
Climate Change Effects in a Mediterranean Forest Following 21 Consecutive Years of Experimental Drought



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Basal Area Determination
2.4. Data Analysis
3. Results
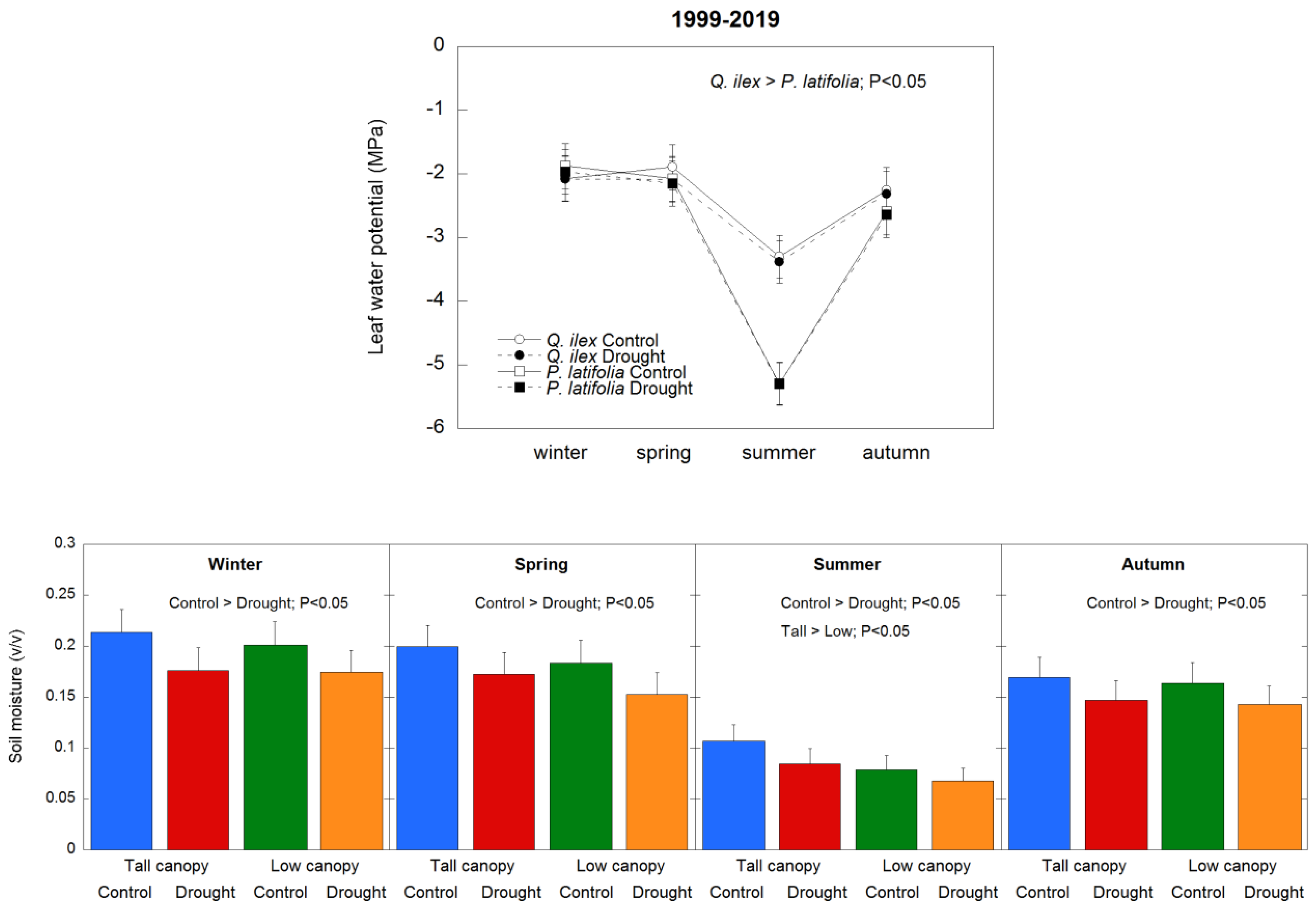
3.1. Soil Moisture and Leaf Water Potential
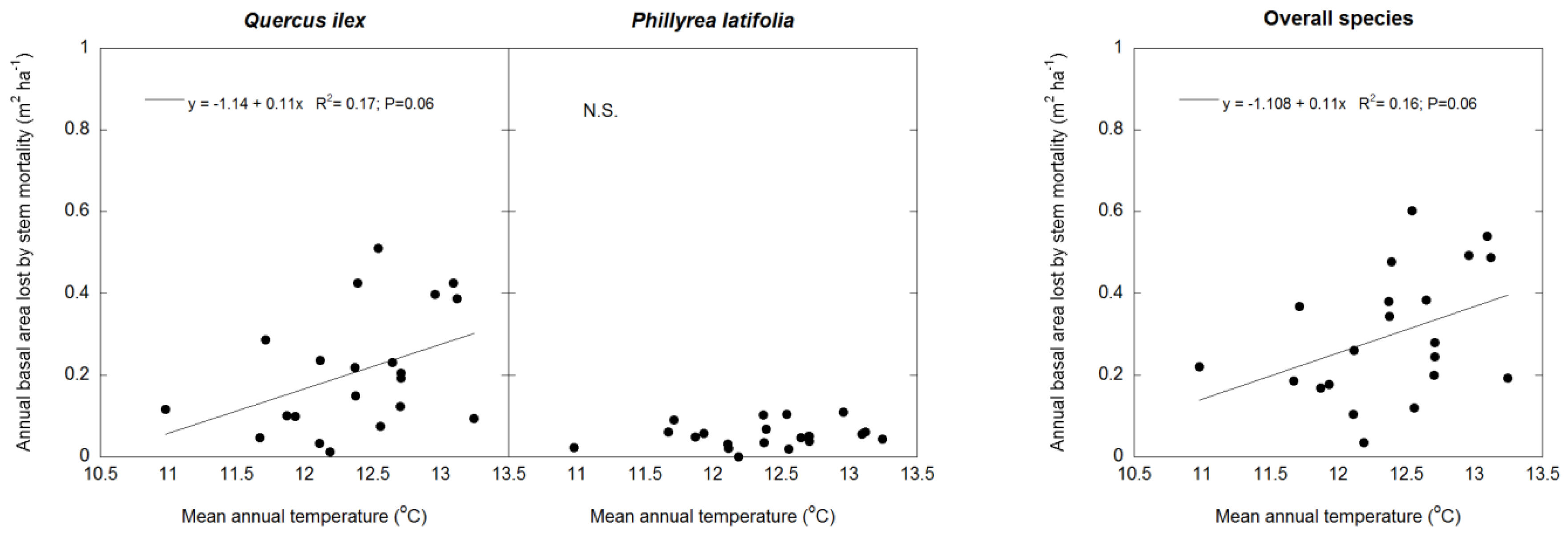
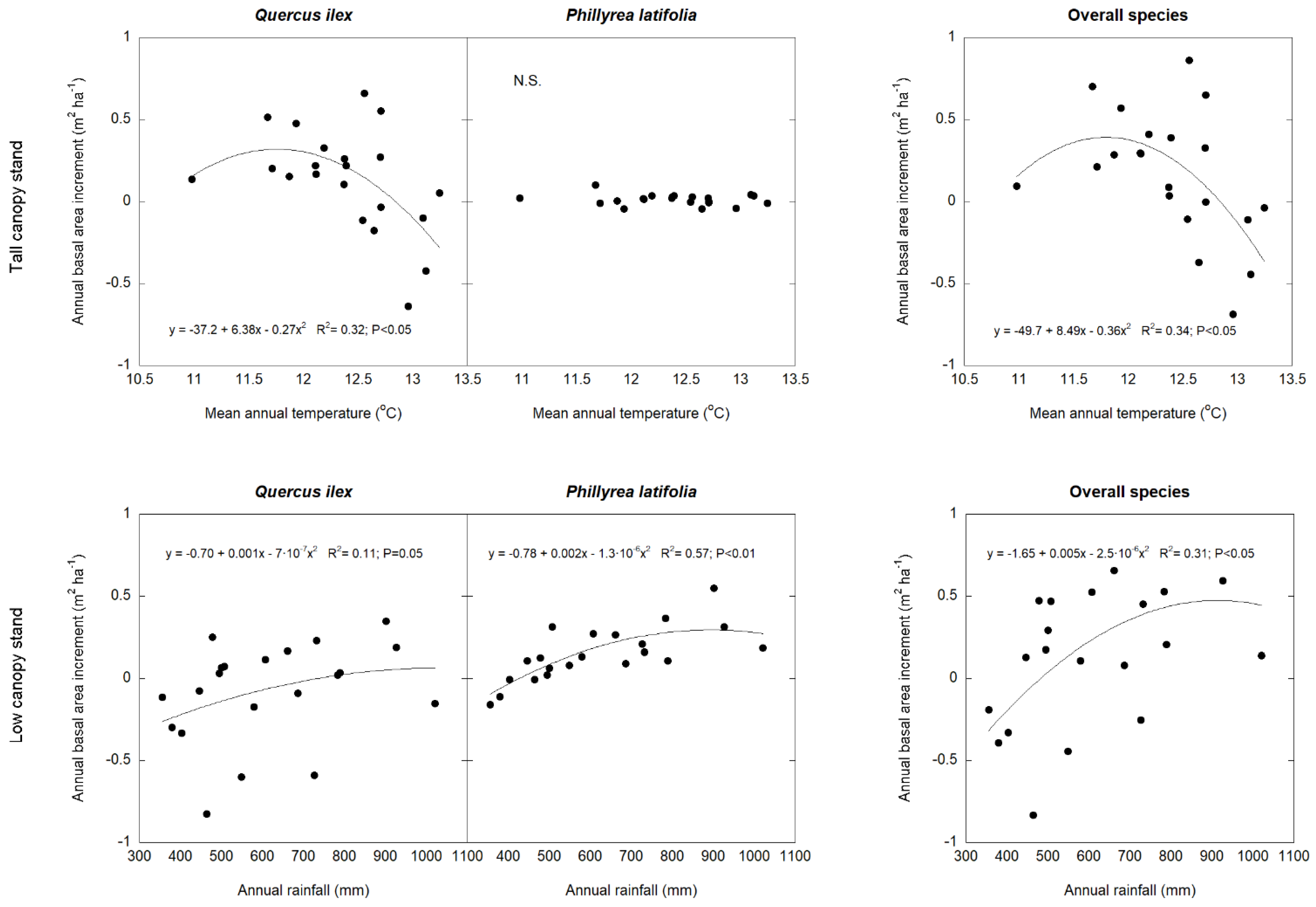
3.2. Climate Effects on Stem Growth
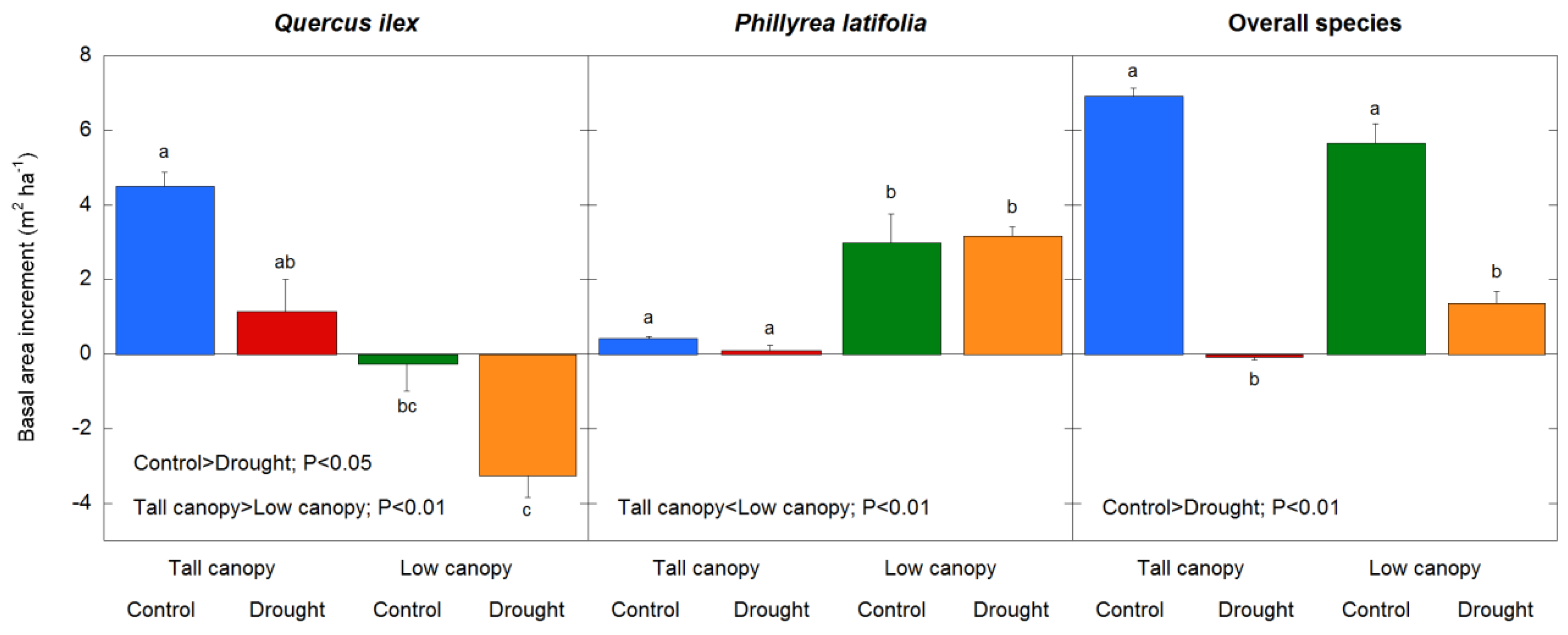
3.3. Drought Effects on Stem Growth
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Plot Type | Basal Area 1999 | Basal Area 2019 |
|---|---|---|---|
| Overall Species | Tall control | 45.83 ± 2.89 | 52.74 ± 2.58 |
| Overall Species | Tall drought | 48.68 ± 3.87 | 48.64 ± 4.02 |
| Overall Species | Low control | 39.19 ± 9.37 | 44.79 ± 10.18 |
| Overall Species | Low drought | 33.25 ± 9.39 | 34.59 ± 9.86 |
| Quercus ilex | Tall control | 37.70 ± 3.67 | 42.18 ± 4.24 |
| Quercus ilex | Tall drought | 37.94 ± 4.91 | 39.08 ± 3.68 |
| Quercus ilex | Low control | 15.99 ± 4.92 | 15.75 ± 3.87 |
| Quercus ilex | Low drought | 19.03 ± 12.14 | 15.78 ± 12.97 |
| Phillyrea latifolia | Tall control | 2.19 ± 1.14 | 2.61 ± 1.23 |
| Phillyrea latifolia | Tall drought | 2.88 ± 0.48 | 2.94 ± 0.23 |
| Phillyrea latifolia | Low control | 15.24 ± 3.37 | 18.20 ± 4.49 |
| Phillyrea latifolia | Low drought | 13.21 ± 2.65 | 16.36 ± 2.29 |
References
- Allen, M.R.; Babiker, M.; Chen, Y.; de Coninck, H.; Connors, S.; van Diemen, R.; Dube, O.P.; Ebi, K.L.; Engelbrecht, F.; Ferrat, M.; et al. IPCC 2018: Summary of Policymakers. In Global Warming of 1.5 °C. An IPCC Special Report on the Impacts of Global Warming of 1. 5 °C Above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Masson-Delmotte, V., Pörtner, H.O., Skea, J., Zhai, P., Roberts, D., Shukla, P.R., Pirani, A., Moufouma-Okia, W., Péan, C., Pidcock, R., et al., Eds.; World Meteorological Organization: Geneva, Switzerland, 2018; p. 32. [Google Scholar]
- Wang, W.; Peng, C.; Kneeshaw, D.D.; Larocque, G.R.; Luo, Z. Drought induced tree mortality: Ecological consequences, causes, and modeling. Environ. Rev. 2012, 20, 109–121. [Google Scholar] [CrossRef]
- Barbeta, A.; Ogaya, R.; Peñuelas, J. Dampening effects of long-term experimental drought on growth and mortality rates of a Holm oak forest. Glob. Chang. Biol. 2013, 19, 3133–3144. [Google Scholar] [CrossRef]
- Liu, D.; Ogaya, R.; Barbeta, A.; Yang, X.; Peñuelas, J. Long-term experimental drought combined with natural extremes accelerate vegetation shift in a Mediterranean holm oak forest. Environ. Exp. Bot. 2018, 151, 1–11. [Google Scholar] [CrossRef]
- Carnicer, J.; Coll, M.; Ninyerola, M.; Pons, X.; Sanchez, G.; Peñuelas, J. Widespread crown condition decline, food web disruption, and amplified tree mortality with increased climate change-type drought. Proc. Natl. Acad. Sci. USA 2011, 108, 1474–1478. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Breshears, D.D.; Cobb, N.S.; Rich, P.M.; Price, K.D.; Allen, C.D.; Balice, R.G.; Romme, W.H.; Kastens, J.H.; Floyd, M.L.; Belnap, J.; et al. Regional vegetation die-off in response to global-change-type drought. Proc. Natl. Acad. Sci. USA 2005, 102, 15144–15148. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.J.P.; Aref, S.; Ho, R.M.; Pippen, J.S.; Hamilton, J.G.; de Lucia, E.H. Annual basal area increment and growth duration of Pinus taeda in response to eight years of free-air carbon dioxide enrichment. Glob. Chang. Biol. 2006, 12, 1367–1377. [Google Scholar] [CrossRef]
- Terradas, J. Holm oak and holm oak forests: An introduction. In Ecology of Mediterranean Evergreen oak Forests; Rodà, F., Retana, J., Gracia, C.A., Bellot, J., Eds.; Springer: Berlin, Germany, 1999; pp. 3–14. [Google Scholar]
- Peñuelas, J.; Filella, I.; Llusià, J.; Siscart, D.; Piñol, J. Comparative field study of spring and summer leaf gas Exchange and photobiology of the Mediterranean trees Quercus ilex and Phillyrea latifolia. J. Exp. Bot. 1998, 49, 229–238. [Google Scholar] [CrossRef]
- Ogaya, R.; Penuelas, J. Wood vs. canopy allocation of aboveground net primary productivity in a Mediterranean forest during 21 years of experimental rainfall exclusion. Forests 2020, 11, 1094. [Google Scholar] [CrossRef]
- Ogaya, R.; Liu, D.; Barbeta, A.; Peñuelas, J. Stem Mortality and Forest Dieback in a 20-Years Experimental Drought in a Mediterranean Holm Oak Forest. Front. For. Glob. Chang. 2020, 2, 89. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J.; Filella, I.; Estiarte, M.; Llusià, J.; Ogaya, R.; Carnicer, J.; Bartrons, M.; Rivas-Ubach, A.; Grau, O.; et al. Impacts of global change on Mediterranean forests and their services. Forests 2017, 8, 463. [Google Scholar] [CrossRef]
- Yin, J.; D’Odorico, P.; Porporato, A. Soil Moisture Dynamics in Water-Limited Ecosystems. In Dryland Ecohydrology; D’Odorico, P., Porporato, A., Wilkinson Runyan, C., Eds.; Springer: Cham, Switzerland, 2019; pp. 31–48. [Google Scholar]
- Kröel-Dulay, G.; Ransijn, J.; Schmidt, I.K.; Beier, C.; de Angelis, P.; de Dato, G.; Dukes, J.S.; Emmett, B.A.; Estiarte, M.; Garadnai, J.; et al. Increased sensitivity to climate change in disturbed ecosystems. Nat. Comm. 2015, 1–7. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, C.; Ogaya, R.; Estiarte, M.; Peñuelas, J. Effects of decadal experimental drought and climate extremes on vegetation growth in Mediterranean forests and shrublands. J. Veg. Sci. 2020, 31, 768–779. [Google Scholar] [CrossRef]
- Leuzinger, S.; Luo, Y.; Beier, C.; Dieleman, W.; Vicca, S.; Körner, C. Do global change experiments overestimate impacts on terrestrial ecosystems? Trends Ecol. Evol. 2011, 26, 236–241. [Google Scholar] [CrossRef]
- Bates, B.C.; Kundzewicz, Z.W.; Wu, S.; Palutikof, J.P. Climate Change and Water. Technical Paper of the Intergovernmental Panel on Climate Change; IPCC Secretariat: Geneva, Switzerland, 2018; p. 210. [Google Scholar]
- Rivas-Ubach, A.; Gargallo-Garriga, A.; Sardans, J.; Oravec, M.; Mateu-Castell, L.; Pérez-Trujillo, M.; Parella, T.; Ogaya, R.; Urban, O.; Peñuelas, J. Drought enhances folivory by shifting foliar metabolomes in Quercus ilex trees. New Phytol. 2014, 202, 874–885. [Google Scholar] [CrossRef]
- Ogaya, R.; Barbeta, A.; Basnou, C.; Peñuelas, J. Satellite data as indicators of tree biomass growth and forest dieback in a Mediterranean holm oak forest. Ann. For. Sci. 2015, 72, 135–144. [Google Scholar] [CrossRef]
- Stephenson, N.L.; Das, A.J.; Condit, R.; Russo, S.E.; Baker, P.J.; Beckman, N.G.; Coomes, D.A.; Lines, E.R.; Morris, W.K.; Rüger, N.; et al. Rate of tree carbon accumulation increases continuously with tree size. Nature 2014, 507, 90–93. [Google Scholar] [CrossRef]
- Trouillier, M.; van der Maaten-Theunissen, M.; Scharnweber, T.; Würth, D.; Burger, A.; Schnittler, M.; Wilmking, M. Size matters–a comparison of three methods to assess age- and size-dependent climate sensitivity of trees. Trees 2019, 33, 183–192. [Google Scholar] [CrossRef]
- Ogaya, R.; Llusià, J.; Barbeta, A.; Asensio, D.; Liu, D.; Alessio, G.A.; Peñuelas, J. Foliar CO2 in a holm oak forest subjected to 15 years of climate change simulation. Plant Sci. 2014, 226, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Ogaya, R.; Peñuelas, J. Contrasting foliar responses to drought in Quercus ilex and Phillyrea latifolia. Biol. Plant. 2006, 50, 373–382. [Google Scholar] [CrossRef]
- Sevanto, S.; McDowell, N.G.; Dickman, L.T.; Pangle, R.; Pockman, W.T. How do trees die? A test of the hydraulic failure and carbon starvation hypotheses. Plant Cell Environ. 2014, 37, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Vilalta, J.; Prat, E.; Oliveras, I.; Piñol, J. Hydraulic properties of roots and stems of nine woody species from a Holm oak forest in NE Spain. Oecologia 2002, 133, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Dijskstra, P.; Koch, G.W.; Peñuelas, J.; Hungate, B.A. Responses of terrestrial ecosystems to temperature and precipitation change: A meta–analysis of experimental manipulation. Glob. Chang. Biol. 2011, 17, 927–942. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J.; Filella, I.; Estiarte, M.; Llusià, J.; Ogaya, R.; Carnicer, J.; Bartrons, M.; Rivas-Ubach, A.; Grau, O.; et al. Assessment of the impacts of climate change on Mediterranean terrestrial ecosystems based on data from field experiments and long-term monitored field gradients in Catalonia. Env. Exp. Bot. 2018, 152, 49–59. [Google Scholar]
- Lloret, F.; Peñuelas, J.; Ogaya, R. Seedling versus sprout recruitment of two Mediterranean trees (Phillyrea latifolia and Quercus ilex) under drought conditions. J. Veg. Sci. 2004, 15, 237–244. [Google Scholar] [CrossRef]
- Ogaya, R.; Peñuelas, J. Changes in leaf δ13C and δ15N and trunk growth for three Mediterranean tree species in relation to soil water availability. Acta Oecol. 2008, 34, 331–338. [Google Scholar] [CrossRef]
- Curiel-Yuste, J.; Peñuelas, J.; Estiarte, M.; Garcia-Mas, J.; Mattana, S.; Ogaya, R.; Pujol, M.; Sardans, J. Drought-resistant fungi control soil organic matter decomposition and its response to temperature. Glob. Chang. Biol. 2011, 17, 1475–1486. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J.; Ogaya, R. Experimental drought reduced acid and alkaline phosphatise activity and increased organic extractable P in soil in a Quercus ilex Mediterranean forest. Eur. J. Soil Biol. 2008, 44, 509–520. [Google Scholar] [CrossRef]
- Ingrisch, J.; Bahn, M. Towards a comparable quantification of resilience. Trends Ecol. Evol. 2018, 33, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Vospernik, S. Basal area increment models accounting for climate and mixture for Austrian tree species. For. Ecol. Manag. 2021, 480, 118725. [Google Scholar] [CrossRef]
- Martín-Benito, D.; Rio, M.; Heinrich, I.; Helle, G.; Cañellas, I. Response of climate-growth relationships and water use efficiency to thinning in a Pinus nigra afforestation. For. Ecol. Manag. 2010, 259, 967–975. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogaya, R.; Peñuelas, J. Climate Change Effects in a Mediterranean Forest Following 21 Consecutive Years of Experimental Drought. Forests 2021, 12, 306. https://doi.org/10.3390/f12030306
Ogaya R, Peñuelas J. Climate Change Effects in a Mediterranean Forest Following 21 Consecutive Years of Experimental Drought. Forests. 2021; 12(3):306. https://doi.org/10.3390/f12030306
Chicago/Turabian StyleOgaya, Romà, and Josep Peñuelas. 2021. "Climate Change Effects in a Mediterranean Forest Following 21 Consecutive Years of Experimental Drought" Forests 12, no. 3: 306. https://doi.org/10.3390/f12030306
APA StyleOgaya, R., & Peñuelas, J. (2021). Climate Change Effects in a Mediterranean Forest Following 21 Consecutive Years of Experimental Drought. Forests, 12(3), 306. https://doi.org/10.3390/f12030306