
Comparative Dynamics of Above-Ground Litter Production and Decomposition from Eucalyptus grandis Hill ex Maiden and Pinus taeda L., and Their Contribution to Soil Organic Carbon


Abstract
1. Introduction
2. Materials and Methods
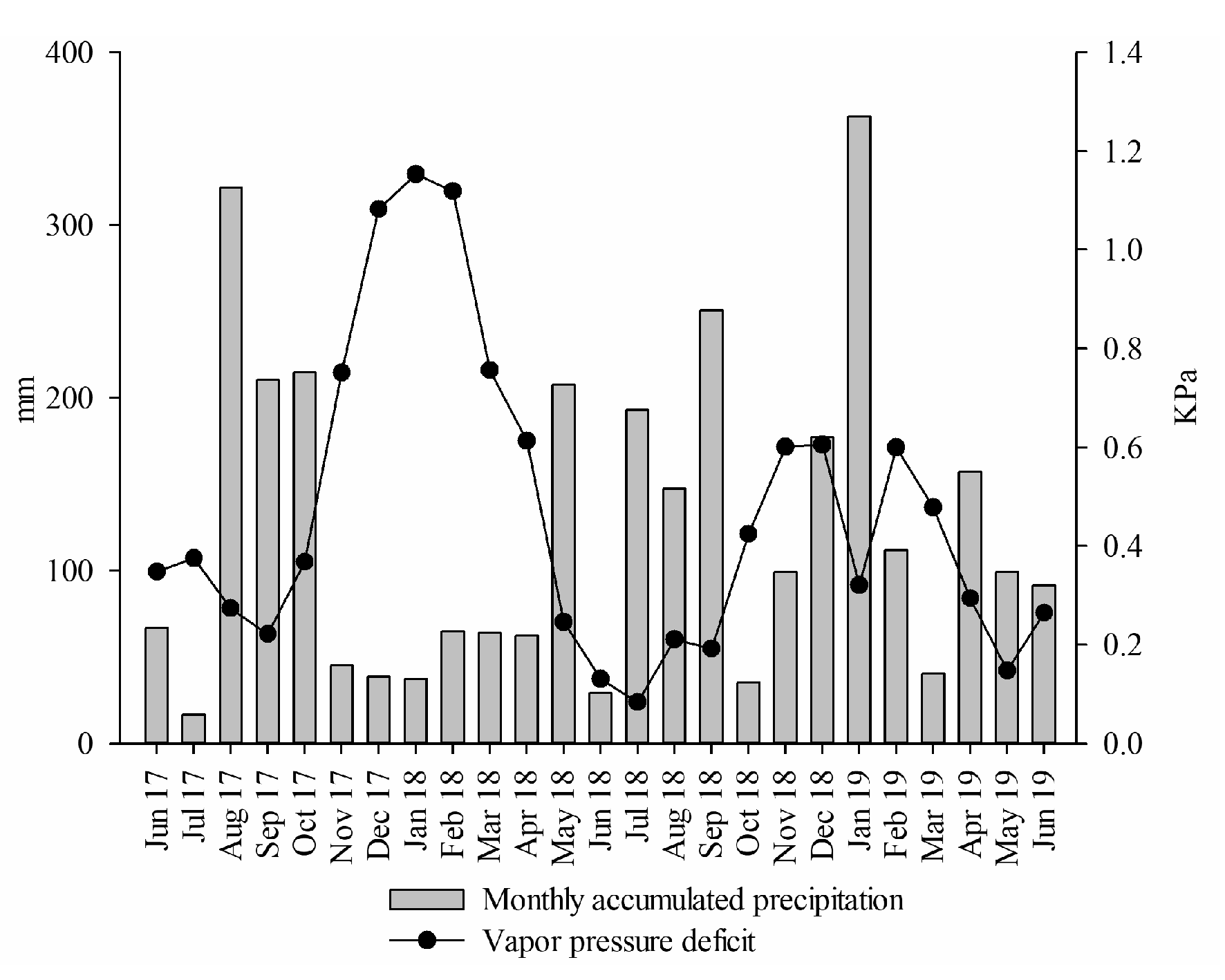
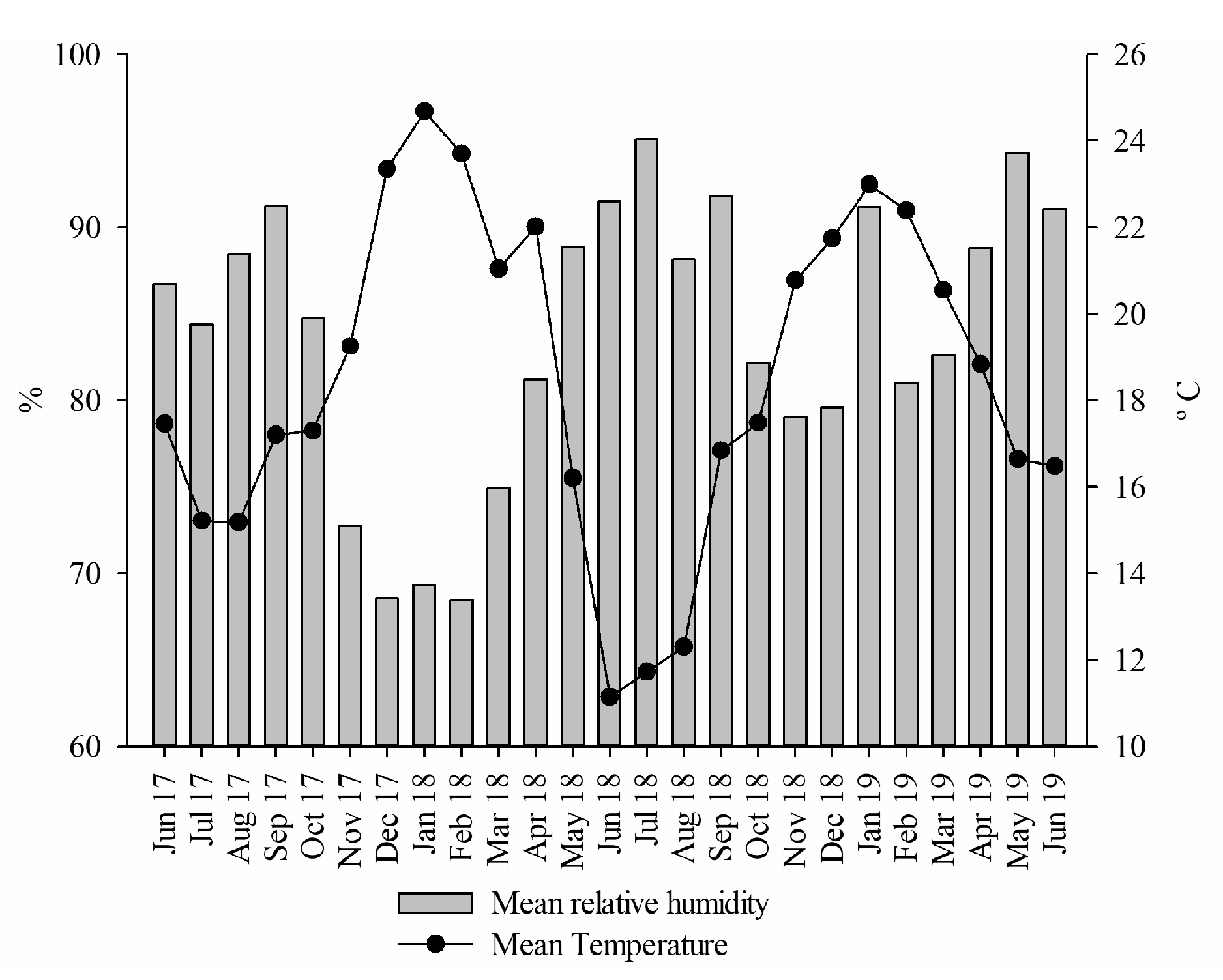
2.1. Characteristics of the Experimental Site
2.2. Litterfall, Decomposition and Accumulation Assessment
2.3. Chemical Analyses
2.4. Calculation of δ13C in Soil and Litter
2.5. Statistical Analysis
3. Results
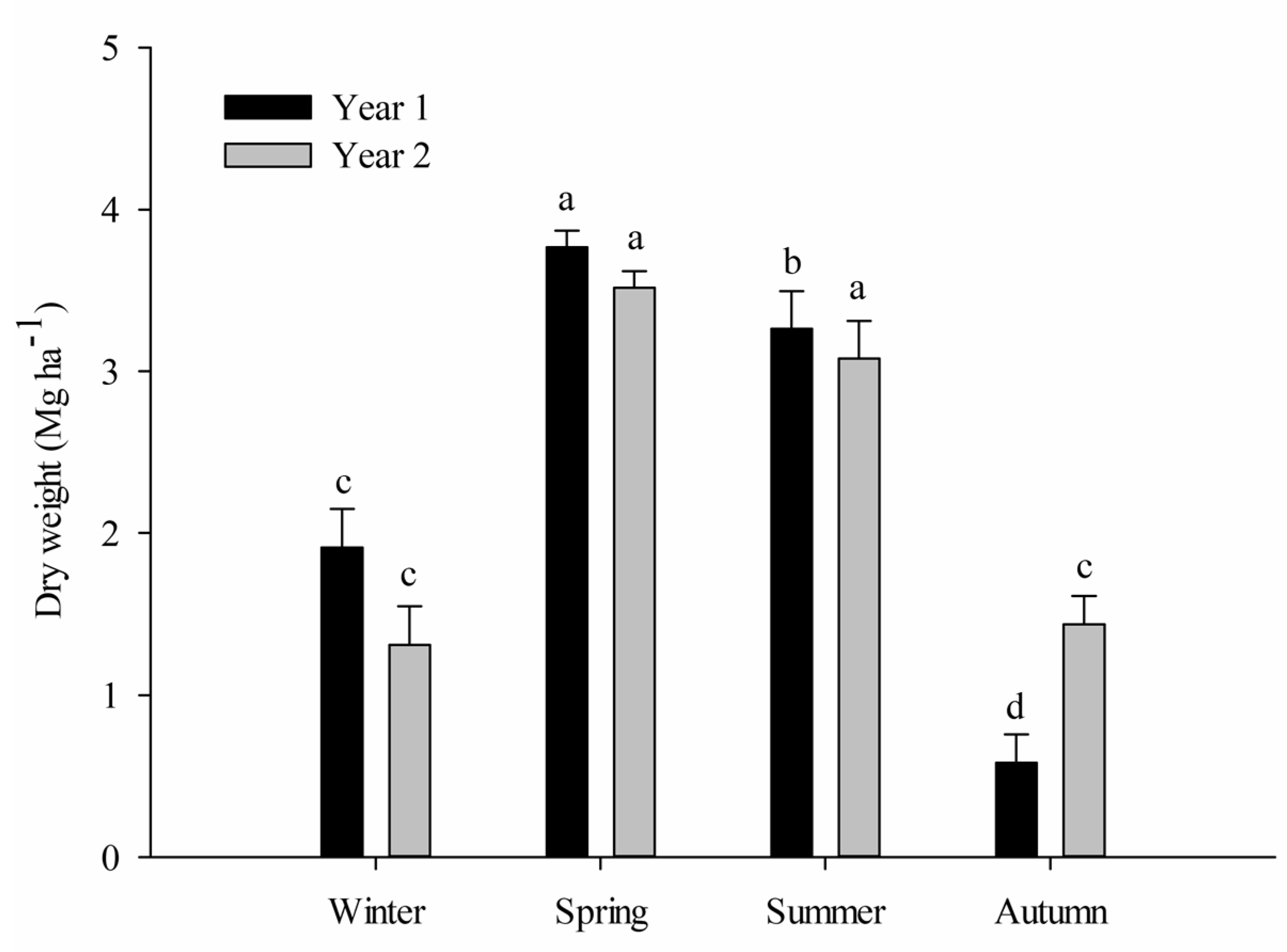
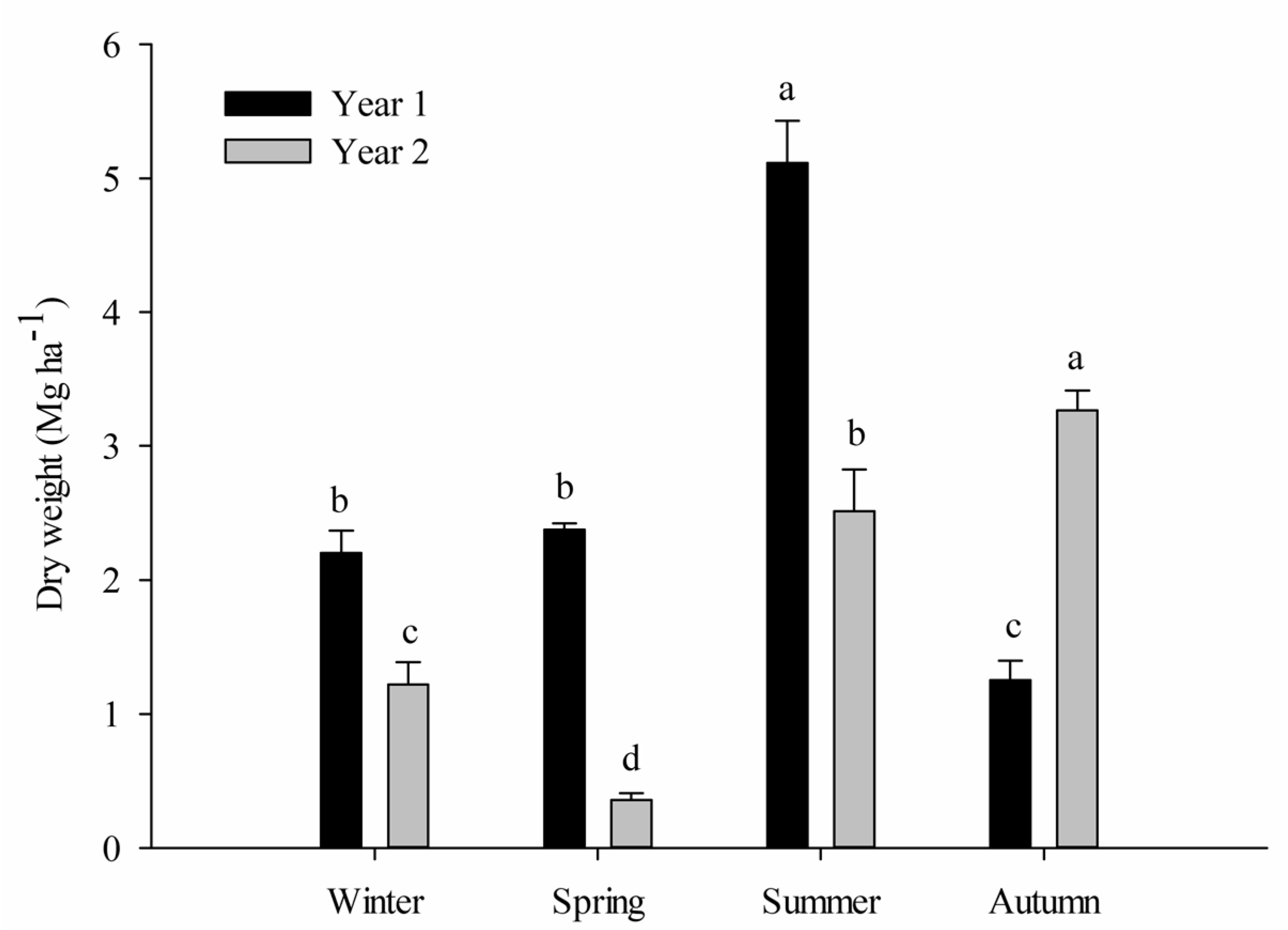
3.1. Litter Production
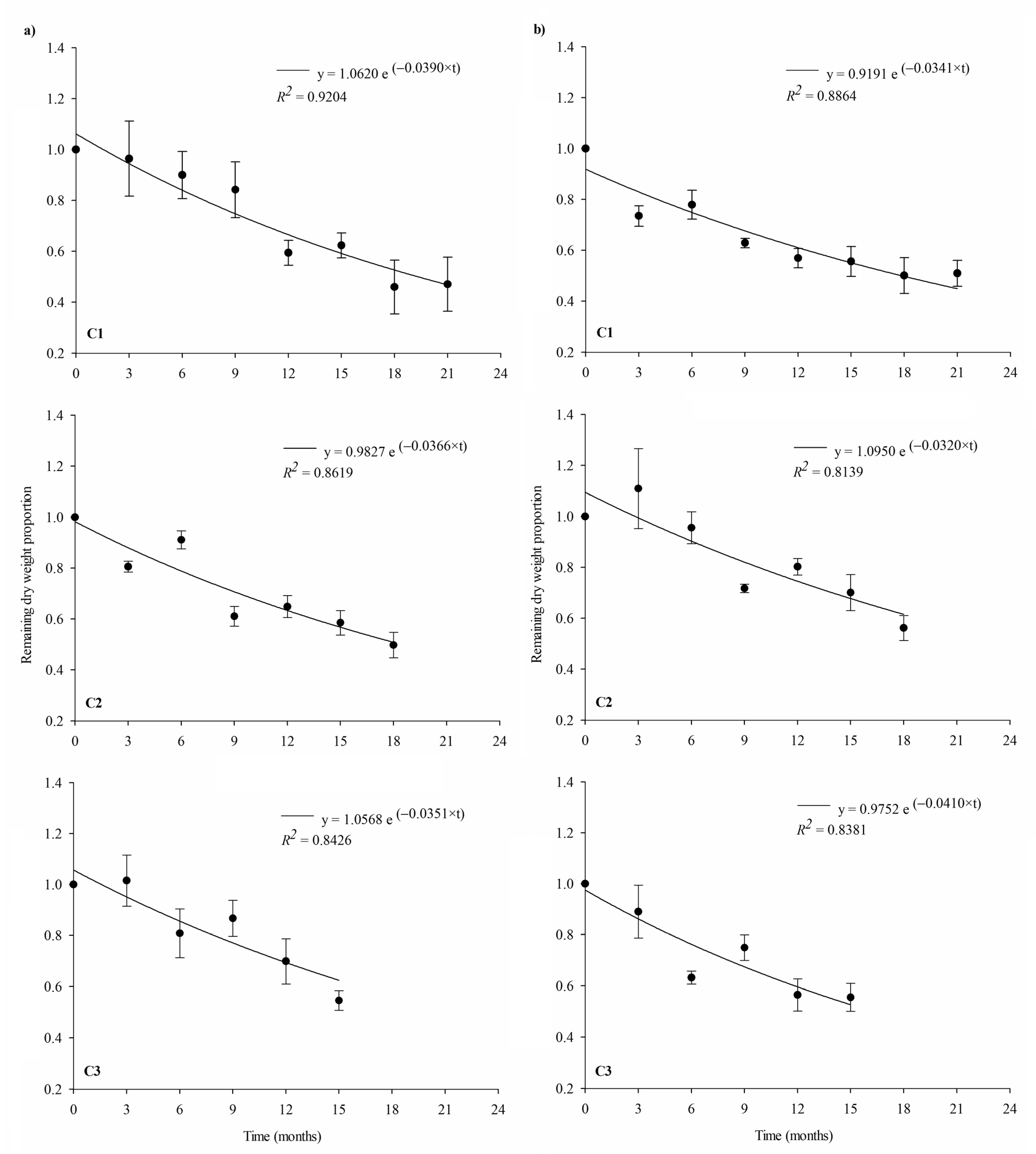
3.2. Litter Decomposition
3.3. Carbon Stock Remaining in Litter
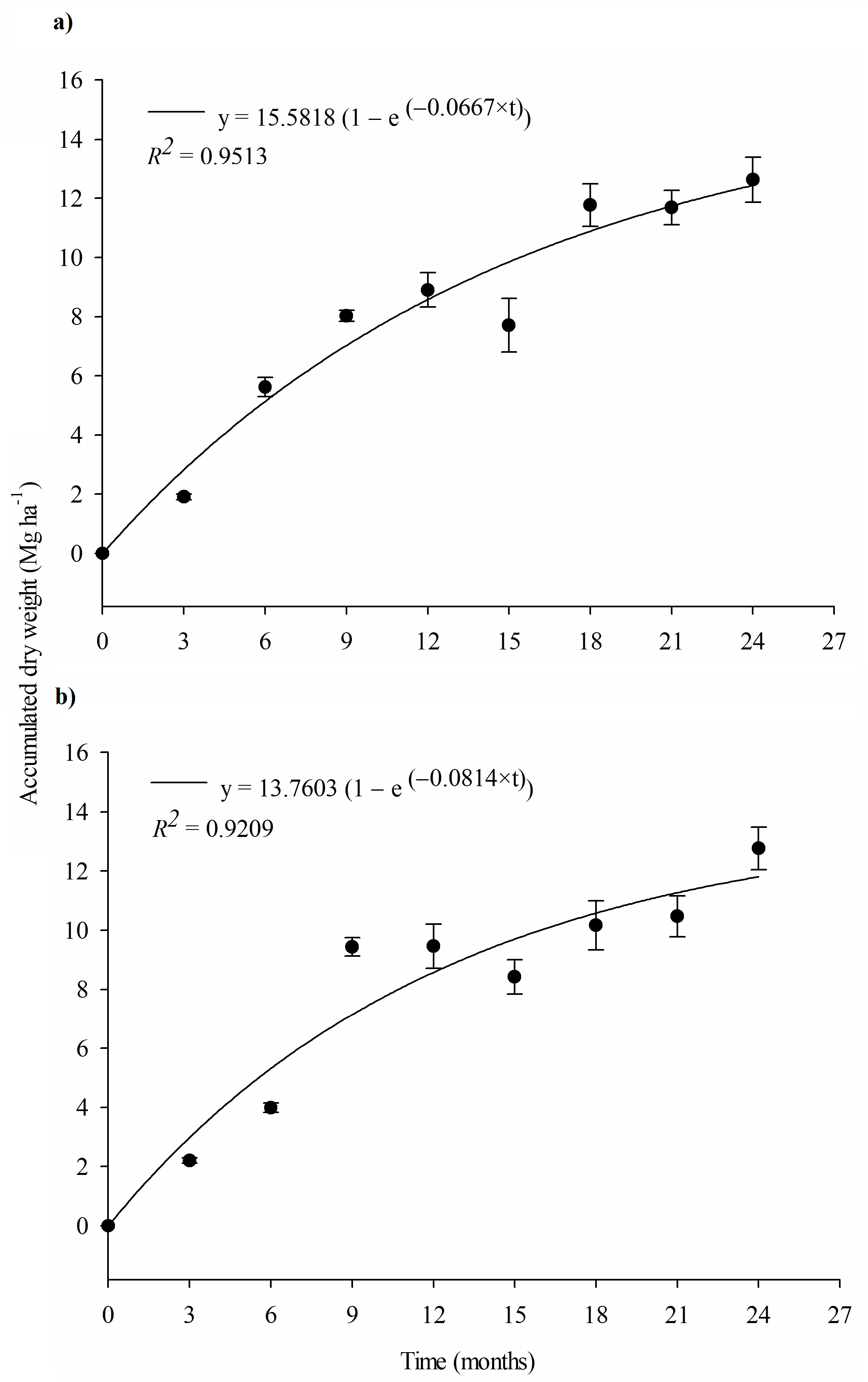
3.4. Litter Accumulation Dynamics
3.5. Change in the Concentration and Stock of C and δ13C Values in the Soil
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thompson, L.M.; Troeh, F.R. Los Suelos y Su Fertilidad, 4th ed.; Reverté: Barcelona, Spain, 1982; ISBN 978-84-291-9001-4. [Google Scholar]
- Goma-Tchimbakala, J.; Bernhard-Reversat, F. Comparison of Litter Dynamics in Three Plantations of an Indigenous Timber-Tree Species (Terminalia Superba) and a Natural Tropical Forest in Mayombe, Congo. For. Ecol. Manag. 2006, 229, 304–313. [Google Scholar] [CrossRef]
- Olson, J.S. Energy Storage and the Balance of Producers and Decomposers in Ecological Systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- Theodorou, C.; Bowen, G.D. Effects of Fertilizer on Litterfall and N and P Release from Decomposing Litter in a Pinus radiata Plantation. For. Ecol. Manag. 1990, 32, 87–102. [Google Scholar] [CrossRef]
- Pook, E.W.; Gill, A.M.; Moore, P.H.R. Long-Term Variation of Litter Fall, Canopy Leaf Area and Flowering in a Eucalyptus maculata Forest on the South Coast of New South Wales. Aust. J. Bot. 1997, 45, 737. [Google Scholar] [CrossRef]
- Li, Z.A.; Zou, B.; Xia, H.; Ren, H.; Mo, J.; Weng, H. Litterfall Dynamics of an Evergreen Broadleaf Forest and a Pine Forest in the Subtropical Region of China. For. Sci. 2005, 51, 608–615. [Google Scholar] [CrossRef]
- Baker, T.G. Dry Matter, Nitrogen, and Phosphorus Content of Litterfall and Branchfall in Pinus radiata and Eucalyptus Forests. N. Z. J. For. Sci. 1983, 13, 205–221. [Google Scholar]
- Abelho, M.; Graça, M.A.S. Effects of Eucalyptus Afforestation on Leaf Litter Dynamics and Macroinvertebrate Community Structure of Streams in Central Portugal. Hydrobiologia 1996, 324, 195–204. [Google Scholar] [CrossRef]
- Demessie, A.; Singh, B.R.; Lal, R.; Strand, L.T. Leaf Litter Fall and Litter Decomposition under Eucalyptus and Coniferous Plantations in Gambo District, Southern Ethiopia. Soil Plant Sci. 2011, 1–10. [Google Scholar] [CrossRef]
- Erkan, N.; Comez, A.; Aydin, A.C.; Denli, O.; Erkan, S. Litterfall in Relation to Stand Parameters and Climatic Factors in Pinus brutia Forests in Turkey. Scand. J. For. Res. 2018, 33, 338–346. [Google Scholar] [CrossRef]
- Swift, M.J.; Heal, O.W.; Anderson, J.M.; Anderson, J.M. Decomposition in Terrestrial Ecosystems; University of California Press: Oakland, CA, USA, 1979; ISBN 978-0-520-04001-4. [Google Scholar]
- Prescott, C.E.; Zabek, L.M.; Staley, C.L.; Kabzems, R. Decomposition of Broadleaf and Needle Litter in Forests of British Columbia: Influences of Litter Type, Forest Type, and Litter Mixtures. Can. J. For. Res. 2000, 30, 1742–1750. [Google Scholar] [CrossRef]
- Canhoto, C.; Graça, M.A.S. Leaf Barriers to Fungal Colonization and Shredders (Tipula lateralis) Consumption of Decomposing Eucalyptus globulus. Microb. Ecol. 1999, 37, 163–172. [Google Scholar] [CrossRef]
- Hernández, J.; del Pino, A.; Salvo, L.; Arrarte, G. Nutrient Export and Harvest Residue Decomposition Patterns of a Eucalyptus dunnii Maiden Plantation in Temperate Climate of Uruguay. For. Ecol. Manag. 2009, 258, 92–99. [Google Scholar] [CrossRef]
- Aerts, R. Climate, Leaf Litter Chemistry and Leaf Litter Decomposition in Terrestrial Ecosystems: A Triangular Relationship. Oikos 1997, 79, 439. [Google Scholar] [CrossRef]
- Berg, B. Litter Decomposition and Organic Matter Turnover in Northern Forest Soils. For. Ecol. Manag. 2000, 133, 13–22. [Google Scholar] [CrossRef]
- Xuluc-Tolosa, F. Leaf Litter Decomposition of Tree Species in Three Successional Phases of Tropical Dry Secondary Forest in Campeche, Mexico. For. Ecol. Manag. 2003, 174, 401–412. [Google Scholar] [CrossRef]
- Hättenschwiler, S. Effects of Tree Species Diversity on Litter Quality and Decomposition. In Forest Diversity and Function; Scherer-Lorenzen, M., Körner, C., Schulze, E.-D., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; Volume 176, pp. 149–164. ISBN 978-3-540-22191-3. [Google Scholar]
- Tateno, R.; Tokuchi, N.; Yamanaka, N.; Du, S.; Otsuki, K.; Shimamura, T.; Xue, Z.; Wang, S.; Hou, Q. Comparison of Litterfall Production and Leaf Litter Decomposition between an Exotic Black Locust Plantation and an Indigenous Oak Forest near Yan’an on the Loess Plateau, China. For. Ecol. Manag. 2007, 241, 84–90. [Google Scholar] [CrossRef]
- Laclau, P. Biomass and Carbon Sequestration of Ponderosa Pine Plantations and Native Cypress Forests in Northwest Patagonia. For. Ecol. Manag. 2003, 180, 317–333. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.; Wang, J.; Shi, Z.; Lu, L.; Guo, W.; Jia, H.; Cai, D. Dynamics and Speciation of Organic Carbon during Decomposition of Leaf Litter and Fine Roots in Four Subtropical Plantations of China. For. Ecol. Manag. 2013, 300, 43–52. [Google Scholar] [CrossRef]
- Hernández, J.; del Pino, A.; Vance, E.D.; Califra, Á.; Del Giorgio, F.; Martínez, L.; González-Barrios, P. Eucalyptus and Pinus Stand Density Effects on Soil Carbon Sequestration. For. Ecol. Manag. 2016, 368, 28–38. [Google Scholar] [CrossRef]
- Trouve, C.; Mariotti, A.; Schwartz, D.; Guillet, B. Soil Organic Carbon Dynamics under Eucalyptus and Pinus Planted on Savannas in the Congo. Soil Biol. Biochem. 1994, 26, 287–295. [Google Scholar] [CrossRef]
- Bubb, K.A.; Xu, Z.H.; Simpson, J.A.; Saffigna, P.G. Some Nutrient Dynamics Associated with Litterfall and Litter Decomposition in Hoop Pine Plantations of Southeast Queensland, Australia. For. Ecol. Manag. 1998, 110, 343–352. [Google Scholar] [CrossRef]
- Blanco, J.A.; Imbert, J.B.; Castillo, F.J. Nutrient Return via Litterfall in Two Contrasting Pinus sylvestris Forests in the Pyrenees under Different Thinning Intensities. For. Ecol. Manag. 2008, 256, 1840–1852. [Google Scholar] [CrossRef]
- Goya, J.F.; Frangi, J.L.; Pérez, C.; Tea, F.D. Decomposition and Nutrient Release from Leaf Litter in Eucalyptus grandis Plantations on Three Different Soils in Entre Ríos, Argentina. Bosque 2008, 29, 217–226. [Google Scholar] [CrossRef]
- Olsson, B.A.; Guedes, B.S.; Dahlin, A.S.; Hyvönen, R. Predicted Long-Term Effects of Decomposition of Leaf Litter from Pinus taeda, Eucalyptus cloeziana and Deciduous Miombo Trees on Soil Carbon Stocks. Glob. Ecol. Conserv. 2019, 17, e00587. [Google Scholar] [CrossRef]
- Castaño, J.P.; Giménez, A.; Ceroni, M.; Furest, J.; Aunchayna, R.; Bidegain, M. Caracterización Agroclimática Del Uruguay 1980-2009; INIA Serie Técnica; Instituto Nacional de Investigación Agropecuaria: Montevideo, Uruguay, 2011; ISBN 978-9974-38-330-2. [Google Scholar]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; Department of Agriculture, Natural Resources Conservation Service: Washington, DC, USA, 2014. [Google Scholar]
- Altamirano, A.; Da Silva, H.; Durán, A.; Echeverría, A.; Panario, D.; Puentes, R. Carta de Reconocimiento de Suelos del Uruguay: Clasificación de Suelos; Ministerio de Ganadería Agricultura y Pesca: Montevideo, Uruguay, 1976. [Google Scholar]
- Schwanninger, M.; Hinterstoisser, B. Klason Lignin: Modifications to Improve the Precision of the Standardized Determination. Holzforschung 2002, 56, 161–166. [Google Scholar] [CrossRef]
- Corbeels, M.; O’Connell, A.M.; Grove, T.S.; Mendham, D.S.; Rance, S.J. Nitrogen Release from Eucalypt Leaves and Legume Residues as Influenced by Their Biochemical Quality and Degree of Contact with Soil. Plant Soil. 2003, 250, 15–28. [Google Scholar] [CrossRef]
- Michener, R.H.; Lajtha, K. (Eds.) Ecological Methods and Concepts Series, 2nd ed.; Blackwell Pub: Malden, MA, USA, 2007; ISBN 978-1-4051-2680-9. [Google Scholar]
- Balesdent, J.; Mariotti, A. Measurement of Soil Organic Matter Turnover Using 13C Natural Abundance. In Mass Spectrometry of Soils; CRC Press: Boca Raton, FL, USA, 1996; pp. 83–111. [Google Scholar]
- Draper, N.R.; Smith, H. Applied Regression Analysis, 3rd ed.; Wiley Series in Probability and Statistics; John Wiley & Sons: Hoboken, NJ, USA, 1998; ISBN 978-0-471-17082-2. [Google Scholar]
- SAS Institute. SAS® Studio 3.3: User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2015. [Google Scholar]
- Harmon, M.E.; Silver, W.L.; Fasth, B.; Chen, H.; Burke, I.C.; Parton, W.J.; Hart, S.C.; Currie, W.S. LIDET Long-Term Patterns of Mass Loss during the Decomposition of Leaf and Fine Root Litter: An Intersite Comparison. Glob. Chang. Biol. 2009, 15, 1320–1338. [Google Scholar] [CrossRef]
- Schlatter, J.E.; Gerding, V.; Calderón, S. Aporte de la Hojarasca al Ciclo Biogeoquímico en Plantaciones de Eucalyptus nitens, X Región, Chile. Bosque 2006, 27. [Google Scholar] [CrossRef]
- De Vargas, G.R.; Marques, R.; Bianchin, J.E.; Teixeira, W.W.R.; Blum, H. Biomass Deposition and Chemical Composition of Litterfall in Clonal Eucalyptus Plantations. Floresta E Ambiente 2019, 26. [Google Scholar] [CrossRef]
- Hernández, J.; del Pino, A.; Califra, A. Eucalyptus and Pine Stand Spacing Density Study and Its Implications for Carbon Sequestration Processes. Final Report; Faculty of Agronomy: Montevideo, Uruguay, 2014. [Google Scholar]
- Momolli, D.R.; Schumacher, M.V.; Viera, M.; Ludvichak, A.A.; do Couto Guimarães, C.; de Souza, H.P. Litterfall and Nutrient Return in Eucalyptus dunnii Maiden in the Pampa Biome, Brazil. J. Agric. Sci. 2019, 11, 362. [Google Scholar] [CrossRef]
- O’Connell, A.M.; Menage, P.M.A. Litter Fall and Nutrient Cycling in Karri (Eucalyptus diversicolor F. Muell.) Forest in Relation to Stand Age. Austral Ecol. 1982, 7, 49–62. [Google Scholar] [CrossRef]
- Nouvellon, Y.; Epron, D.; Marsden, C.; Kinana, A.; Le Maire, G.; Deleporte, P.; Saint-André, L.; Bouillet, J.-P.; Laclau, J.-P. Age-Related Changes in Litter Inputs Explain Annual Trends in Soil CO2 Effluxes over a Full Eucalyptus Rotation after Afforestation of a Tropical Savannah. Biogeochemistry 2012, 111, 515–533. [Google Scholar] [CrossRef]
- Berg, B.; Meentemeyer, V. Litter Fall in Some European Coniferous Forests as Dependent on Climate: A Synthesis. Can. J. For. Res. 2001, 31, 292–301. [Google Scholar] [CrossRef]
- Cromer, R.N.; Tompkins, D.; Barr, N.J.; Williams, E.R.; Stewart, H.T.L. Litter-Fall In a Pinus radiata Forest: The Effect of Irrigation and Fertilizer Treatments. J. Appl. Ecol. 1984, 21, 313. [Google Scholar] [CrossRef]
- Crockford, R.H.; Richardson, D.P. Litterfall, Litter and Associated Chemistry in a Dry Sclerophyll Eucalypt Forest and a Pine Plantation in South-Eastern Australia: 1. Litterfall and Litter. Hydrol. Process. 1998, 12, 20. [Google Scholar] [CrossRef]
- Raison, R.J.; Myers, B.J.; Benson, M.L. Dynamics of Pinus radiata Foliage in Relation to Water and Nitrogen Stress: I. Needle Production and Properties. For. Ecol. Manag. 1992, 52, 139–158. [Google Scholar] [CrossRef]
- Wienand, K.T.; Stock, W.D. Long-Term Phosphorus Fertilization Effects on the Litter Dynamics of an Age Sequence of Pinus elliottii Plantations in the Southern Cape of South Africa. For. Ecol. Manag. 1995, 75, 135–146. [Google Scholar] [CrossRef]
- Ribeiro, C.; Madeira, M.; Araújo, M.C. Decomposition and Nutrient Release from Leaf Litter of Eucalyptus globulus Grown under Different Water and Nutrient Regimes. For. Ecol. Manag. 2002, 171, 31–41. [Google Scholar] [CrossRef]
- Trofymow, J.A.; Moore, T.R.; Titus, B.; Prescott, C.; Morrison, I.; Siltanen, M.; Smith, S.; Fyles, J.; Wein, R.; Camiré, C.; et al. Rates of Litter Decomposition over 6 Years in Canadian Forests: Influence of Litter Quality and Climate. Can. J. For. Res. 2002, 32, 789–804. [Google Scholar] [CrossRef]
- Xu, S.; Liu, Y.; Cui, Y.; Pei, Z. Litter Decomposition in a Subtropical Plantation in Qianyanzhou, China. J. For. Res. 2011, 16, 8–15. [Google Scholar] [CrossRef]
- Harmon, M.E.; Naddlehoffer, K.J.; John, M. Blair Measuring decomposition, nutrient turnover and stores in plant litter. In Standard Soil Methods for Long Term Ecological Research; Robertson, G.P., Bledsoe, C.S., Coleman, D.C., Sollins, P.S., Eds.; Oxford University Press: New York, NY, USA, 1999. [Google Scholar]
- del Pino, A.; Hernández, J.; Arrarte, G. Descomposición de Residuos de Pinus taeda En Uruguay. In Jornadas de Actualización Técnica; Departamento de Suelos y Aguas: Sayago, Uruguay, 2010. [Google Scholar]
- Berg, B.; Johansson, M.-B.; Meentemeyer, V. Litter Decomposition in a Transect of Norway Spruce Forests: Substrate Quality and Climate Control. Can. J. For. Res. 2000, 30, 1136–1147. [Google Scholar] [CrossRef]
- Kim, C.; Jeong, J.; Cho, H.-S.; Son, Y. Carbon and Nitrogen Status of Litterfall, Litter Decomposition and Soil in Even-Aged Larch, Red Pine and Rigitaeda Pine Plantations. J. Plant Res. 2010, 123, 403–409. [Google Scholar] [CrossRef]
- Kiser, L.; Fox, T.; Carlson, C. Foliage and Litter Chemistry, Decomposition, and Nutrient Release in Pinus taeda. Forests 2013, 4, 595–612. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, Y.; Cao, J.; Fu, S.; Hu, S.; Wu, J.; Zhao, J.; Liu, Z. Stand Age and Species Traits Alter the Effects of Understory Removal on Litter Decomposition and Nutrient Dynamics in Subtropical Eucalyptus Plantations. Glob. Ecol. Conserv. 2019, 20, e00693. [Google Scholar] [CrossRef]
- Gholz, H.L.; Wedin, D.A.; Smitherman, S.M.; Harmon, M.E.; Parton, W.J. Long-Term Dynamics of Pine and Hardwood Litter in Contrasting Environments: Toward a Global Model of Decomposition. Glob. Change Biol. 2000, 6, 751–765. [Google Scholar] [CrossRef]
- Zhang, M.; Cheng, X.; Geng, Q.; Shi, Z.; Luo, Y.; Xu, X. Leaf Litter Traits Predominantly Control Litter Decomposition in Streams Worldwide. Glob. Ecol. Biogeogr. 2019, 28, 1469–1486. [Google Scholar] [CrossRef]
- Woo, K.-S.; Fins, L.; McDonald, G.I.; Wenny, D.L.; Eramian, A. Effects of Nursery Environment on Needle Morphology of Pinus monticola Dougl. and Implications for Tree Improvement Programs. New For. 2002, 24, 113–129. [Google Scholar] [CrossRef]
- Sayer, E.J.; Tanner, E.V.J.; Lacey, A.L. Effects of Litter Manipulation on Early-Stage Decomposition and Meso-Arthropod Abundance in a Tropical Moist Forest. For. Ecol. Manag. 2006, 229, 285–293. [Google Scholar] [CrossRef]
- Leff, J.W.; Wieder, W.R.; Taylor, P.G.; Townsend, A.R.; Nemergut, D.R.; Grandy, A.S.; Cleveland, C.C. Experimental Litterfall Manipulation Drives Large and Rapid Changes in Soil Carbon Cycling in a Wet Tropical Forest. Glob. Chang. Biol. 2012, 18, 2969–2979. [Google Scholar] [CrossRef]
- Vasconcelos, S.S.; Zarin, D.J.; Capanu, M.; Littell, R.; Davidson, E.A.; Ishida, F.Y.; Santos, E.B.; Araújo, M.M.; Aragão, D.V.; Rangel-Vasconcelos, L.G.T.; et al. Moisture and Substrate Availability Constrain Soil Trace Gas Fluxes in an Eastern Amazonian Regrowth Forest. Glob. Biogeochem. Cycles 2004, 18. [Google Scholar] [CrossRef]
- Li, Y.; Xu, M.; Sun, O.J.; Cui, W. Effects of Root and Litter Exclusion on Soil CO2 Efflux and Microbial Biomass in Wet Tropical Forests. Soil Biol. Biochem. 2004, 36, 2111–2114. [Google Scholar] [CrossRef]
- Sayer, E.J. Using Experimental Manipulation to Assess the Roles of Leaf Litter in the Functioning of Forest Ecosystems. Biol. Rev. 2005, 81, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Gurmesa, G.A.; Liu, L.; Zhang, T.; Fu, S.; Liu, Z.; Dong, S.; Ma, C.; Mo, J. Effects of Litter Manipulation on Litter Decomposition in a Successional Gradients of Tropical Forests in Southern China. PLoS ONE 2014, 9, e99018. [Google Scholar] [CrossRef]
- Hui, D.; Zhang, D.; Norby, R.J. Field Litter Decomposition Rate Estimation: Does Incubation Starting Time Matter? In Proceedings of the 2011 International Conference on Multimedia Technology, Hangzhou, China, 26–28 July 2011; pp. 1558–1560. [Google Scholar]
- Constantinides, M.; Fownes, J. Nitrogen Mineralization from Leaves and Litter of Tropical Plants: Relationships to Nitrogen, Lignin and Soluble Polyphenol Concentrations. Soil Biol. Biochem. 1994, 26, 49–55. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Vitousek, P.M. The Role of Polyphenols in Terrestrial Ecosystem Nutrient Cycling. Trends Ecol. Evol. 2000, 15, 238–243. [Google Scholar] [CrossRef]
- Palm, C.A.; Gachengo, C.N.; Delve, R.J.; Cadisch, G.; Giller, K.E. Organic Inputs for Soil Fertility Management in Tropical Agroecosystems: Application of an Organic Resource Database. Agric. Ecosyst. Environ. 2001, 83, 27–42. [Google Scholar] [CrossRef]
- Piatek, K.B.; Allen, H.L. Are Forest Floors in Mid-Rotation Stands of Loblolly Pine (Pinus taeda) a Sink for Nitrogen and Phosphorus? Can. J. For. Res. 2001, 31, 1164–1174. [Google Scholar] [CrossRef]
- Chae, H.M.; Choi, S.H.; Lee, S.H.; Cha, S.; Yang, K.C.; Shim, J.K. Effect of Litter Quality on Needle Decomposition for Four Pine Species in Korea. Forests 2019, 10, 371. [Google Scholar] [CrossRef]
- Raison, R.J.; Woods, P.V.; Khanna, P.K. Dynamics of Fine Fuels in Recurrently Burnt Eucalypt Forests. Aust. For. 1983, 46, 294–302. [Google Scholar] [CrossRef]
- Raison, R.J.; Woods, P.V.; Khanna, P.K. Decomposition and Accumulation of Litter after Fire in Sub-Alpine Eucalypt Forests. Austral Ecol. 1986, 11, 9–19. [Google Scholar] [CrossRef]
- Gould, J.S.; Lachlan McCaw, W.; Phillip Cheney, N. Quantifying Fine Fuel Dynamics and Structure in Dry Eucalypt Forest (Eucalyptus marginata) in Western Australia for Fire Management. For. Ecol. Manag. 2011, 262, 531–546. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Pallardy, S.G.; Kramer, P.J. Physiology of Woody Plants, 2nd ed.; Academic Press: San Diego, CA, USA, 1997; ISBN 978-0-12-424162-6. [Google Scholar]
- Finotti, R.; Freitas, S.R.; Cerqueira, R.; Vieira, M.V. A Method to Determine the Minimum Number of Litter Traps in Litterfall Studies. Biotropica 2003, 35, 419–421. [Google Scholar] [CrossRef]
- Hu, Y.-L.; Zeng, D.-H.; Ma, X.-Q.; Chang, S.X. Root Rather than Leaf Litter Input Drives Soil Carbon Sequestration after Afforestation on a Marginal Cropland. For. Ecol. Manag. 2016, 362, 38–45. [Google Scholar] [CrossRef]
- Turner, J.; Lambert, M. Change in Organic Carbon in Forest Plantation Soils in Eastern Australia. For. Ecol. Manag. 2000, 133, 231–247. [Google Scholar] [CrossRef]
- Guo, L.B.; Cowie, A.L.; Montagu, K.D.; Gifford, R.M. Carbon and Nitrogen Stocks in a Native Pasture and an Adjacent 16-Year-Old Pinus radiata D. Don. Plantation in Australia. Agric. Ecosyst. Environ. 2008, 124, 205–218. [Google Scholar] [CrossRef]
- Lima, A.M.N.; Silva, I.R.; Neves, J.C.L.; Novais, R.F.; Barros, N.F.; Mendonça, E.S.; Smyth, T.J.; Moreira, M.S.; Leite, F.P. Soil Organic Carbon Dynamics Following Afforestation of Degraded Pastures with Eucalyptus in Southeastern Brazil. For. Ecol. Manag. 2006, 235, 219–231. [Google Scholar] [CrossRef]
- Smith, P. How Long before a Change in Soil Organic Carbon Can Be Detected? Glob. Change Biol. 2004, 10, 1878–1883. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C.; Santo, A.V.D.; Johnson, D. Humus Buildup in Boreal Forests: Effects of Litter Fall and Its N Concentration. Can. J. For. Res. 2011. [Google Scholar] [CrossRef]
- Prescott, C.E. Litter Decomposition: What Controls It and How Can We Alter It to Sequester More Carbon in Forest Soils? Biogeochemistry 2010, 101, 133–149. [Google Scholar] [CrossRef]
- Paul, K.I.; Polglase, P.J.; Nyakuengama, J.G.; Khanna, P.K. Change in Soil Carbon Following Afforestation. For. Ecol. Manag. 2002, 168, 241–257. [Google Scholar] [CrossRef]
- Epron, D.; Marsden, C.; Thongo M’Bou, A.; Saint-André, L.; d’Annunzio, R.; Nouvellon, Y. Soil Carbon Dynamics Following Afforestation of a Tropical Savannah with Eucalyptus in Congo. Plant Soil 2009, 323, 309–322. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hor. | Depth | Clay | Sand | Silt | C | pH a | Al | Ca | Mg | K | Na | Total Bases | CECe | Base Sat. | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (cm) | (g kg−1) | H2O | KCl | (cmolc kg−1) | (%) | |||||||||||
| A | Mean | 0–48 | 126 | 54 | 820 | 9.36 | 4.7 | 3.8 | 1.60 | 1.33 | 0.78 | 0.26 | 0.35 | 2.73 | 4.33 | 63 |
| Std. D. | - | - | - | - | (1.82) | (0.1) | (0.1) | (0.45) | (0.32) | (0.17) | (0.05) | (0.05) | (0.51) | (0.61) | (9) | |
| AB | Mean | 48–66 | 233 | 87 | 681 | 7.37 | 4.7 | 3.8 | 2.14 | 1.39 | 0.77 | 0.22 | 0.40 | 2.78 | 4.92 | 58 |
| Std. D. | - | - | - | - | (2.21) | (0.1) | (0.1) | (0.87) | (0.32) | (0.21) | (0.05) | (0.02) | (0.52) | (1.04) | (12) | |
| Bt | Mean | 66–90 | 342 | 116 | 543 | 7.15 | 4.7 | 3.7 | 4.01 | 2.08 | 1.31 | 0.26 | 0.43 | 4.08 | 8.09 | 51 |
| Std. D. | - | - | - | - | (1.23) | (0.1) | (0.1) | (1.36) | (0.33) | (0.34) | (0.06) | (0.07) | (0.67) | (1.71) | (9) | |
| Characteristic | Eucalyptus grandis Hill ex Maiden | Pinus taeda L. |
|---|---|---|
| Spacing 1 (m) | 3.5 × 3.5 | 3.5 × 3.5 |
| Initial density (trees ha−1) | 816 | 816 |
| DBH 2 (cm) | 29.70 | 31.70 |
| Total Height (m) | 36.30 | 22.50 |
| Variable | E. grandis | P. taeda |
|---|---|---|
| Accumulated precipitation | 0.19 p = 0.3627 | −0.45 p = 0.0277 |
| Mean relative humidity | −0.75 p < 0.0001 | −0.28 p = 0.1791 |
| Mean minimum relative humidity | −0.60 p = 0.0019 | −0.36 p = 0.0865 |
| Mean maximum relative humidity | −0.59 p = 0.0026 | −0.11 p = 0.5992 |
| Mean temperature | 0.65 p = 0.0006 | 0.41 p = 0.0467 |
| Mean minimum temperature | 0.64 p = 0.0008 | 0.34 p = 0.1024 |
| Mean maximum temperature | 0.25 p = 0.2447 | 0.80 p < 0.0001 |
| Mean vapor pressure deficit | 0.72 p = 0.0001 | 0.38 p = 0.0663 |
| C | N | Soluble C | Soluble Pol. | Lignin | C:N | L:N | |
|---|---|---|---|---|---|---|---|
| (g kg−1) 1 | |||||||
| E. grandis | |||||||
| C1 | 472.53 a (6.24) | 10.63 a (0.62) | 114.37 a (10.62) | 67.32 a (2.24) | 488.80 a (5.06) | 45 c (2) | 46 b (3.03) |
| C2 | 468.79 a (4.50) | 7.63 b (0.50) | 82.18 b (9.76) | 67.32 a (7.09) | 440.67 b (2.79) | 62 b (4) | 58 b (3.39) |
| C3 | 441.04 b (6.10) | 5.70 c (0.40) | 120.33 a (6.61) | 69.39 a (2.25) | 455.20 b (14.67) | 78 a (5) | 80 a (3.89) |
| P. taeda | |||||||
| C1 | 421.34 a (18.47) | 8.07 a (0.52) | 90.04 b (4.69) | 8.33 b (2.42) | 450.00 a (9.00) | 53 a (4) | 56 a (2.74) |
| C2 | 451.12 a (8.44) | 9.27 a (0.92) | 126.77 a (12.88) | 22.31 a (2.90) | 469.07 a (1.54) | 50 a (4) | 51 a (5.39) |
| C3 | 431.61 a (9.51) | 7.07 a (0.62) | 108.88 ab (8.91) | 23.45 a (3.35) | 475.87 a (13.96) | 62 a (4) | 68 a (6.28) |
| Variable | E. grandis | P. taeda |
|---|---|---|
| Accumulated precipitation | −0.24 p = 0.0540 | −0.23 p = 0.0694 |
| Mean relative humidity | −0.41 p = 0.0007 | −0.46 p = 0.0002 |
| Mean minimum relative humidity | −0.61 p < 0.0001 | −0.64 p < 0.0001 |
| Mean maximum relative humidity | −0.48 p = 0.0001 | −0.50 p < 0.0001 |
| Mean temperature | 0.04 p = 0.7794 | 0.13 p = 0.3184 |
| Mean minimum temperature | −0.07 p = 0.5599 | 0.03 p = 0.9815 |
| Mean maximum temperature | 0.02 p = 0.8895 | 0.07 p = 0.5841 |
| Mean vapor pressure deficit | 0.37 p = 0.0027 | 0.41 p = 0.0010 |
| E. grandis1 | P. taeda1 | |||||||
|---|---|---|---|---|---|---|---|---|
| Layers | Ci | Cf | Ci | Cf | Ci | Cf | Ci | Cf |
| (g kg−1) | (Mg ha−1) | (g kg−1) | (Mg ha−1) | |||||
| C1 | 472.53 a (18.47) | 463.37 a (10.90) | 0.90 a (0.05) | 0.41 b (0.03) | 421.34 a (6.24) | 384.79 a (3.80) | 0.93 a (0.04) | 0.43 b (0.08) |
| C2 | 468.79 a (8.44) | 435.68 a (25.53) | 1.77 a (0.01) | 0.82 b (0.08) | 451.12 a (4.50) | 421.30 a (9.97) | 1.07 a (0.06) | 0.57 b (0.09) |
| C3 | 441.04 a (9.51) | 443.53 a (21.95) | 1.44 a (0.01) | 0.79 b (0.16) | 431.61 a (6.10) | 433.91 a (3.85) | 2.21 a (0.09) | 1.24 b (0.03) |
| Depth | NP | E. grandis | P. taeda | Comparisons | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Initial | Final | Initial | Final | NP vs. EI 1 | NP vs. PI 1 | NP vs. EF 1 | NP vs. PF 1 | EI vs. PI 1 | EF vs. PF 1 | EI vs. EF 2 | PI vs. PF 2 | ||
| (cm) | (‰) | ||||||||||||
| 0–3 | −18.03 (0.07) | −22.75 (0.56) | −24.15 (0.22) | −23.72 (0.06) | −23.37 (0.25) | 0.0141 | <0.0001 | <0.0001 | <0.0001 | 0.2290 | 0.0793 | 0.0679 | 0.2707 |
| 3–6 | −17.53 (0.06) | −19.22 (0.48) | −19.83 (0.32) | −19.91 (0.37) | −20.78 (0.39) | 0.0744 | 0.0033 | 0.0022 | 0.0145 | 0.3195 | 0.1341 | 0.5271 | 0.0020 |
| 6–9 | −16.78 (0.26) | −18.60 (0.10) | −18.83 (0.27) | −18.92 (0.32) | −19.38 (0.43) | 0.0029 | 0.0065 | 0.0055 | 0.0067 | 0.3856 | 0.3379 | 0.3161 | 0.2173 |
| 9–12 | −16.26 (0.42) | −17.69 (0.32) | −17.92 (0.46) | −18.46 (0.60) | −18.64 (0.23) | 0.0534 | 0.0400 | 0.0558 | 0.0077 | 0.3204 | 0.2316 | 0.2819 | 0.6665 |
| 12–20 | −14.87 (0.11) | −17.14 (0.23) | −17.47 (0.43) | −17.89 (0.51) | −17.99 (0.48) | 0.0008 | 0.0045 | 0.0044 | 0.0032 | 0.2533 | 0.4640 | 0.3747 | 0.7397 |
| 0–20 | −16.24 (0.05) | −18.95 (0.16) | −19.78 (0.20) | −19.63 (0.44) | −19.75 (0.31) | 0.0001 | 0.0165 | 0.0001 | 0.0004 | 0.2159 | 0.9246 | 0.0320 | 0.5496 |
| Depth | NP | E. grandis | P. taeda | Comparisons | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Initial | Final | Initial | Final | NP vs. EI 1 | NP vs. PI 1 | NP vs. EF 1 | NP vs. PF 1 | EI vs. PI 1 | EF vs. PF 1 | EI vs. EF 2 | PI vs. PF 2 | ||
| (cm) | (g kg−1) | ||||||||||||
| 0–3 | 15.43 (3.06) | 15.77 (1.94) | 18.91 (1.05) | 10.71 (1.84) | 9.44 (1.13) | 0.9299 | 0.2565 | 0.3425 | 0.1398 | 0.1317 | 0.0036 | 0.2113 | 0.3282 |
| 3–6 | 13.80 (3.13) | 9.86 (1.35) | 10.13 (0.70) | 7.53 (0.60) | 6.98 (1.13) | 0.3117 | 0.1206 | 0.3164 | 0.1096 | 0.1907 | 0.0768 | 0.7368 | 0.5198 |
| 6–9 | 11.35 (3.24) | 9.00 (0.79) | 8.93 (0.81) | 7.08 (0.87) | 7.65 (0.62) | 0.5195 | 0.2716 | 0.5086 | 0.3250 | 0.1763 | 0.2807 | 0.6718 | 0.4496 |
| 9–12 | 10.26 (3.14) | 8.42 (0.11) | 8.34 (0.40) | 6.50 (0.46) | 6.91 (0.95) | 0.6177 | 0.3580 | 0.6058 | 0.3646 | 0.0153 | 0.2387 | 0.8603 | 0.5885 |
| 12–20 | 8.57 (2.07) | 7.74 (0.19) | 8.64 (0.53) | 6.25 (0.48) | 6.39 (0.45) | 0.7282 | 0.3358 | 0.9755 | 0.3617 | 0.0453 | 0.0320 | 0.1281 | 0.4123 |
| 0–20 | 11.13 (2.79) | 9.43 (0.60) | 10.18 (0.63) | 7.28 (0.65) | 7.23 (0.75) | 0.5834 | 0.2502 | 0.7569 | 0.2479 | 0.0719 | 0.0400 | 0.1050 | 0.6468 |
| Depth | NP | E. grandis | P. taeda | Comparisons | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Initial | Final | Initial | Final | NP vs. EI 1 | NP vs. PI 1 | NP vs. EF 1 | NP vs. PF 1 | EI vs. PI 1 | EF vs. PF 1 | EI vs. EF 2 | PI vs. PF 2 | ||
| (cm) | (Mg ha−1) | ||||||||||||
| 0–3 | 6.07 (1.20) | 6.20 (0.76) | 7.43 (0.41) | 4.21 (0.72) | 3.71 (0.44) | 0.9299 | 0.2562 | 0.2876 | 0.2347 | 0.1317 | 0.0036 | 0.2113 | 0.3282 |
| 3–6 | 5.54 (1.29) | 3.60 (0.61) | 3.57 (0.30) | 3.04 (0.25) | 2.82 (0.46) | 0.2453 | 0.1295 | 0.2431 | 0.1406 | 0.4458 | 0.2468 | 0.9376 | 0.5199 |
| 6–9 | 4.85 (1.42) | 3.83 (0.33) | 3.79 (0.36) | 3.04 (0.37) | 3.28 (0.27) | 0.5232 | 0.2831 | 0.4971 | 0.3432 | 0.1820 | 0.3247 | 0.5953 | 0.4496 |
| 9–12 | 4.31 (1.33) | 3.55 (0.05) | 3.51 (0.18) | 2.73 (0.19) | 2.90 (0.40) | 0.6257 | 0.3596 | 0.5262 | 0.3259 | 0.0138 | 0.2395 | 0.8200 | 0.5901 |
| 12–20 | 8.51 (2.10) | 7.62 (0.19) | 8.49 (0.52) | 6.15 (0.48) | 6.30 (0.45) | 0.7148 | 0.3341 | 0.5319 | 0.2047 | 0.0330 | 0.0460 | 0.1237 | 0.3978 |
| 0–20 | 29.28 (7.33) | 24.80 (1.58) | 26.79 (1.67) | 19.17 (1.70) | 19.01(1.99) | 0.5834 | 0.2502 | 0.6310 | 0.2303 | 0.0719 | 0.0400 | 0.1050 | 0.6468 |
| Depth | E. grandis | P. taeda | Comparisons 1 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| α (2017) | α (2019) | Dif. | α (2017) | α (2019) | Dif. | EI vs. PI 1 | EF vs. PF 1 | EI vs. EF 2 | PI vs. PF 2 | E Dif vs. P Dif 1 | |
| (cm) | (Mg ha−1) | ||||||||||
| 0–3 | 2.37 (0.05) | 3.78 (0.11) | 1.42 (0.15) | 2.12 (0.37) | 1.73 (0.15) | −0.38 (0.22) | 0.5688 | 0.0004 | 0.0112 | 0.2270 | 0.0026 |
| 3–6 | 0.46 (0.10) | 0.66 (0.13) | 0.21 (0.22) | 0.62 (0.13) | 0.77 (0.16) | 0.15 (0.09) | 0.3724 | 0.6330 | 0.4389 | 0.2396 | 0.8116 |
| 6–9 | 0.52 (0.03) | 0.58 (0.02) | 0.06 (0.03) | 0.51 (0.10) | 0.69 (0.19) | 0.18 (0.10) | 0.9298 | 0.6110 | 0.2290 | 0.1991 | 0.2957 |
| 9–12 | 0.37 (0.07) | 0.43 (0.09) | 0.06 (0.05) | 0.45 (0.05) | 0.53 (0.11) | 0.08 (0.10) | 0.3906 | 0.5031 | 0.3630 | 0.5071 | 0.8921 |
| 12–20 | 1.14 (0.14) | 1.46 (0.32) | 0.32 (0.19) | 1.24 (0.14) | 1.32 (0.14) | 0.08 (0.11) | 0.6490 | 0.7062 | 0.2365 | 0.5385 | 0.3298 |
| 0–20 | 4.86 (0.15) | 6.92 (0.64) | 2.06 (0.49) | 4.95 (0.71) | 5.05 (0.53) | 0.11 (0.23) | 0.9086 | 0.0873 | 0.0527 | 0.6851 | 0.0232 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baietto, A.; Hernández, J.; del Pino, A. Comparative Dynamics of Above-Ground Litter Production and Decomposition from Eucalyptus grandis Hill ex Maiden and Pinus taeda L., and Their Contribution to Soil Organic Carbon. Forests 2021, 12, 349. https://doi.org/10.3390/f12030349
Baietto A, Hernández J, del Pino A. Comparative Dynamics of Above-Ground Litter Production and Decomposition from Eucalyptus grandis Hill ex Maiden and Pinus taeda L., and Their Contribution to Soil Organic Carbon. Forests. 2021; 12(3):349. https://doi.org/10.3390/f12030349
Chicago/Turabian StyleBaietto, Andrés, Jorge Hernández, and Amabelia del Pino. 2021. "Comparative Dynamics of Above-Ground Litter Production and Decomposition from Eucalyptus grandis Hill ex Maiden and Pinus taeda L., and Their Contribution to Soil Organic Carbon" Forests 12, no. 3: 349. https://doi.org/10.3390/f12030349
APA StyleBaietto, A., Hernández, J., & del Pino, A. (2021). Comparative Dynamics of Above-Ground Litter Production and Decomposition from Eucalyptus grandis Hill ex Maiden and Pinus taeda L., and Their Contribution to Soil Organic Carbon. Forests, 12(3), 349. https://doi.org/10.3390/f12030349