
Xylem and Phloem Formation Dynamics in Quercus ilex L. at a Dry Site in Southern Italy




Abstract

1. Introduction
2. Materials and Methods
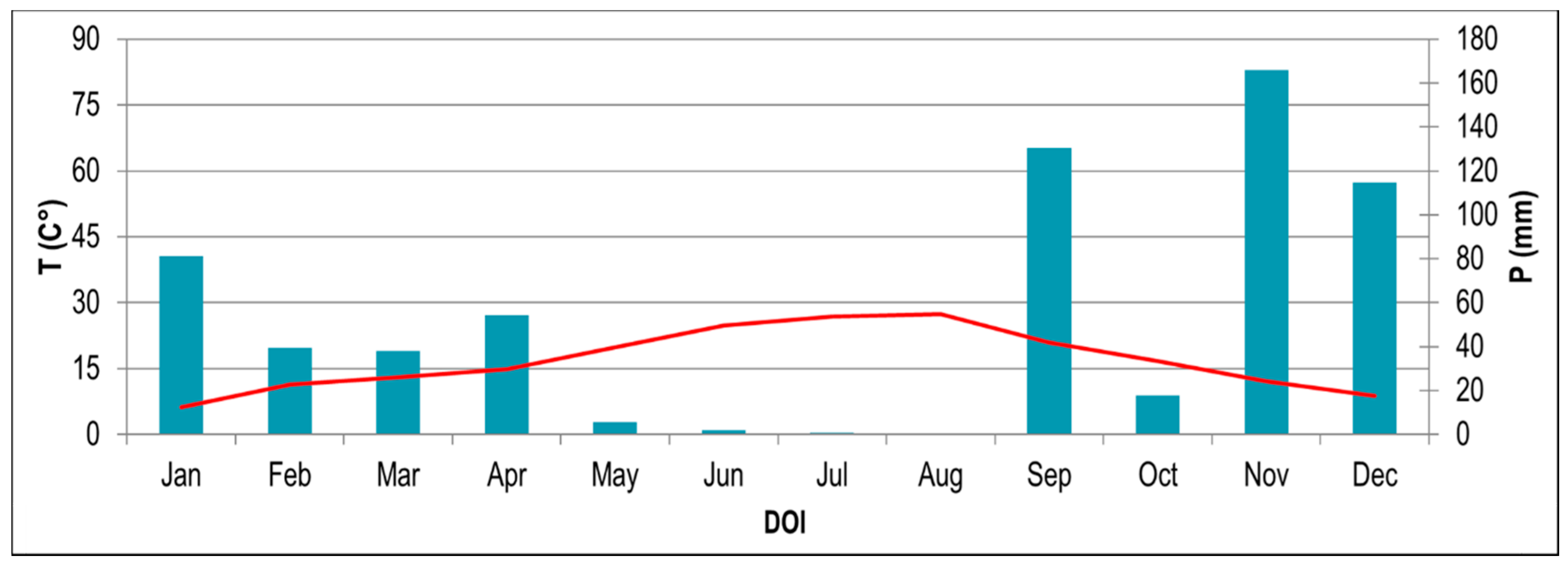
2.1. Study Site
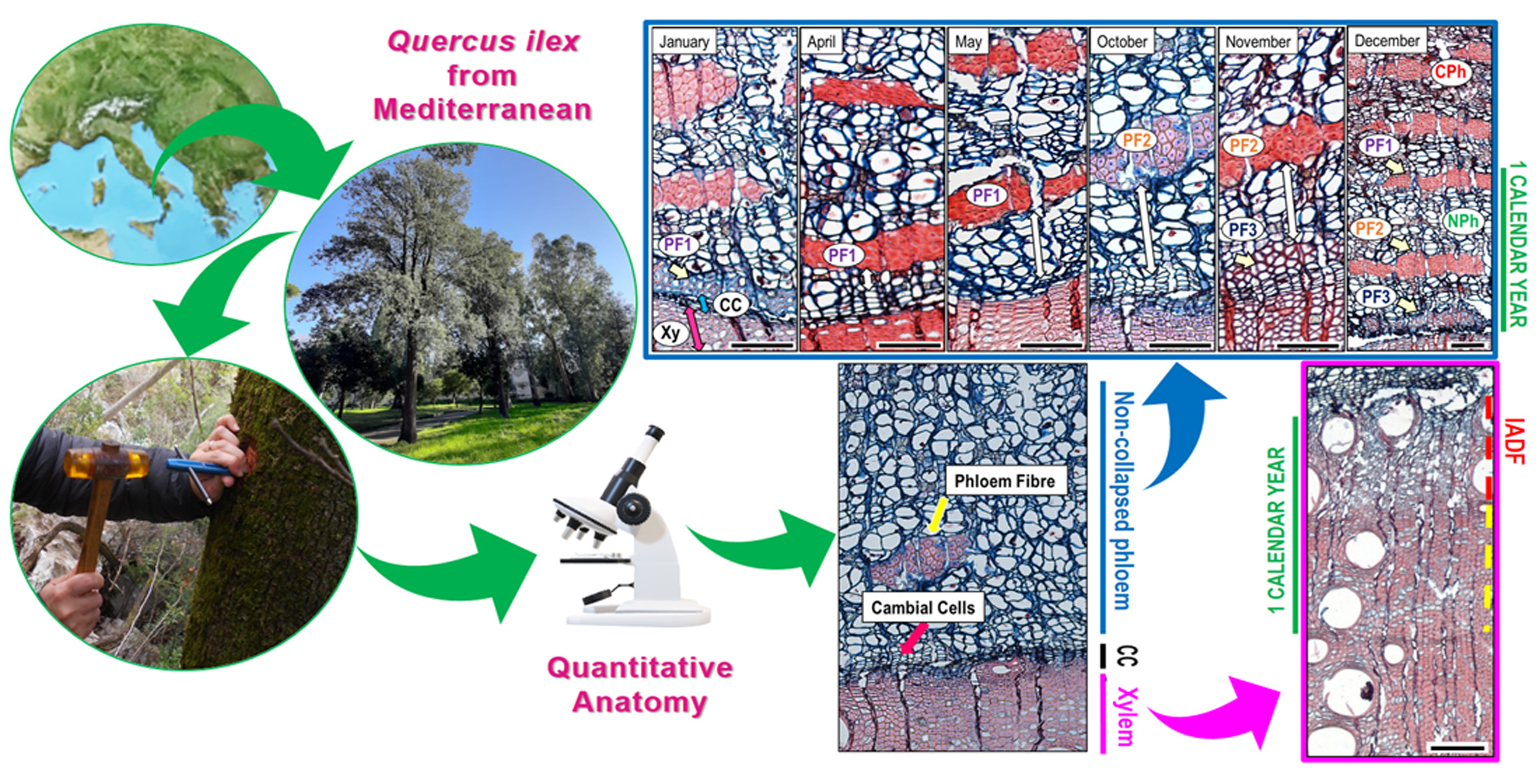
2.2. Sampling, Microscopy, and Quantitative Wood Anatomy
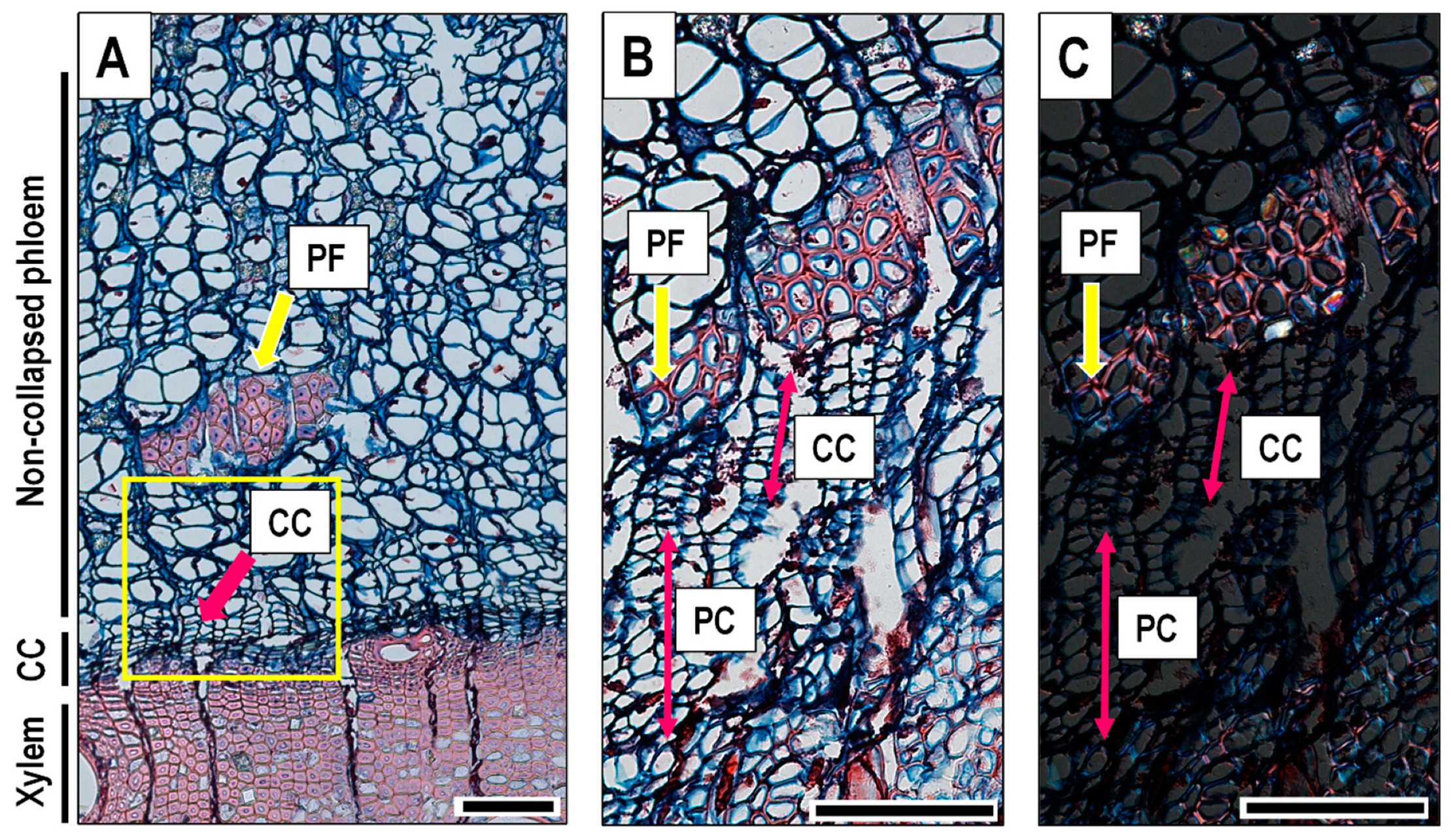
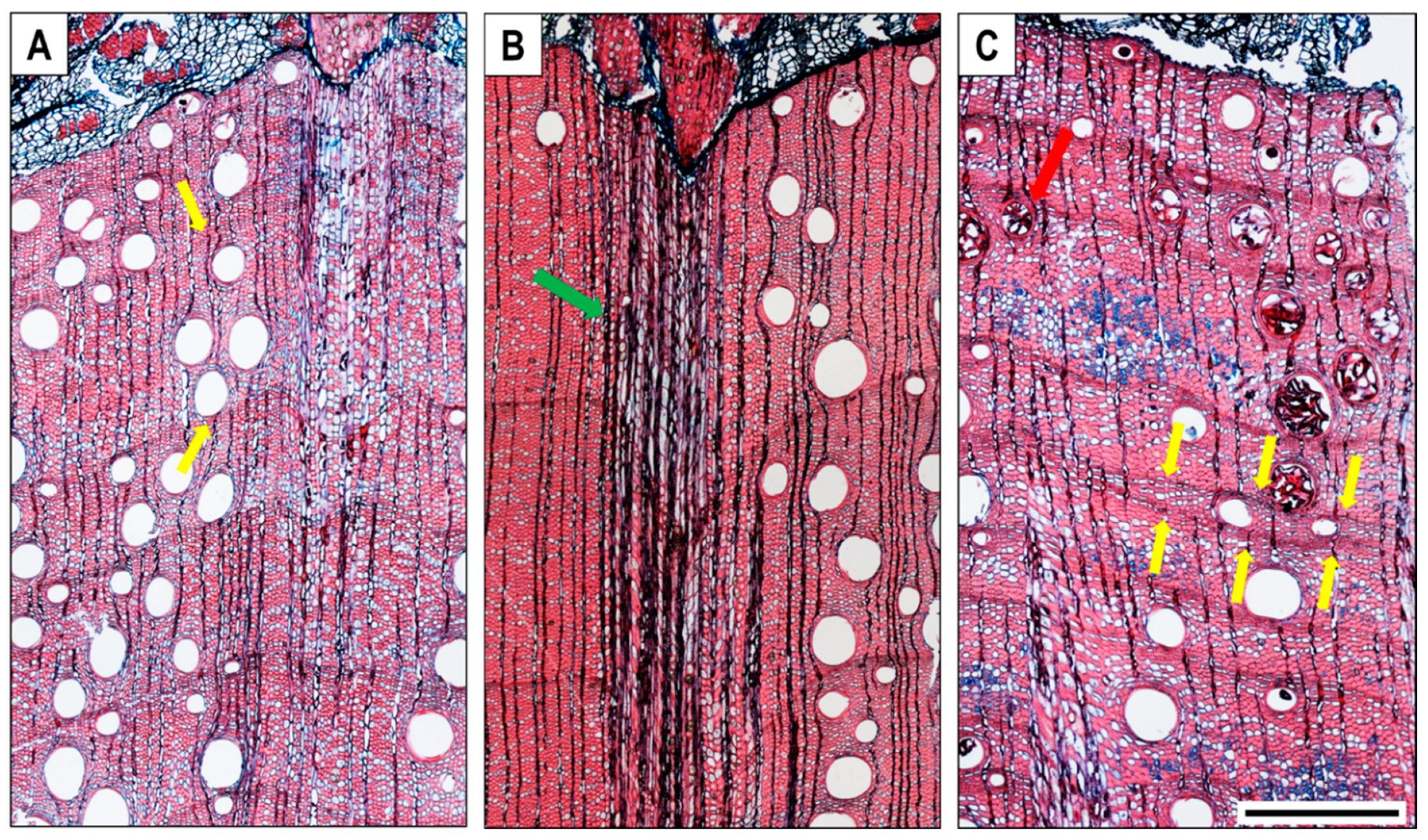
3. Results
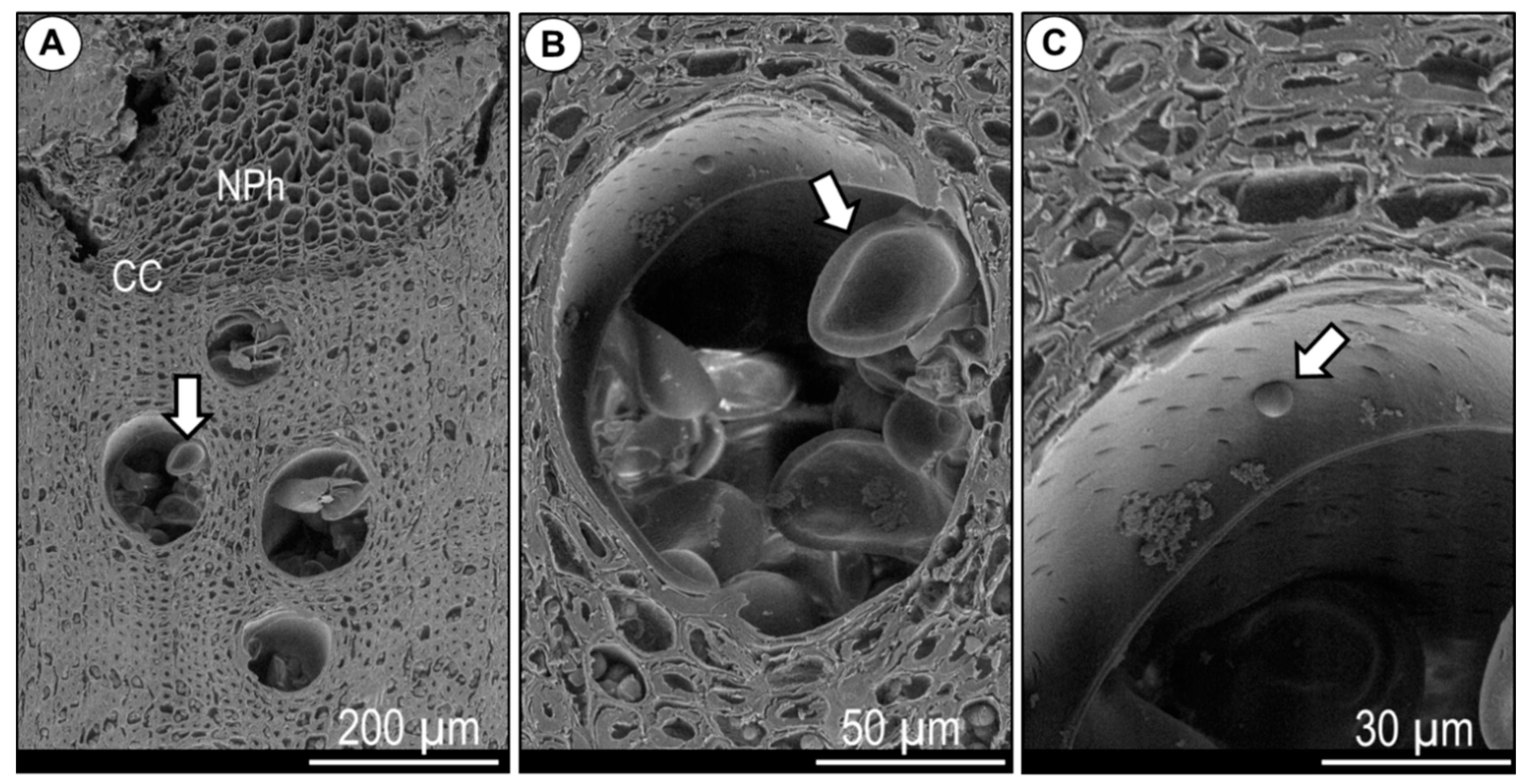
3.1. Xylem Anatomical Features
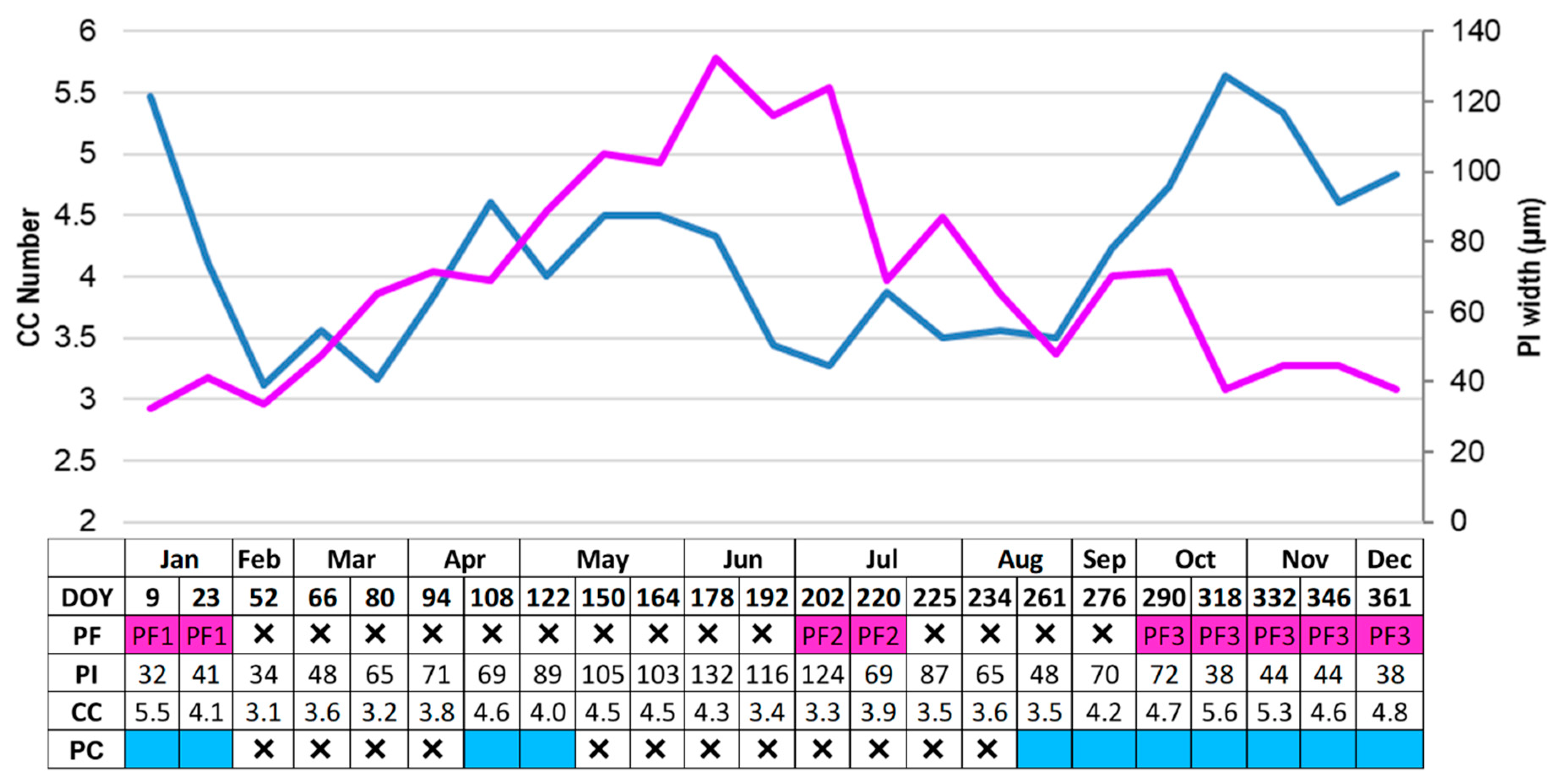
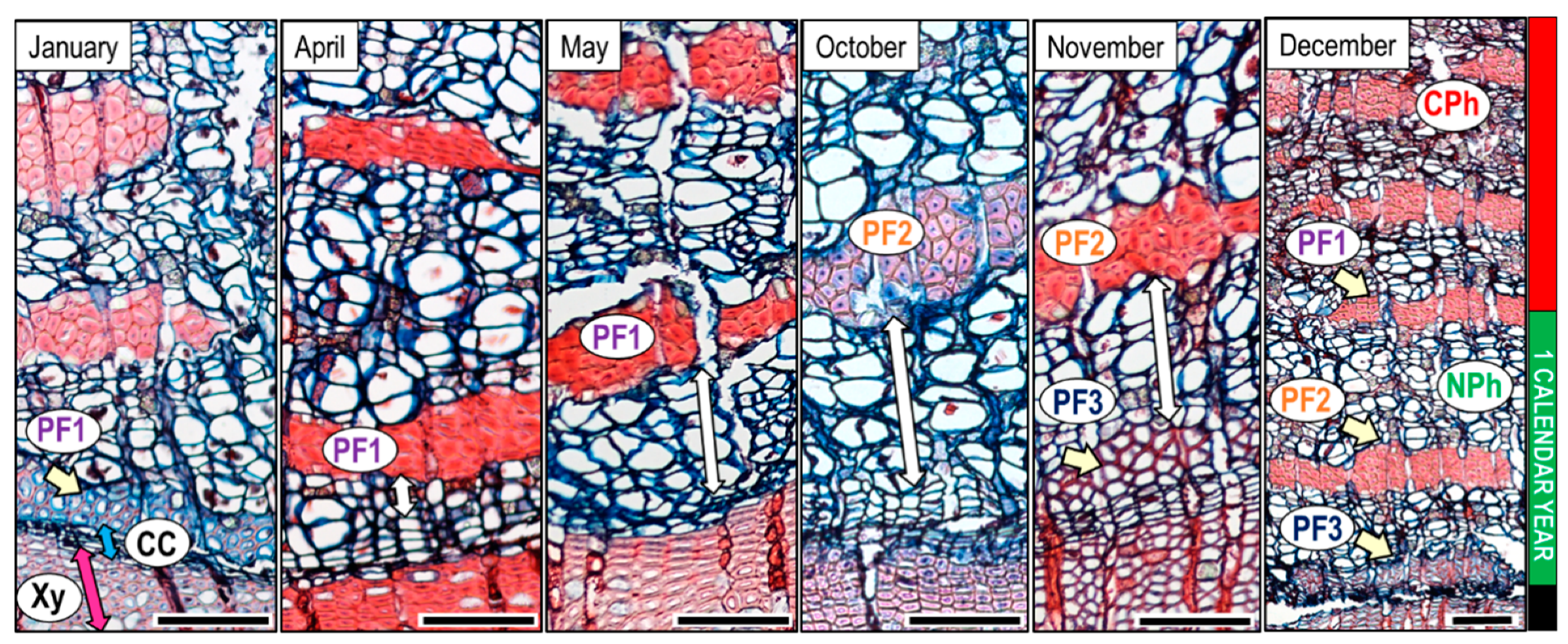
3.2. Cambial Activity and Wood Increments Formed Throughout the Year
4. Discussion
4.1. Cambial Activity and Xylem Increment
4.2. Phloem Formation
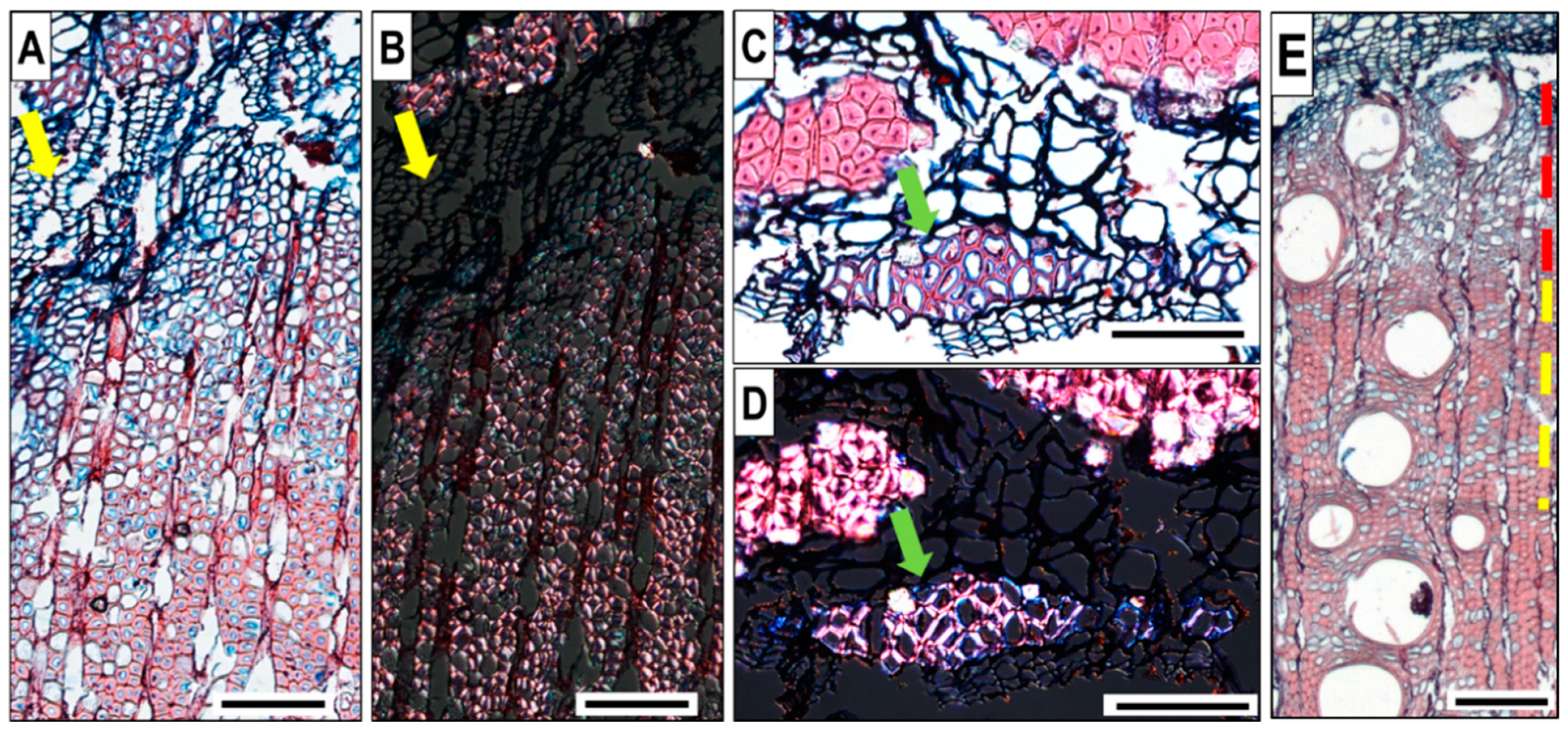
4.3. Technical Notes to Improve Sample Preparation of Q. ilex
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Masson-Delmotte, V.; Zhai, P.; Pörtner, H.O.; Roberts, D.; Skea, J.; Shukla, P.R.; Pirani, A.; Moufouma-Okia, W.; Péan, C.; Pidcock, R.; et al. Global warming of 1.5 C. IPCC Spec. Rep. Impacts Glob. Warm. 1 2018, 1, 1–9. [Google Scholar]
- Ogaya, R.; Liu, D.; Barbeta, A.; Peñuelas, J. Stem mortality and forest dieback in a 20-year experimental drought in a Mediterranean holm oak forest. Front. For. Glob. Chang. 2019, 2, 89. [Google Scholar] [CrossRef]
- De Marco, A.; Arena, C.; Giordano, M.; De Santo, A.V. Impact of the invasive tree black locust on soil properties of Mediterranean stone pine-holm oak forests. Plant Soil 2013, 372, 473–486. [Google Scholar] [CrossRef]
- Barbero, M.; Loisel, R.; Quézel, P. Biogeography, ecology and history of Mediterranean Quercus ilex ecosystems. In Quercus ilex L. Ecosystems: Function, Dynamics and Management; Springer: Dordrecht, The Netherlands, 1992; pp. 19–34. [Google Scholar]
- Canadell, J.; Roda, F. Root biomass of Quercus ilex in a montane Mediterranean forest. Can. J. For. Res. 1991, 21, 1771–1778. [Google Scholar] [CrossRef][Green Version]
- Savé, R.; Castell, C.; Terradas, J. Gas exchange and water relations. In Ecology of Mediterranean Evergreen Oak Forests; Springer: Berlin/Heidelberg, Germany, 1999; pp. 135–147. [Google Scholar]
- Infante, J.M.; Domingo, F.; Alés, R.F.; Joffre, R.; Rambal, S. Quercus ilex transpiration as affected by a prolonged drought period. Biol. Plant 2003, 46, 49–55. [Google Scholar] [CrossRef]
- Tognetti, R.; Longobucco, A.; Raschi, A. Vulnerability of xylem to embolism in relation to plant hydraulic resistance in Quercus pubescens and Quercus ilex co-occurring in a Mediterranean coppice stand in central Italy. New Phytol. 1998, 139, 437–447. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Mangirón, M.; Ogaya, R.; Sauret, M.; Serrano, L.; Peñuelas, J.; Piñol, J. Sap flow of three co-occurring Mediterranean woody species under varying atmospheric and soil water conditions. Tree Physiol. 2003, 23, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Sade, N.; Gebremedhin, A.; Moshellon, M. Risk taking plants. Anisohydric behaviour as a stress resistance trait. Plant Signal Behav. 2012, 7, 767–770. [Google Scholar] [CrossRef]
- Corcuera, L.; Camarero, J.J.; Gil-Pelegrín, E. Effects of a severe drought on Quercus ilex radial growth and xylem anatomy. Trees 2004, 18, 83–92. [Google Scholar] [CrossRef]
- Ogaya, R.; Peñuelas, J. Species-specific drought effects on flower and fruit production in a Mediterranean holm oak forest. Forestry 2007, 80, 351–357. [Google Scholar] [CrossRef]
- Barbeta, A.; Ogaya, R.; Peñuelas, J. Dampening effects of long-term experimental drought on growth and mortality rates of a Holm oak forest. Glob. Chang. Biol. 2013, 19, 3133–3144. [Google Scholar] [CrossRef]
- Liu, D.; Ogaya, R.; Barbeta, A.; Yang, X.; Peñuelas, J. Long-term experimental drought combined with natural extremes accelerate vegetation shift in a Mediterranean holm oak forest. Environ. Exp. Bot. 2018, 151, 1–11. [Google Scholar] [CrossRef]
- Lloret, F.; Siscart, D. Drought demographic effects in Holm Oak populations. Cuad. Soc. Esp. Cien. For. 1995, 2, 77–81. [Google Scholar]
- Peñuelas, J.; Filella, I.; Lloret, F.; Piñol, J.; Siscart, D. Effects of a severe drought on water and nitrogen use by Quercus ilex and Phyllirea latifolia. Biol. Plant 2000, 43, 47–53. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Piñol, J.; Beven, K. A hydraulic model to predict drought-induced mortality in woody plants: An application to climate change in the Mediterranean. Ecol. Model. 2002, 155, 127–147. [Google Scholar] [CrossRef]
- Lorenz, M.; Fischer, R.; Becher, G.; Mues, V.; Seidling, W.; Kraft, P.; Nagel, H.D. Forest Condition in Europe: 2006 Technical Report of ICP Forests (No. 2006/1); Work Report of the Institute for World Forestry: Hamburg, Germany, 2006; Available online: http://hdl.handle.net/10419/96611 (accessed on 1 September 2006).
- Bonal, R.; Muñoz, A. Multi-trophic effects of ungulate intraguild predation on acorn weevils. Oecologia 2007, 152, 533–540. [Google Scholar] [CrossRef] [PubMed]
- De Micco, V.; Carrer, M.; Rathgeber, C.B.K.; Camarero, J.J.; Voltas, J.; Cherubini, P.; Battipaglia, G. From xylogenesis to tree rings: Wood traits to investigate tree response to environmental changes. IAWA J. 2019, 40, 155–182. [Google Scholar] [CrossRef]
- Prislan, P.; Čufar, K.; De Luis, M.; Gričar, J. Precipitation is not limiting for xylem formation dynamics and vessel development in European beech from two temperate forest sites. Tree Physiol. 2018, 38, 186–197. [Google Scholar] [CrossRef]
- Cherubini, P.; Gartner, B.L.; Tognetti, R.; Braeker, O.U.; Schoch, W.; Innes, J.L. Identification, measurement and interpretation of tree rings in woody species from Mediterranean climates. Biol. Rev. 2003, 78, 119–148. [Google Scholar] [CrossRef] [PubMed]
- De Micco, V.; Campelo, F.; De Luis, M.; Bräuning, A.; Grabner, M.; Battipaglia, G.; Cherubini, P. Intra-annual density fluctuations in tree rings: How, when, where, and why? IAWA J. 2016, 37, 232–259. [Google Scholar] [CrossRef]
- Terradas, J.; Savé, R. The influence of summer and winter stress and water relationships on the distribution of Quercus ilex L. In Quercus ilex L. Ecosystems: Function, Dynamics and Management; Springer: Dordrecht, The Netherlands, 1992; pp. 137–145. [Google Scholar]
- Campelo, F.; Gutiérrez, E.; Ribas, M.; Nabais, C.; Freitas, H. Relationships between climate and double rings in Quercus ilex from northeast Spain. Can. J. For. Res. 2007, 37, 1915–1923. [Google Scholar] [CrossRef]
- Gea-Izquierdo, G.; Martín-Benito, D.; Cherubini, P.; Isabel, C. Climate-growth variability in Quercus ilex L. west Iberian open woodlands of different stand density. Ann. For. Sci. 2009, 66, 802. [Google Scholar] [CrossRef]
- Gea-Izquierdo, G.; Cherubini, P.; Cañellas, I. Tree-rings reflect the impact of climate change on Quercus ilex L. along a temperature gradient in Spain over the last 100 years. For. Ecol. Manag. 2011, 262, 1807–1816. [Google Scholar] [CrossRef]
- Gutiérrez, E.; Campelo, F.; Camarero, J.J.; Ribas, M.; Muntán, E.; Nabais, C.; Freitas, H. Climate controls act at different scales on the seasonal pattern of Quercus ilex L. stem radial increments in NE Spain. Trees 2011, 25, 637–646. [Google Scholar] [CrossRef]
- Zalloni, E.; Battipaglia, G.; Cherubini, P.; Saurer, M.; De Micco, V. Contrasting physiological responses to Mediterranean climate variability are revealed by intra-annual density fluctuations in tree rings of Quercus ilex L. and Pinus pinea L. Tree Physiol. 2018, 38, 1213–1224. [Google Scholar] [CrossRef] [PubMed]
- Balzano, A.; Čufar, K.; Battipaglia, G.; Merela, M.; Prislan, P.; Aronne, G.; De Micco, V. Xylogenesis reveals the genesis and ecological signal of IADFs in Pinus pinea L. and Arbutus unedo L. Ann. Bot. 2018, 121, 1231–1242. [Google Scholar] [CrossRef]
- Balzano, A.; De Micco, V.; Merela, M.; Čufar, K. Tree-rings in Mediterranean pines–can we ascribe them to calendar years? Les/Wood 2019, 68, 5–14. [Google Scholar] [CrossRef]
- De Luis, M.; Gričar, J.; Čufar, K.; Raventós, J. Seasonal dynamics of wood formation in Pinus halepensis from dry and semi-arid ecosystems in Spain. IAWA J. 2007, 28, 389–404. [Google Scholar] [CrossRef]
- Novak, K.; De Luis, M.; Gričar, J.; Prislan, P.; Merela, M.; Smith, K.T.; Čufar, K. Missing and dark rings associated with drought in Pinus halepensis. IAWA J. 2016, 37, 260–274. [Google Scholar] [CrossRef]
- De Micco, V.; Balzano, A.; Čufar, K.; Aronne, G.; Gričar, J.; Merela, M.; Battipaglia, G. Timing of false ring formation in Pinus halepensis and Arbutus unedo in Southern Italy: Outlook from an analysis of xylogenesis and tree-ring chronologies. Front. Plant Sci. 2016, 7, 705. [Google Scholar] [CrossRef]
- Balzano, A.; Battipaglia, G.; Cherubini, P.; De Micco, V. Xylem Plasticity in Pinus pinaster and Quercus ilex Growing at Sites with Different Water Availability in the Mediterranean Region: Relations between Intra-Annual Density Fluctuations and Environmental Conditions. Forests 2020, 11, 379. [Google Scholar] [CrossRef]
- Zhang, S.H.; Romane, F. Diameter growth of Quercus ilex L. and the interannual variability of climatic characteristics. Ann. Sci. For. 1991, 48, 225–234. [Google Scholar] [CrossRef]
- Zalloni, E.; Battipaglia, G.; Cherubini, P.; De Micco, V. Site conditions influence the climate signal of intra-annual density fluctuations in tree rings of Q. ilex L. Ann. For. Sci. 2018, 75, 68. [Google Scholar] [CrossRef]
- Čufar, K.; Prislan, P.; De Luis, M.; Gričar, J. Tree-ring variation, wood formation and phenology of beech (Fagus sylvatica) from a representative site in Slovenia, SE Central Europe. Trees 2008, 22, 749–758. [Google Scholar] [CrossRef]
- Prislan, P.; Gričar, J.; De Luis, M.; Smith, K.T.; Čufar, K. Phenological variation in xylem and phloem formation in Fagus sylvatica from two contrasting sites. Agric. For. Metereol. 2013, 180, 142–151. [Google Scholar] [CrossRef]
- Prislan, P.; Gričar, J.; De Luis, M.; Novak, K.; Martinez del Castillo, E.; Schmitt, U.; Koch, G.; Štrus, J.; Mrak, P.; Žnidarič, M.T.; et al. Annual cambial rhythm in Pinus halepensis and Pinus sylvestris as indicator for climate adaptation. Front. Plant Sci. 2016, 7, 1923. [Google Scholar] [CrossRef]
- Prislan, P.; Gričar, J.; Čufar, K.; De Luis, M.; Merela, M.; Rossi, S. Growing season and radial growth predicted for Fagus sylvatica under climate change. Clim. Chang. 2019, 153, 181–197. [Google Scholar] [CrossRef]
- Gričar, J.; Prislan, P.; De Luis, M.; Gryc, V.; Hacurová, J.; Vavrčík, H.; Čufar, K. Plasticity in variation of xylem and phloem cell characteristics of Norway spruce under different local conditions. Front. Plant Sci. 2015, 6, 730. [Google Scholar] [CrossRef]
- Del Castillo, E.M.; Prislan, P.; Gričar, J.; Gryc, V.; Merela, M.; Giagli, K.; De Luis, M.; Vavrčík, H.; Čufar, K. Challenges for growth of beech and co-occurring conifers in a changing climate context. Dendrochronologia 2018, 52, 1–10. [Google Scholar] [CrossRef]
- Gričar, J. Cambial cell production and structure of xylem and phloem as an indicator of tree vitality: A review. In Sustainable Forest Management-Current Research; IntechOpen: Rijeka, Hrvatska, 2012; pp. 111–134. [Google Scholar]
- Gričar, J.; Jagodic, Š.; Šefc, B.; Trajković, J.; Eler, K. Can the structure of dormant cambium and the widths of phloem and xylem increments be used as indicators for tree vitality? Eur. J. For. Res. 2014, 133, 551–562. [Google Scholar] [CrossRef]
- Gričar, J.; Krže, L.; Čufar, K. Number of cells in xylem, phloem and dormant cambium in silver fir (Abies alba), in trees of different vitality. IAWA J. 2009, 30, 121–133. [Google Scholar] [CrossRef]
- Gričar, J.; Prislan, P.; De Luis, M.; Novak, K.; Longares, L.A.; del Castillo, E.M.; Čufar, K. Lack of annual periodicity in cambial production of phloem in trees from Mediterranean areas. IAWA J. 2016, 37, 349–364. [Google Scholar] [CrossRef]
- Balzano, A.; De Micco, V.; Čufar, K.; De Luis, M.; Gričar, J. Intra-seasonal trends in phloem traits in Pinus spp. from drought-prone environments. IAWA J. 2020, 41, 219–235. [Google Scholar] [CrossRef]
- Sökücü, A.; Güney, A. Vitality versus cambium, xylem, and phloem characteristics in Cedrus libani. IAWA J. 2020, 1, 1–15. [Google Scholar] [CrossRef]
- Gričar, J. Xylem and phloem formation in sessile oak from Slovenia in 2007. Wood Res. 2007, 55, 15–22. [Google Scholar]
- Gričar, J.; Jagodic, Š.; Prislan, P. Structure and subsequent seasonal changes in the bark of sessile oak (Quercus petraea). Trees 2015, 29, 747–757. [Google Scholar] [CrossRef]
- Teobaldelli, M.; Cona, F.; Stinca, A.; Saulino, L.; Anzano, E.; Giordano, D.; Migliozzi, A.; Bonanomi, G.; D’Urso, G.; Mazzoleni, S.; et al. Improving resilience of an old-growth urban forest in Southern Italy: Lesson (s) from a stand-replacing windstorm. Urban For. Urban Green. 2020, 47, 126521. [Google Scholar] [CrossRef]
- Ricciardi, M.; Mazzoleni, S.; Pizzolongo, P. Il bosco del parco Gussone in Portici. Aspetti naturalistici e problemi di conservazione. In Parchi e Giardini Storici, Parchi Letterari. Conoscenza, tutela e valorizzazione. Atti III Convegno (primo internazionale)“Paesaggi e Giardini del Mediterraneo”. 2-La valorizzazione; Ministero per i Beni Culturali e Ambientali. Comitato Nazionale per lo studio e la conservazione dei Giardini Storici. Soprintendenza Archeologica di Pompei, Tipolitografia GRG: Salerno, Italy, 1993. [Google Scholar]
- Stinca, A.; Motti, R. The vascular flora of the Royal Park of Portici (Naples, Italy). Webbia 2009, 64, 235–266. [Google Scholar] [CrossRef]
- Rossi, S.; Anfodillo, T.; Menardi, R. Trephor: A new tool for sampling microcores from tree stems. IAWA J. 2006, 27, 89–97. [Google Scholar] [CrossRef]
- Van der Werf, G.W.; Sass-Klaassen, U.G.; Mohren GM, J. The impact of the 2003 summer drought on the intra-annual growth pattern of beech (Fagus sylvatica L.) and oak (Quercus robur L.) on a dry site in the Netherlands. Dendrochronologia 2007, 25, 103–112. [Google Scholar] [CrossRef]
- De Luis, M.; Novak, K.; Raventós, J.; Gričar, J.; Prislan, P.; Čufar, K. Climate factors promoting intra-annual density fluctuations in Aleppo pine (Pinus halepensis) from semiarid sites. Dendrochronologia 2011, 29, 163–169. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Morin, H.; Saracino, A.; Motta, R.; Borghetti, M. Conifers in cold environments synchronize maximum growth rate of tree-ring formation with day length. New Phytol. 2006, 170, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Čufar, K.; Cherubini, M.; Gričar, J.; Prislan, P.; Spina, S.; Romagnoli, M. Xylem and phloem formation in chestnut (Castanea sativa Mill.) during the 2008 growing season. Dendrochronologia 2011, 29, 127–134. [Google Scholar] [CrossRef]
- Trockenbrodt, M. Survey and discussion of the terminology used in bark anatomy. IAWA J. 1990, 11, 141–166. [Google Scholar] [CrossRef]
- Merela, M.; Thaler, N.; Balzano, A.; Plavčak, D. Optimal Surface Preparation for Wood Anatomy Research of Invasive Species by Scanning Electron Microscopy. Drv. Ind. 2020, 71, 117–127. [Google Scholar] [CrossRef]
- Allard, V.; Ourcival, J.M.; Rambal, S.; Joffre, R.; Rocheteau, A. Seasonal and annual variation of carbon exchange in an evergreen Mediterranean forest in southern France. Glob. Chang. Biol. 2008, 14, 714–725. [Google Scholar] [CrossRef]
- Montagnoli, A.; Dumroese, R.K.; Terzaghi, M.; Onelli, E.; Scippa, G.S.; Chiatante, D. Seasonality of fine root dynamics and activity of root and shoot vascular cambium in a Quercus ilex L. forest (Italy). For. Ecol. Manag. 2019, 431, 26–34. [Google Scholar] [CrossRef]
- Zalloni, E.; Battipaglia, G.; Cherubini, P.; Saurer, M.; De Micco, V. Wood growth in pure and mixed Quercus ilex L. forests: Drought influence depends on site conditions. Front. Plant Sci. 2019, 10, 397. [Google Scholar] [CrossRef]
- Güney, A.; Kerr, D.; Sökücü, A.; Zimmermann, R.; Küppers, M. Cambial activity and xylogenesis in stems of Cedrus libani A. Rich at different altitudes. Bot. Stud. 2015, 56, 20. [Google Scholar] [CrossRef] [PubMed]
- Zalloni, E.; de Luis, M.; Campelo, F.; Novak, K.; De Micco, V.; Di Filippo, A.; Vieira, J.; Nabais, C.; Rozas, V.; Battipaglia, G. Climatic signals from intra-annual density fluctuation frequency in Mediterranean pines at a regional scale. Front. Plant Sci. 2016, 7, 579. [Google Scholar] [CrossRef] [PubMed]
- Worbes, M. Wood anatomy and tree-ring structure and their importance for tropical dendrochronology. In Amazonian Floodplain Forests; Springer: Dordrecht, The Netherlands, 2010; pp. 329–346. [Google Scholar] [CrossRef]
- De Micco, V.; Balzano, A.; Wheeler, E.A.; Baas, P. Tyloses and gums: A review of structure, function and occurrence of vessel occlusions. IAWA J. 2016, 37, 186–205. [Google Scholar] [CrossRef]
- Oven, P.; Merela, M.; Mikac, U.; Serša, I. 3D magnetic resonance microscopy of a wounded beech branch. Holzforschung 2008, 62, 322–328. [Google Scholar] [CrossRef]
- Oven, P.; Merela, M.; Mikac, U.; Serša, I. Application of 3D magnetic resonance microscopy to the anatomy of woody tissues. IAWA J. 2011, 32, 401–414. [Google Scholar] [CrossRef]
- Savi, T.; Casolo, V.; Luglio, J.; Bertuzzi, S.; Gullo, M.A.L.; Nardini, A. Species-specific reversal of stem xylem embolism after a prolonged drought correlates to endpoint concentration of soluble sugars. Plant Physiol. Biochem. 2016, 106, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Pagliarani, C.; Casolo, V.; Ashofteh Beiragi, M.; Cavalletto, S.; Siciliano, I.; Schubert, A.; Gullino, M.L.; Zwieniecki, M.A.; Secchi, F. Priming xylem for stress recovery depends on coordinated activity of sugar metabolic pathways and changes in xylem sap pH. Plant Cell Environ. 2019, 42, 1775–1787. [Google Scholar] [CrossRef]
- Dwyer, J.P.; Cutter, B.E.; Wetteroff, J.J. A dendrochronological study of black and scarlet oak decline in the Missouri Ozarks. For. Ecol. Manag. 1995, 75, 69–75. [Google Scholar] [CrossRef]
- Voelker, S.L.; Muzika, R.M.; Guyette, R.P. Individual tree and stand level influences on the growth, vigor, and decline of red oaks in the Ozarks. For. Sci. 2008, 54, 8–20. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree | Total Xylem | Non-Collapsed | Phloem/Xylem | IADF |
|---|---|---|---|---|
| Increment | Phloem Increment | Increment Ratio | Occurrence | |
| Number | µm | µm | Number | |
| 1 | 808.9 | 314.4 | 0.389 | 1 |
| 2 | 682.0 | 260.0 | 0.381 | 1 |
| 3 | 188.0 | 230.0 | 1.223 | 1 |
| 4 | 0.0 | 204.3 | -- | - |
| 5 | 0.0 | 408.0 | -- | - |
| 6 | 682.0 | 394.0 | 0.578 | 1 |
| 7 | 632.0 | 420.0 | 0.665 | 1 |
| 8 | 803.0 | 362.0 | 0.451 | 1 |
| Average | 474.5 | 324.1 | 0.614 | 0.4 |
| STD | 351.1 | 84.5 | 0.318 | 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balzano, A.; Čufar, K.; De Micco, V. Xylem and Phloem Formation Dynamics in Quercus ilex L. at a Dry Site in Southern Italy. Forests 2021, 12, 188. https://doi.org/10.3390/f12020188
Balzano A, Čufar K, De Micco V. Xylem and Phloem Formation Dynamics in Quercus ilex L. at a Dry Site in Southern Italy. Forests. 2021; 12(2):188. https://doi.org/10.3390/f12020188
Chicago/Turabian StyleBalzano, Angela, Katarina Čufar, and Veronica De Micco. 2021. "Xylem and Phloem Formation Dynamics in Quercus ilex L. at a Dry Site in Southern Italy" Forests 12, no. 2: 188. https://doi.org/10.3390/f12020188
APA StyleBalzano, A., Čufar, K., & De Micco, V. (2021). Xylem and Phloem Formation Dynamics in Quercus ilex L. at a Dry Site in Southern Italy. Forests, 12(2), 188. https://doi.org/10.3390/f12020188