
Forests, Water, and Land Use Change across the Central American Isthmus: Mapping the Evidence Base for Terrestrial Holocene Palaeoenvironmental Proxies

 ,
, 
Abstract
:1. Introduction
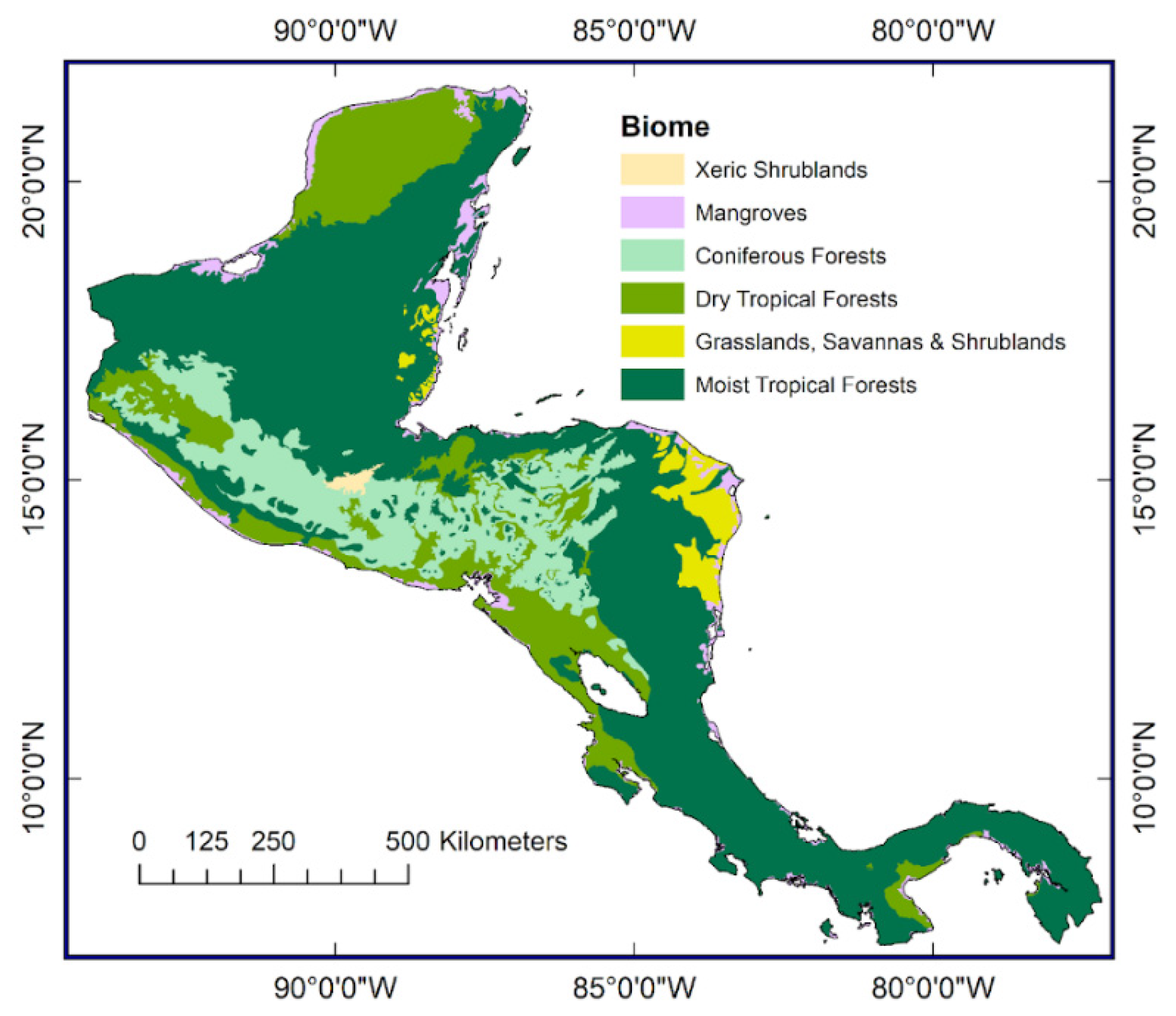
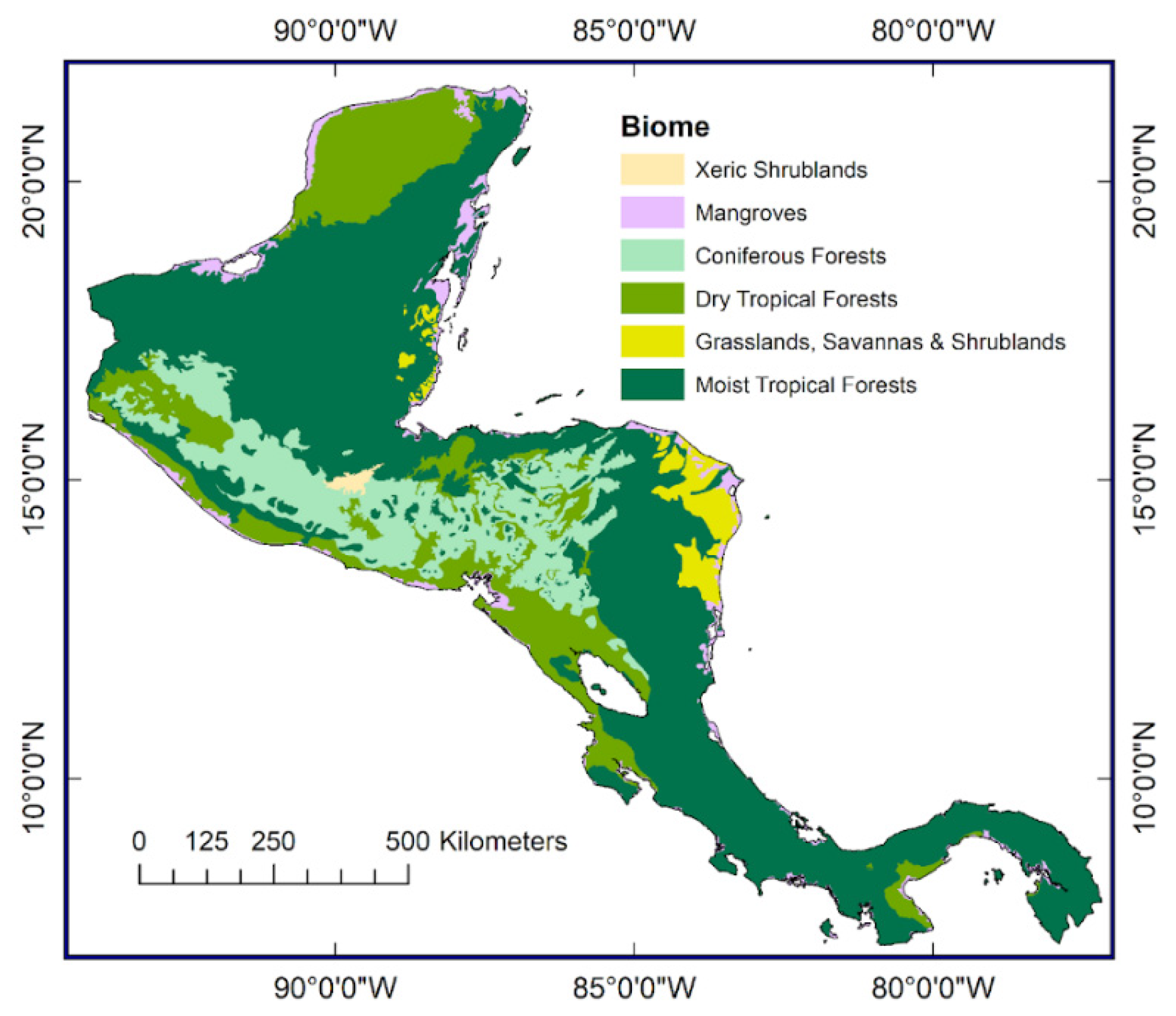
1.1. Forests of the Central American Isthmus
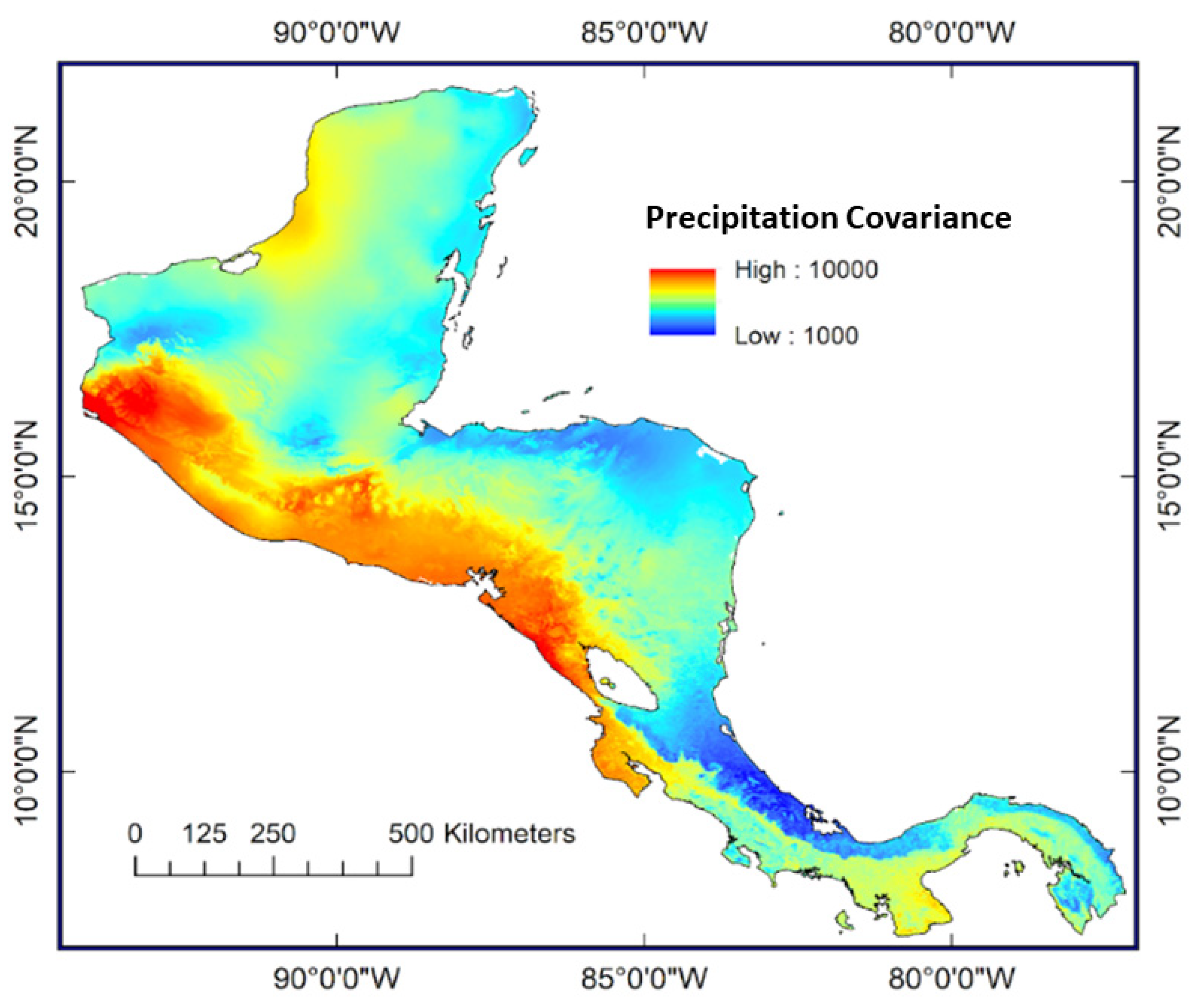
1.2. Water and Climate
1.3. Fire and Burning
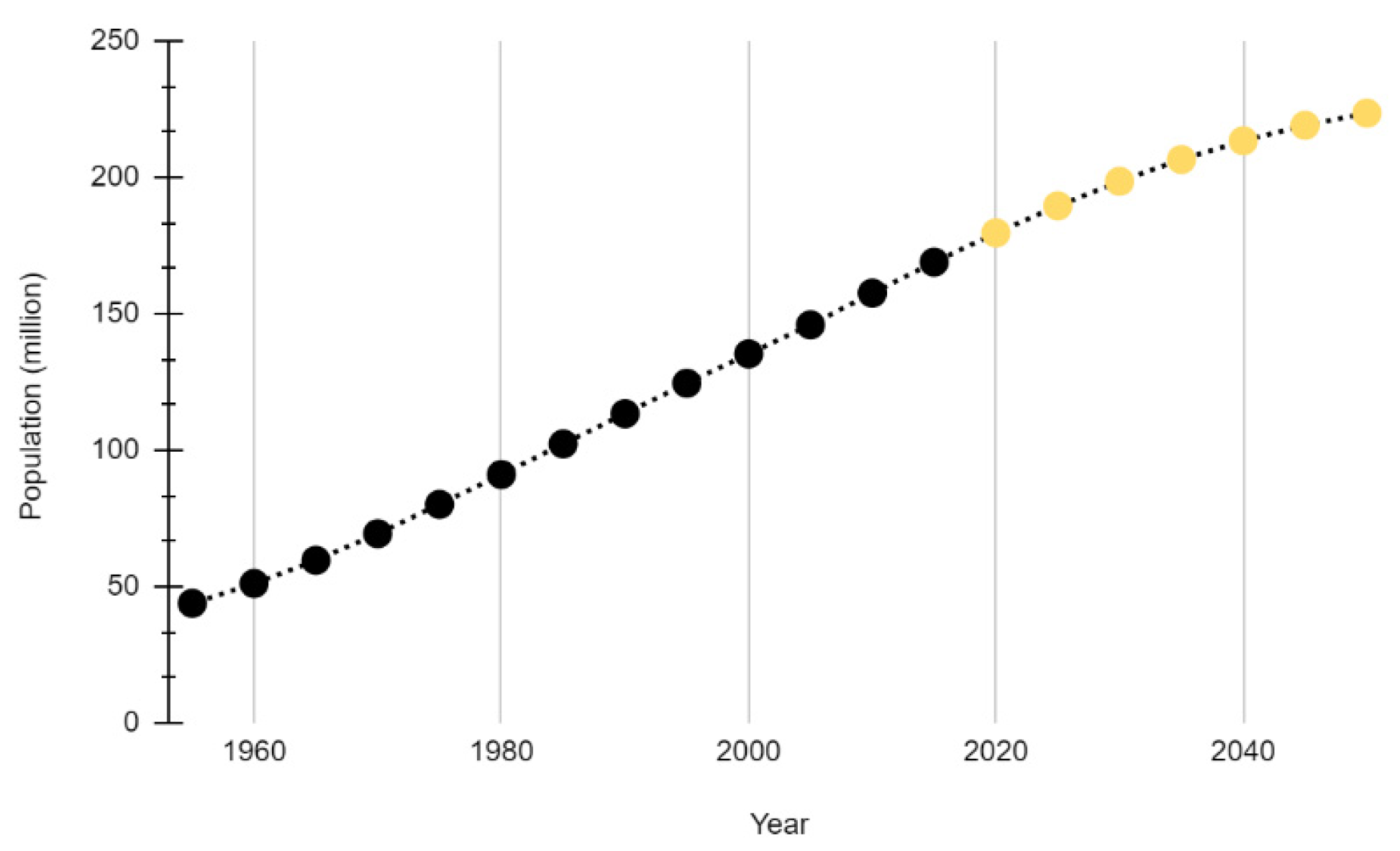
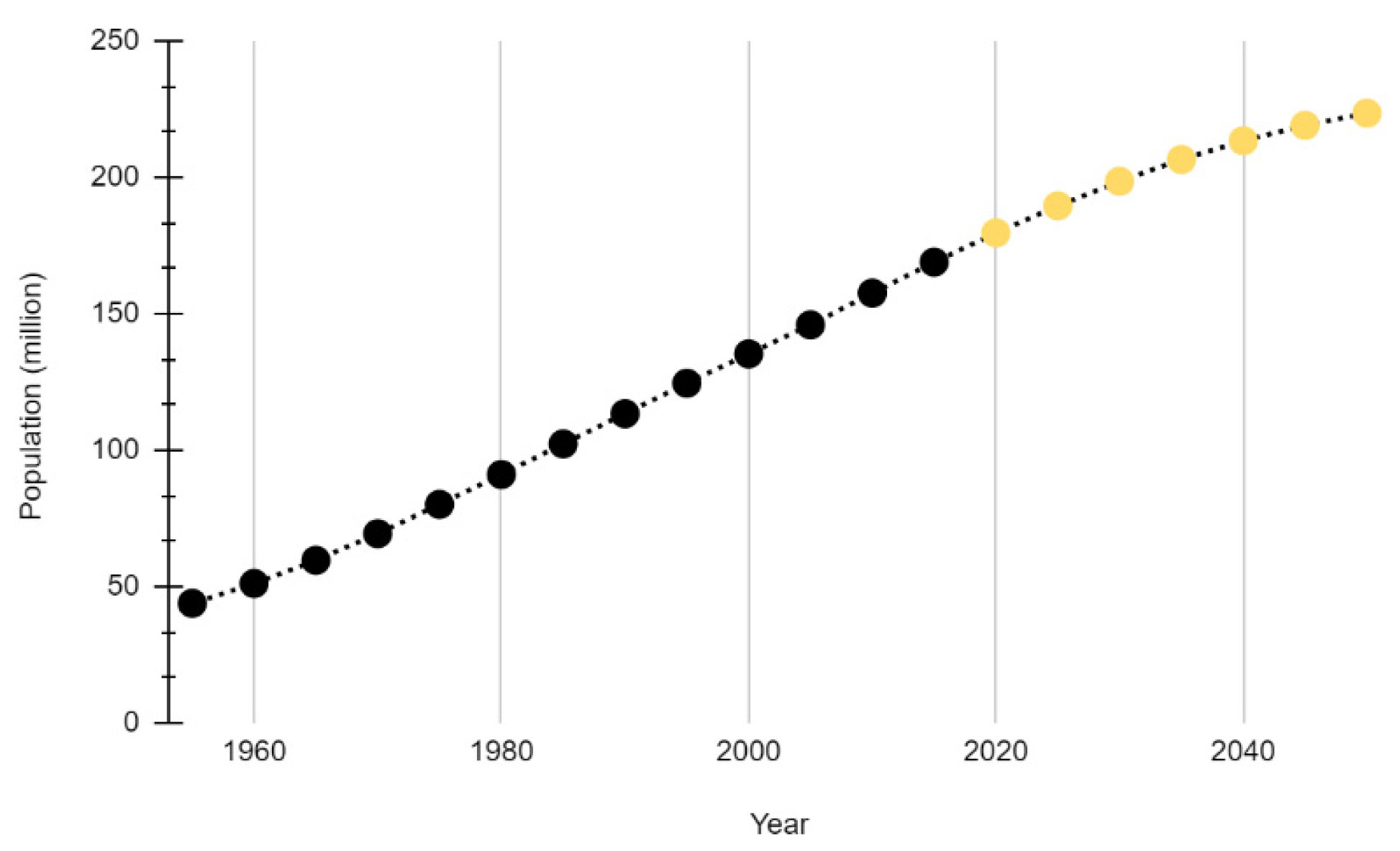
1.4. Population Growth and Land Use Change
1.5. Previous Reviews
1.6. Review Question
2. Materials and Methods
2.1. Systematic Mapping
2.2. PECO Framework
2.3. Literature Search Strategy
2.4. Comprehensiveness of Search
2.5. Inclusion Criteria
2.6. Article Screening
2.7. Article Coding and Data Extraction
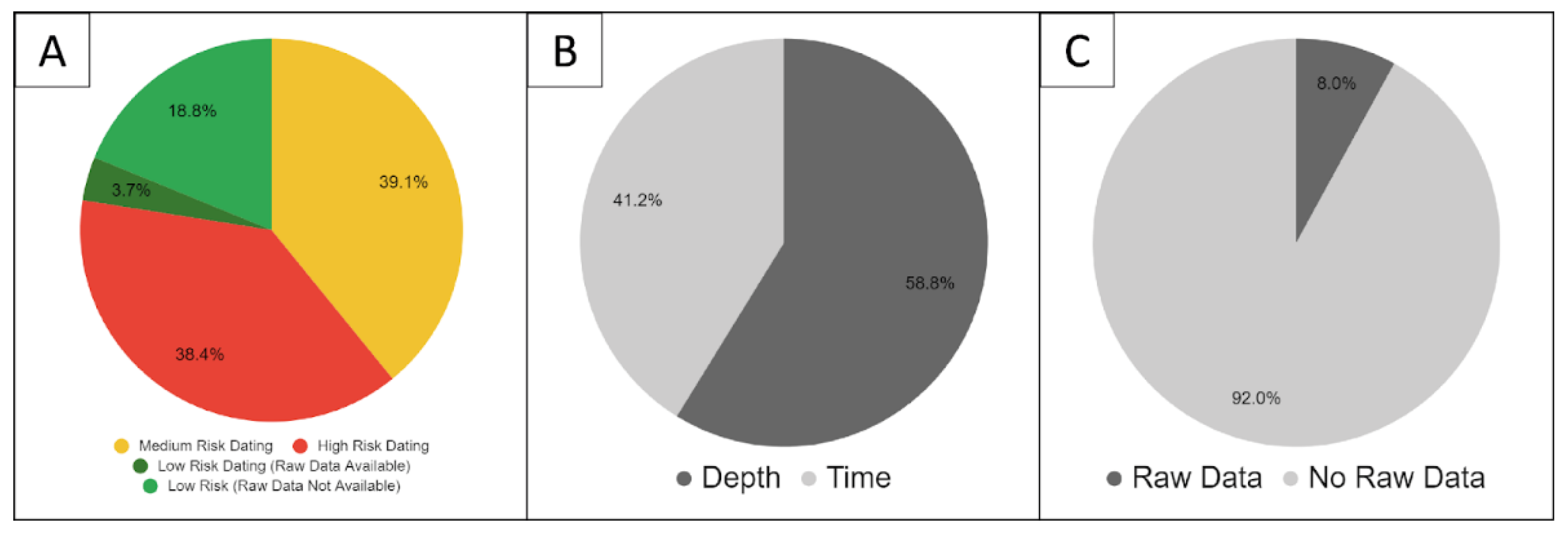
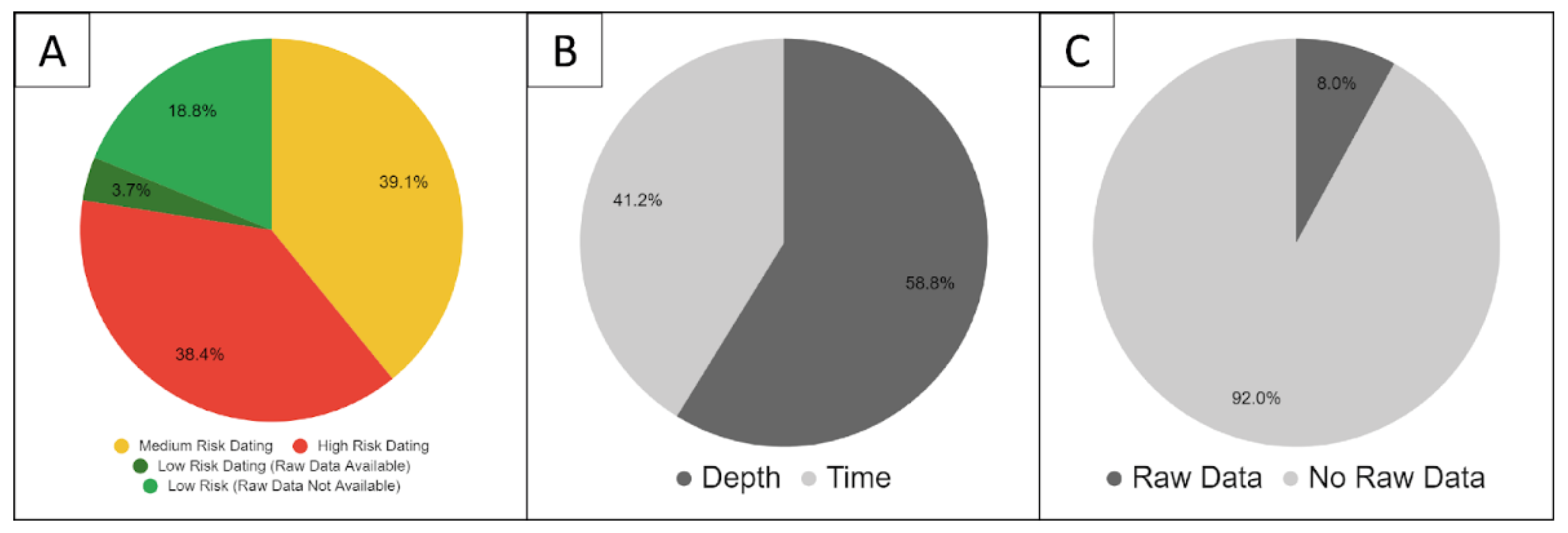
2.8. Study Quality
3. Results
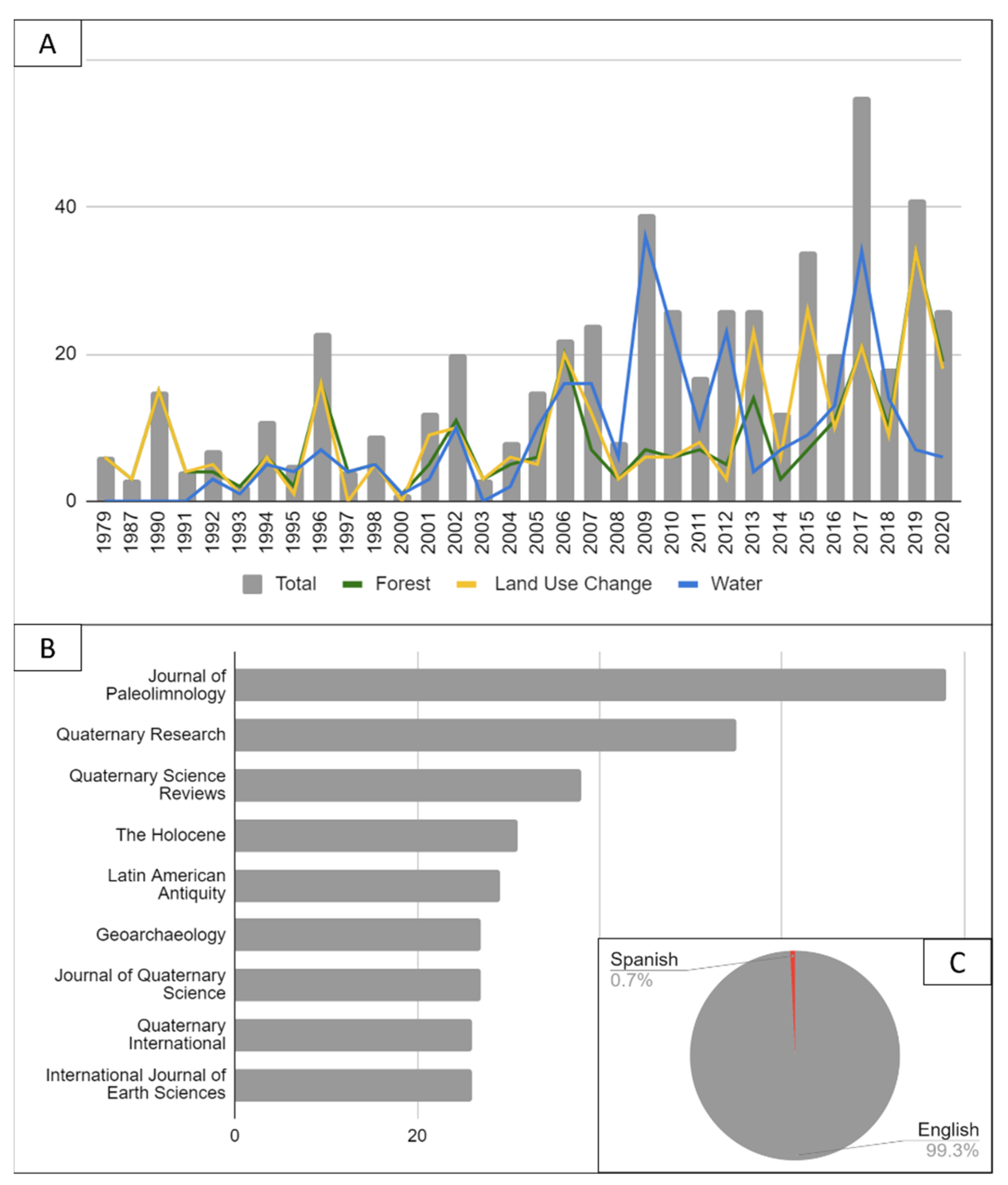
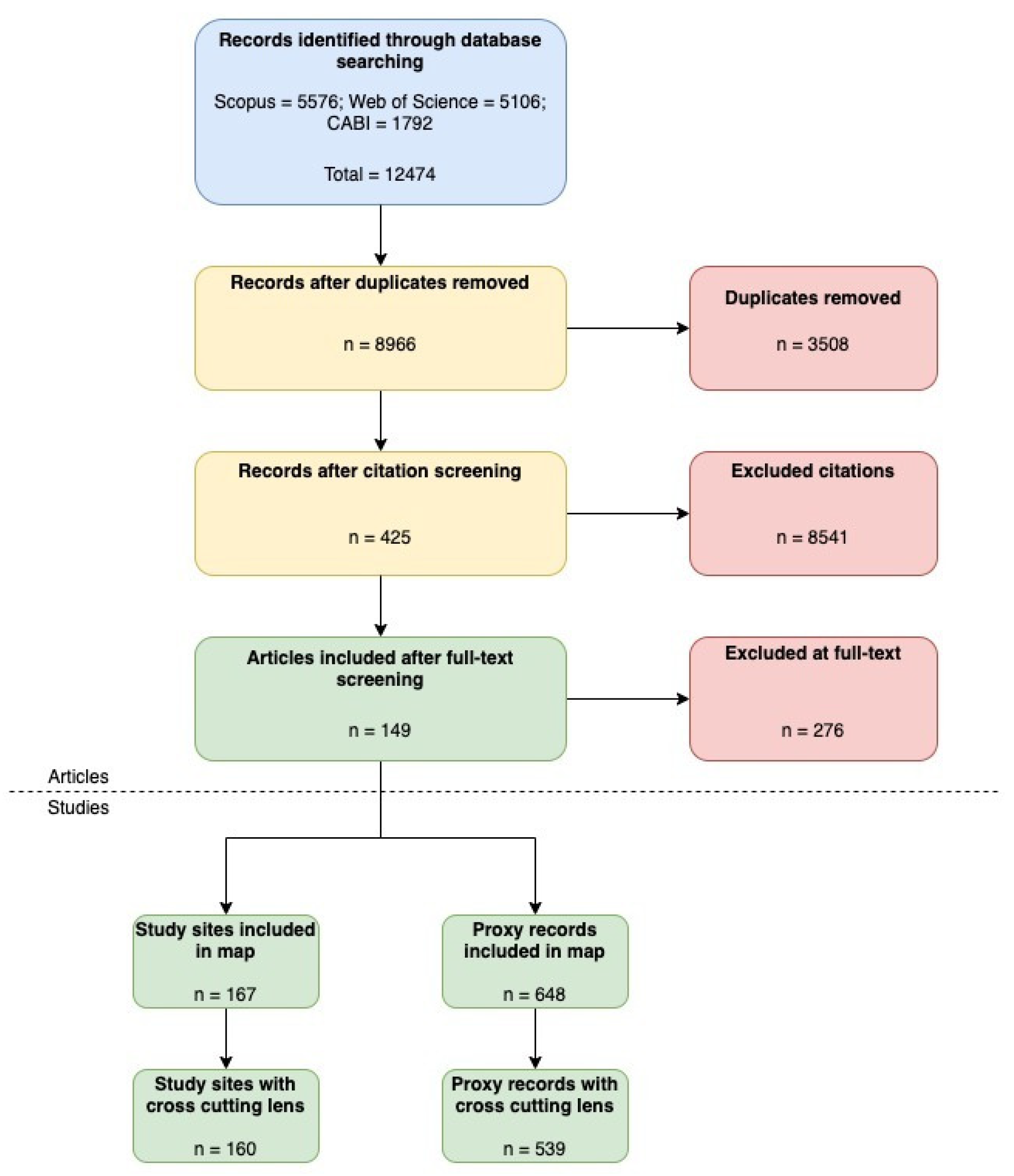
3.1. Selection of Articles
3.2. Interactive Systematic Map
3.3. Source of Studies
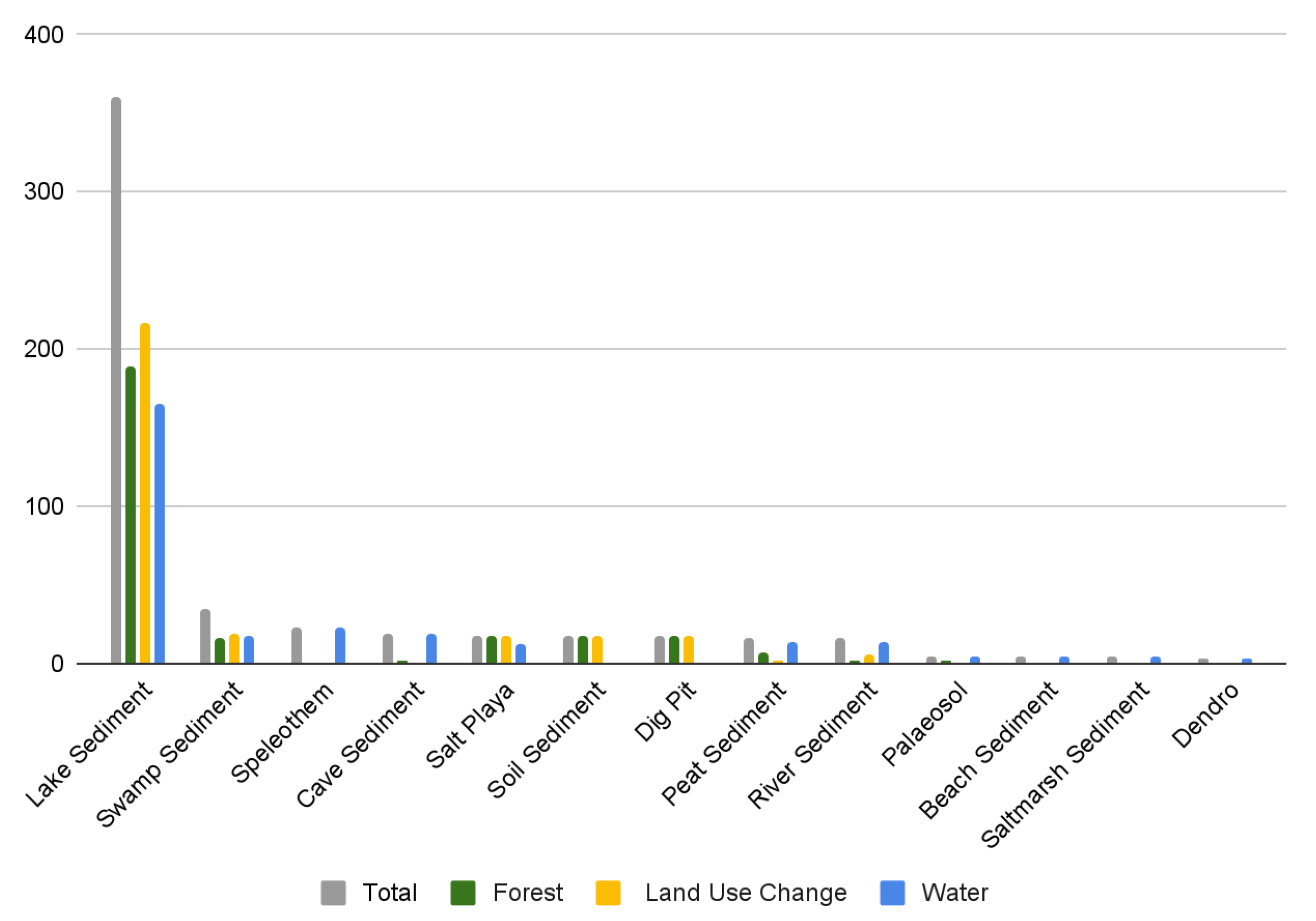
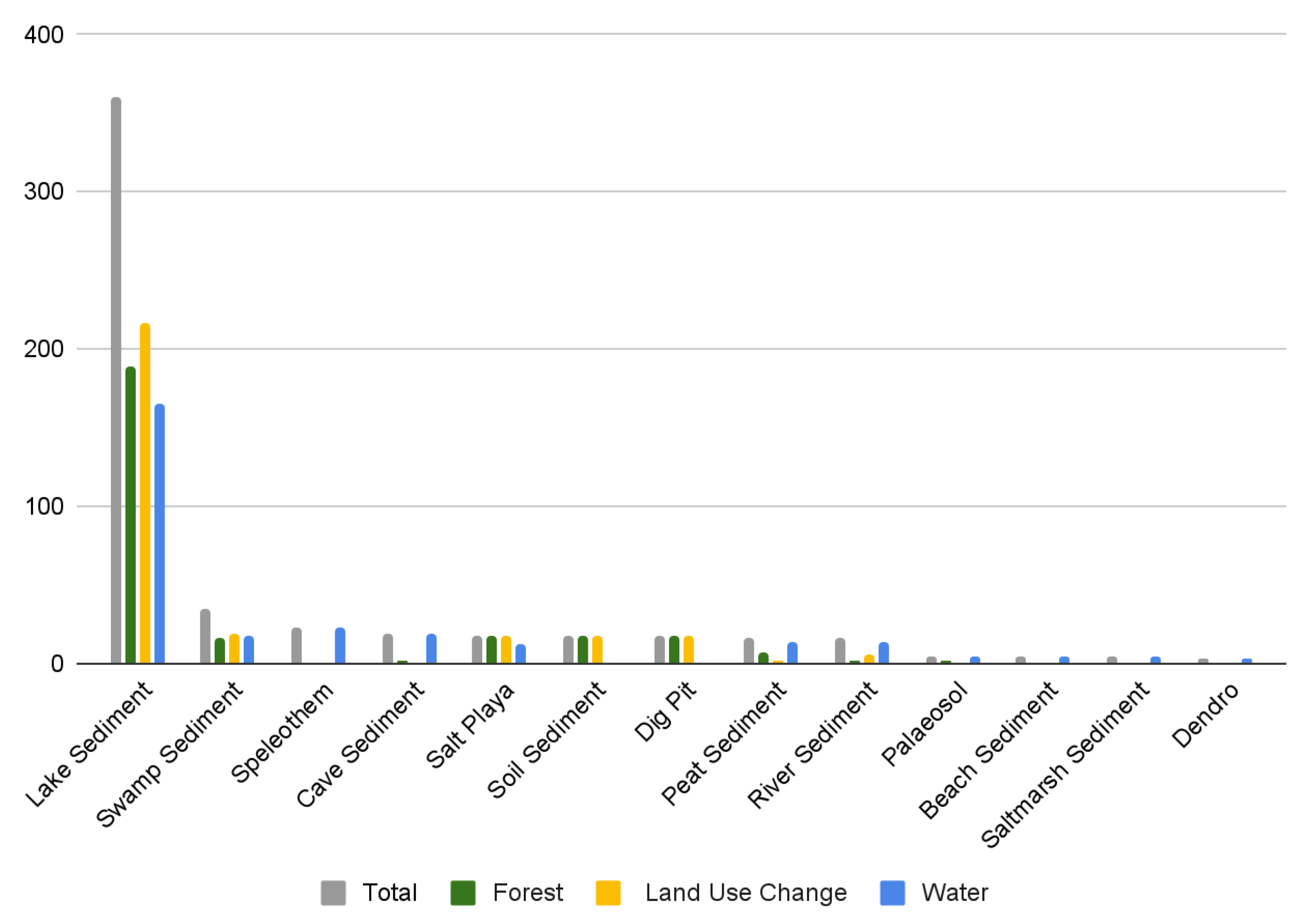
3.4. Type of Environmental Archive
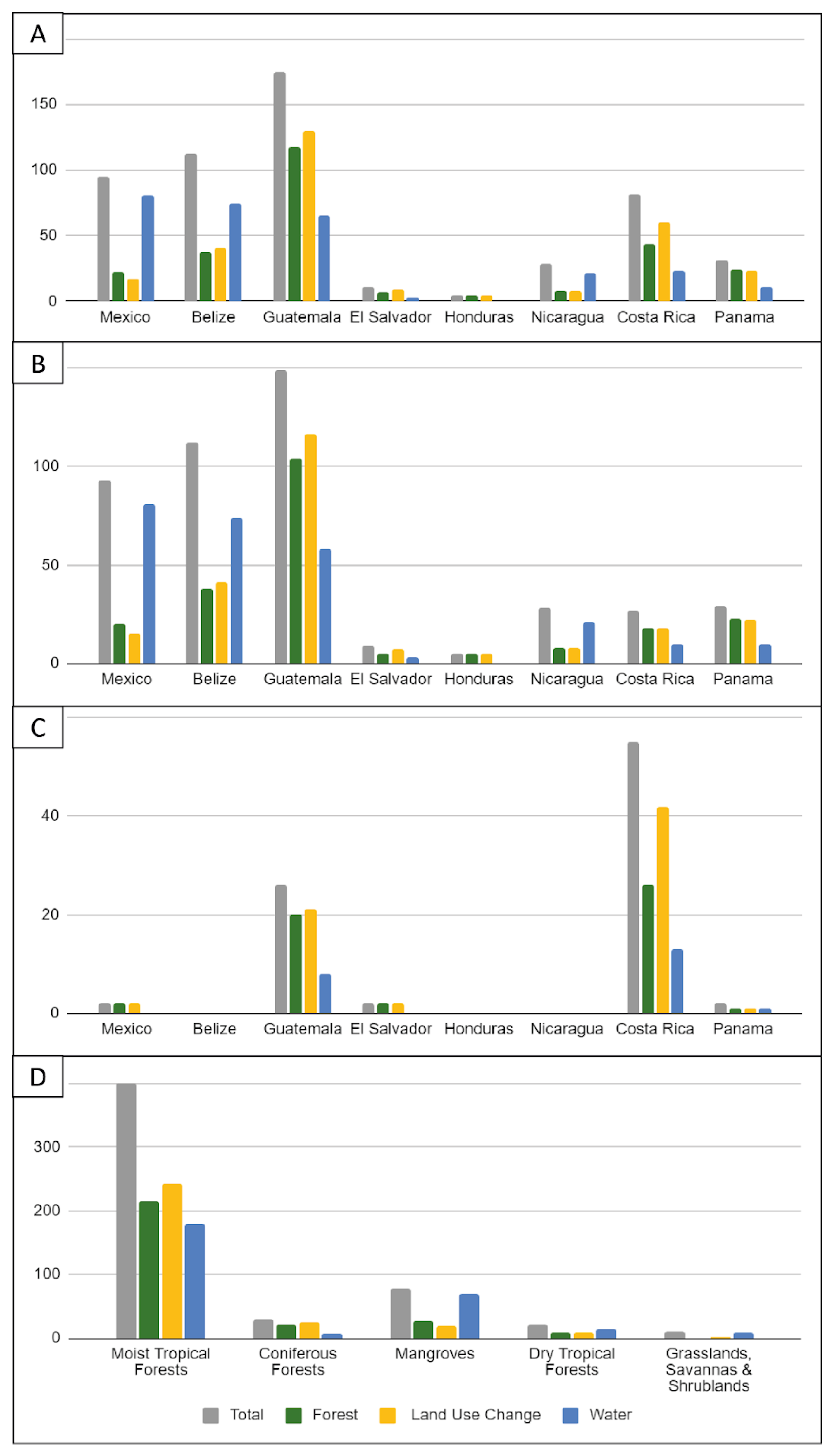
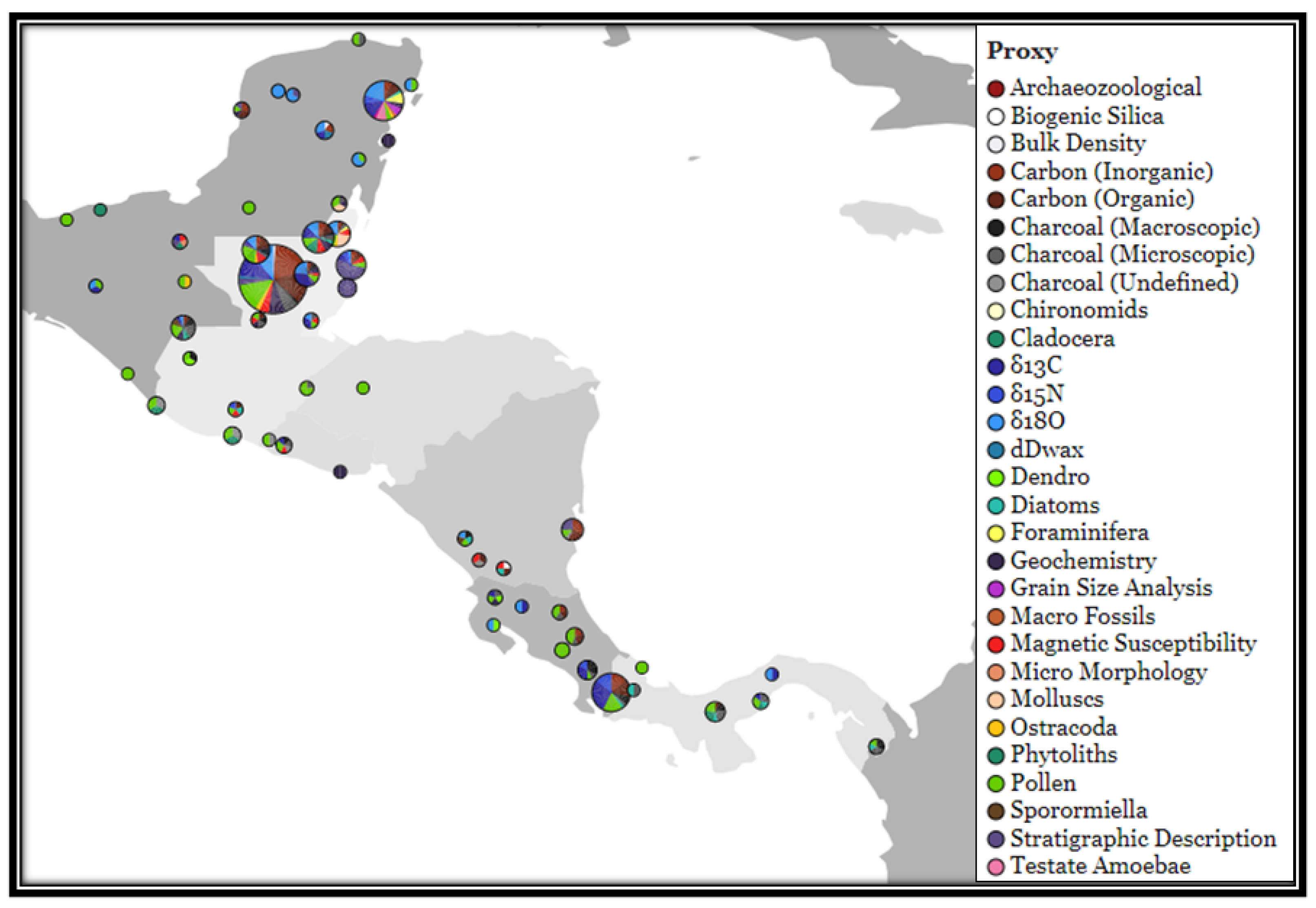
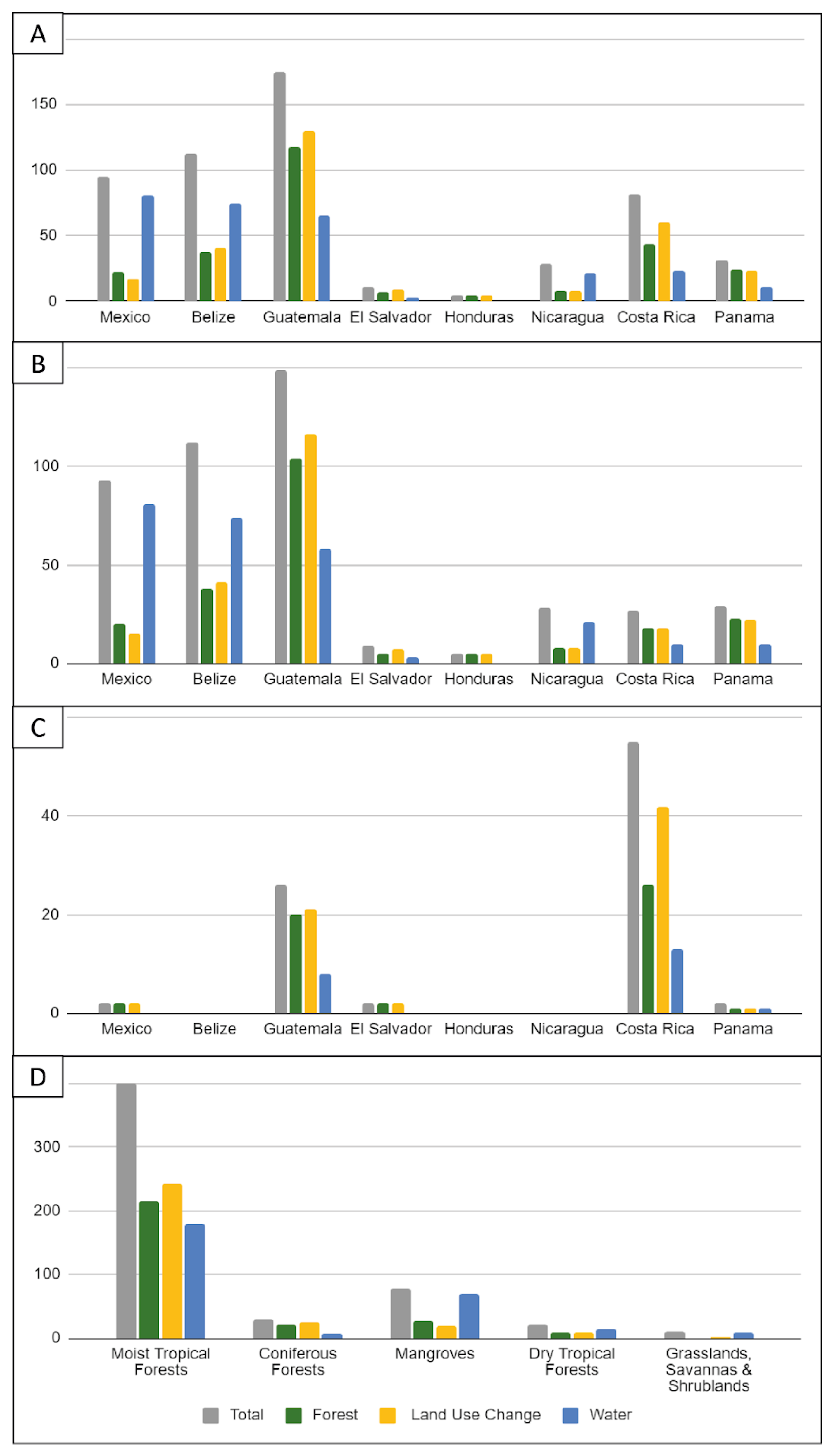
3.5. Location of Studies
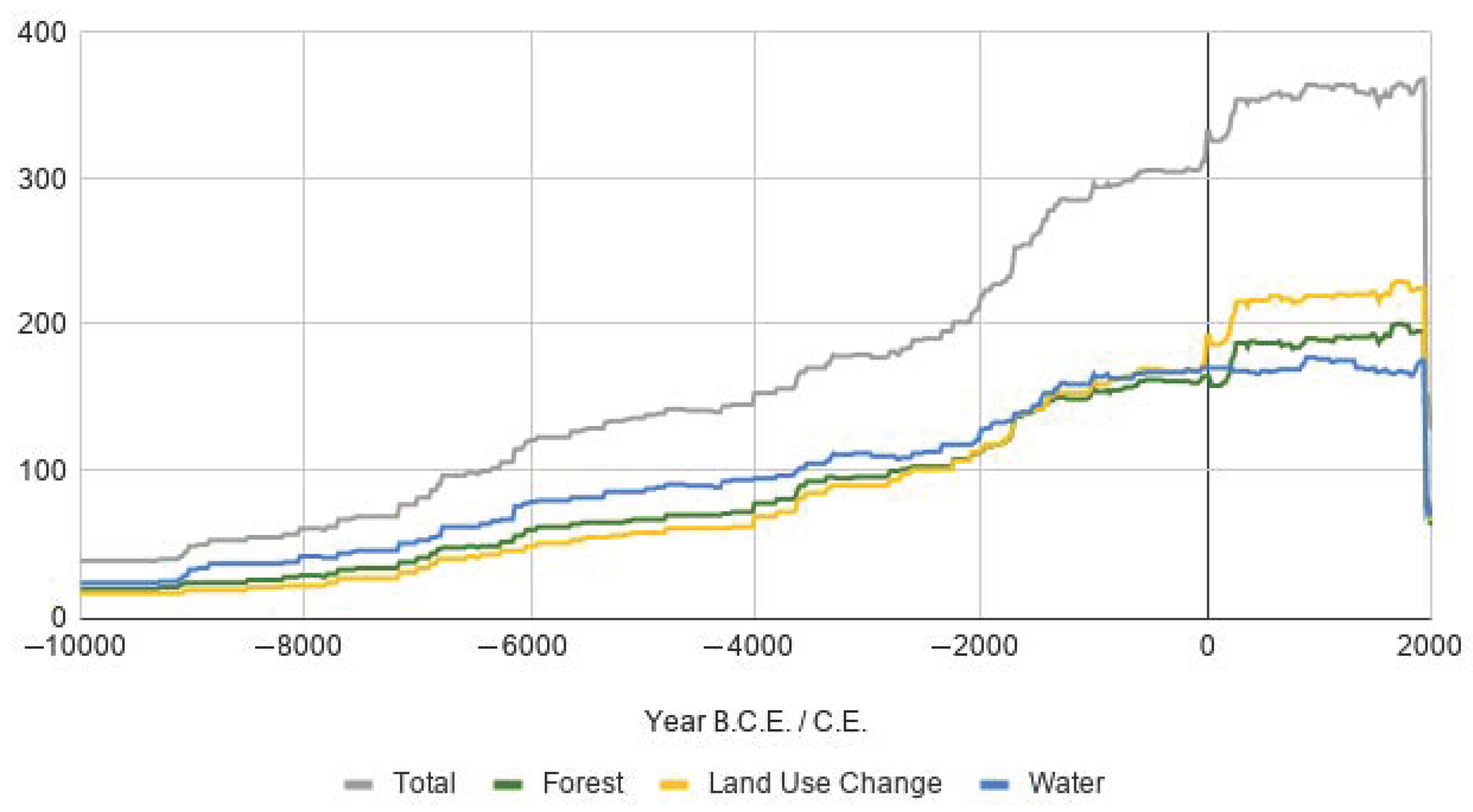
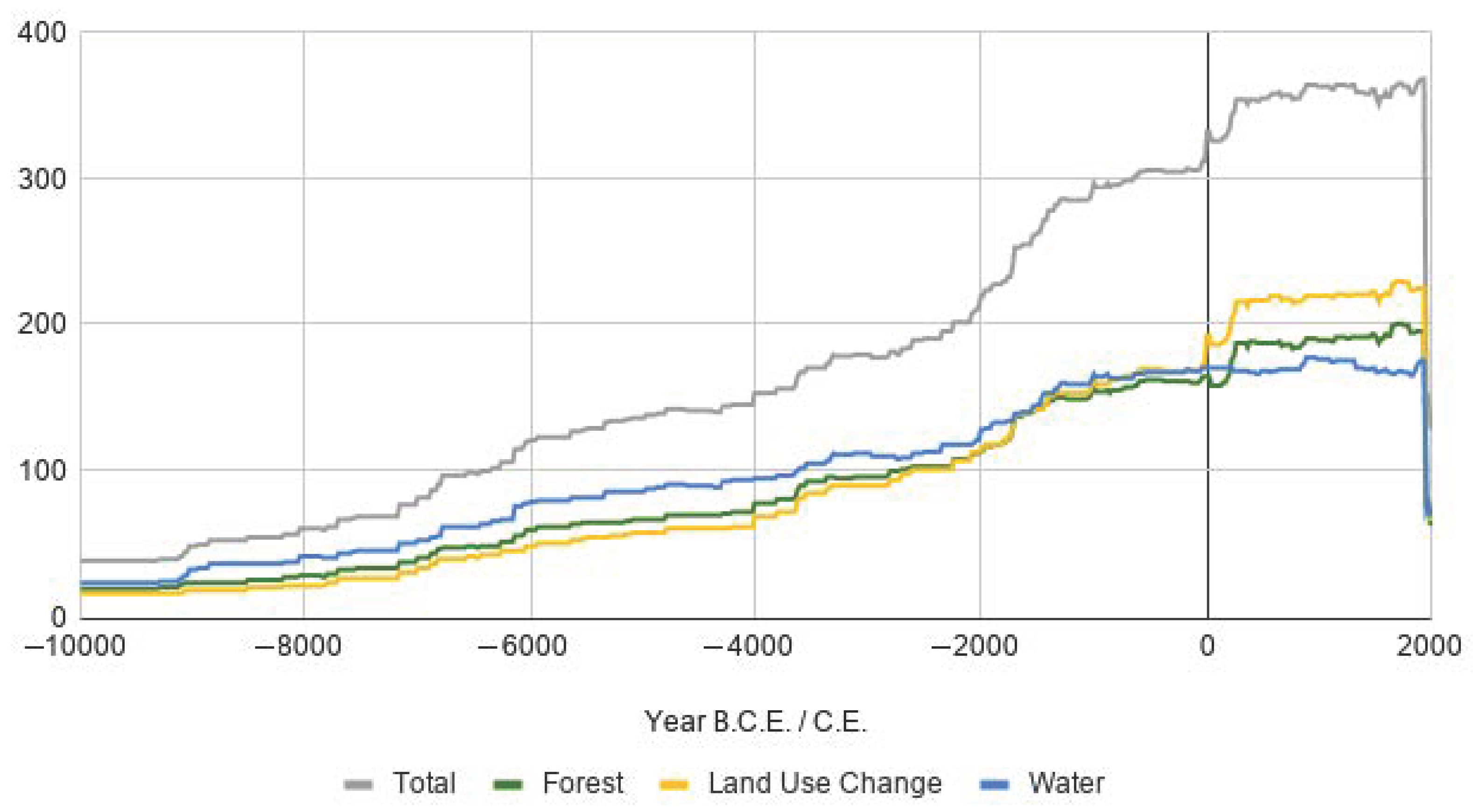
3.6. Temporal Coverage
3.7. Environmental Proxies
3.8. Forest Evidence
3.9. Water Evidence
3.10. Land Use Change Evidence
4. Discussion
4.1. Knowledge Gaps in the Evidence Base
4.2. Future Analyses and Research
4.2.1. Dating Uncertainty and Standardising Reporting
4.2.2. Land Use Change across Space and Time
4.2.3. Dispersal Pathways of Agriculture
4.2.4. The Role and Impacts of Fire and Burning
4.2.5. Changes in Hydro-Climate, Water Availability, and the Risk of Tropical Storms
4.2.6. Forest Resilience and Recovery
4.3. Limitations of This Review
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Search Terms
Appendix B. Test Library
References
- Harvey, C.A.; Komar, O.; Chazdon, R.; Ferguson, B.; Finegan, B.; Griffith, D.; Martínez-Ramos, M.; Morales, H.; Nigh, R.; Soto-Pinto, L.; et al. Integrating Agricultural Landscapes with Biodiversity Conservation in the Mesoamerican Hotspot. Conserv. Biol. 2008, 22, 8–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redo, D.J.; Grau, H.R.; Aide, T.M.; Clark, M.L. Asymmetric forest transition driven by the interaction of soci-oeconomic development and environmental heterogeneity in Central America. Proc. Natl. Acad. Sci. USA 2012, 109, 8839–8844. [Google Scholar] [CrossRef] [Green Version]
- Chiabai, A. Climate Change Impacts on Tropical Forests in Central America: An Ecosystem Service Perspective; Routledge: London, UK, 2015. [Google Scholar]
- IPCC (Intergovernmental Panel on Climate Change). IPCC (2014) Climate Change 2014 Mitigation of Climate Change. Working Group 3 Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (IPCC). Technical Summary and Chapter 6 (Assessing Transformation Pathways). 2014. Available online: https://www.eea.europa.eu/data-and-maps/indicators/atmospheric-greenhouse-gasconcentrations-10/ipcc-2014 (accessed on 26 April 2018).
- DiNezio, P.; Clement, A.; Vecchi, G. Reconciling Differing Views of Tropical Pacific Climate Change. Eos 2010, 91, 141–152. [Google Scholar] [CrossRef] [Green Version]
- World Food Programme. El Nino in Latin America and the Caribbean. 2016. Available online: https://reliefweb.int/sites/reliefweb.int/files/resources/WFP%20El%20Nino%20in%20Latin%20America%20and%20the%20Caribbean%20External%20Situation%20Report%20%231%2030%20May%202016_0.pdf. (accessed on 22 April 2018).
- CI (Conservation International). Biodiversity Hotspots: The Most Remarkable Places on Earth Are also the Most Threatened. CI, Arlington, Virginia. 2007. Available online: http://www.biodiversityhotspots.org (accessed on 26 April 2018).
- Dinerstein, E.; Olson, D.; Joshi, A.; Vynne, C.; Burgess, N.D.; Wikramanayake, E.; Hahn, N.; Palminteri, S.; Hedao, P.; Noss, R.; et al. An Ecoregion-Based Approach to Protecting Half the Terrestrial Realm. Bioscience 2017, 67, 534–545. [Google Scholar] [CrossRef] [PubMed]
- Pither, R.; Kellman, M. Tree species diversity in small, tropical riparian forest fragments in Belize, Central America. Biodivers. Conserv. 2002, 11, 1623–1636. [Google Scholar] [CrossRef]
- Geist, H.J.; Lambin, E.F. What Drives Tropical Deforestation. A Meta-Analysis of Proximate and Underlying Causes of Deforestation Based on Subnational Case Study Evidence; LUCC Report Series, 4; FAO: Rome, Italy, 2001. [Google Scholar]
- Corrales, L.; Bouroncle, C.; Zamora, J.C. An overview of forest biomes and ecoregions of Central America. In Climate Change Impacts on Tropical Forests in Central America; Routledge: London, UK, 2015; pp. 17–38. [Google Scholar]
- Janzen, D.H. Management of Habitat Fragments in a Tropical Dry Forest: Growth. Ann. Mo. Bot. Gard. 1988, 75, 105. [Google Scholar] [CrossRef]
- Sabogal, C. Regeneration of tropical dry forests in Central America, with examples from Nicaragua. J. Veg. Sci. 1992, 3, 407–416. [Google Scholar] [CrossRef]
- Griscom, H.P.; Ashton, M.S. Restoration of dry tropical forests in Central America: A review of pattern and process. For. Ecol. Manag. 2011, 261, 1564–1579. [Google Scholar] [CrossRef]
- Gillespie, T.W.; Grijalva, A.; Farris, C.N. Diversity, composition, and structure of tropical dry forests in Central America. Plan Ecol. 2000, 147, 37–47. [Google Scholar] [CrossRef]
- Muller, C.H. The Central American Species of Quercus (No. 477); US Government Printing Office: Washington, DC, USA, 1942.
- Kappelle, M. Neotropical montane oak forests: Overview and outlook. In Ecology and Conservation of Neotropical Montane Oak Forests; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Rzedowski, J. Vegetación de México, 1st ed.; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: México City, Mexico, 2006; p. 504. [Google Scholar]
- Veblen, T.T. Forest Preservation in the Western Highlands of Guatemala. Geogr. Rev. 1978, 68, 417. [Google Scholar] [CrossRef]
- Parsons, J.J. The miskito pine savanna of nicaragua and honduras. Ann. Assoc. Am. Geogr. 1955, 45, 36–63. [Google Scholar] [CrossRef]
- Nájera Acevedo, A. The conservation of the thorn scrub and dry forest of the Motagua Valley, Guatemala: Promoting the protection of a unique ecoregion. Lyonia 2006, 9, 7–19. [Google Scholar]
- Young, C.A. Belize’s ecosystems: Threats and challenges to conservation in Belize. Trop. Conserv. Sci. 2008, 1, 18–33. [Google Scholar] [CrossRef] [Green Version]
- Alfaro, E.J. Some characteristics of the precipitation annual cycle in Central America and their relationships with its surrounding tropical oceans. Top. Meteor. Oceanog. 2000, 7, 99–115. [Google Scholar]
- Metcalfe, S.E.; Barron, J.A.; Davies, S.J. The Holocene history of the North American Monsoon: ‘known knowns’ and ‘known unknowns’ in understanding its spatial and temporal complexity. Quat. Sci. Rev. 2015, 120, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Giannini, A.; Chiang, J.C.; Cane, M.A.; Kushnir, Y.; Seager, R. The ENSO teleconnection to the Tropical Atlantic Ocean: Contributions of the remote and local SSTs to rainfall variability in the tropical Americas. J. Clim. 2001, 14, 4530–4544. [Google Scholar] [CrossRef]
- Enfield, D.B.; Alfaro, E.J. The dependence of Caribbean rainfall on the interaction of the tropical Atlantic and Pacific Oceans. J. Clim. 1999, 12, 2093–2103. [Google Scholar] [CrossRef] [Green Version]
- GardaWorld. Nicaragua: Major Wildfire in Indio Maíz Biological Reserve. 2018. Available online: https://www.garda.com/crisis24/news-alerts/108451/nicaragua-major-wildfire-inindio-maiz-biological-reserve-update-1 (accessed on 22 April 2018).
- Harvey, W.J.; Stansell, N.; Nogué, S.; Willis, K.J. The Apparent Resilience of the Dry Tropical Forests of the Nicaraguan Region of the Central American Dry Corridor to Variations in Climate Over the Last C. 1200 Years. Quaternary 2019, 2, 25. [Google Scholar] [CrossRef] [Green Version]
- Sparks, A.; Hengl, T.; Nelson, A. Global Surface Summary of the Day—GSOD. 2017. Available online: https://data.noaa.gov/dataset/dataset/global-surface-summary-of-the-daygsod (accessed on 13 March 2017).
- Bowman, D.M.J.S.; Balch, J.; Artaxo, P.; Bond, W.; Cochrane, M.A.; D’Antonio, C.M.; DeFries, R.; Johnston, F.H.; Keeley, J.E.; Krawchuk, M.A.; et al. The human dimension of fire regimes on Earth. J. Biogeogr. 2011, 38, 2223–2236. [Google Scholar] [CrossRef] [Green Version]
- Anderson, L.; Wahl, D. Two Holocene paleofire records from Peten, Guatemala: Implications for natural fire regime and prehispanic Maya land use. Glob. Planet. Chang. 2016, 138, 82–92. [Google Scholar] [CrossRef]
- Murphy, P.G.; Lugo, A. Ecology of Tropical Dry Forest. Annu. Rev. Ecol. Syst. 1986, 17, 67–88. [Google Scholar] [CrossRef]
- Urquhart, G.R. Paleoecological record of hurricane disturbance and forest regeneration in Nicaragua. Quat. Int. 2009, 195, 88–97. [Google Scholar] [CrossRef] [Green Version]
- Cochrane, M.A.; Alencar, A.; Schulze, M.D.; Souza, C.M.; Nepstad, D.C.; Lefebvre, P.; Davidson, E.A. Positive feedbacks in the fire dynamic of closed canopy tropical forests. Science 1999, 284, 1832–1835. [Google Scholar] [CrossRef]
- Scott, A.C.; Bowman, D.M.; Bond, W.J.; Pyne, S.J.; Alexander, M.E. Fire on earth: An introduction; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Pyne, S.J. Fire management in the United States. In Introduction to Wildland Fire; John Wiley & Sons: Hoboken, NJ, USA, 1984. [Google Scholar]
- Uhl, C.; Kauffman, J.B. Deforestation, Fire Susceptibility, and Potential Tree Responses to Fire in the Eastern Amazon. Ecology 1990, 71, 437–449. [Google Scholar] [CrossRef]
- Holdsworth, A.R.; Uhl, C. Fire in Amazonian selectively logged rain forest and the potential for fire reduction. Ecol. Appl. 1997, 7, 713–725. [Google Scholar] [CrossRef]
- Rothermel, R.C. How to Predict the Spread and Intensity of Forest and Range Fires; USDA Forest Service General Technical Report INT; Intermountain Forest and Range Experiment Station: Washington, DC, USA, 1983.
- Peterson, D.L.; Ryan, K.C. Modeling postfire conifer mortality for long-range planning. Environ. Manag. 1986, 10, 797–808. [Google Scholar] [CrossRef]
- Joaquin-Chamorro. Lessons Learned from Indio Maíz. La Prensa. 2018. Available online: https://www.theguardian.com/world/2018/apr/11/nicaragua-rainforest-fire-costa-rica (accessed on 25 April 2018).
- The Guardian. Nicaragua Fires: Aid from Costa Rica Rejected as Blaze Destroys Rainforest. 2018. Available online: https://www.theguardian.com/world/2018/apr/11/nicaraguarainforest-fire-costa-rica (accessed on 25 April 2018).
- Carr, D. Population and deforestation: Why rural migration matters. Prog. Hum. Geogr. 2008, 33, 355–378. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT (Food and Agriculture Organisation of the United Nations). Food and Agriculture Data. 2011. Available online: http://faostat.fao.org/default.aspx (accessed on 12 May 2021).
- Worldometers. Central America Population. 2021. Available online: http://www.worldometers.info/world-population/central-america-population/ (accessed on 23 May 2021).
- Bilsborrow, R.E.; Carr, D.L. Tradeoffs or Synergies? Agricultural Intensification, Economic Development and the Environment; Lee, D.R., Barrett, C.B., Eds.; CABI Publishing: Wallingford, UK, 2001; pp. 35–55. [Google Scholar]
- Rueda, X. Understanding deforestation in the southern Yucatán: Insights from a sub-regional, multi-temporal analysis. Reg. Environ. Chang. 2010, 10, 175–189. [Google Scholar] [CrossRef]
- Bray, D.B.; Duran, E.; Ramos, V.H.; Mas, J.F.; Velazquez, A.; McNab, R.B.; Barry, D.; Radachowsky, J. Tropical Deforestation, Community Forests, and Protected Areas in the Maya Forest. Ecol. Soc. 2008, 13, 56. [Google Scholar] [CrossRef]
- White, C.D. Reconstructing Ancient Maya Diet; University of Utah Press: Salt Lake City, UT, USA, 1999. [Google Scholar]
- Dull, R.A. An 8000-year record of vegetation, climate, and human disturbance from the Sierra de Apaneca, El Salvador. Quat. Res. 2004, 61, 159–167. [Google Scholar] [CrossRef]
- Franco-Gaviria, F.; Correa-Metrio, A.; Cordero-Oviedo, C.; López-Pérez, M.; Cárdenes-Sandí, G.; Romero, F. Effects of late Holocene climate variability and anthropogenic stressors on the vegetation of the Maya highlands. Quat. Sci. Rev. 2018, 189, 76–90. [Google Scholar] [CrossRef]
- Dull, R.A. A Holocene record of Neotropical savanna dynamics from El Salvador. J. Paleolimnol. 2004, 32, 219–231. [Google Scholar] [CrossRef]
- Dull, R.A. Evidence for Forest Clearance, Agriculture, and Human-Induced Erosion in Precolumbian El Salvador. Ann. Assoc. Am. Geogr. 2007, 97, 127–141. [Google Scholar] [CrossRef]
- Velez, M.I.; Curtis, J.H.; Brenner, M.; Escobar, J.; Leyden, B.W.; De Hatch, M.P. Environmental and cultural changes in highland Guatemala inferred from Lake Amatitlán sediments. Geoarchaeology 2011, 26, 346–364. [Google Scholar] [CrossRef]
- Harvey, W.J.; Nogué, S.; Stansell, N.; Petrokofsky, G.; Steinman, B.; Willis, K.J. The Legacy of Pre-Columbian Fire on the Pine–Oak Forests of Upland Guatemala. Front. For. Glob. Chang. 2019, 2, 34. [Google Scholar] [CrossRef] [Green Version]
- Harvey, W.J.; Nogué, S.; Stansell, N.; Adolf, C.; Long, P.R.; Willis, K. A palynological perspective on the im-pacts of European contact: Historic deforestation, ranching and agriculture surrounding the Cuchumatanes Highlands, Guatemala. Veg. Hist. Archaeobotany 2021, 30, 395–408. [Google Scholar] [CrossRef]
- Dull, R.A. The Maize Revolution A View from El Salvador. In Histories of Maize in Mesoamerica: Multidisciplinary Approaches; Left Coast Press: Walnut Creek, CA, USA, 2016. [Google Scholar]
- Anchukaitis, K.J.; Horn, S.P. A 2000-year reconstruction of forest disturbance from southern Pacific Costa Rica. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 221, 35–54. [Google Scholar] [CrossRef]
- Schüpbach, S.; Kirchgeorg, T.; Colombaroli, D.; Beffa, G.; Radaelli, M.; Kehrwald, N.M.; Barbante, C. Combining charcoal sediment and molecular markers to infer a Holocene fire history in the Maya Lowlands of Petén, Guatemala. Quat. Sci. Rev. 2015, 115, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Piperno, D.R.; Bush, M.; Colinvaux, P.A. Paleoenvironments and Human Occupation in Late-Glacial Panama. Quat. Res. 1990, 33, 108–116. [Google Scholar] [CrossRef]
- Piperno, D.R.; Bush, M.B.; Colinvuax, P.A. Paleoecological perspectives on human adaptation in central Panama. II the Holocene. Geoarchaeology 1991, 6, 227–250. [Google Scholar] [CrossRef]
- Neff, H.; Pearsall, D.M.; Jones, J.G.; Arroyo, B.; Collins, S.K.; Freidel, D.E. Early Maya Adaptive Patterns: Mid-Late Holocene Paleoenvironmental Evidence from Pacific Guatemala. Lat. Am. Antiq. 2006, 17, 287–315. [Google Scholar] [CrossRef]
- Ford, A.; Nigh, R. Origins of the Maya Forest Garden: Maya Resource Management. J. Ethnobiol. 2009, 29, 213–236. [Google Scholar] [CrossRef]
- Piperno, D.R. Quaternary environmental history and agricultural impact on vegetation in Central America. Ann. Mo. Bot. Gard. 2006, 93, 274–296. [Google Scholar] [CrossRef]
- Islebe, G.A.; Torrescano-Valle, N.; Aragón-Moreno, A.A.; Vela-Peláez, A.A.; Valdez-Hernández, M. The Paleoanthropocene of the Yucatán Peninsula: Palynological evidence of environmental change. Boletín Soc. Geológica Mex. 2018, 70, 49–60. [Google Scholar] [CrossRef]
- Metcalfe, S.E.; O’Hara, S.L.; Caballero, M.; Davies, S.J. Records of Late Pleistocene–Holocene climatic change in Mexico—A review. Quat. Sci. Rev. 2000, 19, 699–721. [Google Scholar] [CrossRef]
- Beach, T.; Luzzadder-Beach, S.; Cook, D.; Dunning, N.; Kennett, D.J.; Krause, S.; Terry, R.; Trein, D.; Valdez, F. Ancient Maya impacts on the Earth’s surface: An Early Anthropocene analog? Quat. Sci. Rev. 2015, 124, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Douglas, P.M.; Demarest, A.A.; Brenner, M.; Canuto, M.A. Impacts of climate change on the collapse of low-land Maya civilization. Annu. Rev. Earth Planet. Sci. 2016, 44, 613–645. [Google Scholar] [CrossRef]
- Haug, G.H.; Hughen, K.A.; Sigman, D.M.; Peterson, L.C.; Röhl, U. Southward Migration of the Intertropical Convergence Zone Through the Holocene. Science 2001, 293, 1304–1308. [Google Scholar] [CrossRef]
- League, B.L.; Horn, S.P. A 10,000 year record of Paramo fires in Costa Rica. J. Trop. Ecol. 2000, 16, 747–752. [Google Scholar] [CrossRef]
- Lane, C.S.; Horn, S.P.; Mora, C.I.; Orvis, K.H.; Finkelstein, D.B. Sedimentary stable carbon isotope evidence of late Quaternary vegetation and climate change in highland Costa Rica. J. Paleolimnol. 2011, 45, 323–338. [Google Scholar] [CrossRef]
- Correa-Metrio, A.; Bush, M.; Cabrera, K.R.; Sully, S.; Brenner, M.; Hodell, D.; Escobar, J.; Guilderson, T. Rapid climate change and no-analog vegetation in lowland Central America during the last 86,000 years. Quat. Sci. Rev. 2012, 38, 63–75. [Google Scholar] [CrossRef]
- CEE (Collaboration for Environmental Evidence). Guidelines and Standards for Evidence synthesis in Environmental Management; Pullin, A.S., Frampton, G.K., Livoreil, B., Petrokofsky, G., Eds.; Version 5.0; 2018; Available online: www.environmentalevidence.org/information-for-authors (accessed on 24 May 2021).
- Haddaway, N.R.; Bethel, A.; Dicks, L.; Koricheva, J.; Macura, B.; Petrokofsky, G.; Pullin, A.S.; Savilaakso, S.; Stewart, G.B. Eight problems with literature reviews and how to fix them. Nat. Ecol. Evol. 2020, 4, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Livoreil, B.; Glanville, J.; Haddaway, N.R.; Bayliss, H.; Bethel, A.; De Lachapelle, F.F.; Robalino, S.; Savilaakso, S.; Zhou, W.; Petrokofsky, G.; et al. Systematic searching for environmental evidence using multiple tools and sources. Environ. Évid. 2017, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Horn, S.P.; Rodgers, J.C., III; Orvis, K.H.; Northrop, L.A. Recent land use and vegetation history from soil pollen analysis: Testing the potential in the lowland humid tropics. Palynology 1998, 22, 167–180. [Google Scholar] [CrossRef]
- Frampton, G.K.; Livoreil, B.; Petrokofsky, G. Eligibility screening in evidence synthesis of environmental management topics. Environ. Evid. 2017, 6, 27. [Google Scholar] [CrossRef] [Green Version]
- Wallace, B. Abstrackr. 2018. Available online: http://abstrackr.cebm.brown.edu/ (accessed on 9 September 2017).
- Altman, D.G. Measuring agreement. In Practical Statistics for Medical Research; Altman, D.G., Ed.; Chapman and Hall: London, UK, 1991. [Google Scholar]
- Martin, A.C. Thalloo Evidence-Mapping: A Jekyll Theme. Available online: https://github.com/AndrewIOM/thalloo (accessed on 1 October 2018).
- Roth, N.; Jaramillo, F.; Wang-Erlandsson, L.; Zamora, D.; Palomino-Ángel, S.; Cousins, S.A. A call for consistency with the terms ‘wetter’ and ‘drier’ in climate change studies. Environ. Evid. 2021, 10, 1–7. [Google Scholar] [CrossRef]
- Adolf, C.; Tovar, C.; Kühn, N.; Behling, H.; Berrío, J.C.; Dominguez-Vázquez, G.; Figueroa-Rangel, B.; Gonzalez-Carranza, Z.; Islebe, G.A.; Hooghiemstra, H.; et al. Identifying drivers of forest resilience in long-term records from the Neotropics. Biol. Lett. 2020, 16, 20200005. [Google Scholar] [CrossRef]
- Cole, L.; Bhagwat, S.A.; Willis, K. Recovery and resilience of tropical forests after disturbance. Nat. Commun. 2014, 5, 3906. [Google Scholar] [CrossRef] [Green Version]
- Heidorn, P.B. Shedding Light on the Dark Data in the Long Tail of Science. Libr. Trends 2008, 57, 280–299. [Google Scholar] [CrossRef]
- Mahood, Q.; Van Eerd, D.; Irvin, E. Searching for grey literature for systematic reviews: Challenges and benefits. Res. Synth. Methods 2014, 5, 221–234. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Systematic Evidence Evaluation | Literature Review | |
|---|---|---|
| Definition | High-level overview of primary research on a focused question that identifies, selects, synthesises, and appraises all high-quality research evidence relevant to that question. | Qualitatively summarises evidence on a topic using informal or subjective methods to collect and interpret studies. |
| Aim | Answers a focused question. Reduces bias. | Provide summary or overview of topic. |
| Question | Clearly defined and answerable question. Uses a recommended framework, e.g., PICO or PECO. | Can be a general topic or a specific question. |
| Components | Pre-specified eligibility criteria. Systematic search strategy. Assessment of the validity of findings. Interpretation and presentation of results. Discussion. Summary of key findings. | Introduction. Methods. Discussion. Conclusion. |
| Number of Authors | Two or more. | One or more. |
| Timeline | Months to years (average eighteen months). | Weeks to months. |
| Requirement | Thorough knowledge of the topic. Perform searches of all relevant databases. | Understanding of topic. Perform searches of one or more databases. |
| Value | Connects practicing scientists to high-quality evidence. Supports evidence-based practice. | Provides summary of literature on the topic. |
| PECO | Definition | Description |
|---|---|---|
| Population | Populations of subject(s) of relevance to the review question. | Terrestrial palaeoenvironmental proxies interpreted to reconstruct forests, water, or land use change during the Holocene and within the Central American Isthmus. |
| Exposure | Environmental variable impacting the populations or to which the subject populations are exposed. | A window of time. |
| Counterfactual | What the exposure is compared to. Either a control with no exposure or an alternative counterfactual scenario. | A different window of time. |
| Outcome | Consequences of the exposure. All relevant variables that can be reliably measured. | Instances of change relating to forests, water, or land use change. |
| Database | Type of Literature | Description of Search |
|---|---|---|
| Web of Science | Platform of bibliographic databases | University of Oxford Core Collection of databases, 1945–2020. |
| Scopus | Bibliographic database | All records to 2020. |
| Cab Abstracts | Bibliographic database | All records included in the 1910–2020 databases. |
| PECO | Inclusion Criteria |
|---|---|
| Population | All studies that present primary palaeoenvironmental proxy data collected from a terrestrial environmental archive situated within the Central American Isthmus (terrestrial records only). Studies must represent a period of time within the Holocene. |
| Exposure | The environmental proxy data must be representative of a point in time. |
| Counterfactual | The environmental proxy data must be representative of a different point in time. |
| Outcomes | Palaeoenvironmental proxy data must have been interpreted to represent an environmental variable. Studies reconstruct an aspect of the past environment relating to forests, and water or land use change. |
| Coded Elements | Description |
|---|---|
| Article metadata | Authors, Title, Journal, Year of publication, Abstract, etc. |
| Study area metadata | Information about the study site including precise location. |
| Study proxy | Information about the study proxy, including data collection and interpretation. |
| Cross Cutting Lens | Study evidence for forests, and land use change or water. |
| Time data | Information about the time period examined, dating methods, materials, number of dates, constrained sequences, calibration, and reported errors. |
| Presentation of data | Temporal or depositional presentation of proxy data |
| Availability of original data | Accountability of the presented data. Are the graphically presented data available with the published article? |
| Forests | Hydro Climate | Sea Level | Tropical Storms | Agriculture | Land Use Change | |
|---|---|---|---|---|---|---|
| Charcoal (Macroscopic) | X | X | ||||
| Charcoal (Microscopic) | X | X | ||||
| Diatoms | X | X | ||||
| Foraminifera | X | X | ||||
| Geochemistry | X | X | ||||
| Grain Size Analysis | X | X | ||||
| Inorganic Carbon | X | X | X | X | X | |
| Magnetic Susceptibility | X | X | X | |||
| Organic Matter | X | X | X | X | X | |
| Ostracoda | X | X | ||||
| Phytoliths | X | X | X | |||
| Pollen | X | X | X | X | X | X |
| Stratigraphic Description | X | X | X | |||
| Testate Amoebae | X | X | ||||
| δ13C | X | X | X | X | X | X |
| δ15N | X | X | X | X | X | |
| δ18O | X | X | X |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harvey, W.J.; Petrokofsky, G.; Stansell, N.; Nogué, S.; Petrokofsky, L.; Willis, K.J. Forests, Water, and Land Use Change across the Central American Isthmus: Mapping the Evidence Base for Terrestrial Holocene Palaeoenvironmental Proxies. Forests 2021, 12, 1057. https://doi.org/10.3390/f12081057
Harvey WJ, Petrokofsky G, Stansell N, Nogué S, Petrokofsky L, Willis KJ. Forests, Water, and Land Use Change across the Central American Isthmus: Mapping the Evidence Base for Terrestrial Holocene Palaeoenvironmental Proxies. Forests. 2021; 12(8):1057. https://doi.org/10.3390/f12081057
Chicago/Turabian StyleHarvey, William J., Gillian Petrokofsky, Nathan Stansell, Sandra Nogué, Leo Petrokofsky, and Katherine J. Willis. 2021. "Forests, Water, and Land Use Change across the Central American Isthmus: Mapping the Evidence Base for Terrestrial Holocene Palaeoenvironmental Proxies" Forests 12, no. 8: 1057. https://doi.org/10.3390/f12081057
APA StyleHarvey, W. J., Petrokofsky, G., Stansell, N., Nogué, S., Petrokofsky, L., & Willis, K. J. (2021). Forests, Water, and Land Use Change across the Central American Isthmus: Mapping the Evidence Base for Terrestrial Holocene Palaeoenvironmental Proxies. Forests, 12(8), 1057. https://doi.org/10.3390/f12081057