
Long-Term Tree-Ring Response to Drought and Frost in Two Pinus halepensis Populations Growing under Contrasting Environmental Conditions in Peninsular Italy




Abstract
1. Introduction
2. Materials and Methods
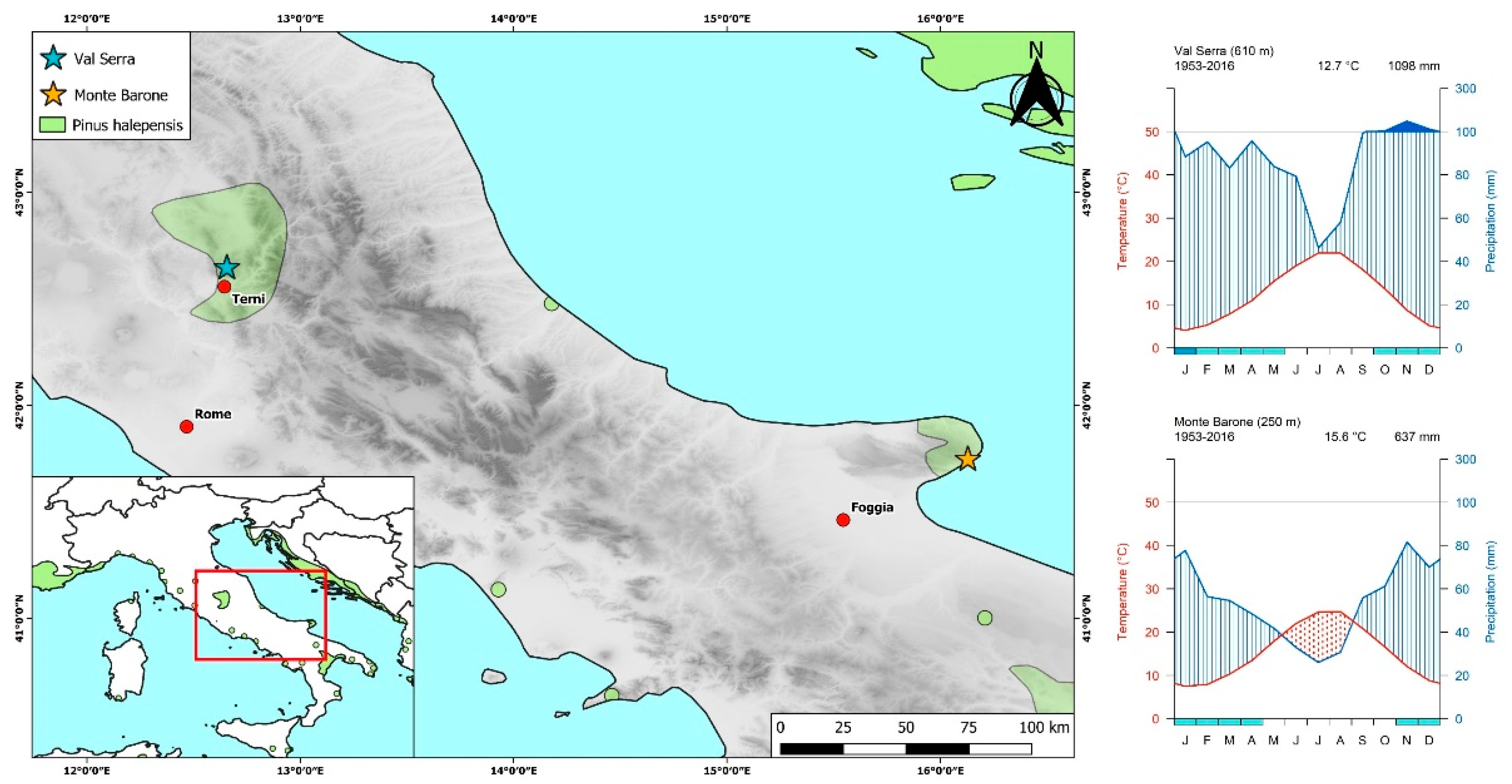
2.1. Study Sites
2.2. Tree-Ring Data Collection and Analysis
2.3. The Climate-Growth Relationships
2.4. Satellite Data Analysis
3. Results
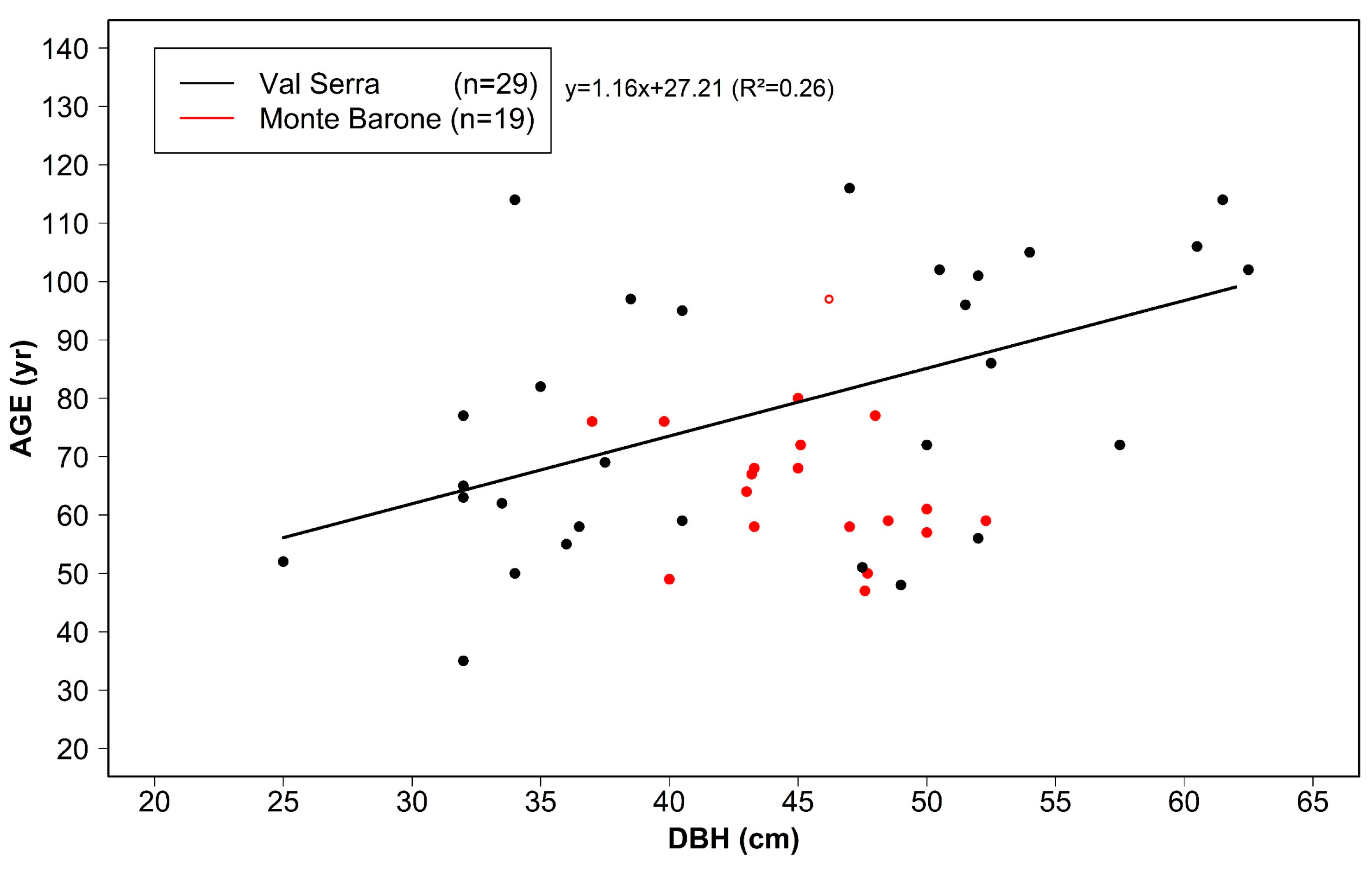
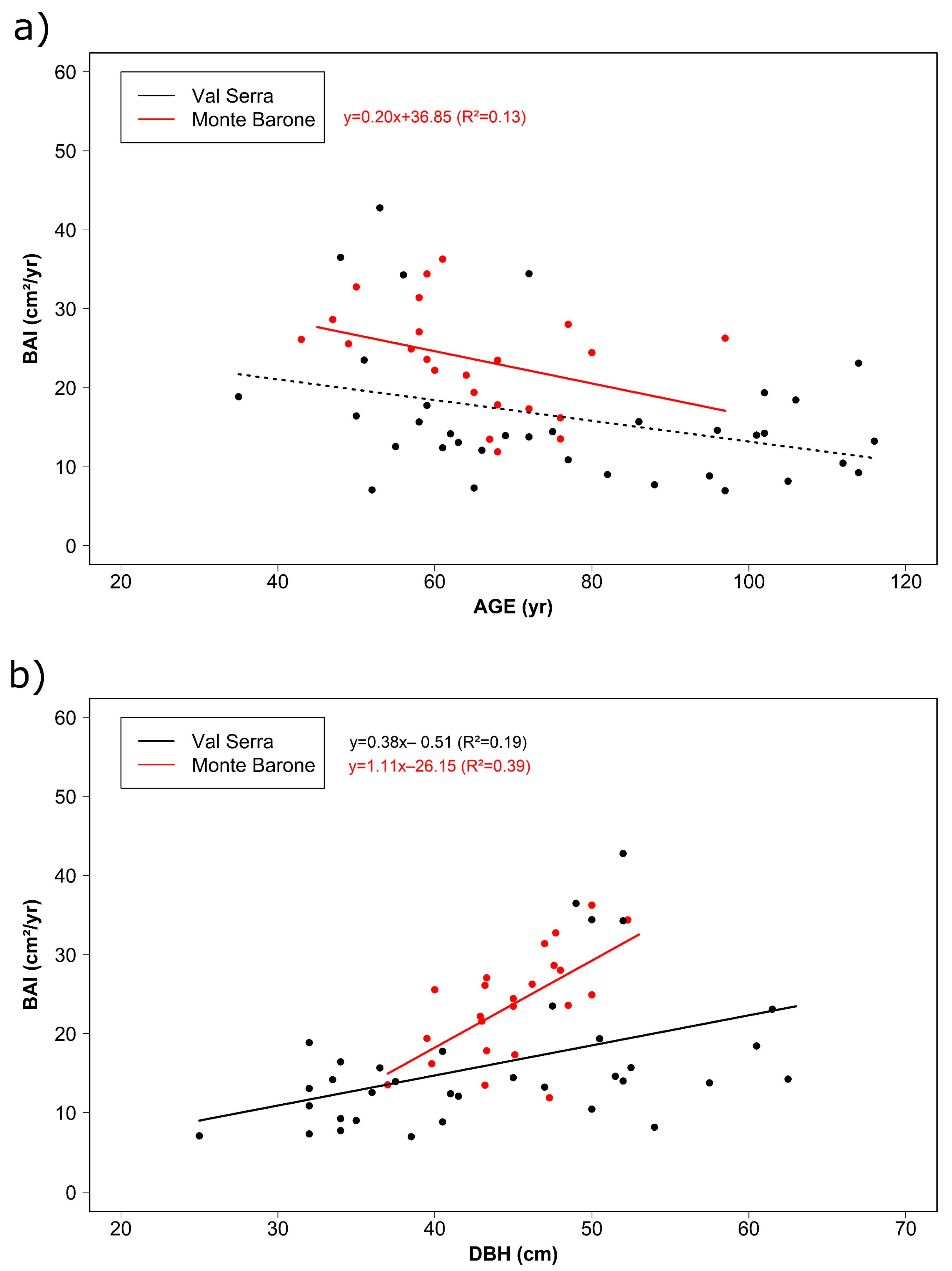
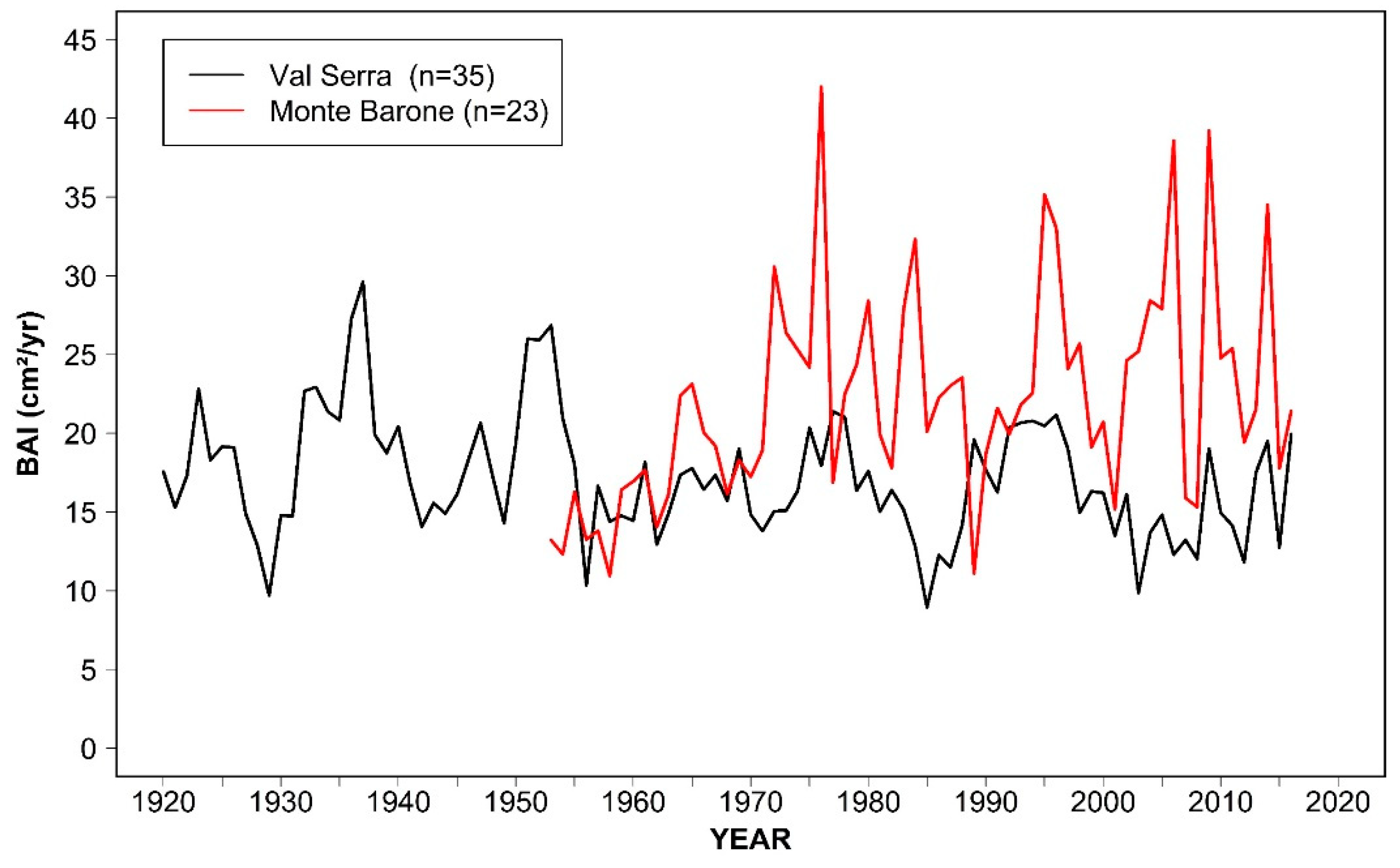
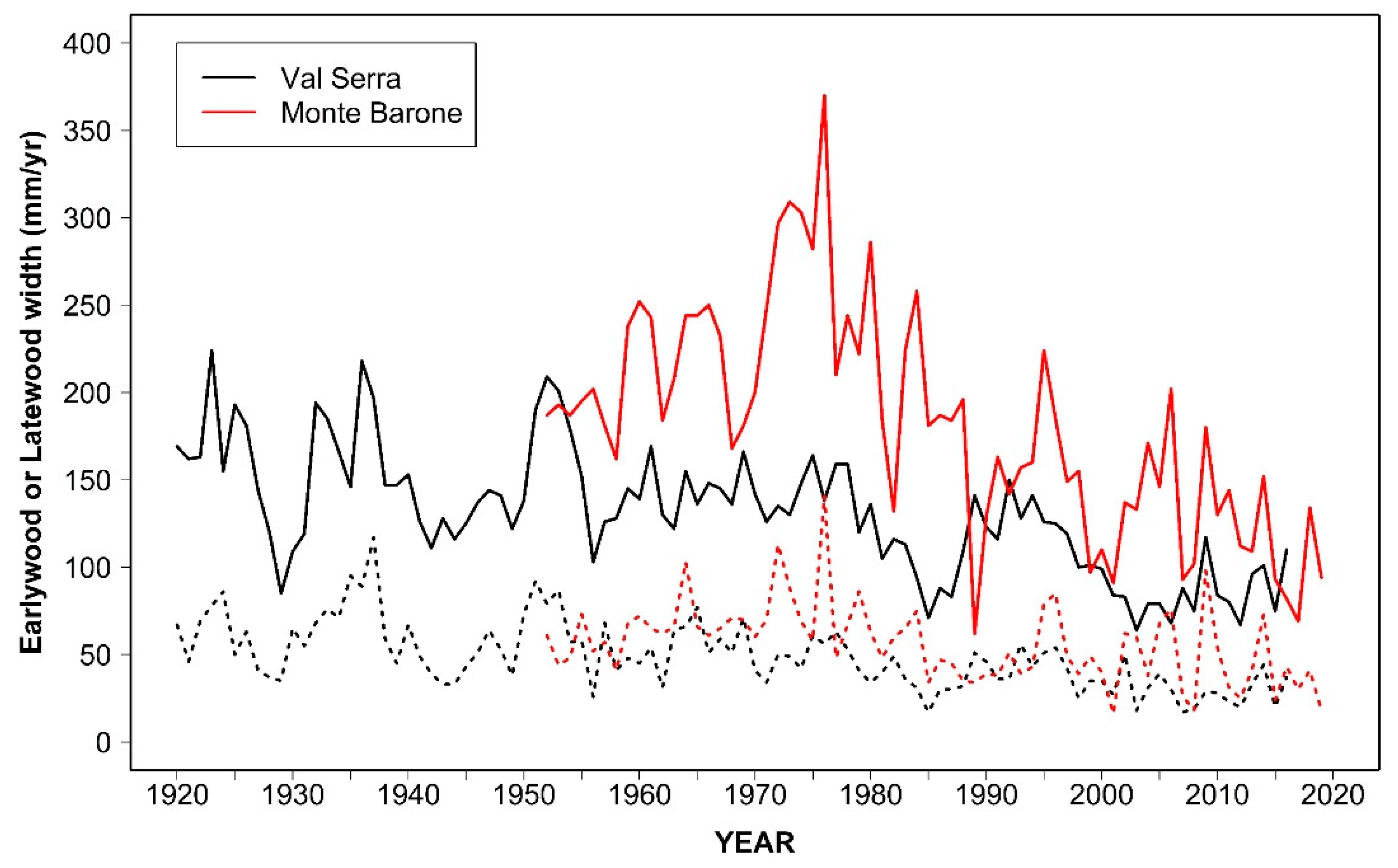
3.1. Tree Age, Size, and Growth Rates
3.2. Climate-Growth Relationships
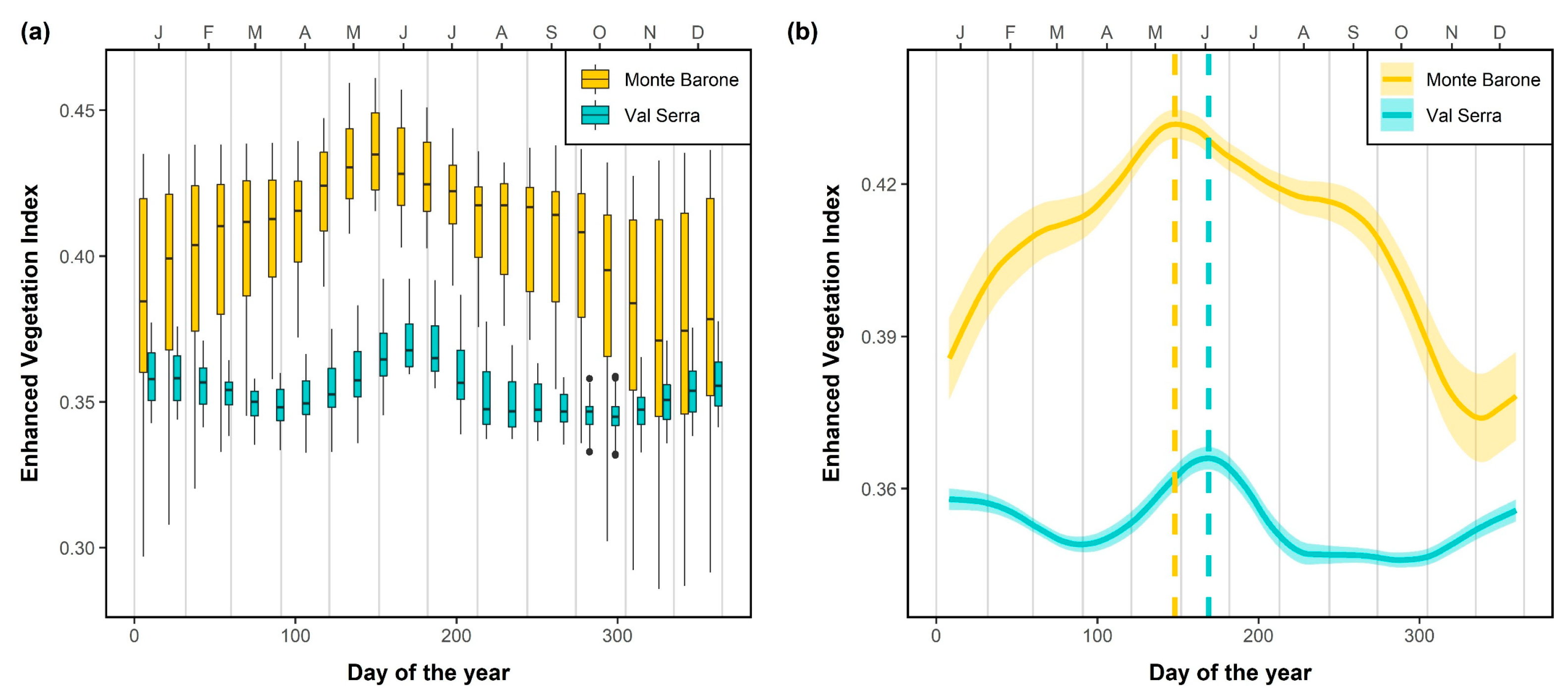
3.3. Satellite Data Analysis
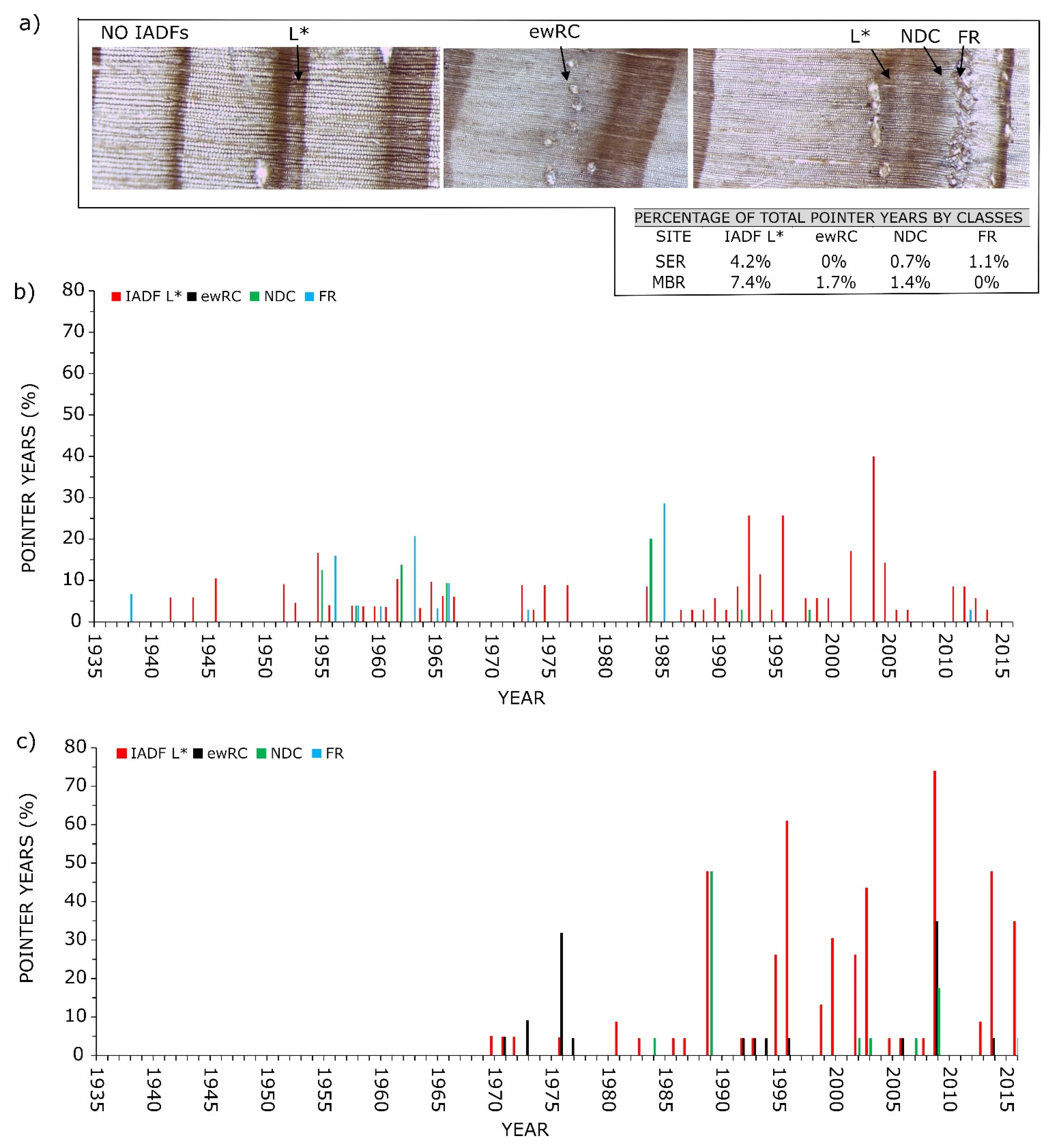
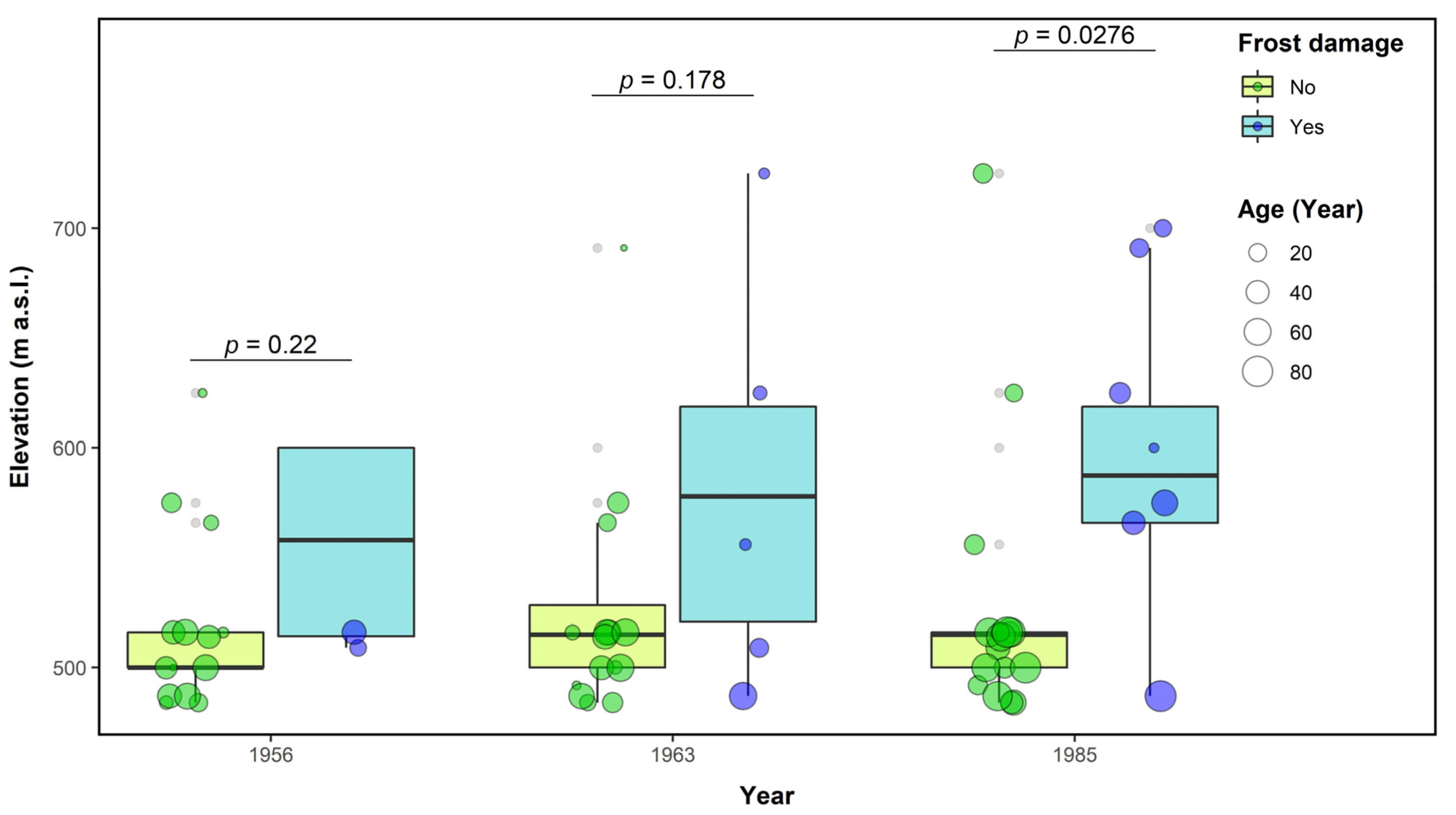
3.4. Pointer Years and Frost Events
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cuttelod, A.; García, N.; Malak, D.A.; Temple, H.; Katariya, V. The Mediterranean: A biodiversity hotspot under threat. In Wildlife in a Changing World—An Analysis of the 2008 IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2008. [Google Scholar]
- Hoerling, M.; Eischeid, J.; Perlwitz, J.; Quan, X.; Zhang, T.; Pegion, P. On the increased frequency of mediterranean drought. J. Clim. 2012, 25, 2146–2161. [Google Scholar] [CrossRef]
- Lionello, P.; Scarascia, L. The relation between climate change in the Mediterranean region and global warming. Reg. Environ. Chang. 2018, 18, 1481–1493. [Google Scholar] [CrossRef]
- Sánchez, E.; Gallardo, C.; Gaertner, M.A.; Arribas, A.; Castro, M. Future climate extreme events in the Mediterranean simulated by a regional climate model: A first approach. Glob. Planet. Chang. 2004, 44, 163–180. [Google Scholar] [CrossRef]
- Chambel, M.R.; Climent, J.; Pichot, C.; Ducci, F. Mediterranean Pines (Pinus halepensis Mill. and brutia Ten.). In Forest Tree Breeding in Europe; Springer: Dordrecht, The Netherlands, 2013; ISBN 978-94-007-6145-2. [Google Scholar]
- De Luis, M.; Čufar, K.; Di Filippo, A.; Novak, K.; Papadopoulos, A.; Piovesan, G.; Rathgeber, C.B.K.; Raventós, J.; Saz, M.A.; Smith, K.T. Plasticity in dendroclimatic response across the distribution range of Aleppo pine (Pinus halepensis). PLoS ONE 2013, 8, e83550. [Google Scholar] [CrossRef] [PubMed]
- Borghetti, M.; Cinnirella, S.; Magnani, F.; Saracino, A. Impact of long-term drought on xylem embolism and growth in Pinus halepensis Mill. Trees Struct. Funct. 1998, 12, 187–195. [Google Scholar] [CrossRef]
- Klein, T. Eco-Physiology of Water Use in Pinus halepensis: From Leaf to Forest Scale. Ph.D. Thesis, Scientific Council of the Weizmann Institute of Science, Rehovot, Israel, 2012. [Google Scholar]
- Hover, A.; Buissart, F.; Caraglio, Y.; Heinz, C.; Pailler, F.; Ramel, M.; Vennetier, M.; Prévosto, B.; Sabatier, S. Growth phenology in Pinus halepensis Mill.: Apical shoot bud content and shoot elongation. Ann. For. Sci. 2017, 74. [Google Scholar] [CrossRef]
- Pardos, M.; Climent, J.; Gil, L.; Pardos, J.A. Shoot growth components and flowering phenology in grafted Pinus halepensis Mill. Trees Struct. Funct. 2003, 17, 442–450. [Google Scholar] [CrossRef]
- Prislan, P.; Gričar, J.; de Luis, M.; Novak, K.; Del Castillo, E.M.; Schmitt, U.; Koch, G.; Štrus, J.; Mrak, P.; Žnidarič, M.T.; et al. Annual cambial rhythm in Pinus halepensis and Pinus sylvestris as indicator for climate adaptation. Front. Plant. Sci. 2016, 7, 1923. [Google Scholar] [CrossRef]
- Baquedano, F.J.; Valladares, F.; Castillo, F.J. Phenotypic plasticity blurs ecotypic divergence in the response of Quercus coccifera and Pinus halepensis to water stress. Eur. J. For. Res. 2008, 127, 495–506. [Google Scholar] [CrossRef]
- Camarero, J.J.; Olano, J.M.; Parras, A. Plastic bimodal xylogenesis in conifers from continental Mediterranean climates. New Phytol. 2010, 185, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Campelo, F.; Nabais, C.; Freitas, H.; Gutiérrez, E. Climatic significance of tree-ring width and intra-annual density fluctuations in Pinus pinea from a dry Mediterranean area in Portugal. Ann. For. Sci. 2007, 64, 229–238. [Google Scholar] [CrossRef]
- Zalloni, E.; de Luis, M.; Campelo, F.; Novak, K.; De Micco, V.; Di Filippo, A.; Vieira, J.; Nabais, C.; Rozas, V.; Battipaglia, G. Climatic signals from intra-annual density fluctuation frequency in mediterranean pines at a regional scale. Front. Plant. Sci. 2016, 7, 579. [Google Scholar] [CrossRef] [PubMed]
- Novak, K.; de Luis, M.; Saz, M.A.; Longares, L.A.; Serrano-Notivoli, R.; Raventós, J.; Čufar, K.; Gričar, J.; Di Filippo, A.; Piovesan, G.; et al. Missing rings in Pinus halepensis—The missing link to relate the tree-ring record to extreme climatic events. Front. Plant. Sci. 2016, 7, 727. [Google Scholar] [CrossRef] [PubMed]
- Fritts, H.C.; Swetnam, T.W. Dendroecology: A tool for evaluating variations in past and present forest environments. Adv. Ecol. Res. 1989, 19, 111–188. [Google Scholar]
- Piermattei, A.; Crivellaro, A.; Carrer, M.; Urbinati, C. The “blue ring”: Anatomy and formation hypothesis of a new tree-ring anomaly in conifers. Trees Struct. Funct. 2015, 29, 613–620. [Google Scholar] [CrossRef]
- Montwé, D.; Isaac-Renton, M.; Hamann, A.; Spiecker, H. Cold adaptation recorded in tree rings highlights risks associated with climate change and assisted migration. Nat. Commun. 2018, 9, 1–7. [Google Scholar] [CrossRef]
- Barinov, V.V.; Myglan, V.S.; Taynik, A.V.; Oydupa, O.C.; Vaganov, E.A. Extreme climatic events in the Republic of Tuva according to tree-ring analysis. Contemp. Probl. Ecol. 2015, 8, 414–422. [Google Scholar] [CrossRef]
- Pedrotti, F. Notizia di ricerche fitosociologiche sulle pinete a Pino d’Aleppo della valle del serra (Terni). Mitt. Ostalp. Pflanz. Arb. 1967, 7, 139–142. [Google Scholar]
- Caudullo, G.; Welk, E.; San-Miguel-Ayanz, J. Chorological maps for the main European woody species. Data Br. 2017, 12, 662–666. [Google Scholar] [CrossRef]
- Novak, K.; Čufar, K.; De Luis, M.; Sánchez, M.A.S.; Raventós, J. Age, climate and intra-annual density fluctuations in Pinus halepensis in Spain. IAWA J. 2013, 34, 459–474. [Google Scholar] [CrossRef]
- Di Filippo, A.; Biondi, F.; Maugeri, M.; Schirone, B.; Piovesan, G. Bioclimate and growth history affect beech lifespan in the Italian Alps and Apennines. Glob. Chang. Biol. 2012, 18, 960–972. [Google Scholar] [CrossRef]
- Bunn, A.; Korpela, M.; Biondi, F.; Campelo, F.; Mérian, P.; Qeadan, F.; Zang, C.; Buras, A.; Cecile, J.; Mudelsee, M.; et al. (Eds.) Package “dplR”, Version 1.7.1. In Dendrochronology Program Library in R; R Foundation: Vienna, Austria, 2015. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Di Filippo, A.; Biondi, F.; Čufar, K.; De Luis, M.; Grabner, M.; Maugeri, M.; Presutti Saba, E.; Schirone, B.; Piovesan, G. Bioclimatology of beech (Fagus sylvatica L.) in the Eastern Alps: Spatial and altitudinal climatic signals identified through a tree-ring network. J. Biogeogr. 2007, 34, 1873–1892. [Google Scholar] [CrossRef]
- Brunetti, M.; Maugeri, M.; Nanni, T.; Simolo, C.; Spinoni, J. High-resolution temperature climatology for Italy: Interpolation method intercomparison. Int. J. Climatol. 2014, 34, 1278–1296. [Google Scholar] [CrossRef]
- Louis, J.; Debaecker, V.; Pflug, B.; Main-Knorn, M.; Bieniarz, J.; Mueller-Wilm, U.; Cadau, E.; Gascon, F. Sentinel-2 SEN2COR: L2A processor for users. Eur. Sp. Agency (Spec. Publ.) ESA SP 2016, SP-740, 9–13. [Google Scholar]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Aybar, C.; Wu, Q.; Bautista, L.; Yali, R.; Barja, A. rgee: An R package for interacting with Google Earth Engine. J. Open Source Softw. 2020, 5, 2272. [Google Scholar] [CrossRef]
- Huete, A.R.; Liu, H.Q.; Batchily, K.; van Leeuwen, W. A comparison of vegetation indices over a global set of TM images for EOS-MODIS. Remote Sens. Environ. 1997, 59, 440–451. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Cai, Z.; Jönsson, P.; Jin, H.; Eklundh, L. Performance of smoothing methods for reconstructing NDVI time-series and estimating vegetation phenology from MODIS data. Remote Sens. 2017, 9, 1271. [Google Scholar] [CrossRef]
- Golub, G.H.; Heath, M.; Wahba, G. Generalized cross-validation as a method for choosing a good ridge parameter. Technometrics 1979, 21, 215–223. [Google Scholar] [CrossRef]
- Aragones, D.; Rodriguez-Galiano, V.F.; Caparros-Santiago, J.A.; Navarro-Cerrillo, R.M. Could land surface phenology be used to discriminate Mediterranean pine species? Int. J. Appl. Earth Obs. Geoinf. 2019, 78, 281–294. [Google Scholar] [CrossRef]
- Karamihalaki, M.; Stagakis, S.; Sykioti, O.; Kyparissis, A.; Parcharidis, I. Monitoring drought effects on mediteranean conifer forests using spot-vegetation NDVI and NDWI timeseries. Eur. Sp. Agency (Spec. Publ.) ESA SP 2016, SP-740, 603. [Google Scholar]
- Maselli, F. Monitoring forest conditions in a protected Mediterranean coastal area by the analysis of multiyear NDVI data. Remote Sens. Environ. 2004, 89, 423–433. [Google Scholar] [CrossRef]
- Zeng, L.; Wardlow, B.D.; Xiang, D.; Hu, S.; Li, D. A review of vegetation phenological metrics extraction using time-series, multispectral satellite data. Remote Sens. Environ. 2020, 237, 111511. [Google Scholar] [CrossRef]
- Chen, R.; Yin, G.; Liu, G.; Li, J.; Verger, A. Evaluation and normalization of topographic effects on vegetation indices. Remote Sens. 2020, 12, 2290. [Google Scholar] [CrossRef]
- Di Filippo, A.; Pederson, N.; Baliva, M.; Brunetti, M.; Dinella, A.; Kitamura, K.; Knapp, H.D.; Schirone, B.; Piovesan, G. The longevity of broadleaf deciduous trees in Northern Hemisphere temperate forests: Insights from tree-ring series. Front. Ecol. Evol. 2015, 3. [Google Scholar] [CrossRef]
- Piovesan, G.; Biondi, F.; Baliva, M.; Dinella, A.; Di Fiore, L.; Marchiano, V.; Presutti Saba, E.; De Vivo, G.; Schettino, A.; Di Filippo, A. Tree growth patterns associated with extreme longevity: Implications for the ecology and conservation of primeval trees in Mediterranean mountains. Anthropocene 2019, 26, 100199. [Google Scholar] [CrossRef]
- Brienen, R.J.W.; Caldwell, L.; Duchesne, L.; Voelker, S.; Barichivich, J.; Baliva, M.; Ceccantini, G.; Di Filippo, A.; Helama, S.; Locosselli, G.M.; et al. Forest carbon sink neutralized by pervasive growth-lifespan trade-offs. Nat. Commun. 2020, 11, 4241. [Google Scholar] [CrossRef]
- Linares, J.C.; Delgado-Huertas, A.; Carreira, J.A. Climatic trends and different drought adaptive capacity and vulnerability in a mixed Abies pinsapo-Pinus halepensis forest. Clim. Chang. 2011, 105, 67–90. [Google Scholar] [CrossRef]
- Stephenson, N.L.; Das, A.J.; Condit, R.; Russo, S.E.; Baker, P.J.; Beckman, N.G.; Coomes, D.A.; Lines, E.R.; Morris, W.K.; Rüger, N.; et al. Rate of tree carbon accumulation increases continuously with tree size. Nature 2014, 507, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Rowland, L.; Da Costa, A.C.L.; Galbraith, D.R.; Oliveira, R.S.; Binks, O.J.; Oliveira, A.A.R.; Pullen, A.M.; Doughty, C.E.; Metcalfe, D.B.; Vasconcelos, S.S.; et al. Death from drought in tropical forests is triggered by hydraulics not carbon starvation. Nature 2015, 528, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Piovesan, G.; Biondi, F.; Di Filippo, A.; Alessandrini, A.; Maugeri, M. Drought-driven growth reduction in old beech (Fagus sylvatica L.) forests of the central Apennines, Italy. Glob. Chang. Biol. 2008, 14, 1265–1281. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Oliva, J.; Vicente-Serrano, S.M. To die or not to die: Early warnings of tree dieback in response to a severe drought. J. Ecol. 2015, 103, 44–57. [Google Scholar] [CrossRef]
- Rathgeber, C.B.K.; Misson, L.; Nicault, A.; Guiot, J. Bioclimatic model of tree radial growth: Application to the French Mediterranean Aleppo pine forests. Trees Struct. Funct. 2005, 19, 162–176. [Google Scholar] [CrossRef]
- Trenberth, K.E.; Fasullo, J.T. An apparent hiatus in global warming? Earth’s Future 2013, 1, 19–32. [Google Scholar] [CrossRef]
- Galli, M.; Guadalupi, M.; Nanni, T.; Ruggiero, L.; Zuanni, F. Ravenna pine trees as monitors of winter severity in N-E Italy. Theor. Appl. Climatol. 1992, 45, 217–224. [Google Scholar] [CrossRef]
- Hadad, M.; Tardif, J.C.; Conciatori, F.; Waito, J.; Westwood, A. Climate and atmospheric circulation related to frost-ring formation in Picea mariana trees from the Boreal Plains, interior North America. Weather Clim. Extrem. 2020, 29, 100264. [Google Scholar] [CrossRef]
- Calamassi, R.; Paoletti, E.; Strati, S. Frost hardening and resistance in three Aleppo pine (Pinus halepensis Mill.) provenances. Isr. J. Plant. Sci. 2001, 49, 179–186. [Google Scholar] [CrossRef]
- Camuffo, D.; Bertolin, C.; Craievich, A.; Granziero, R.; Enzi, S. When the Lagoon was frozen over in Venice from A.D. 604 to 2012: Evidence from written documentary sources, visual arts and instrumental readings. Méditerranée. Rev. Géographique Des Pays Méditerranéens/J. Mediterr. Geogr. 2017. Available online: https://journals.openedition.org/mediterranee/7983 (accessed on 4 March 2021).
- Pignatti, S. I Boschi d'Italia. Sinecologia e Biodiversità; UTET: Torino, Italy, 1998. [Google Scholar]
- Mensing, S.; Tunno, I.; Cifani, G.; Passigli, S.; Noble, P.; Archer, C.; Piovesan, G. Human and climatically induced environmental change in the Mediterranean during the Medieval Climate Anomaly and Little Ice Age: A case from central Italy. Anthropocene 2016, 15, 49–59. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Samples | Latitude (°) | Longitude (°) | Elevation Range (m a.s.l.) | Slope (°) | Aspect |
|---|---|---|---|---|---|---|
| Val Serra | 35 | 42.6448 | 12.6565 | 490–730 | 26.8 | SW |
| Monte Barone | 23 | 41.7450 | 16.1303 | 200–300 | 12.9 | NE |
| Site | N Samples | DBH 1 (cm) | Age 1 (yr) | BAI 1 (cm²/yr) | Earlywood Width ¹ (mm/yr) | Latewood Width ¹ (mm/yr) | EPS > 0.85 | N Trees 2 |
|---|---|---|---|---|---|---|---|---|
| Val Serra | 35 | 44 ± 10 (25–62) | 79 ± 24 (35–122) | 16.1 ± 3.2 (7.0–42.1) | 1.13 ± 0.70 (0.10–7.26) | 0.39 ± 0.24 (0.02–3.12) | 1920–2016 | 10 |
| Monte Barone | 23 | 45 ± 4 (37–52) | 64 ± 12 (43–97) | 23.7 ± 6.6 (12.0–35.8) | 1.79 ± 0.69 (0.15–9.68) | 0.56 ± 0.22 (0.06–4.53) | 1952–2019 | 10 |
| Year Preceding Growth | Year of Growth | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| May | Jun | Jul | Aug | Sep | Oct | Nov | Dec | JAN | FEB | MAR | APR | MAY | JUN | JUL | AUG | SEP | OCT | NOV | DEC | |
| VAL SERRA | ||||||||||||||||||||
| P | 0.10 | −0.03 | −0.14 | −0.30 | −0.06 | 0.13 | −0.05 | 0.08 | −0.11 | −0.09 | −0.13 | 0.05 | 0.10 | 0.29 | 0.21 | 0.24 | 0.18 | 0.06 | −0.05 | 0.09 |
| Tmax | 0.05 | 0.08 | 0.34 | 0.36 | −0.02 | 0.02 | −0.21 | −0.14 | 0.05 | 0.28 | 0.27 | 0.20 | −0.05 | −0.22 | −0.19 | −0.15 | −0.13 | −0.01 | 0.23 | 0.09 |
| Tmin | 0.03 | 0.13 | 0.30 | 0.30 | 0.02 | 0.10 | −0.12 | −0.30 | −0.03 | 0.19 | 0.18 | 0.24 | 0.04 | −0.07 | −0.07 | −0.05 | −0.04 | 0.05 | 0.14 | 0.21 |
| MONTE BARONE | ||||||||||||||||||||
| P | −0.07 | −0.09 | −0.08 | 0.08 | −0.07 | −0.07 | 0.20 | 0.16 | 0.25 | 0.06 | 0.07 | 0.24 | 0.17 | 0.25 | 0.30 | 0.46 | 0.10 | 0.13 | −0.18 | −0.01 |
| Tmax | 0.16 | 0.05 | 0.05 | 0.02 | 0.05 | −0.10 | 0.00 | 0.09 | −0.09 | −0.07 | −0.27 | −0.20 | −0.09 | −0.09 | −0.20 | −0.41 | −0.27 | −0.11 | 0.06 | 0.17 |
| Tmin | 0.09 | 0.10 | 0.14 | 0.10 | 0.08 | −0.15 | 0.12 | 0.22 | 0.09 | 0.04 | −0.21 | 0.00 | −0.08 | −0.05 | −0.05 | −0.31 | −0.11 | −0.12 | −0.04 | 0.23 |
| Year Preceding Growth | Year of Growth | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| May | Jun | Jul | Aug | Sep | Oct | Nov | Dec | JAN | FEB | MAR | APR | MAY | JUN | JUL | AUG | SEP | OCT | NOV | DEC | |
| VAL SERRA | ||||||||||||||||||||
| (a) | ||||||||||||||||||||
| P | 0.08 | 0.00 | −0.13 | −0.12 | 0.06 | 0.02 | −0.04 | 0.05 | −0. 09 | −0.14 | −0.14 | 0.06 | 0.11 | 0.33 | 0.06 | 0.07 | 0.00 | 0.11 | −0.10 | −0.05 |
| Tmax | −0.02 | 0.07 | 0.27 | 0.22 | −0.09 | 0.01 | −0.14 | −0.05 | 0.15 | 0.37 | 0.29 | 0.11 | −0.09 | −0.26 | −0.20 | −0.04 | 0.02 | −0.08 | 0.08 | 0.01 |
| Tmin | −0.01 | 0.16 | 0.25 | 0.19 | 0.03 | 0.11 | −0.09 | −0.16 | 0.12 | 0.29 | 0.20 | 0.15 | 0.07 | −0.06 | −0.13 | 0.04 | 0.05 | 0.03 | −0.05 | 0.05 |
| (b) | ||||||||||||||||||||
| P | 0.15 | 0.05 | −0.08 | −0.27 | −0.09 | 0.08 | −0.01 | 0.03 | −0.07 | −0.01 | −0.17 | 0.00 | 0.03 | 0.11 | 0.38 | 0.49 | 0.27 | −0.07 | −0.03 | 0.19 |
| Tmax | 0.05 | −0.11 | 0.19 | 0.33 | −0.08 | 0.04 | −0.26 | −0.22 | −0.13 | 0.22 | 0.24 | 0.08 | −0.10 | −0.09 | −0.30 | −0.43 | −0.34 | 0.06 | 0.25 | 0.21 |
| Tmin | 0.11 | −0.04 | 0.18 | 0.30 | −0.06 | 0.06 | −0.20 | −0.30 | −0.19 | 0.17 | 0.08 | 0.08 | 0.00 | −0.02 | −0.08 | −0.26 | −0.11 | 0.11 | 0.22 | 0.44 |
| MONTE BARONE | ||||||||||||||||||||
| (a) | ||||||||||||||||||||
| P | −0.15 | −0.07 | −0.05 | 0.09 | −0.06 | −0.01 | 0.20 | 0.13 | 0.23 | 0.16 | 0.10 | 0.26 | 0.18 | 0.19 | 0.09 | 0.26 | −0.10 | −0.01 | −0.06 | −0.03 |
| Tmax | 0.22 | 0.03 | 0.03 | 0.00 | 0.04 | −0.16 | 0.08 | 0.09 | −0.09 | −0.13 | −0.34 | −0.21 | −0.17 | −0.16 | −0.08 | −0.22 | −0.05 | 0.01 | 0.02 | 0.08 |
| Tmin | 0.08 | 0.11 | 0.09 | 0.03 | 0.09 | −0.17 | 0.18 | 0.19 | 0.04 | −0.02 | −0.24 | −0.04 | −0.13 | −0.09 | −0.03 | −0.20 | −0.01 | −0.03 | −0.08 | 0.12 |
| (b) | ||||||||||||||||||||
| P | −0.01 | −0.04 | 0.01 | 0.18 | 0.05 | −0.04 | 0.09 | 0.08 | 0.18 | 0.08 | 0.11 | 0.08 | 0.03 | 0.25 | 0.35 | 0.48 | 0.33 | 0.27 | −0.21 | 0.08 |
| Tmax | 0.08 | −0.02 | 0.05 | −0.07 | −0.05 | −0.06 | 0.01 | 0.02 | −0.09 | −0.01 | −0.27 | −0.10 | 0.11 | 0 | −0.31 | −0.41 | −0.39 | −0.32 | 0.15 | 0.11 |
| Tmin | 0.02 | 0 | 0.10 | 0 | 0.01 | −0.15 | 0 | 0.19 | 0.10 | 0.13 | −0.19 | 0.03 | 0.10 | 0.07 | −0.11 | 0.30 | −0.13 | −0.21 | 0.05 | 0.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Filippo, A.; Baliva, M.; Brunetti, M.; Di Fiore, L. Long-Term Tree-Ring Response to Drought and Frost in Two Pinus halepensis Populations Growing under Contrasting Environmental Conditions in Peninsular Italy. Forests 2021, 12, 305. https://doi.org/10.3390/f12030305
Di Filippo A, Baliva M, Brunetti M, Di Fiore L. Long-Term Tree-Ring Response to Drought and Frost in Two Pinus halepensis Populations Growing under Contrasting Environmental Conditions in Peninsular Italy. Forests. 2021; 12(3):305. https://doi.org/10.3390/f12030305
Chicago/Turabian StyleDi Filippo, Alfredo, Michele Baliva, Michele Brunetti, and Luca Di Fiore. 2021. "Long-Term Tree-Ring Response to Drought and Frost in Two Pinus halepensis Populations Growing under Contrasting Environmental Conditions in Peninsular Italy" Forests 12, no. 3: 305. https://doi.org/10.3390/f12030305
APA StyleDi Filippo, A., Baliva, M., Brunetti, M., & Di Fiore, L. (2021). Long-Term Tree-Ring Response to Drought and Frost in Two Pinus halepensis Populations Growing under Contrasting Environmental Conditions in Peninsular Italy. Forests, 12(3), 305. https://doi.org/10.3390/f12030305