
Comparison and Phylogenetic Analyses of Nine Complete Chloroplast Genomes of Zingibereae

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and DNA Sequencing
2.2. The Genomes of Plastome Assembly, Annotation and Structure
2.3. The Analysis of Codon Usage
2.4. Complete Chloroplast Genome Comparison and Molecular Marker Identification
2.5. The Analysis of Long Repetitive Sequences and Simple Sequence Repeats (SSRs)
2.6. Phylogenetic Analysis
3. Results
3.1. The Genome Structure and Content of Nine Zingibereae Species
3.2. Condon Usage Bias
3.3. Comparative Genomic Analysis
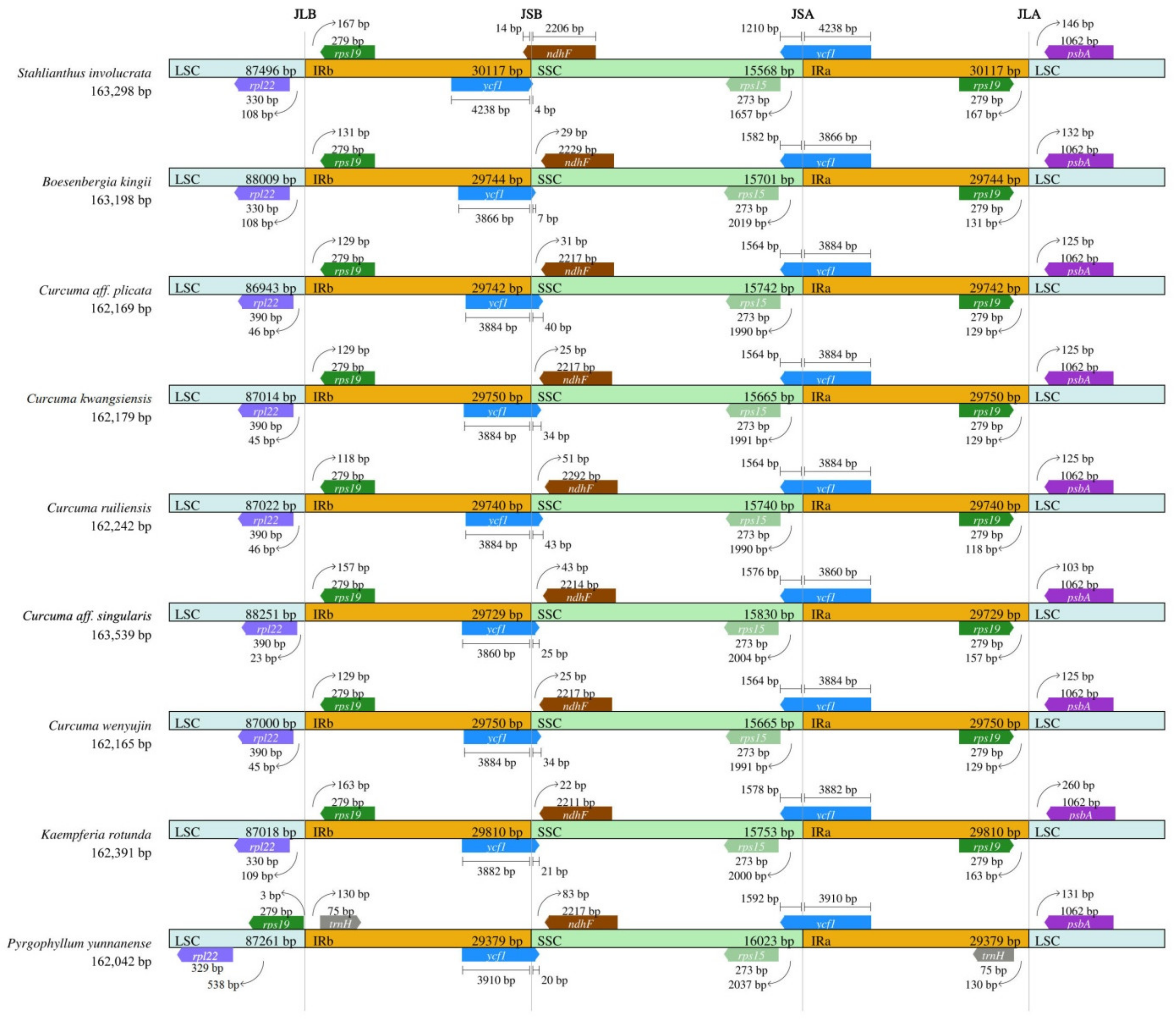
3.4. Expansion and Contraction of Inverted Repeats (IRs)
3.5. Repeat Structure and SSR Analysis
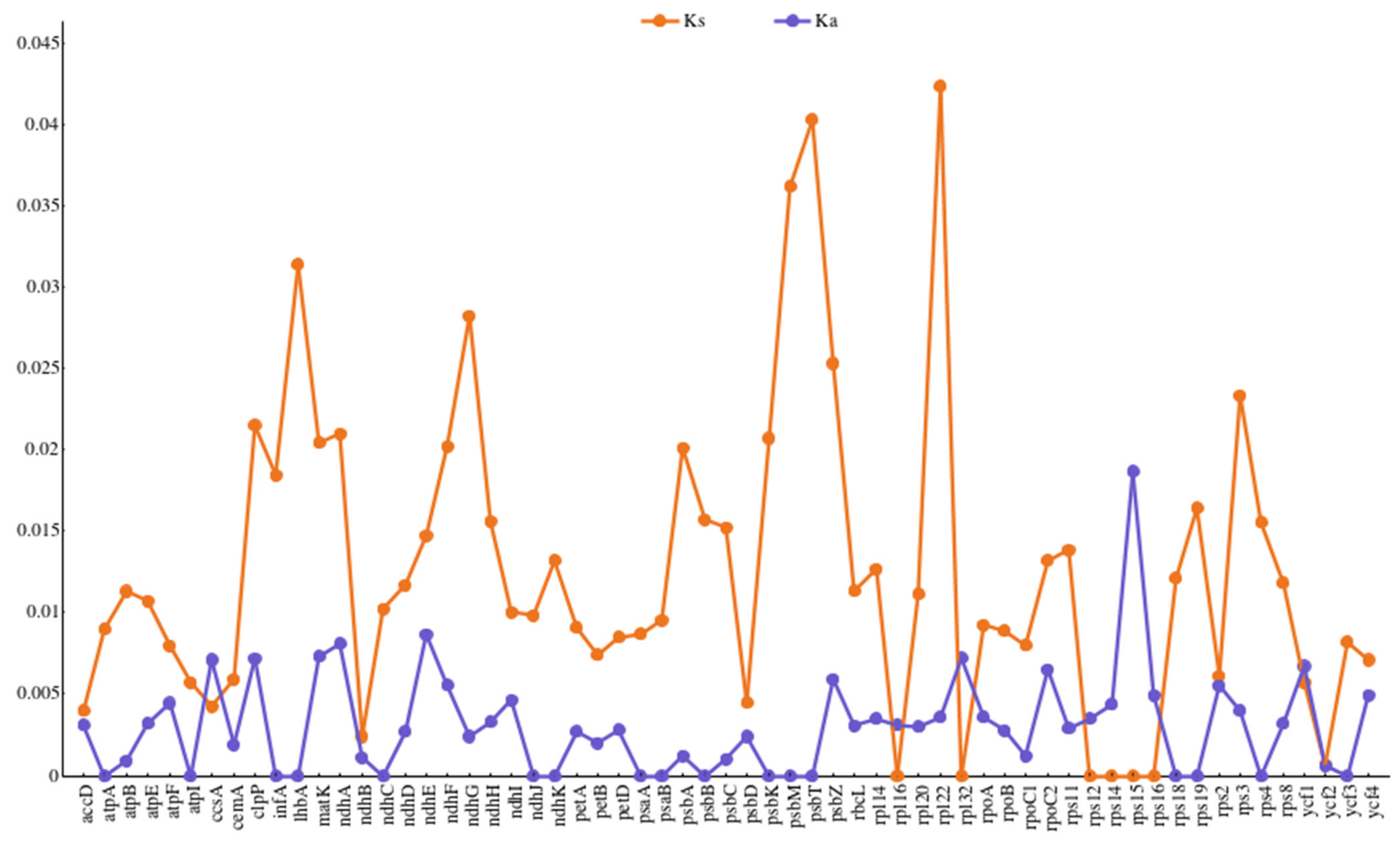
3.6. Sequence Divergence Hotspots
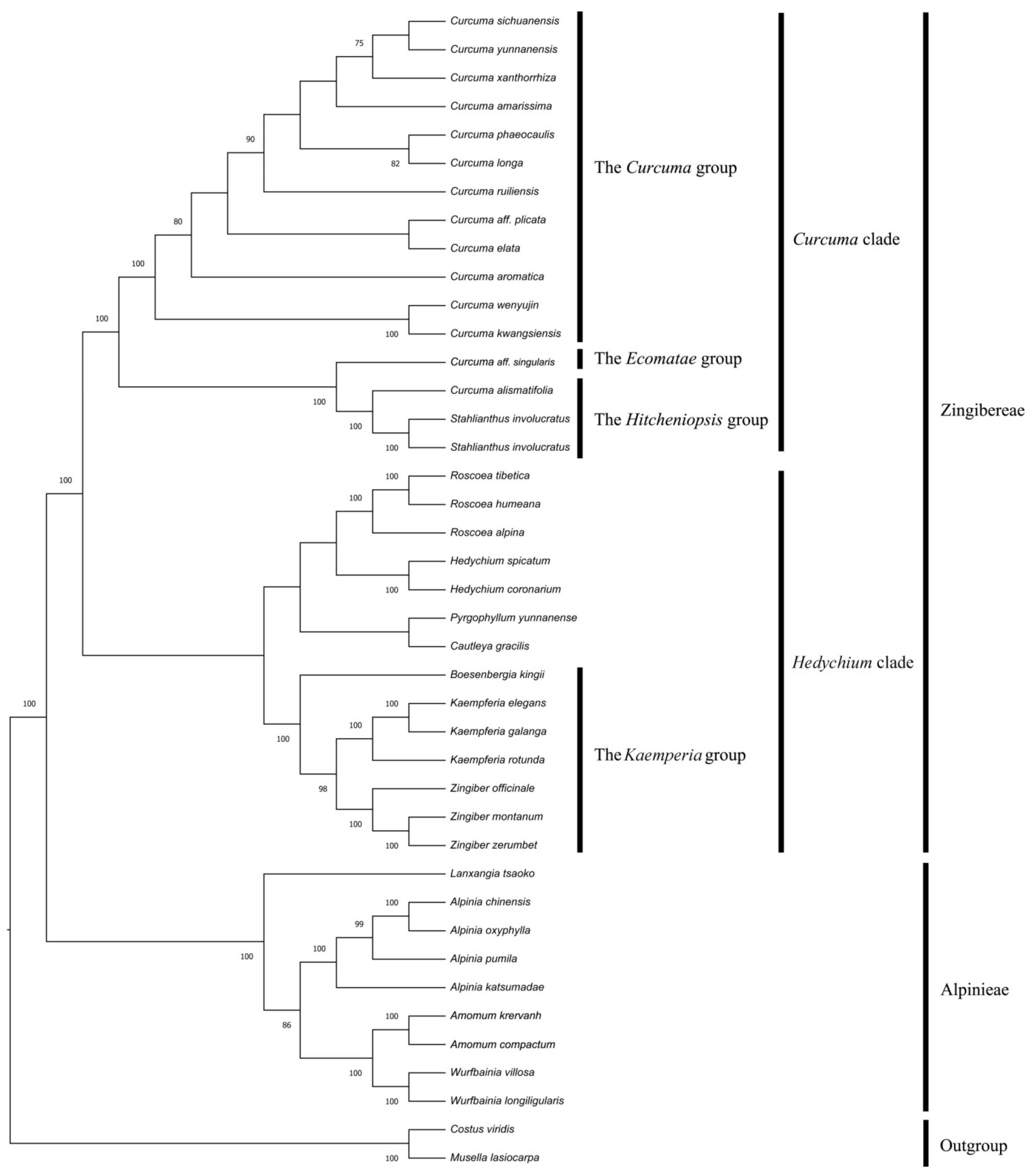
3.7. Phylogenetic Analysis Based on Chloroplast Genomes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CTAB | Cetyltrimethy lammonium Ammonium Bromide |
| CP | Chloroplast |
| IRs | Inverted repeats |
| Ka | Non-synonymous site |
| Ks | Synonymous site |
| Ka/Ks | the ratio of non-synonymous site and synonymous site |
| LSC | Large single-copy region |
| ML | Maximum likelihood |
| mono- | Mononucleotides |
| Pi | Nucleotide diversity values |
| rRNA | Ribosomal RNAs |
| RSCU | Relative synonymous codon usage |
| SSC | Small single-copy region |
| SSRs | Simple-sequence repeats |
| tRNA | Transfer RNAs |
References
- Kress, W.J.; Prince, L.M.; Williams, K.J. The phylogeny and a new classification of the gingers (Zingiberaceae): Evidence from molecular data. Am. J. Bot. 2002, 89, 1682–1696. [Google Scholar] [CrossRef]
- Wu, T.L.; Larsen, K. Flora of China; Botanical Garden Press: St. Louis, MO, USA; Science Press: Beijing, China, 2000; Volume 24, pp. 322–377. [Google Scholar]
- Liang, H.; Wang, Q.; Ding, C.; Zhang, L.; Yang, R. Chemical composition, antioxidant and antibacterial activities of essential oil of Curcuma phaeocaulis Valeton. Bangladesh J. Bot. 2020, 49, 531–540. [Google Scholar] [CrossRef]
- Yenjai, C.; Prasanphen, K.; Daodee, S.; Wongpanich, V.; Kittakoop, P. Bioactive flavonoids from Kaempferia parviflora. Fitoterapia 2004, 75, 89–92. [Google Scholar] [CrossRef]
- Tuchinda, P.; Reutrakul, V.; Claeson, P.; Pongprayoon, U.; Sematong, T.; Santisuk, T.; Taylor, W.C. Anti-inflammatory cyclohexenyl chalcone derivatives in Boesenbergia pandurata. Phytochemistry 2002, 59, 169–173. [Google Scholar] [CrossRef]
- Ngamriabsakul, C.; Newman, M.F.; Cronk, Q.C.B. The phylogeny of tribe Zingibereae (Zingiberaceae) based on ITS (nrDNA) and trnL-F (cpDNA) sequences. Edinb. J. Bot. 2004, 60, 483–507. [Google Scholar] [CrossRef]
- Linnaeus, C. Species Plantarum; Forgotten Books: London, UK, 1753. [Google Scholar]
- Záveská, E.; Fer, T.; Šída, O.; Krak, K.; Marhold, K.; Leong-Škorničková, J. Phylogeny of Curcuma (Zingiberaceae) based on plastid and nuclear sequences: Proposal of the new subgenus Ecomata. Taxon 2012, 61, 747–763. [Google Scholar] [CrossRef]
- Leong-Škorničková, J.; Otakar, Š.; Záveská, E.; Marhold, K. History of infrageneric classification, typification of supraspecific names and outstanding transfers in Curcuma (Zingiberaceae). Taxon 2015, 64, 362–373. [Google Scholar] [CrossRef]
- Gagnepain, M.F. Zingibéracées Nouvelles Ou Mal Connues De L’Herbier Du Muséum. Bull. Soc. Bot. France 1901, 48, 205. [Google Scholar] [CrossRef]
- Schumann, K.M. Zingiberaceae. In Das Pflanzenreich; Engler, A., Ed.; Engelmann: Berlin, Germany, 1904; Volume 46, pp. 1–20. [Google Scholar]
- Larsen, K.; Smith, R.M. Notes on Caulokaempferia. Notes Roy. Bot. Gard. Edinb. 1972, 31, 287–295. [Google Scholar]
- Wu, T.; Chen, Y. Pyrgophyllum, a new genus of Zingiberaceae from China. J. Univ. Chin. Acad. Sci. 1989, 27, 124–128. [Google Scholar]
- Mood, J.; MacKechnie, L.; Veldkamp, J.-F.; Dey, S. The history and identity of Boesenbergia longiflora (Zingiberaceae) and descriptions of five related taxa. Gard. Bull. Singap. 2013, 65, 47–95. [Google Scholar]
- Leong-Škorničková, J.; Lý, N.-S.; Poulsen, A.D.; Tosh, J.; Forrest, A. Newmania: A new ginger genus from central Vietnam. Taxon 2011, 60, 1386–1396. [Google Scholar] [CrossRef]
- Smith, R.M. A new genus of Zingiberaceae from N. Burma. Notes Notes Roy. Bot. Gard. Edinb. 1980, 38, 13–17. [Google Scholar]
- Sabu, M.; Prasanthkumar, M.G.; Leong-Škorničková, J.; Jayasree, S. Transfer of Kaempferia siphonantha Baker to Boesenbergia Kuntze (Zingiberaceae). Rheedea 2004, 14, 55–59. [Google Scholar]
- Techaprasan, J.; Ngamriabsakul, C.; Klinbunga, S.; Chusacultanachai, S.; Jenjittikul, T. Genetic variation and species identification of Thai Boesenbergia (Zingiberaceae) analyzed by chloroplast DNA polymorphism. J. Biochem. Mol. Biol. 2006, 39, 361–370. [Google Scholar] [CrossRef]
- Wicke, S.; Schneeweiss, G.M.; Depamphilis, C.W.; Müller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef]
- Shinozaki, K.; Ohme, M.; Tanaka, M.; Wakasugi, T.; Hayashida, N.; Matsubayashi, T.; Zaita, N.; Chunwongse, J.; Obokata, J.; Yamaguchi-Shinozaki, K.; et al. The complete nucleotide sequence of the tobacco chloroplast genome: Its gene organization and expression. EMBO J. 1986, 5, 2043–2049. [Google Scholar] [CrossRef] [PubMed]
- Bock, R. Plastid biotechnology: Prospects for herbicide and insect resistance, metabolic engineering and molecular farming. Curr. Opin. Biotechnol. 2007, 18, 100–106. [Google Scholar] [CrossRef]
- Ma, J.; Yang, B.; Zhu, W.; Sun, L.; Tian, J.; Wang, X. The complete chloroplast genome sequence of Mahonia bealei (Berberidaceae) reveals a significant expansion of the inverted repeat and phylogenetic relationship with other angiosperms. Gene 2013, 528, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ma, J.; Yang, B.; Li, R.; Zhu, W.; Sun, L.; Tian, J.; Zhang, L. The complete chloroplast genome sequence of Taxus chinensis var. mairei (Taxaceae): Loss of an inverted repeat region and comparative analysis with related species. Gene 2014, 540, 201–209. [Google Scholar] [CrossRef]
- Liang, H.; Zhang, Y.; Deng, J.; Gao, G.; Ding, C.; Zhang, L.; Yang, R. The complete chloroplast genome sequences of 14 Curcuma species: Insights into genome evolution and phylogenetic relationships within Zingiberales. Front. Genet. 2020, 11. [Google Scholar] [CrossRef]
- Reith, M.; Munholland, J. Complete nucleotide sequence of the Porphyra purpurea chloroplast genome. Plant Mol. Biol. Rep. 1995, 13, 333–335. [Google Scholar] [CrossRef]
- Hallick, R.; Hong, L.; Drager, R.; Favreau, M.; Monfort, A.; Orsat, B.; Spielmann, A.; Stutz, E. Complete sequence of Euglena gracilis chloroplast DNA. Nucl. Acids Res. 1993, 21, 3537–3544. [Google Scholar] [CrossRef]
- Alwadani, K.G.; Janes, J.K.; Andrew, R.L. Chloroplast genome analysis of box-ironbark Eucalyptus. Mol. Phylogenet. Evol. 2019, 136, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, D.J.; Simpson, B.B.; Ortiz, E.M.; Shimizu, G.H.; Jansen, R.K. Incongruence between gene trees and species trees and phylogenetic signal variation in plastid genes. Mol. Phylogenet. Evol. 2019, 138, 219–232. [Google Scholar] [CrossRef]
- Xue, S.; Shi, T.; Luo, W.; Ni, X.; Iqbal, S.; Ni, Z.; Huang, X.; Yao, D.; Shen, Z.; Gao, Z. Comparative analysis of the complete chloroplast genome among Prunus mume, P. armeniaca, and P. salicina. Hortic. Res. 2019, 6, 89. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhou, T.; Kanwal, N.; Zhao, Y.; Bai, G.; Zhao, G. Completion of eight Gynostemma BL.(Cucurbitaceae) chloroplast genomes: Characterization, comparative analysis, and phylogenetic relationships. Front. Plant Sci. 2017, 8, 1583. [Google Scholar] [CrossRef]
- Patel, R.K.; Jain, M. NGS QC Toolkit: A toolkit for quality control of next generation sequencing data. PLoS ONE 2012, 7, e30619. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Huang, D.I.; Cronk, Q.C.B. Plann: A command-line application for annotating plastome sequences. Appl. Plant Sci. 2015, 3, 1500026. [Google Scholar] [CrossRef]
- Wyman, S.K.; Jansen, R.K.; Boore, J.L. Automatic annotation of organellar genomes with DOGMA. Bioinformatics 2004, 20, 3252–3255. [Google Scholar] [CrossRef]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucl. Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef]
- Xia, X. DAMBE5: A comprehensive software package for data analysis in molecular biology and evolution. Mol. Biol. Evol. 2013, 30, 1720–1728. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucl. Acids Res. 2004, 32, 273–279. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The manifold applications of repeat analysis on a genomic scale. Nucl. Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (* and Other Methods); Version 4.0 a149; Sinauer Associates: Sunderland, UK, 2002. [Google Scholar]
- Chen, J.; Hao, Z.; Xu, H.; Yang, L.; Liu, G.; Sheng, Y.; Zheng, C.; Zheng, W.; Cheng, T.; Shi, J. The complete chloroplast genome sequence of the relict woody plant Metasequoia glyptostroboides. Front. Plant Sci. 2015, 6, 447. [Google Scholar] [CrossRef] [PubMed]
- Powell, W.; Morgante, M.; McDevitt, R.; Vendramin, G.; Rafalski, J. Polymorphic simple sequence repeat regions in chloroplast genomes: Applications to the population genetics of pines. Proc. Natl. Acad. Sci. USA 1995, 92, 7759–7763. [Google Scholar] [CrossRef]
- Park, H.; Kim, C.; Lee, Y.M.; Kim, J.H. Development of chloroplast microsatellite markers for the endangered Maianthemum bicolor (Asparagaceae sl). Appl. Plant Sci. 2016, 4, 1600032. [Google Scholar] [CrossRef]
- Yan, Y.D.; Li, X.Y.; Worth, J.R.; Lin, X.Y.; Ruhsam, M.; Chen, L.; Wu, X.T.; Wang, M.q.; Thomas, P.I.; Wen, Y.F. Development of chloroplast microsatellite markers for Glyptostrobus pensilis (Cupressaceae). Appl. Plant Sci. 2019, 7, e11277. [Google Scholar] [CrossRef]
- Bremer, B.; Bremer, K.; Chase, M.W.; Fay, M.F.; Reveal, J.L.; Soltis, D.E.; Soltis, P.S.; Stevens, P.F.; Anderberg, A.A.; Moore, M.J.; et al. An update of the angiosperm phylogeny group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar]
- Du, Y.; Bi, Y.; Yang, F.; Zhang, M.; Chen, X.; Xue, J.; Zhang, X. Complete chloroplast genome sequences of Lilium: Insights into evolutionary dynamics and phylogenetic analyses. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Ree, R.H. Uplift-driven diversification in the Hengduan Mountains, a temperate biodiversity hotspot. Proc. Natl. Acad. Sci. USA 2017, 114, 3444–3451. [Google Scholar] [CrossRef] [PubMed]
- Li, D.M.; Xu, Y.C.; Zhu, G.F. Complete chloroplast genome of the plant Stahlianthus Involucratus (Zingiberaceae). Mitochondrial DNA Part B Resour. 2019, 4, 2702–2703. [Google Scholar] [CrossRef]
- Cui, Y.; Chen, X.; Nie, L.; Sun, W.; Hu, H.; Lin, Y.; Li, H.; Zheng, X.; Song, J.; Yao, H. Comparison and phylogenetic analysis of chloroplast genomes of three medicinal and edible Amomum species. Int. J. Mol. Sci. 2019, 20, 4040. [Google Scholar] [CrossRef]
- Cui, Y.; Nie, L.; Sun, W.; Xu, Z.; Wang, Y.; Yu, J.; Song, J.; Yao, H. Comparative and phylogenetic analyses of ginger (Zingiber officinale) in the family Zingiberaceae based on the complete chloroplast genome. Plants 2019, 8, 283. [Google Scholar] [CrossRef]
- Gu, C.; Ma, L.; Wu, Z.; Chen, K.; Wang, Y. Comparative analyses of chloroplast genomes from 22 Lythraceae species: Inferences for phylogenetic relationships and genome evolution within Myrtales. BMC Plant Biol. 2019, 19, 281. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhao, C.; Liu, X. Complete chloroplast genome sequences of Kaempferia galanga and Kaempferia elegans: Molecular structures and comparative analysis. Molecules 2019, 24, 474. [Google Scholar] [CrossRef]
- Wu, C.S.; Wang, Y.N.; Liu, S.M.; Chaw, S.M. Chloroplast genome (cpDNA) of Cycas taitungensis and 56 cp protein-coding genes of Gnetum parvifolium: Insights into cpDNA evolution and phylogeny of extant seed plants. Mol. Biol. Evol. 2007, 24, 1366–1379. [Google Scholar] [CrossRef] [PubMed]
- Downie, S.R.; Jansen, R.K. A comparative analysis of whole plastid genomes from the apiales: Expansion and contraction of the inverted repeat, mitochondrial to plastid transfer of DNA, and identification of highly divergent noncoding regions. Syst. Bot. 2015, 40, 336–351. [Google Scholar] [CrossRef]
- Williams, A.M.; Friso, G.; Van Wijk, K.J.; Sloan, D.B. Extreme variation in rates of evolution in the plastid Clp protease complex. Plant J. 2019, 98, 243–259. [Google Scholar] [CrossRef]
- Chen, Y.; Zhong, H.; Zhu, Y.; Huang, Y.; Wu, S.; Liu, Z.; Lan, S.; Zhai, J. Plastome structure and adaptive evolution of Calanthe s.l. species. PeerJ 2020, 8, e10051. [Google Scholar] [CrossRef]
- Zhou, T.; Ruhsam, M.; Wang, J.; Zhu, H.; Li, W.; Zhang, X.; Xu, Y.; Xu, F.; Wang, X. The complete chloroplast genome of Euphrasia regelii, pseudogenization of ndh genes and the phylogenetic relationships within Orobanchaceae. Front. Genet. 2019, 10, 444. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, Z.; Sablok, G.; Daskalova, E.; Zahmanova, G.; Apostolova, E.; Yahubyan, G.; Baev, V. Chloroplast genome analysis of resurrection tertiary relict haberlea rhodopensis highlights genes important for desiccation stress response. Front. Plant Sci. 2017, 8, 204. [Google Scholar] [CrossRef]
- Piot, A.; Hackel, J.; Christin, P.-A.; Besnard, G. One-third of the plastid genes evolved under positive selection in PACMAD grasses. Planta 2018, 247, 255–266. [Google Scholar] [CrossRef]
- Parks, M.; Cronn, R.; Liston, A. Increasing phylogenetic resolution at low taxonomic levels using massively parallel sequencing of chloroplast genomes. BMC Biol. 2009, 7. [Google Scholar] [CrossRef]
- Wheeler, G.L.; Dorman, H.E.; Buchanan, A.; Challagundla, L.; Wallace, L.E. A review of the prevalence, utility, and caveats of using chloroplast simple sequence repeats for studies of plant biology. Appl. Plant Sci. 2014, 2, 1400059. [Google Scholar] [CrossRef]
- Shahzadi, I.; Abdullah; Mehmood, F.; Ali, Z.; Ahmed, I.; Mirza, B. Chloroplast genome sequences of Artemisia maritima and Artemisia absinthium: Comparative analyses, mutational hotspots in genus Artemisia and phylogeny in family Asteraceae. Genomics 2020, 112, 1454–1463. [Google Scholar] [CrossRef] [PubMed]
- Wood, T.H.; Whitten, W.M.; Williams, N.H. Phylogeny of Hedychium and related genera (Zingiberaceae) based on ITS sequence data. Edinb. J. Bot. 2000, 57, 261–270. [Google Scholar] [CrossRef]
- Newman, M.F. Distichochlamys, a new genus from Vietnam. Edinb. J. Bot. 1995, 52, 65–69. [Google Scholar] [CrossRef]
- Searle, R.J.; Hedderson, T.A.J. A preliminary phylogeny of the Hedychieae tribe (Zingiberaceae) based on ITS sequences of the nuclear rRNA cistron. In Monocots: Systematics and Evolution; Wilson, K.L.M., Ed.; CSIRO Publishing: Collingwood, Australia, 2000; pp. 710–718. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Species | Voucher Number | Locality | Accession Number |
|---|---|---|---|---|
| 1 | Boesenbergia kingii | 17081502 (IBSC) | Yunnan, China | MW326451 |
| 2 | Curcuma ruiliensis | 17082304 (IBSC) | Yunnan, China | MW326454 |
| 3 | Curcuma aff. singularis | 1722 (IBSC) | Chiangmai, Thailand | MW326455 |
| 4 | Curcuma aff. plicata | 17081107 (IBSC) | Yunnan, China | MW326452 |
| 5 | Curcuma kwangsiensis | 17083001 (IBSC) | Guangxi, China | MW326453 |
| 6 | Curcuma wenyujin | 201544 (IBSC) | Guangdong, China | MW326456 |
| 7 | Kaempferia rotunda | 17081102 (IBSC) | Yunnan, China | MW326457 |
| 8 | Pyrgophyllum yunnanense | 2014106 (IBSC) | Sichuan, China | MW326458 |
| 9 | Stahlianthusinvolucratus | 19031203 (IBSC) | Laos | MW326459 |
| Species | Size | PCGs | tRNAs | rRNAs | Genes | GC% | Length (LSC) | Length (SSC) | Length (IR) | GC% (LSC) | GC% (SSC) | GC% (IR) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B. kingii | 163,198 | 88 | 38 | 8 | 134 | 36.16 | 88,009 | 15,701 | 29,744 | 33.95 | 29.66 | 41.15 |
| C. aff. plicata | 162,169 | 87 | 38 | 8 | 133 | 36.20 | 86,943 | 15,742 | 29,742 | 34.01 | 29.62 | 41.14 |
| C. ruiliensis | 162,242 | 87 | 38 | 8 | 133 | 36.19 | 87,022 | 15,740 | 29,740 | 33.99 | 29.56 | 41.16 |
| C. aff. singularis | 163,539 | 87 | 38 | 8 | 133 | 36.07 | 88,251 | 15,830 | 29,729 | 33.83 | 29.51 | 41.15 |
| C. kwangsiensis | 162,179 | 87 | 38 | 8 | 133 | 36.18 | 87,014 | 15,665 | 29,750 | 33.97 | 29.65 | 41.13 |
| C. wenyujin | 162,165 | 87 | 38 | 8 | 133 | 36.19 | 87,000 | 15,665 | 29,750 | 33.98 | 29.66 | 41.13 |
| K. rotunda | 162,391 | 87 | 38 | 8 | 133 | 36.25 | 87,018 | 15,753 | 29,810 | 34.11 | 29.66 | 41.12 |
| P. yunnanense | 162,042 | 86 | 38 | 8 | 132 | 36.06 | 87,261 | 16,023 | 29,379 | 33.81 | 29.14 | 41.30 |
| S. involucratus | 163,298 | 88 | 38 | 8 | 134 | 36.00 | 87,496 | 15,568 | 30,117 | 33.78 | 29.60 | 40.89 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, H.; Chen, J. Comparison and Phylogenetic Analyses of Nine Complete Chloroplast Genomes of Zingibereae. Forests 2021, 12, 710. https://doi.org/10.3390/f12060710
Liang H, Chen J. Comparison and Phylogenetic Analyses of Nine Complete Chloroplast Genomes of Zingibereae. Forests. 2021; 12(6):710. https://doi.org/10.3390/f12060710
Chicago/Turabian StyleLiang, Heng, and Juan Chen. 2021. "Comparison and Phylogenetic Analyses of Nine Complete Chloroplast Genomes of Zingibereae" Forests 12, no. 6: 710. https://doi.org/10.3390/f12060710
APA StyleLiang, H., & Chen, J. (2021). Comparison and Phylogenetic Analyses of Nine Complete Chloroplast Genomes of Zingibereae. Forests, 12(6), 710. https://doi.org/10.3390/f12060710