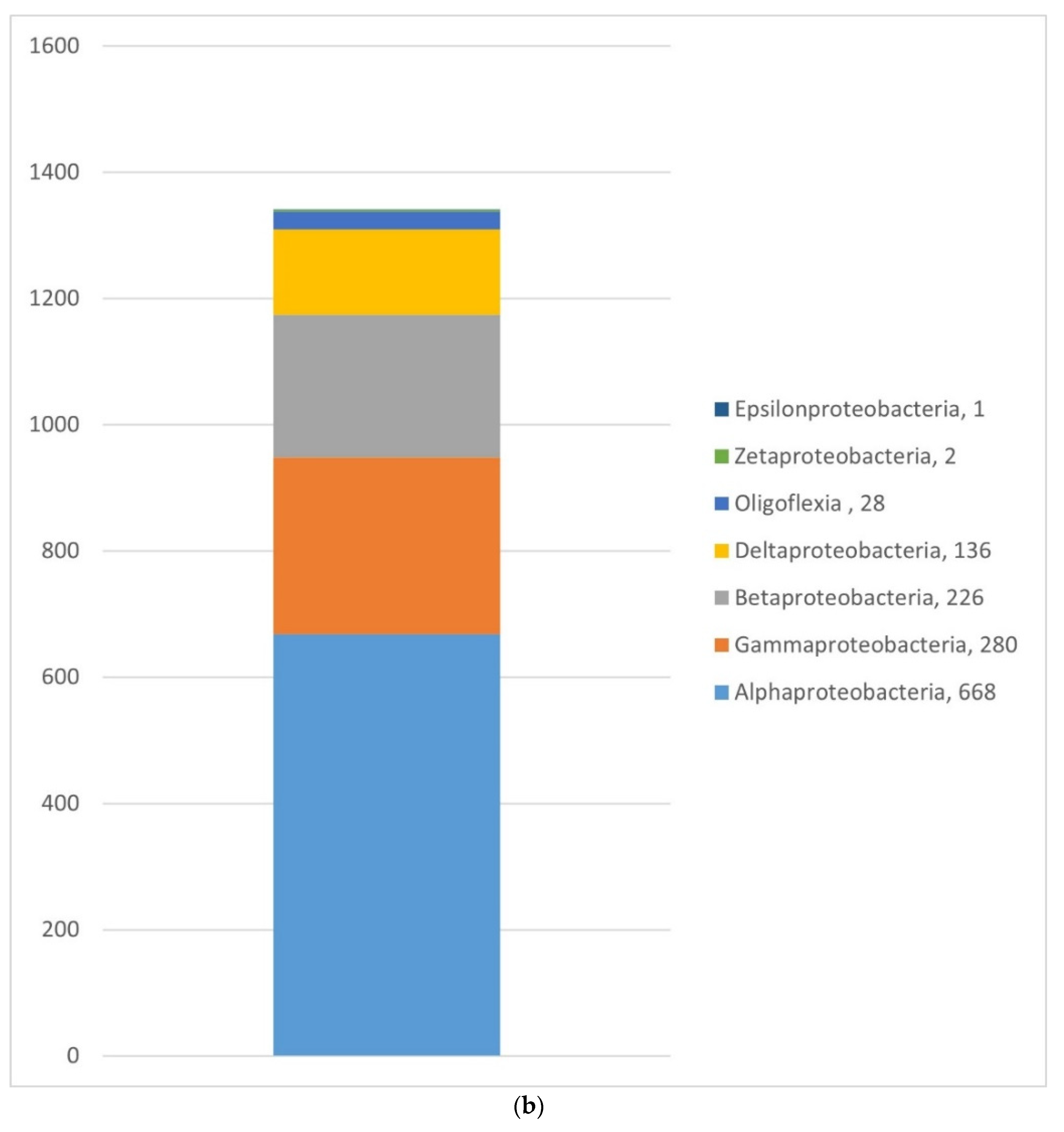
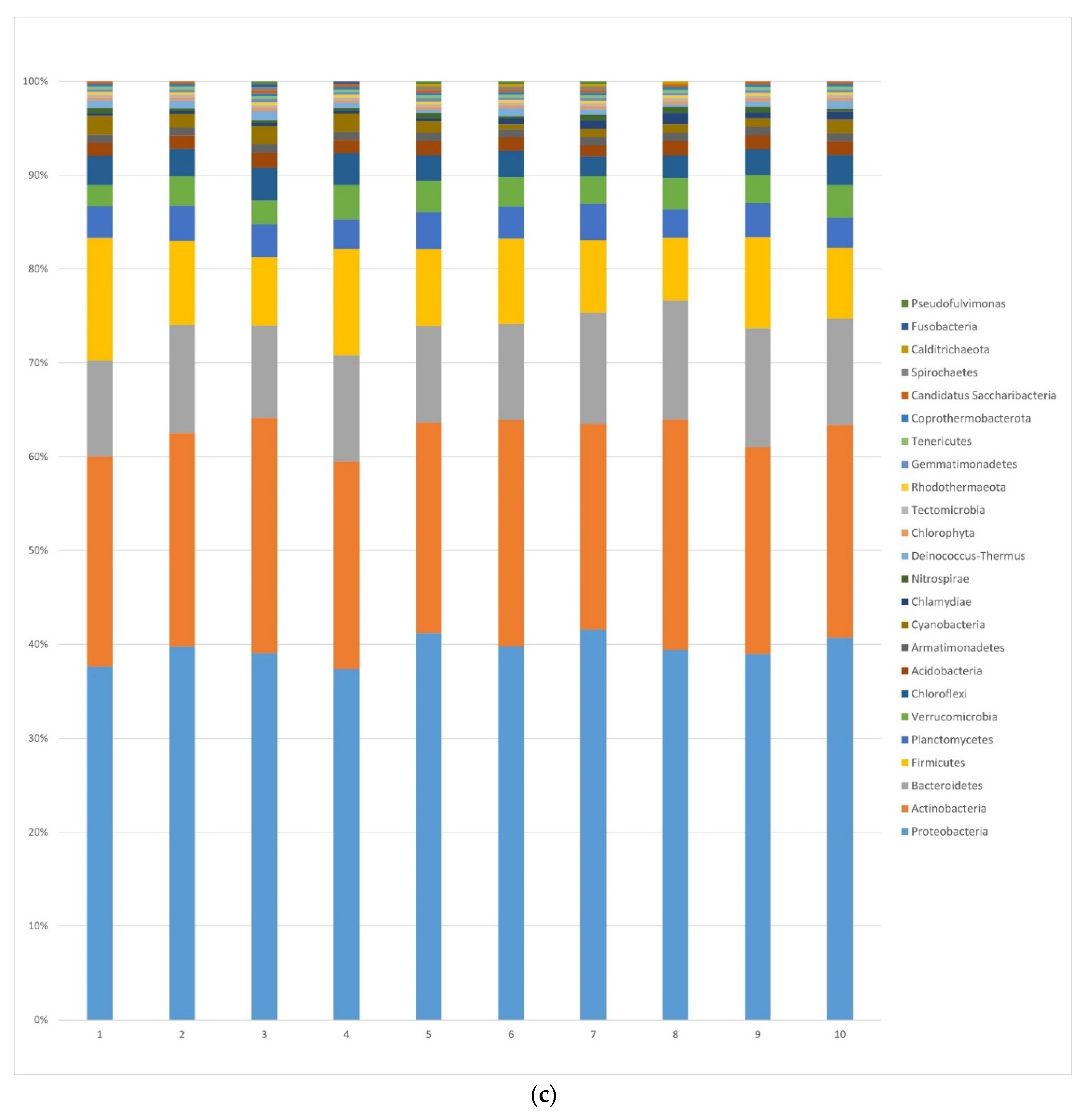
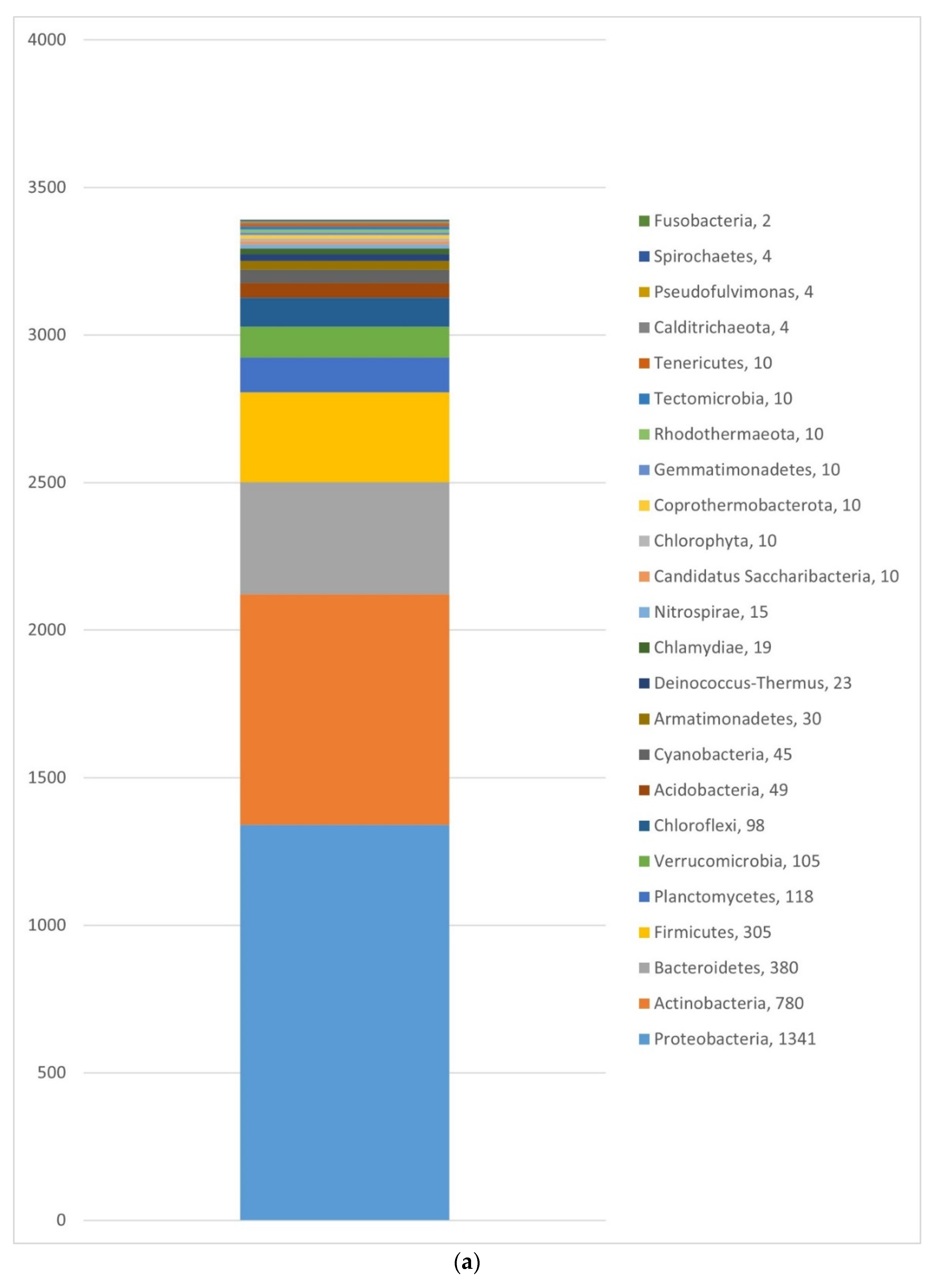
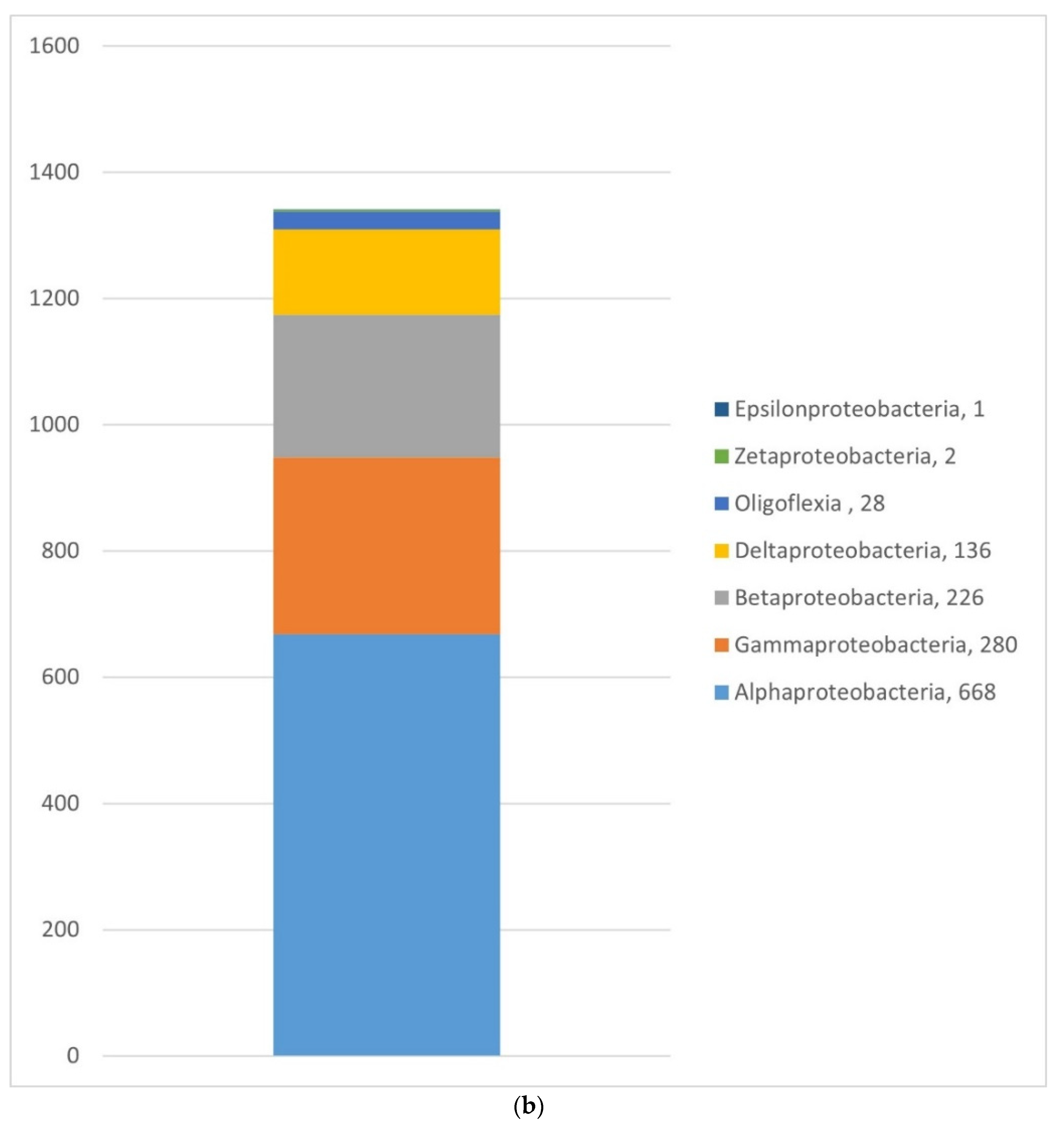
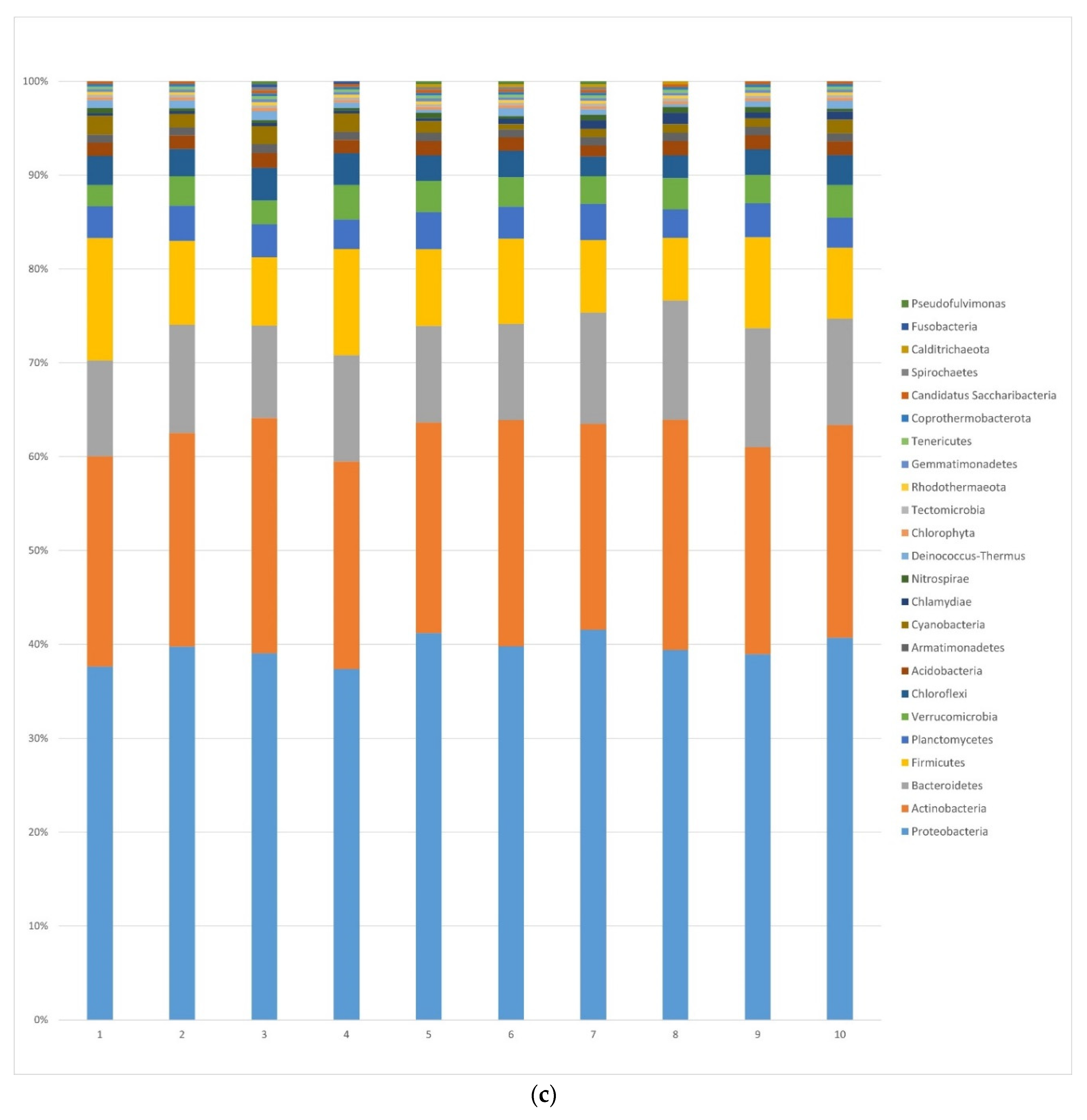
3.1.2. Bacteria
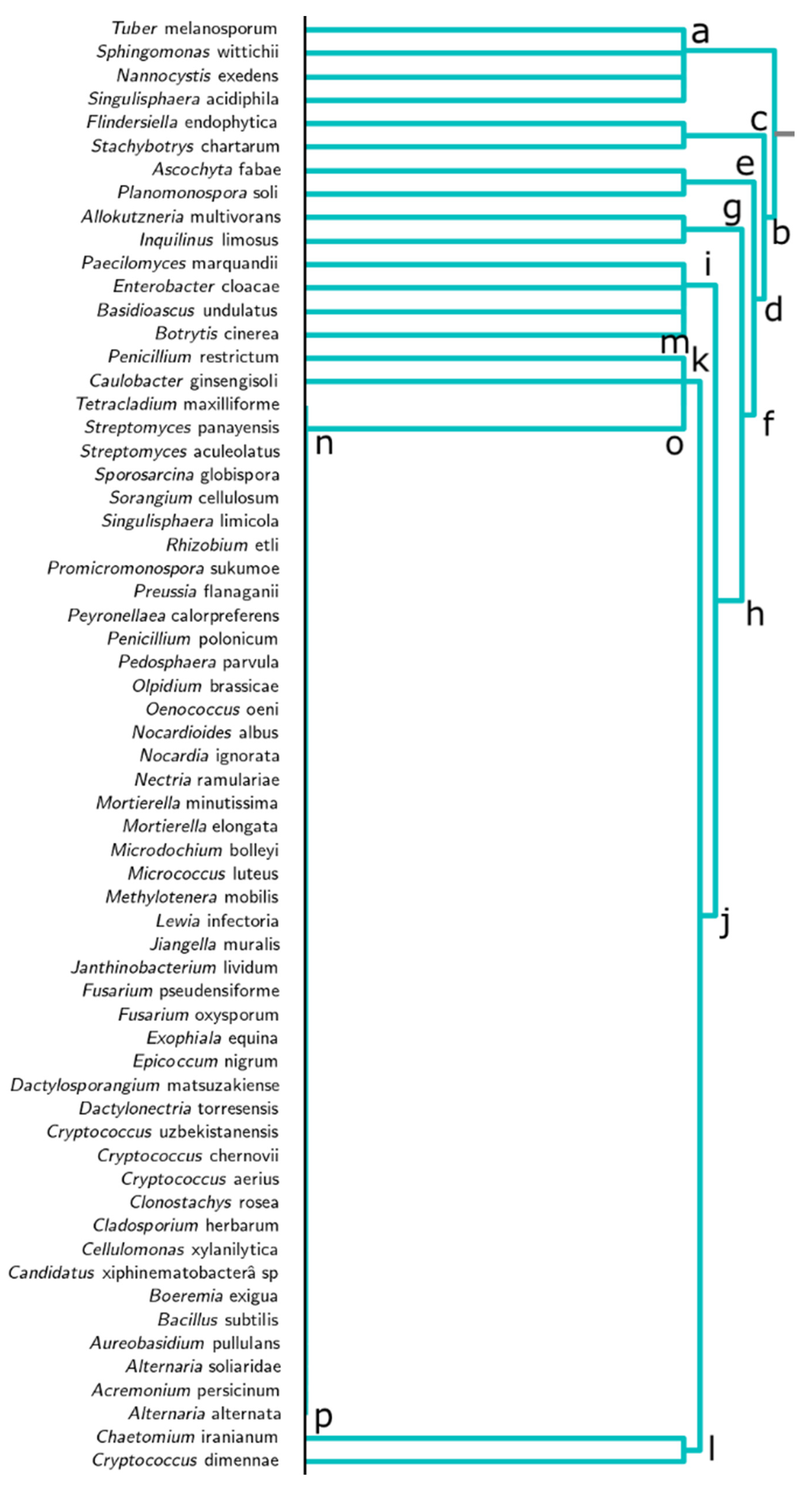
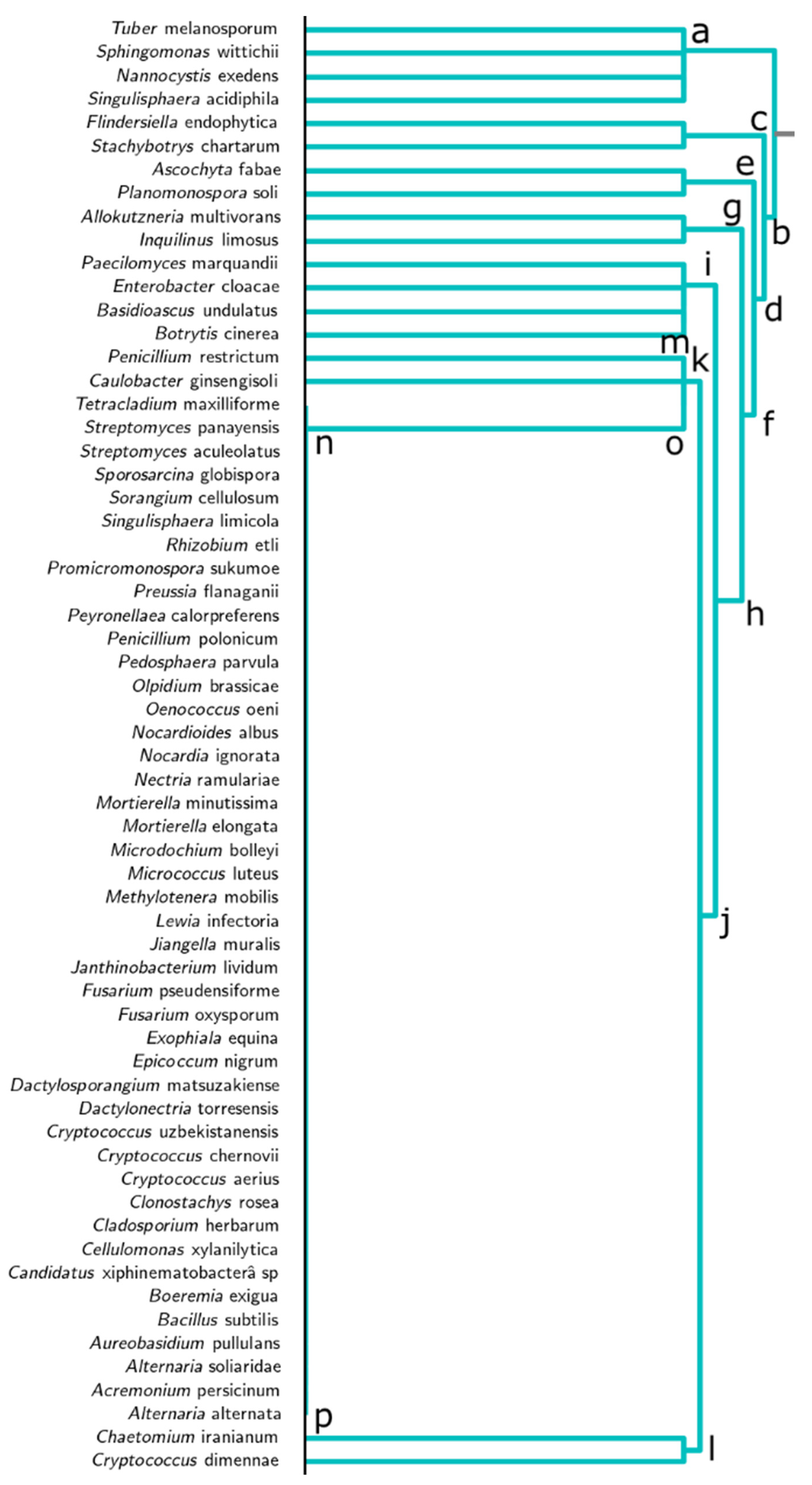
3.2. Dendrogram Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mello, A.; Ding, G.C.; Piceno, Y.M.; Napoli, C.; Tom, L.M.; DeSantis, T.Z.; Andersen, G.L.; Smalla, K.; Bonfante, P. Truffle Brûlés Have an Impact on the Diversity of Soil Bacterial Communities. PLoS ONE 2013, 8, e61945. [Google Scholar] [CrossRef]
- Büntgen, U.; Oliach, D.; Martínez-Peña, F.; Latorre, J.; Egli, S.; Krusic, P.J. Black truffle winter production depends on Mediterranean summer precipitation. Environ. Res. Lett. 2019, 14, 74004. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Barreda, S.; Sánchez, S.; Marco, P.; Serrano-Notivoli, R. Agro-climatic zoning of Spanish forests naturally producing black truffle. Agric. For. Meteorol. 2019, 269–270, 231–238. [Google Scholar] [CrossRef]
- Splivallo, R.; Vahdatzadeh, M.; MacIá-Vicente, J.G.; Molinier, V.; Peter, M.; Egli, S.; Uroz, S.; Paolocci, F.; Deveau, A. Orchard conditions and fruiting body characteristics drive the microbiome of the black truffle tuber aestivum. Front. Microbiol. 2019, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Oliach, D.; Castaño, C.; Fischer, C.R.; Barry-Etienne, D.; Bonet, J.A.; Colinas, C.; Oliva, J. Soil fungal community and mating type development of Tuber melanosporum in a 20-year chronosequence of black truffle plantations. Soil Biol. Biochem. 2022, 165, 108510. [Google Scholar] [CrossRef]
- De la Varga, H.; le Tacon, F.; Lagoguet, M.; Todesco, F.; Varga, T.; Miquel, I.; Barry-Etienne, D.; Robin, C.; Halkett, F.; Martin, F.; et al. Five years investigation of female and male genotypes in périgord black truffle (Tuber melanosporum Vittad.) revealed contrasted reproduction strategies. Environ. Microbiol. 2017, 19, 2604–2615. [Google Scholar] [CrossRef]
- Oliach, D.; Vidale, E.; Brenko, A.; Marois, O.; Andrighetto, N.; Stara, K.; de Aragón, J.M.; Colinas, C.; Bonet, J.A. Truffle market evolution: An application of the delphi method. Forests 2021, 12, 1174. [Google Scholar] [CrossRef]
- Murat, C. Forty years of inoculating seedlings with truffle fungi: Past and future perspectives. Mycorrhiza 2015, 25, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Reyna, S.; Garcia-Barreda, S. Black truffle cultivation: A global reality. For. Syst. 2014, 23, 317–328. [Google Scholar] [CrossRef]
- Deveau, A.; Clowez, P.; Petit, F.; Maurice, J.P.; Todesco, F.; Murat, C.; Harroué, M.; Ruelle, J.; le Tacon, F. New insights into black truffle biology: Discovery of the potential connecting structure between a Tuber aestivum ascocarp and its host root. Mycorrhiza 2019, 29, 219–226. [Google Scholar] [CrossRef]
- Callot, G. La Truffe, la Terre, la Vie; INRA: Paris, France, 1999. [Google Scholar]
- Olivier, J.; Savignac, J.; Sourzat, P. Truffe et Trufficulture; FANLAC: Périgueux, France, 2012. [Google Scholar]
- Barbieri, E.; Ceccaroli, P.; Agostini, D.; Zeppa, S.D.; Gioacchini, A.M.; Stocchi, V. Truffle-associated bacteria: Extrapolation from diversity to function. In True Truffle (Tuber spp.) in the World; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; Volume 47, p. 436. ISBN 331931436X. [Google Scholar]
- Luoma, D.L.; Eberhart, J.L.; Molina, R.; Amaranthus, M.P. Response of ectomycorrhizal fungus sporocarp production to varying levels and patterns of green-tree retention. For. Ecol. Manag. 2004, 202, 337–354. [Google Scholar] [CrossRef]
- Brundrett, M.C. Mycorrhizal associations and other means of nutrition of vascular plants: Understanding the global diversity of host plants by resolving conflicting information and developing reliable means of diagnosis. Plant Soil 2009, 320, 37–77. [Google Scholar] [CrossRef]
- Courty, P.E.; Walder, F.; Boller, T.; Ineichen, K.; Wiemken, A.; Rousteau, A.; Selosse, M.A. Carbon and nitrogen metabolism in mycorrhizal networks and mycoheterotrophic plants of tropical forests: A stable isotope analysis. Plant Physiol. 2011, 156, 952–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, D.D.; Neal, A.L.; van Wees, S.C.M.; Ton, J. Mycorrhiza-induced resistance: More than the sum of its parts? Trends Plant Sci. 2014, 18, 539–545. [Google Scholar] [CrossRef] [Green Version]
- Barbieri, E.; Bertini, L.; Rossi, I.; Ceccaroli, P.; Saltarelli, R.; Guidi, C.; Zambonelli, A.; Stocchi, V. New evidence for bacterial diversity in the ascoma of the ectomycorrhizal fungus Tuber borchii Vittad. FEMS Microbiol. Lett. 2005, 247, 23–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonfante, P.; Anca, I. Plants, Mycorrhizal Fungi, and Bacteria: A Network of Interactions. Annu. Rev. Microbiol. 2009, 63, 363–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frey-Klett, P.; Garbaye, J.; Tarkka, M. The mycorrhiza helper bacteria revisited. New Phytol. 2007, 176, 22–36. [Google Scholar] [CrossRef]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every base matters: Assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Mediavilla, O.; Geml, J.; Olaizola, J.; Oria-de-Rueda, J.A.; Baldrian, P.; Martín-Pinto, P. Effect of forest fire prevention treatments on bacterial communities associated with productive Boletus edulis sites. Microb. Biotechnol. 2019, 12, 1188–1198. [Google Scholar] [CrossRef] [Green Version]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Forest soil bacteria: Diversity, involvement in ecosystem processes, and response to global change. Microbiol. Mol. Biol. Rev. 2017, 81, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Aspray, T.J.; Frey-Klett, P.; Jones, J.E.; Whipps, J.M.; Garbaye, J.; Bending, G.D. Mycorrhization helper bacteria: A case of specificity for altering ectomycorrhiza architecture but not ectomycorrhiza formation. Mycorrhiza 2006, 16, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Splivallo, R.; Deveau, A.; Valdez, N.; Kirchhoff, N.; Frey-Klett, P.; Karlovsky, P. Bacteria associated with truffle-fruiting bodies contribute to truffle aroma. Environ. Microbiol. 2015, 17, 2647–2660. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D.J. Structure and development of ectomycorrhizal roots. In Molecular Mycorrhizal Symbiosis; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Fuhrman, J.A. Microbial community structure and its functional implications. Nature 2009, 459, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Taschen, E.; Sauve, M.; Vincent, B.; Parladé, J.; van Tuinen, D.; Aumeeruddy-Thomas, Y.; Assenat, B.; Selosse, M.A.; Richard, F. Insight into the truffle brûlé: Tripartite interactions between the black truffle (Tuber melanosporum), holm oak (Quercus ilex) and arbuscular mycorrhizal plants. Plant Soil 2020, 446, 577–594. [Google Scholar] [CrossRef]
- Baldrián, P.; Kolarik, M.; Stursova, M.; Kopecky, J.; Valaskova, V.; Vetrovsky, T.; Zifcáková, L.; Snajdr, J.; Ridl, J.; Vlcek, C.; et al. Active and total microbial communities in forest soil are largely different and highly stratified during decomposition. ISME J. 2012, 6, 248–258. [Google Scholar] [CrossRef] [Green Version]
- Karimi, B.; Maron, P.A.; Chemidlin-Prevost Boure, N.; Bernard, N.; Gilbert, D.; Ranjard, L. Microbial diversity and ecological networks as indicators of environmental quality. Environ. Chem. Lett. 2017, 15, 265–281. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Sokal, R.R.; Michener, C.D. A Statistical Methods for Evaluating Relationships. Univ. Kansas Sci. Bull. 1958, 38, 1409–1448. [Google Scholar]
- Jaccard, P. Etude comparative de la distribution florale dans une portion des Alpes et du Jura. Bull. Société Vaudoise Sci. Nat. 1901, 37, 547–579. [Google Scholar]
- Russel, P.; Rao, T.R. On habitat and association of species of anopheline larvae in south-eastern Madras. J. Malar. Inst. India 1940, 3, 154–178. [Google Scholar]
- Legendre, L.; Legendre, P. Écologie Numérique: La Structure des Données Écologiques, 2nd ed.; Collection D’écologie; Masson et Le Presses de L’université du Québec: Québec City, QC, Canada, 1984. [Google Scholar]
- Clifford, H.T.; Stephenson, W. An Introduction to Numerical Classification; Academic Press: New York, NY, USA, 1975. [Google Scholar]
- Ward, J.H. Hierarchical Grouping to Optimize an Objective Function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Sneath, P.H.; Sokal, R. Numerical Taxonomy: The Principles and Practice of Numerical Classification; Freeman and Company: San Francisco, CA, USA, 1973. [Google Scholar]
- Carrasco, J.; Preston, G.M. Growing edible mushrooms: A conversation between bacteria and fungi. Environ. Microbiol. 2020, 22, 858–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mello, A.; Miozzi, L.; Vizzini, A.; Napoli, C.; Kowalchuk, G.; Bonfante, P. Bacterial and fungal communities associated with Tuber magnatum-productive niches. Plant Biosyst. 2010, 144, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Xiaolin, L.; Quiang, L.; Haowei, W.; Chuan, X.; Qi, G.; Honghu, S.; Qun, S. Soil microbial communities of three major Chinese truffles in Southwest China. Can. J. Microbiol. 2016, 62, 348–349. [Google Scholar] [CrossRef] [Green Version]
- Tamayo-Velez, A.; Osorio, N.W. Co-inoculation with an arbuscular mycorrhizal fungus and a phosphate-solubilizing fungus promotes the plant growth and phosphate uptake of avocado plantlets in a nursery. Botany 2017, 95, 539–545. [Google Scholar] [CrossRef] [Green Version]
- Osorio, N.W.; Habte, M. Synergistic influence of an arbuscular mycorrhizal fungus and a P solubilizing fungus on growth and P uptake of Leucaena leucocephala in an oxisol. Arid. Land Res. Manag. 2001, 15, 263–274. [Google Scholar] [CrossRef]
- Zacchi, L.; Vaughan-Martini, A.; Angelini, P. Yeast distribution in a truffle-field ecosystem. Ann. Microbiol. 2003, 53, 275–282. [Google Scholar]
- Antony-Babu, S.; Deveau, A.; van Nostrand, J.D.; Zhou, J.; le Tacon, F.; Robin, C.; Frey-Klett, P.; Uroz, S. Black truffle-associated bacterial communities during the development and maturation of Tuber melanosporum ascocarps and putative functional roles. Environ. Microbiol. 2014, 16, 2831–2847. [Google Scholar] [CrossRef]
- Zhang, X.; Li, X.; Ye, L.; Huang, Y.; Kang, Z.; Zhang, B. Colonization by Tuber melanosporum and Tuber indicum affects the growth of Pinus armandii and phoD alkaline phosphatase encoding bacterial community in the rhizosphere. Microbiol. Res. 2020, 239, 126520. [Google Scholar] [CrossRef]
- Yang, M.; Zou, J.; Liu, C.; Xiao, Y.; Zhang, X.; Yan, L.; Ye, L.; Tang, P.; Li, X. Chinese white truffles shape the ectomycorrhizal microbial communities of Corylus avellana. Ann. Microbiol. 2019, 69, 553–565. [Google Scholar] [CrossRef]
- Ettema, T.; Anderson, G. The α-Proteobacteria: The Darwin finches of the bacterial world. Biol. Lett. 2009, 5, 429–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbieri, E.; Guidi, C.; Bertaux, J.; Frey-klett, P.; Garbaye, J.; Ceccaroli, P.; Saltarelli, R.; Zambonelli, A.; Stocchi, V.; Arbres-microorganismes, I.; et al. Occurrence and diversity of bacterial communities in Tuber magnatum during truffle maturation. Environ. Microbiol. 2007, 9, 2234–2246. [Google Scholar] [CrossRef] [PubMed]
- Hibbing, M.E.; Fuqua, C.; Parsek, M.R.; Brook Peterson, S. Bacterial competition: Surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 2010, 8, 15–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deveau, A.; Antony-Babu, S.; le Tacon, F.; Robin, C.; Frey-Klett, P.; Uroz, S. Temporal changes of bacterial communities in the Tuber melanosporum ectomycorrhizosphere during ascocarp development. Mycorrhiza 2016, 26, 389–399. [Google Scholar] [CrossRef]
- Garbaye, J. Helper bacteria: A new dimension to the mycorrhizal symbiosis. New Phytol. 1994, 128, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Brulé, C.; Frey-Klett, P.; Pierrat, J.C.; Courrier, S.; Gérard, F.; Lemoine, M.C.; Rousselet, J.L.; Sommer, G.; Garbaye, J. Survival in the soil of the ectomycorrhizal fungus Laccaria bicolor and the effects of a mycorrhiza helper Pseudomonas fluorescens. Soil Biol. Biochem. 2001, 33, 1683–1694. [Google Scholar] [CrossRef]
- Kurth, V.J.; Fransioli, N.; Fulé, P.Z.; Hart, S.C.; Gehring, C.A. Stand-replacing wildfires alter the community structure of wood-inhabiting fungi in southwestern ponderosa pine forests of the USA. Fungal Ecol. 2013, 6, 192–204. [Google Scholar] [CrossRef]
- Mamoun, M.; Olivier, J.M. Effect of soil Pseudomonas on colonization of hazel roots by the ecto-mycorrhizal species Tuber melanosporum and its competitors. Plant. Soil 1992, 139, 265–273. [Google Scholar] [CrossRef]
- Dominguez, J.A.; Martin, A.; Anriquez, A.; Albanesi, A. The combined effects of Pseudomonas fluorescens and Tuber melanosporum on the quality of Pinus halepensis seedlings. Mycorrhiza 2012, 22, 429–436. [Google Scholar] [CrossRef]
- Barbieri, E.; Ceccaroli, P.; Saltarelli, R.; Guidi, C.; Potenza, L.; Basaglia, M.; Fontana, F.; Baldan, E.; Casella, S.; Ryahi, O.; et al. New evidence for nitrogen fixation within the Italian white truffle Tuber magnatum. Fungal Biol. 2010, 114, 936–942. [Google Scholar] [CrossRef]
- Streiblová, E.; Gryndlerová, H.; Gryndler, M. Lanýže: Mykolog-ické téma v evropském kontextu (Truffles: A mycological topic in the European context, in Czech). Mykol. List. 2012, 120, 15–24. [Google Scholar]
- Mello, A.; Lumini, E.; Napoli, C.; Bianciotto, V.; Bonfante, P. Arbuscular mycorrhizal fungal diversity in the Tuber melanosporum brûlé. Fungal Biol. 2015, 119, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Napoli, C.; Mello, A.; Borra, A.; Vizzini, A.; Sourzat, P.; Bonfante, P. Tuber melanosporum, when dominant, affects fungal dynamics in truffle grounds. New Phytol. 2010, 185, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zhou, M.; Zhao, Q.; Wang, F.; Gao, J.; Sheng, H.; An, L. Complete genome sequence of Sphingomonas sp. Cra20, a drought resistant and plant growth promoting rhizobacteria. Genomics 2020, 112, 3648–3657. [Google Scholar] [CrossRef]
- Reichenbach, H. Nannocystis exedens gen. nov., spec. nov., a new myxobacterium of the family Sorangiaceae. Arch. Mikrobiol. 1970, 70, 119–138. [Google Scholar] [CrossRef] [PubMed]
- Taylor, W.J.; Draughon, F.A. Nannocystis exedens: A potential biocompetitive agent against Aspergillus flavus and Aspergillus parasiticus. J. Food Prot. 2001, 64, 1030–1034. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Wang, G.; Li, H.M.; Zhong, J.J.; Tang, Y.J. Volatile organic compounds from a Tuber melanosporum fermentation system. Food Chem. 2012, 135, 2628–2637. [Google Scholar] [CrossRef]
- Effmert, U.; Kalderás, J.; Warnke, R.; Piechulla, B. Volatile Mediated Interactions Between Bacteria and Fungi in the Soil. J. Chem. Ecol. 2012, 38, 665–703. [Google Scholar] [CrossRef]
- Seipke, R.F.; Kaltenpoth, M.; Hutchings, M.I. Streptomyces as symbionts: An emerging and widespread theme? FEMS Microbiol. Rev. 2012, 36, 862–876. [Google Scholar] [CrossRef] [Green Version]
- Tarkka, M.T.; Feldhahn, L.; Buscot, F.; Wubet, T. Genome sequence of the mycorrhiza helper bacterium Streptomyces sp. strain AcH 505. Genome Announc. 2016, 3, 10–11. [Google Scholar] [CrossRef] [Green Version]
- Mabrouk, Y.; Hemissi, I.; Ben Salem, I.; Mejri, S.; Saidi, M.; Belhadj, O. Potential of Rhizobia in Improving Nitrogen Fixation and Yields of Legumes. Symbiosis 2018, 107, 73495. [Google Scholar]
- Li, S.; Hartman, G.L.; Jarvis, B.B.; Tak, H. A Stachybotrys chartarum isolate from soybean. Mycopathologia 2002, 154, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, Y.; Ding, C.; Jia, Z.; He, Z.; Zhang, T.; Wang, X. Declined soil suppressiveness to Fusarium oxysporum by rhizosphere microflora of cotton in soil sickness. Biol. Fertil. Soils 2015, 51, 935–946. [Google Scholar] [CrossRef]
- Luppi Mosca, A.M. The mycoflora of the rhizosphere in truffle sites. IV. Microfungi in the root region of some poplars mycorrhized with Tuber magnatum. [La micoflora della rizosfera nelle tartufaie. IV. Microfunghi da radici di pioppo micorrizate da Tuber magnatum]. Allionia 1973, 19, 29–33. [Google Scholar]
- Pacioni, G.; Leonardi, M.; Aimola, P.; Ragnelli, A.M.; Rubini, A.; Paolocci, F. Isolation and characterization of some mycelia inhabiting Tuber ascomata. Mycol. Res. 2007, 111, 1450–1460. [Google Scholar] [CrossRef] [PubMed]
- Zambonelli, A.; Iotti, M.; Murat, C. True Truffle (Tuber app.) in the World; Springer: Berlin/Heidelberg, Germany, 2016; Volume 47, ISBN 978-3-319-31434-1. [Google Scholar]
- Booth, C. The Genus Fusarium; Commonwealth Mycological Institute: Kew, UK, 1971. [Google Scholar]
- Oliach, D.; Colinas, C.; Castaño, C.; Fischer, C.R.; Bolaño, F.; Bonet, J.A.; Oliva, J. The influence of forest surroundings on the soil fungal community of black truffle (Tuber melanosporum) plantations. For. Ecol. Manag. 2020, 469, 118212. [Google Scholar] [CrossRef]
- Liu, B.; Fischer, C.R.; Bonet, J.A.; Castaño, C.; Colinas, C. Shifts in soil fungal communities in Tuber melanosporum plantations over a 20-year transition from agriculture fields to oak woodlands. For. Syst. 2016, 25, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Bascompte, J.; Jordano, P.; Melián, C.J.; Olesen, J.M. The nested assembly of plant-animal mutualistic networks. Proc. Natl. Acad. Sci. USA 2003, 100, 9383–9387. [Google Scholar] [CrossRef] [Green Version]
- Toju, H.; Kishida, O.; Katayama, N.; Takagi, K. Networks depicting the fine-scale co-occurrences of fungi in soil horizons. PLoS ONE 2016, 11, e0165987. [Google Scholar] [CrossRef] [Green Version]
- Calvaruso, C.; Turpault, M.P.; Leclerc, E.; Ranger, J.; Garbaye, J.; Uroz, S.; Frey-Klett, P. Influence of Forest Trees on the Distribution of Mineral Weathering-Associated Bacterial Communities of the Scleroderma citrinum Mycorrhizosphere. Appl. Environ. Microbiol. 2010, 76, 4780–4787. [Google Scholar] [CrossRef] [Green Version]
- Izumi, H.; Finlay, R.D. Ectomycorrhizal roots select distinctive bacterial and ascomycete communities in Swedish subarctic forest. Environ. Microbiol. 2011, 13, 819–830. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dissimilarity Level Cut | |||
|---|---|---|---|
| Coefficient | Reference | Ward | UPGMA |
| Jaccard | Jaccard (1901) | 0.2 | 0.4 |
| Russell & Rao | Russell and Rao (1940) | 0.1 | 0.2 |
| Simple Matching | Sokal and Michener (1958) | 0.2 | 0.2 |
| Fungi | Bacteria |
|---|---|
| Cryptococcus aerius (Saito) Nann. (10.9) | Escherichia sp. (4.0) |
| Tuber melanosporum (8.9) | Gaiella sp. (2.4) |
| Mortierella sp. (4.1) | Rubrobacter sp. (2.3) |
| Alternaria soliaridae E.G. Simmons (3.3) | Novosphingobium sp. (1.6) |
| Lewia infectoria (Fuckel) M.E. Barr & E.G. Simmons (2.8) | Gemmata sp. (1.4) |
| Fusarium sp. (2.4) | Skermanella sp. (1.4) |
| Tetracladium sp. (1.6) | Blastocatella sp. (1.4) |
| Peyronellaea calorpreferens (Boerema, Gruyter & Noordel) (1.5) | Solirubrobacter sp. (1.3) |
| Preussia sp. (1.0) | Gemmatimonas sp. (1.2) |
| Tuber melosporum (1.0) | Arthrobacter sp. (1.1) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrero de Aza, C.; Armenteros, S.; McDermott, J.; Mauceri, S.; Olaizola, J.; Hernández-Rodríguez, M.; Mediavilla, O. Fungal and Bacterial Communities in Tuber melanosporum Plantations from Northern Spain. Forests 2022, 13, 385. https://doi.org/10.3390/f13030385
Herrero de Aza C, Armenteros S, McDermott J, Mauceri S, Olaizola J, Hernández-Rodríguez M, Mediavilla O. Fungal and Bacterial Communities in Tuber melanosporum Plantations from Northern Spain. Forests. 2022; 13(3):385. https://doi.org/10.3390/f13030385
Chicago/Turabian StyleHerrero de Aza, Celia, Sergio Armenteros, James McDermott, Stefano Mauceri, Jaime Olaizola, María Hernández-Rodríguez, and Olaya Mediavilla. 2022. "Fungal and Bacterial Communities in Tuber melanosporum Plantations from Northern Spain" Forests 13, no. 3: 385. https://doi.org/10.3390/f13030385
APA StyleHerrero de Aza, C., Armenteros, S., McDermott, J., Mauceri, S., Olaizola, J., Hernández-Rodríguez, M., & Mediavilla, O. (2022). Fungal and Bacterial Communities in Tuber melanosporum Plantations from Northern Spain. Forests, 13(3), 385. https://doi.org/10.3390/f13030385