
Tree Species Composition in Mixed Plantations Influences Plant Growth, Intrinsic Water Use Efficiency and Soil Carbon Stock

 , ,
, ,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
- Poplar and walnut (plot PJ).
- Poplar, walnut intercropped with hazel (plot PJC).
- Poplar, walnut intercropped with autumn-olive (plot PJE).
- Poplar, walnut intercropped with Italian alder (plot PJA).
2.2. Sampling and Dendrochronological Processing
2.3. Soil Sampling and Laboratory Analysis
2.4. Intrinsic Water Use Efficiency Determination
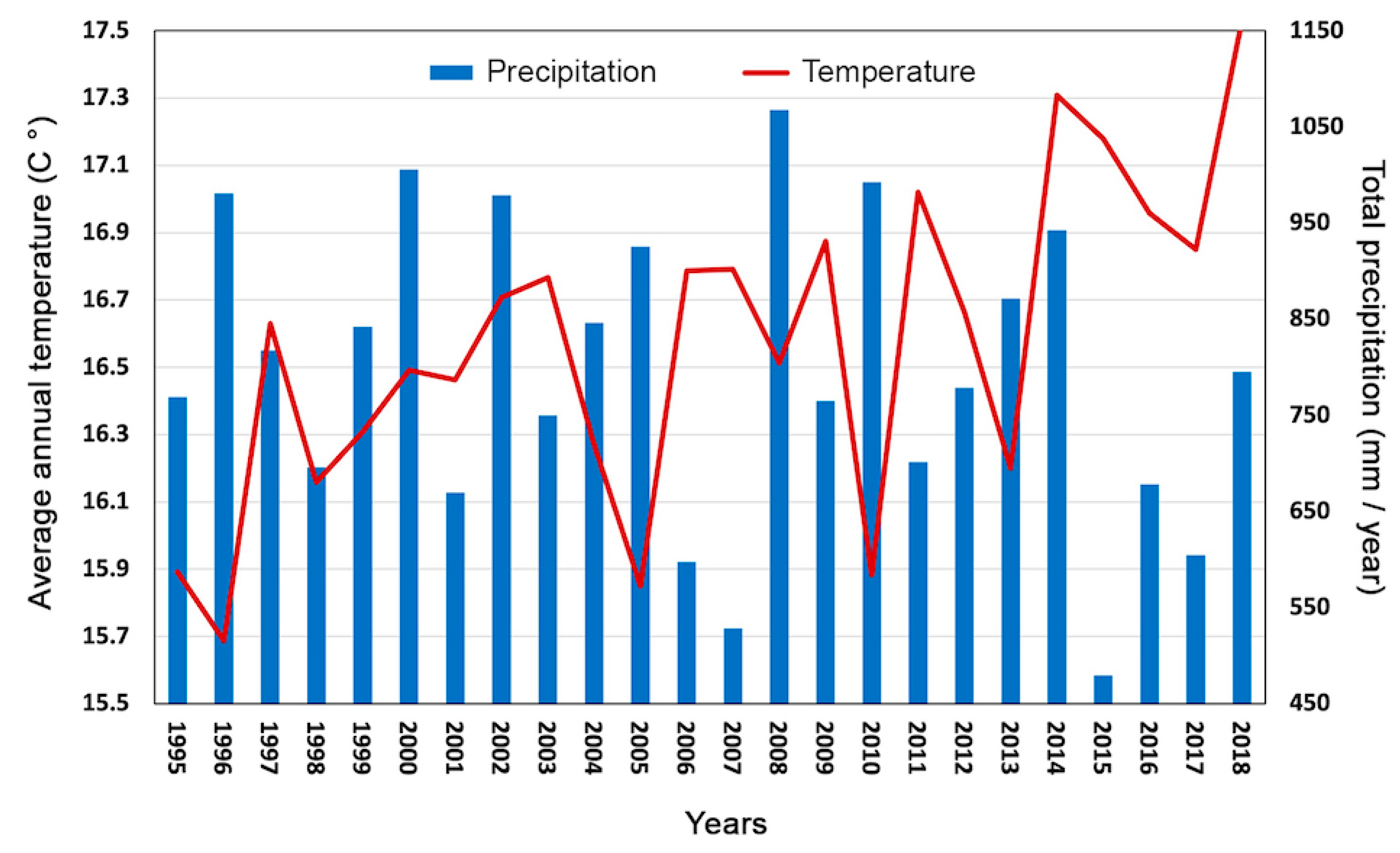
2.5. Climate Analysis
2.6. Data Analyses
3. Results
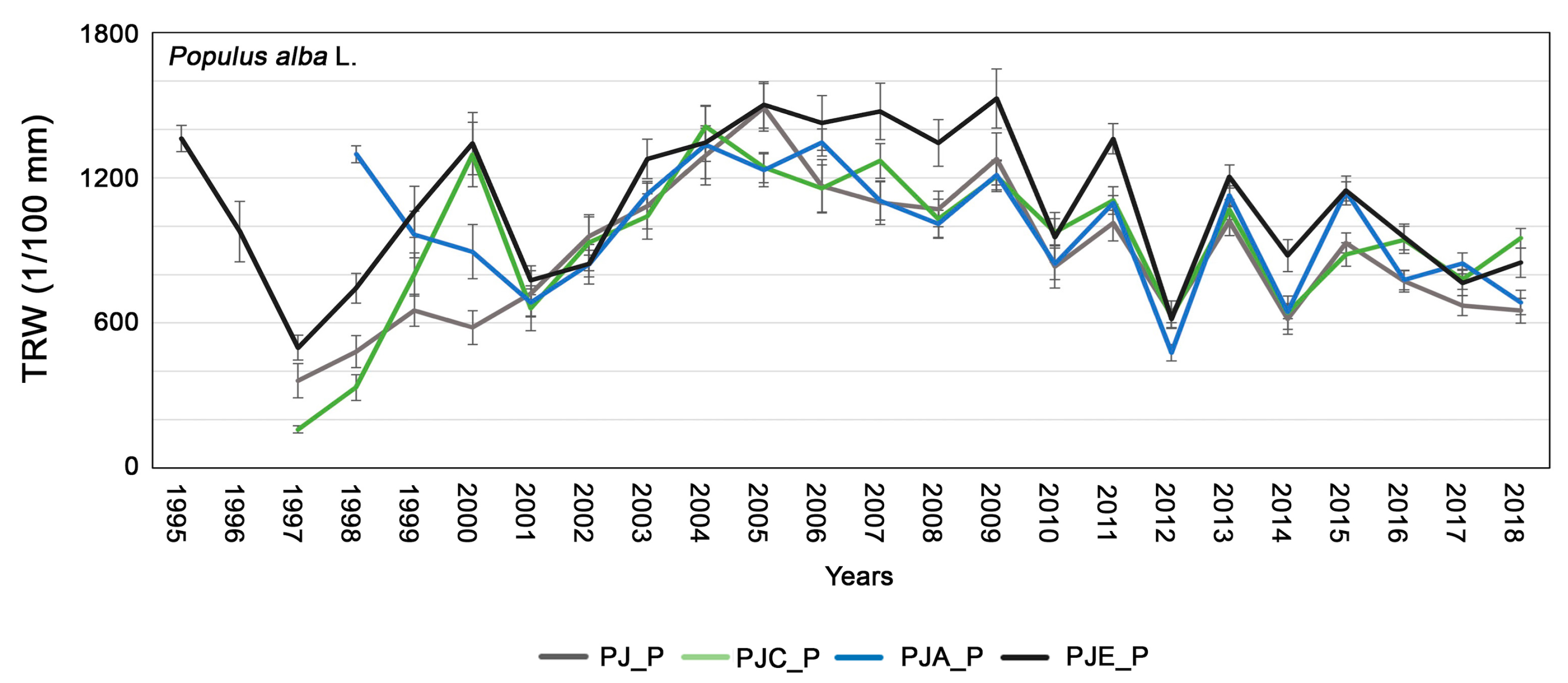
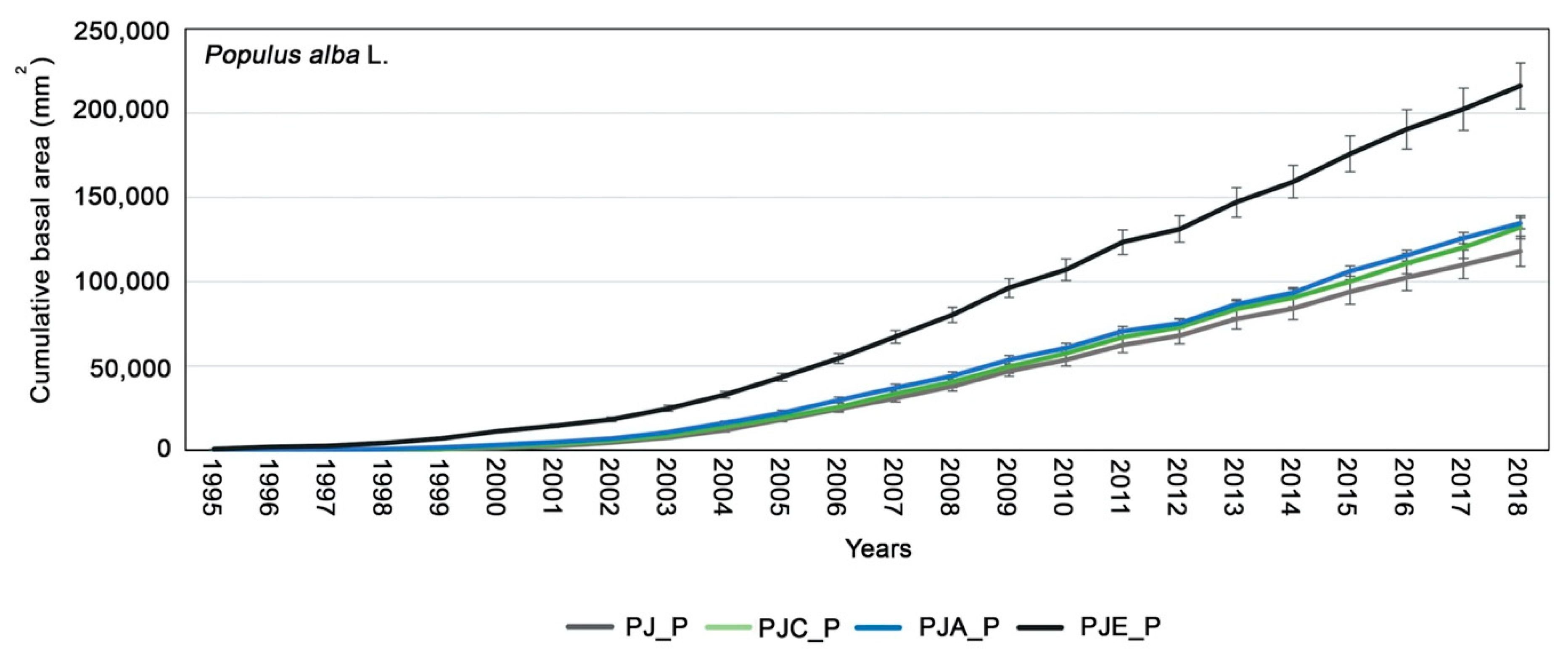
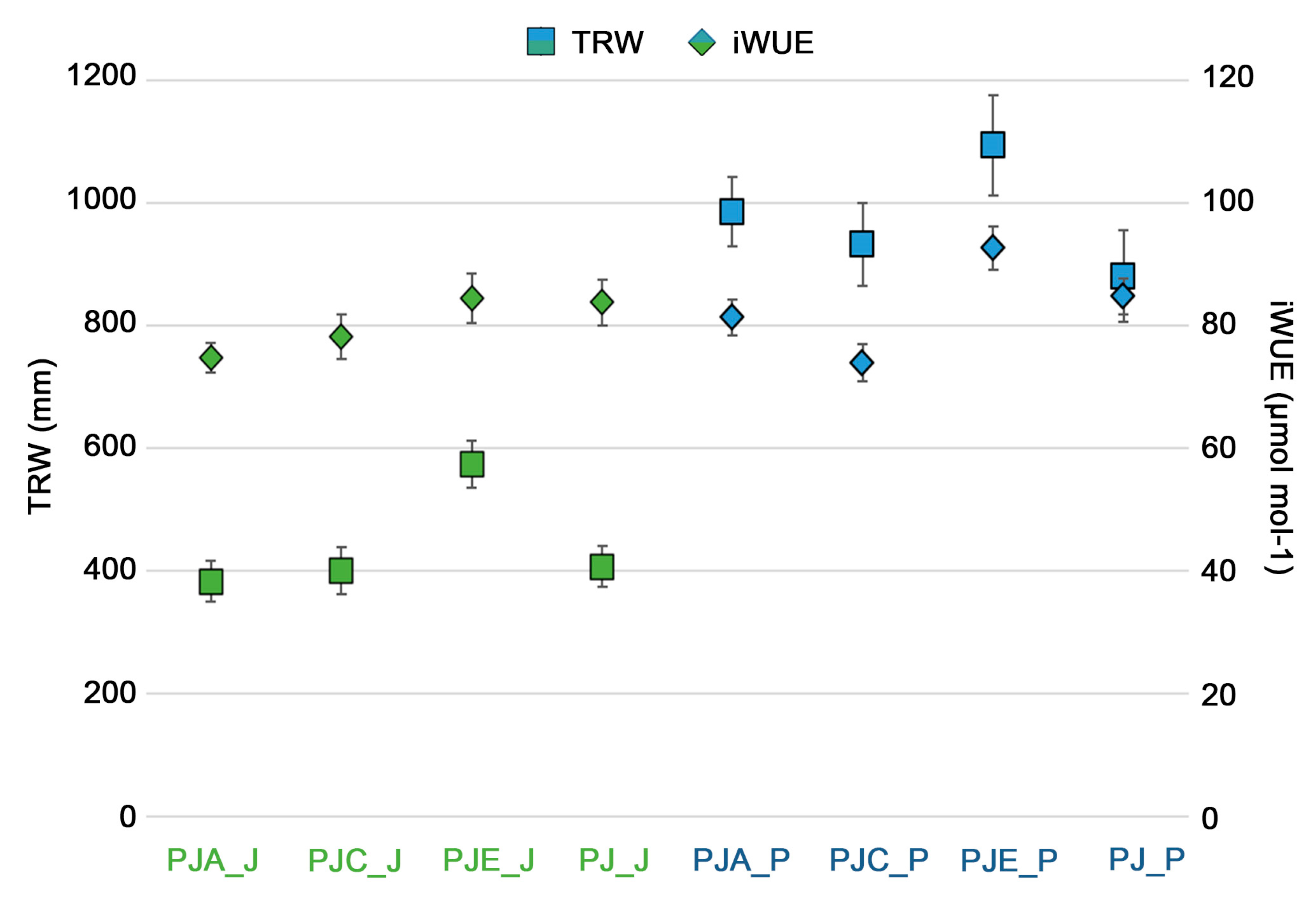
3.1. Dendrochronological Analysis of Populus Alba
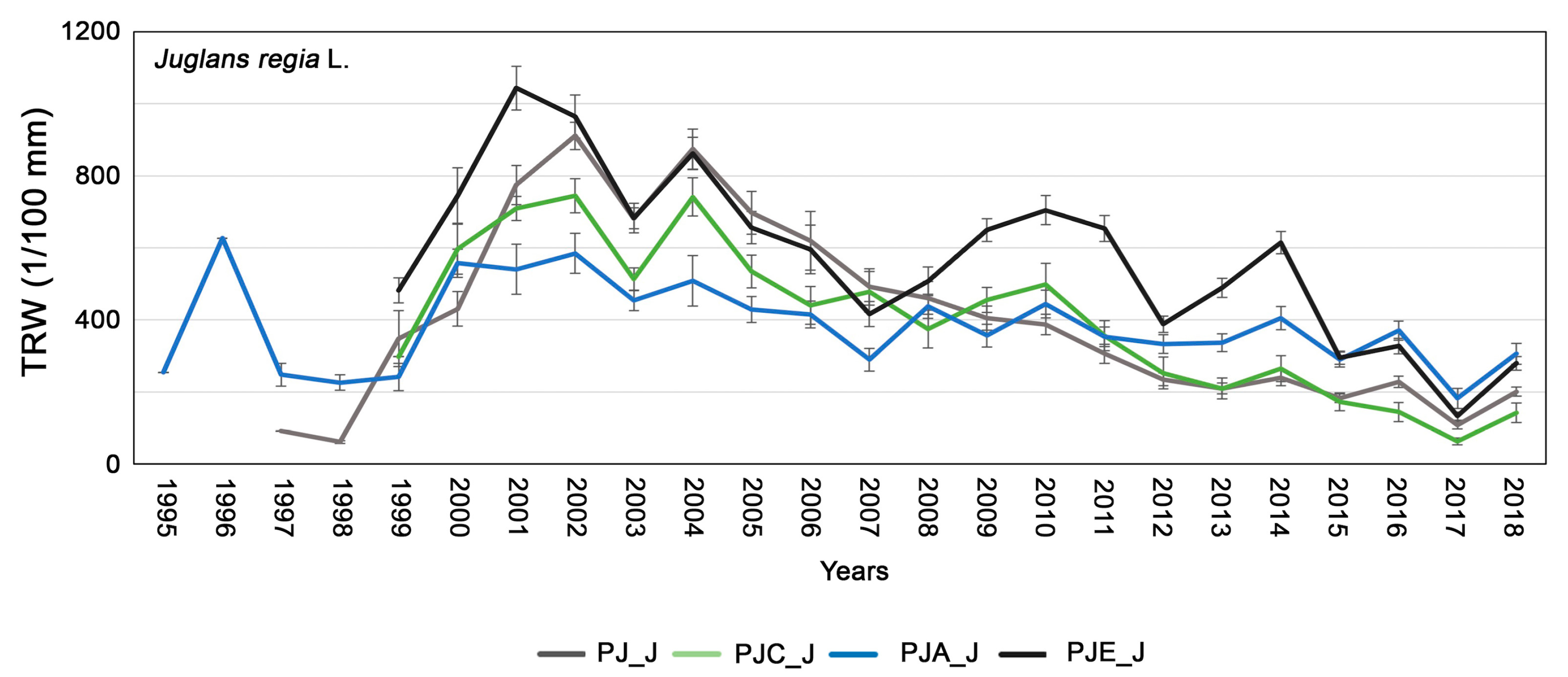
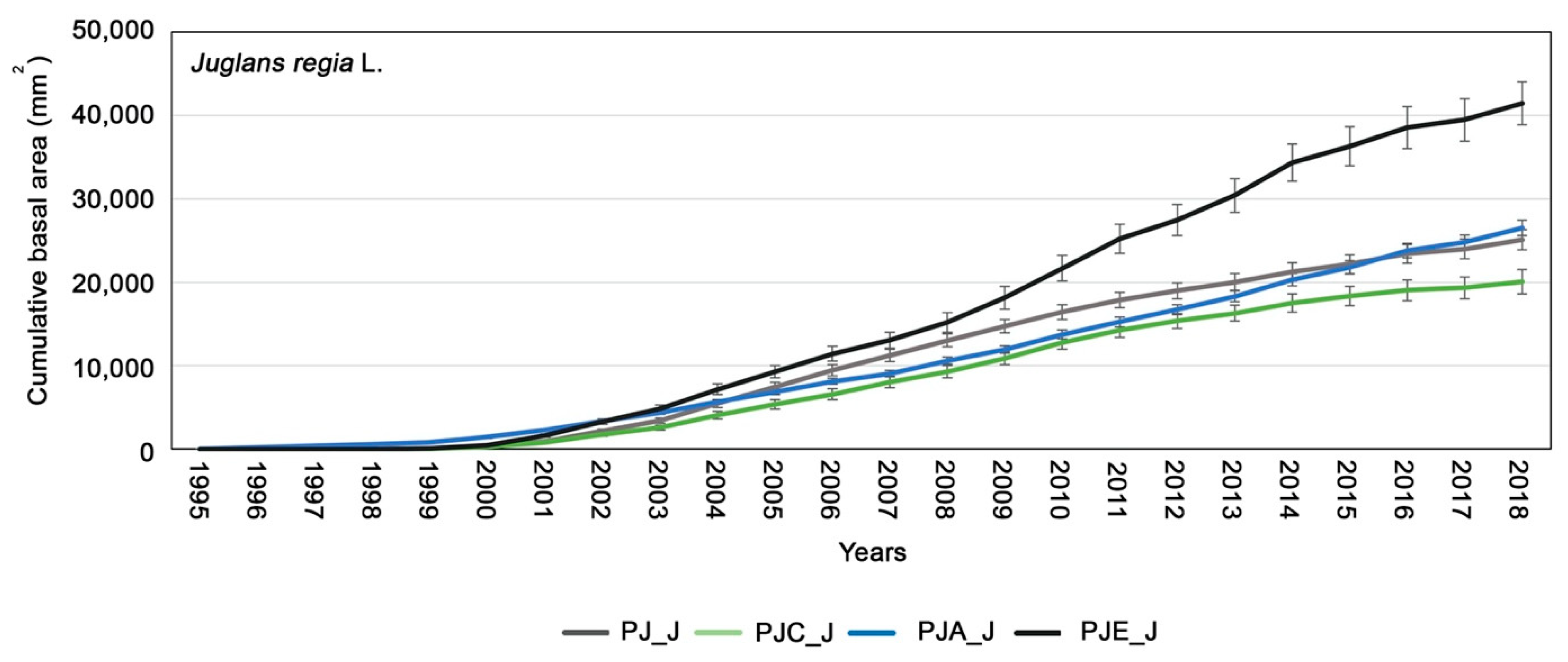
3.2. Dendrochronological Analysis of Juglans Regia
3.3. Relationship between Growth, iWUE and Climate
3.4. Soil Properties
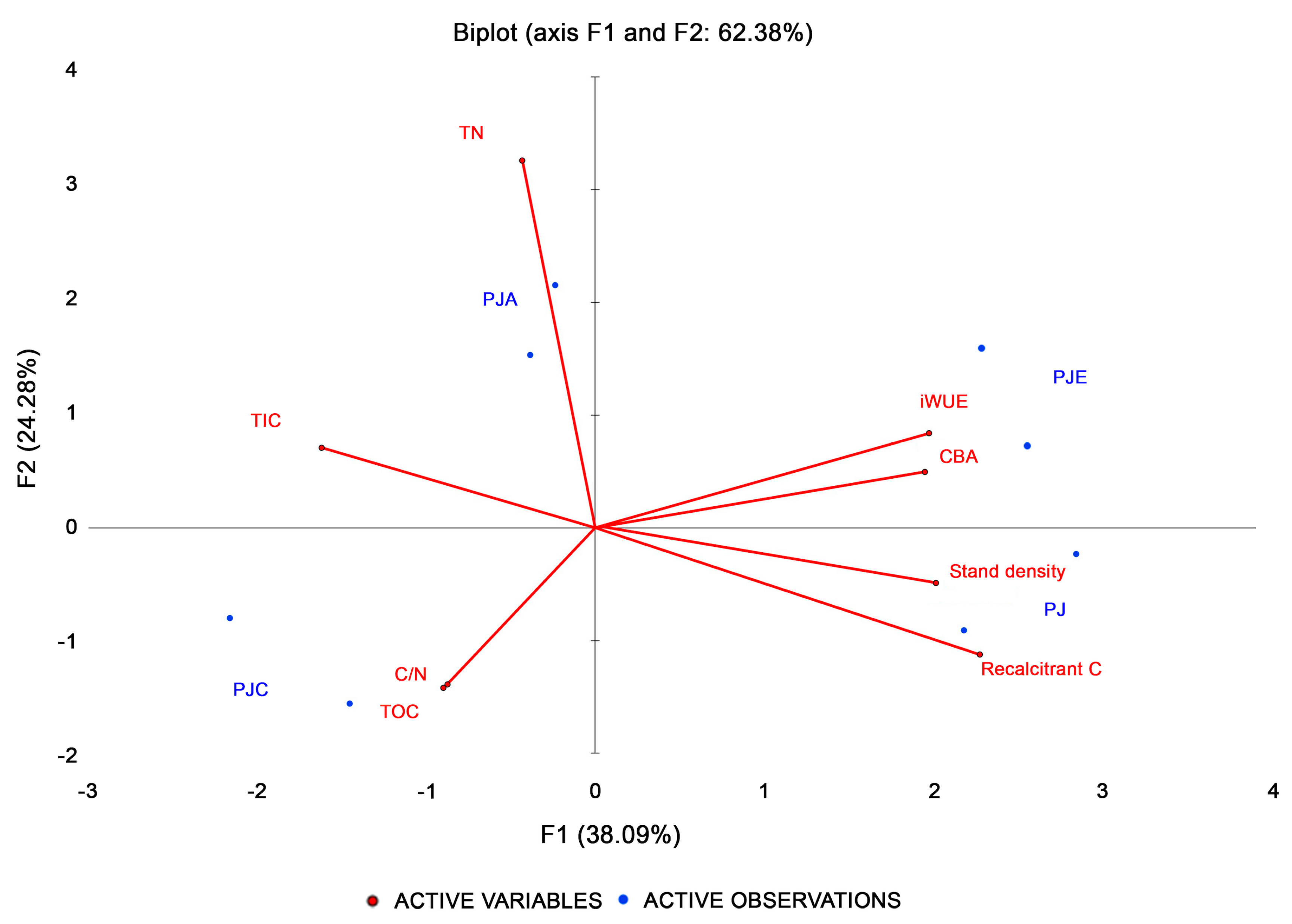
3.5. Multivariate Analysis
4. Discussion
4.1. Influence of the Intercropping on Productivity
4.2. Influence on the iWUE
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organsiation of the United Nations. Global Forest Resources Assessment 2020; FAO: Rome, Italy, 2020. [Google Scholar]
- Zhao, M.; Yang, J.; Zhao, N.; Liu, Y.; Wang, Y.; Wilson, J.P.; Yue, T. Estimation of China’s forest stand biomass carbon sequestration based on the continuous biomass expansion factor model and seven forest inventories from 1977 to 2013. For. Ecol. Manag. 2019, 448, 528–534. [Google Scholar] [CrossRef]
- Agnoletti, M. The degradation of traditional landscape in a mountain area of Tuscany during the 19th and 20th centuries: Implications for biodiversity and sustainable management. For. Ecol. Manag. 2007, 249, 5–17. [Google Scholar] [CrossRef]
- Ferretti, M.; Bacaro, G.; Brunialti, G.; Confalonieri, M.; Cristofolini, F.; Cristofori, A.; Frati, L.; Finco, A.; Gerosa, G.; Maccherini, S.; et al. Scarce evidence of ozone effect on recent health and productivity of alpine forests—A case study in Trentino, N. Italy. Environ. Sci. Pollut. Res. 2018, 25, 8217–8232. [Google Scholar] [CrossRef]
- Evans, J. Planted Forests: Uses, Impacts and Sustainability; FAO: Rome, Italy, 2009; ISBN 9781845935641. [Google Scholar]
- Food and Agriculture Organsiation of the United Nations. Global Forest Resources Assessment 2015; FAO: Rome, Italy, 2015; ISBN 9789251088210. [Google Scholar]
- IFD. Evaluation of the Community Aid Scheme for Forestry Measures in Agriculture of Regulation No 2080/92; Institute for Forestry Development: Joensuu, Finland, 2001. [Google Scholar]
- Kelty, M.J. The role of species mixtures in plantation forestry. For. Ecol. Manag. 2006, 233, 195–204. [Google Scholar] [CrossRef]
- Piotto, D. A meta-analysis comparing tree growth in monocultures and mixed plantations. For. Ecol. Manag. 2008, 255, 781–786. [Google Scholar] [CrossRef]
- Richards, A.E.; Forrester, D.I.; Bauhus, J.; Scherer-Lorenzen, M. The influence of mixed tree plantations on the nutrition of individual species: A review. Tree Physiol. 2010, 30, 1192–1208. [Google Scholar] [CrossRef] [Green Version]
- Alem, S.; Pavlis, J.; Urban, J.; Kucera, J. Pure and Mixed Plantations of Eucalyptus camaldulensis and Cupressus lusitanica: Their Growth Interactions and Effect on Diversity and Density of Undergrowth Woody Plants in Relation to Light. Open J. For. 2015, 5, 375–386. [Google Scholar] [CrossRef] [Green Version]
- Khanna, P.K. Comparison of growth and nutrition of young monocultures and mixed stands of Eucalyptus globulus and Acacia mearnsii. For. Ecol. Manag. 1997, 94, 105–113. [Google Scholar] [CrossRef]
- Liu, D.; Wang, Z.; Zhang, B.; Song, K.; Li, X.; Li, J.; Li, F.; Duan, H. Spatial distribution of soil organic carbon and analysis of related factors in croplands of the black soil region, Northeast China. Agric. Ecosyst. Environ. 2006, 113, 73–81. [Google Scholar] [CrossRef]
- Altieri, M.A. The Ecological Impacts of Large-Scale Agrofuel Monoculture Production Systems in the Americas. Bull. Sci. Technol. Soc. 2009, 29, 236–244. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L.; Vanclay, J.K. Mixed-species plantations of Eucalyptus with nitrogen-fixing trees: A review. For. Ecol. Manag. 2006, 233, 211–230. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.L.C.; Kuchma, O.; Krutovsky, K.V. Mixed-species versus monocultures in plantation forestry: Development, benefits, ecosystem services and perspectives for the future. Glob. Ecol. Conserv. 2018, 15, e00419. [Google Scholar] [CrossRef]
- Pelleri, F.; Pelleri, F.; Castro, G.; Marchi, M.; Fernandez-Moya, J.; Chiararbaglio, P.M.; Giorcelli, A.; Bergante, S.; Gennaro, M.; Manetti, M.C.; et al. The walnut plantations (Juglans spp.) in Italy and Spain: Main factors affecting growth. Ann. Silvic. Res. 2020, 44, 14–23. [Google Scholar] [CrossRef]
- Pelleri, F.; Ravagni, S.; Bianchetto, E.; Bidini, C. Comparing growth rate in a mixed plantation (walnut, poplar and nurse trees) with different planting designs: Results from an experimental plantation in Northern Italy. Ann. Silvic. Res. 2013, 37, 13–21. [Google Scholar] [CrossRef]
- Booth, T.H.; Williams, K.J. Developing biodiverse plantings suitable for changing climatic conditions 1: Underpinning scientific methods. Ecol. Manag. Restor. 2012, 13, 267–273. [Google Scholar] [CrossRef]
- Hulvey, K.B.; Hobbs, R.J.; Standish, R.J.; Lindenmayer, D.B.; Lach, L.; Perring, M.P. Benefits of tree mixes in carbon plantings. Nat. Clim. Chang. 2013, 3, 869–874. [Google Scholar] [CrossRef]
- Bauhus, J.; Forrester, D.I.; Pretzsch, H. From Observations to Evidence About Effects of Mixed-Species Stands BT—Mixed-Species Forests: Ecology and Management; Pretzsch, H., Forrester, D.I., Bauhus, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 27–71. ISBN 978-3-662-54553-9. [Google Scholar]
- Vandermeer, J. The Ecology of Intercropping; Cambridge University Press: Cambridge, UK, 1992; ISBN 0521346894. [Google Scholar]
- Butterfield, R.P. Tropical Timber Species in the Atlantic Low-Lands of Costa Rica and Wood Variation of Two Native Species; North Carolina State University: Raleigh, NC, USA, 1993. [Google Scholar]
- Crews, T.E.; Peoples, M.B. Legume versus fertilizer sources of nitrogen: Ecological tradeoffs and human needs. Agric. Ecosyst. Environ. 2004, 102, 279–297. [Google Scholar] [CrossRef]
- Vidal, D.F.; Trichet, P.; Puzos, L.; Bakker, M.R.; Delerue, F.; Augusto, L. Intercropping N-fixing shrubs in pine plantation forestry as an ecologically sustainable management option. For. Ecol. Manag. 2019, 437, 175–187. [Google Scholar] [CrossRef]
- Danise, T.; Andriuzzi, W.S.; Battipaglia, G.; Certini, G.; Guggenberger, G.; Innangi, M.; Mastrolonardo, G.; Niccoli, F.; Pelleri, F.; Fioretto, A. Mixed-species plantation effects on soil biological and chemical quality and tree growth of a former agricultural land. Forests 2021, 12, 842. [Google Scholar] [CrossRef]
- Torbert, J.L.; Burger, J.A. Forest land reclamation. In Reclamation of Drastically Disturbed Lands; Barnhisel, R.I., Darmody, R.G., Daniels, W.L., Eds.; Agronomy 41 American Society of Agronomy: Madison, WI, USA, 2000; pp. 371–398. [Google Scholar]
- Chiti, T.; Certini, G.; Puglisi, A.; Sanesi, G.; Capperucci, A.; Forte, C. Effects of associating a N-fixer species to monotypic oak plantations on the quantity and quality of organic matter in minesoils. Geoderma 2007, 138, 162–169. [Google Scholar] [CrossRef]
- Voigtlaender, M.; Laclau, J.P.; de Gonçalves, J.L.M.; de Piccolo, M.C.; Moreira, M.Z.; Nouvellon, Y.; Ranger, J.; Bouillet, J.P. Introducing Acacia mangium trees in Eucalyptus grandis plantations: Consequences for soil organic matter stocks and nitrogen mineralization. Plant Soil 2012, 352, 99–111. [Google Scholar] [CrossRef]
- Hoogmoed, M.; Cunningham, S.C.; Baker, P.J.; Beringer, J.; Cavagnaro, T.R. Is there more soil carbon under nitrogen-fixing trees than under non-nitrogen-fixing trees in mixed-species restoration plantings? Agric. Ecosyst. Environ. 2014, 188, 80–84. [Google Scholar] [CrossRef]
- Johnson, D.W.; Curtis, P.S. Effects of forest management on soil C and N storage: Meta analysis. For. Ecol. Manag. 2001, 140, 227–238. [Google Scholar] [CrossRef]
- Marron, N.; Epron, D. Are mixed-tree plantations including a nitrogen-fixing species more productive than monocultures? For. Ecol. Manag. 2019, 441, 242–252. [Google Scholar] [CrossRef]
- Bertness, M.D.; Callaway, R. Positive interactions in communities. Trends Ecol. Evol. 1994, 9, 191–193. [Google Scholar] [CrossRef]
- Forrester, D.I. The spatial and temporal dynamics of species interactions in mixed-species forests: From pattern to process. For. Ecol. Manag. 2014, 312, 282–292. [Google Scholar] [CrossRef]
- Farquhar, G.D.; O’Leary, M.H.; Berry, J.A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Aust. J. Plant. Physiol. 1982, 9, 121. [Google Scholar] [CrossRef]
- Vanclay, J.K. Managing water use from forest plantations. For. Ecol. Manag. 2009, 257, 385–389. [Google Scholar] [CrossRef] [Green Version]
- Siegwolf, R.T.W.; Matyssek, R.; Saurer, M.; Maurer, S.; Günthardt-Goerg, M.S.; Schmutz, P.; Bucher, J.B. Stable isotope analysis reveals differential effects of soil nitrogen and nitrogen dioxide on the water use efficiency in hybrid poplar leaves. New Phytol. 2001, 149, 233–246. [Google Scholar] [CrossRef]
- Ripullone, F.; Lauteri, M.; Grassi, G.; Amato, M.; Borghetti, M. Variation in nitrogen supply changes water-use efficiency of Pseudotsuga menziesii and Populus x euroamericana; a comparison of three approaches to determine water-use efficiency. Tree Physiol. 2004, 24, 671–679. [Google Scholar] [CrossRef]
- Guerrieri, R.; Siegwolf, R.; Saurer, M.; Ripullone, F.; Mencuccini, M.; Borghetti, M. Anthropogenic NOx emissions alter the intrinsic water-use efficiency (WUEi) for Quercus cerris stands under Mediterranean climate conditions. Environ. Pollut. 2010, 158, 2841–2847. [Google Scholar] [CrossRef]
- Battipaglia, G.; Pelleri, F.; Lombardi, F.; Altieri, S.; Vitone, A.; Conte, E.; Tognetti, R. Effects of associating Quercus robur L. and Alnus cordata Loisel. on plantation productivity and water use efficiency. For. Ecol. Manag. 2017, 391, 106–114. [Google Scholar] [CrossRef]
- Nichols, E.; Larsen, T.; Spector, S.; Davis, A.L.; Escobar, F.; Favila, M.; Vulinec, K. Global dung beetle response to tropical forest modification and fragmentation: A quantitative literature review and meta-analysis. Biol. Conserv. 2007, 137, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Manson, D.G.; Schmidt, S.; Bristow, M.; Erskine, P.D.; Vanclay, J.K. Species-site matching in mixed species plantations of native trees in tropical Australia. Agrofor. Syst. 2013, 87, 233–250. [Google Scholar] [CrossRef]
- Nguyen, H.; Firn, J.; Lamb, D.; Herbohn, J. Wood density: A tool to find complementary species for the design of mixed species plantations. For. Ecol. Manag. 2014, 334, 106–113. [Google Scholar] [CrossRef]
- Eckstein, D.; Bauch, J. Beitrag zur Rationalisierung eines dendrochronologischen Verfahrens und zur Analyse seiner Aussagesicherheit. Forstwiss. Cent. 1969, 88, 230–250. [Google Scholar] [CrossRef]
- Martín-Benito, D.; Del Río, M.; Heinrich, I.; Helle, G.; Cañellas, I. Response of climate-growth relationships and water use efficiency to thinning in a Pinus nigra afforestation. For. Ecol. Manag. 2010, 259, 967–975. [Google Scholar] [CrossRef]
- Biondi, F. Comparing tree-ring chronologies and repeated timber inventories as forest monitoring tools. Ecol. Appl. 1999, 9, 216. [Google Scholar] [CrossRef]
- Battipaglia, G.; Zalloni, E.; Castaldi, S.; Marzaioli, F.; Cazzolla-Gatti, R.; Lasserre, B.; Tognetti, R.; Marchetti, M.; Valentini, R. Long tree-ring chronologies provide evidence of recent tree growth decrease in a central african tropical forest. PLoS ONE 2015, 10, e0120962. [Google Scholar] [CrossRef] [PubMed]
- Niccoli, F.; Pelleri, F.; Manetti, M.C.; Sansone, D.; Battipaglia, G. Effects of thinning intensity on productivity and water use efficiency of Quercus robur L. For. Ecol. Manag. 2020, 473, 118282. [Google Scholar] [CrossRef]
- Gee, G.W.; Bauder, J.W. Particle-size analysis. In Methods of Soil Analysis; Klute, A., Ed.; Soil Science Society of America: Madison, WI, USA, 1986; pp. 383–411. [Google Scholar]
- Santi, C.; Certini, G.; D’Acqui, L.P. Direct determination of organic carbon by dry combustion in soils with carbonates. Commun. Soil Sci. Plant Anal. 2006, 37, 155–162. [Google Scholar] [CrossRef]
- Zimmermann, M.; Leifeld, J.; Abiven, S.; Schmidt, M.W.I.; Fuhrer, J. Sodium hypochlorite separates an older soil organic matter fraction than acid hydrolysis. Geoderma 2007, 139, 171–179. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon Isotope Discrimination and Photosynthesis. Annu. Rev. Plant. Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- McCarroll, D.; Loader, N.J. Stable isotopes in tree rings. Quat. Sci. Rev. 2004, 23, 771–801. [Google Scholar] [CrossRef]
- Harris, I.; Jones, P.D.; Osborn, T.J.; Lister, D.H. Updated high-resolution grids of monthly climatic observations—The CRU TS3.10 Dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef] [Green Version]
- Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology: Applications in the Environmental Sciences; Springer: Dordrecht, The Netherlands, 1990; ISBN 0-7923-0586-8. [Google Scholar]
- Podani, J. Introduction to the Exploration of Multivariate Biological Data; Backhuys Publishers: Leiden, The Netherlands, 2000; ISBN 90-5782-067-6. [Google Scholar]
- Podani, J. Multivariate Data Analysis in Ecology and Systematics: A Methodological Guide to the SYN-TAX 5.0 Package; SPB Academic Publishing: Amsterdam, The Netherlands, 1994; ISBN 9051030940. [Google Scholar]
- McLauchlan, K. The Nature and Longevity of Agricultural Impacts on Soil Carbon and Nutrients: A Review. Ecosystems 2006, 9, 1364–1382. [Google Scholar] [CrossRef]
- Matus, F.J. Fine silt and clay content is the main factor defining maximal C and N accumulations in soils: A meta-analysis. Sci. Rep. 2021, 11, 6438. [Google Scholar] [CrossRef]
- Balzano, A.; Čufar, K.; Battipaglia, G.; Merela, M.; Prislan, P.; Aronne, G.; De Micco, V. Xylogenesis reveals the genesis and ecological signal of IADFs in Pinus pinea L. and Arbutus unedo L. Ann. Bot. 2018, 121, 1231–1242. [Google Scholar] [CrossRef]
- Balzano, A.; Battipaglia, G.; Cherubini, P.; De Micco, V. Xylem plasticity in pinus pinaster and quercus ilex growing at sites with different water availability in the mediterranean region: Relations between intra-annual density fluctuations and environmental conditions. Forests 2020, 11, 379. [Google Scholar] [CrossRef] [Green Version]
- Loreau, M.; Hector, A. Partitioning selection and complementarity in biodiversity experiments. Nature 2001, 412, 72–76. [Google Scholar] [CrossRef]
- Tobner, C.M.; Paquette, A.; Reich, P.B.; Gravel, D.; Messier, C. Advancing biodiversity-ecosystem functioning science using high-density tree-based experiments over functional diversity gradients. Oecologia 2014, 174, 609–621. [Google Scholar] [CrossRef]
- Kelty, M.J.; Cameron, I. Plot designs for the analysis of species interactions in mixed stands. Commonw. For. Rev. 1995, 74, 322–332. [Google Scholar]
- Pretzsch, H. Canopy space filling and tree crown morphology in mixed-species stands compared with monocultures. For. Ecol. Manag. 2014, 327, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Fang, C.; Smith, P.; Moncrieff, J.B.; Smith, J.U. Similar response of labile and resistant soil organic matter pools to changes in temperature. Nature 2005, 433, 57–59. [Google Scholar] [CrossRef]
- von Lützow, M.; Kögel-Knabner, I.; Ekschmitt, K.; Flessa, H.; Guggenberger, G.; Matzner, E.; Marschner, B. SOM fractionation methods: Relevance to functional pools and to stabilization mechanisms. Soil Biol. Biochem. 2007, 39, 2183–2207. [Google Scholar] [CrossRef]
- Becquey, J.; Vidal, C. Quels accompagnements ligneux choisir pour les plantations de noyer? Forêt Entrep. 1997, 170, 35–38. [Google Scholar]
- Bianchetto, E.; Vitone, A.; Bidini, C.; Pelleri, F. Effetto di differenti tipologie di consociazione sull’accrescimento e sulla qualità del noce comune (Juglans regia L.) in un impianto di arboricoltura da legno nell’Italia centrale. Ann. Silvic. Res. 2013, 37, 38. [Google Scholar] [CrossRef]
- Mohr, D.; Topp, W. Hazel improves soil quality of sloping oak stands in a German low mountain range. Ann. For. Sci. 2005, 62, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Frouz, J.; Keplin, B.; Pižl, V.; Tajovský, K.; Starý, J.; Lukešová, A.; Nováková, A.; Balík, V.; Háněl, L.; Materna, J.; et al. Soil biota and upper soil layer development in two contrasting post-mining chronosequences. Ecol. Eng. 2001, 17, 275–284. [Google Scholar] [CrossRef]
- Becciolini, R.; Pelleri, F. Consociazione tra farnia e ontano napoletano: Valutazione degli effetti in un impianto prima e dopo il diradamento. Sherwood For. Alberi Oggi 2006, 119, 11–16. [Google Scholar]
- Tani, A.; Maltoni, A.; Mariotti, B.; Buresti Lattes, E. Juglans regia L. tree plantations for wood production in mining area of S. Barbara (AR). Evaluation of N-fixing accessory trees effect. For. Riv. Selvic. Ecol. For. 2006, 3, 588–597. [Google Scholar] [CrossRef] [Green Version]
- Corazzesi, A.; Tani, A.; Pelleri, F. Effetto della consociazione e del diradamento in un impianto di arboricoltura da legno con latifoglie di pregio dopo oltre. Ann. Silvic. Res. 2010, 36, 37–48. [Google Scholar] [CrossRef]
- Buresti, E.; Domenach, A.M.; Bosco, M.; Moiroud, A. Comparison Between Quercus Robur/Alnus Cordata Mixed Plantation and Quercus Robur in Monoculture BT—Nitrogen Fixation. In Proceedings of the Fifth International Symposium on Nitrogen Fixation with Non-Legumes, Florence, Italy, 10–14 September 1990; Polsinelli, M., Materassi, R., Vincenzini, M., Polsinelli, M., Materassi, R., Vincenzini, M., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 1991; pp. 651–652, ISBN 978-94-011-3486-6. [Google Scholar]
- Cutini, A.; Giannini, T. Effetti della consociazione con Alnus cordata sulla funzionalità di impianti di noce comune (Juglans regia L.) sottoposti a diradamento. J. Silvic. For. Ecol. 2009, 6, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Schlesinger, R.C.; Williams, R.D. Growth response of black walnut to interplanted trees. For. Ecol. Manag. 1984, 9, 235–243. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.R.; Hemery, G.E.; Savill, P.S. Early growth and form of common walnut (Juglans regia L.) in mixture with tree and shrub nurse species in southern England. Forestry 2008, 81, 631–644. [Google Scholar] [CrossRef]
- Mohni, C.; Pelleri, F.; Hemery, G.E. The modern silviculture of Juglans regia L.: A literature review. Bodenkultur 2009, 60, 21–34. [Google Scholar]
- Ponder, F. Weed control and autumn-olive affect early growth and survival of black walnut in a hardwood clearcut. New For. 1988, 2, 195–201. [Google Scholar] [CrossRef]
- Hemery, G.E. Growing walnut in mixed stands. Q. J. For. 2001, 95, 31–36. [Google Scholar]
- Loewenstein, N.J.; Pallardy, S.G. Drought tolerance, xylem sap abscisic acid and stomatal conductance during soil drying: A comparison of canopy trees of three temperate deciduous angiosperms. Tree Physiol. 1998, 18, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Monclus, R.; Dreyer, E.; Villar, M.; Delmotte, F.M.; Delay, D.; Petit, J.-M.; Barbaroux, C.; Le Thiec, D.; Brechet, C.; Brignolas, F. Impact of drought on productivity and water use efficiency in 29 genotypes of Populus deltoides x Populus nigra. New Phytol. 2006, 169, 765–777. [Google Scholar] [CrossRef]
- Palandrani, C.; Battipaglia, G.; Alberti, G. Influence of tree species richness on tree growth and intrinsic water-use efficiency after drought in tree plantations in north-eastern Italy. Eur. J. For. Res. 2020, 139, 869–877. [Google Scholar] [CrossRef]
- Roden, J.; Van Volkenburgh, E.; Hinckley, T.M. Cellular basis for limitation of poplar leaf growth by water deficit. Tree Physiol. 1990, 6, 211–219. [Google Scholar] [CrossRef]
- DesRochers, A.; Van Den Driessche, R.; Thomas, B.R. The interaction between nitrogen source, soil pH, and drought in the growth and physiology of three poplar clones. Can. J. Bot. 2007, 85, 1046–1057. [Google Scholar] [CrossRef]
- Edwards, C.E.; Ewers, B.E.; McClung, C.R.; Lou, P.; Weinig, C. Quantitative variation in water-use efficiency across water regimes and its relationship with circadian, vegetative, reproductive, and leaf gas-exchange traits. Mol. Plant. 2012, 5, 653–668. [Google Scholar] [CrossRef] [Green Version]
- Lampinen, B.; Buchner, R.P.; Fulton, A.; Grant, J.; Mills, N.; Prichard, T.; Schwankl, L.; Shackel, K.; Gilles, C.; Little, C.; et al. Irrigation Management in Walnut Using Evapotranspiration, Soil and Plant. Based Data; California Walnut Board and Commission: Folsom, CA, USA, 2003. [Google Scholar]
- Buchner, R.P.; Lindow, S.E.; Adaskaveg, J.E.; Pickel, C.; Gilles, C.K.; Koutsoukis, R.; Smith, S.G. Walnut Blight Control. Investigations Tehama 2008; California Walnut Board and Commission: Folsom, CA, USA, 2008. [Google Scholar]
- Caldwell, M.M.; Richards, J.H. Hydraulic lift: Water efflux from upper roots improves effectiveness of water uptake by deep roots. Oecologia 1989, 79, 1–5. [Google Scholar] [CrossRef]
- Battipaglia, G.; Saurer, M.; Cherubini, P.; Calfapietra, C.; Mccarthy, H.R.; Norby, R.J.; Francesca Cotrufo, M. Elevated CO2 increases tree-level intrinsic water use efficiency: Insights from carbon and oxygen isotope analyses in tree rings across three forest FACE sites. New Phytol. 2013, 197, 544–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, A.P.; De Kauwe, M.G.; Bastos, A.; Belmecheri, S.; Georgiou, K.; Keeling, R.F.; McMahon, S.M.; Medlyn, B.E.; Moore, D.J.P.; Norby, R.J.; et al. Integrating the evidence for a terrestrial carbon sink caused by increasing atmospheric CO2. New Phytol. 2021, 229, 2413–2445. [Google Scholar] [CrossRef]
- Lauteri, M.; Alessio, G.A.; Paris, P. Using oxygen stable isotopes to investigate the soil-plant-atmosphere hydraulic continuum in complex stands of walnut. Acta Hortic. 2006, 705, 223–230. [Google Scholar] [CrossRef]
- Boudru, M. Forêt et Sylviculture: Sylviculture Appliquée; Presses Agronomiques De Gembloux: Gembloux, Belgium, 1986; ISBN 9782870160305. [Google Scholar]
- Becquey, J. Les Noyers à Bois; Forestier, Institut Pour Le Developpement: Paris, France, 1997; ISBN 9782904740558. [Google Scholar]
- Bernetti, G. Selvicoltura Speciale; UTET: Torino, Italy, 1995.
- Stanescu, V.; Sofletea, N.; Popescu, O. Flora Forestiera Lemnoasa a României; Ceres: Bucuresti, Romania, 1997; ISBN 9789734003839. [Google Scholar]
- Salbitano, F.; Buresti Lattes, E.; Tani, A. Analisi Comparata Dell’architettura di Apparati Ipogei ed Epigei di Piante di Juglans spp.; Prefettura di Firenze: Firenze, Italy, 2001.
- Ehleringer, J.R.; Hall, A.E.; Farquhar, G.D. Stable Isotopes and Plant. Carbon-Water Relations; Academic Press: San Diego, CA, USA, 1993. [Google Scholar]
- Aussenac, G. Interactions between forest stands and microclimate: Ecophysiological aspects and consequences for silviculture. Ann. For. Sci. 2000, 57, 287–301. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CBA PJ_J | CBA PJC_J | CBA PJA_J | CBA PJE_J | CBA PJ_P | CBA PJC_P | CBA PJA_P | CBA PJE_P | |
|---|---|---|---|---|---|---|---|---|
| Tmin MAM | ns | ns | ns | ns | ns | ns | ns | ns |
| Tmin JJA | 0.46 * | 0.42 * | 0.44 * | 0.40 * | 0.51 * | 0.43 * | 0.44 * | 0.45 * |
| Tmin SON | ns | ns | ns | ns | ns | ns | ns | ns |
| Tmax MAM | ns | ns | ns | ns | ns | ns | ns | ns |
| Tmax JJA | 0.45 * | 0.50 * | 0.57 ** | 0.46 * | 0.48 * | 0.51 * | 0.55 * | 0.48 * |
| Tmax SON | 0.44 * | 0.55 * | 0.45 * | 0.50 * | 0.43 * | 0.45 * | 0.48 * | 0.50 * |
| Tmedium MAM | ns | ns | ns | ns | ns | ns | ns | ns |
| Tmedium JJA | 0.43 * | 0.45 * | 0.51 * | 0.47 * | 0.46 * | 0.43 * | 0.52 * | 0.52 * |
| Tmedium SON | 0.50 * | 0.43 * | 0.42 * | 0.45 * | 0.42 * | 0.40 * | 0.43 * | 0.44 * |
| P MAM | ns | ns | ns | ns | ns | ns | ns | ns |
| P JJA | ns | ns | ns | ns | ns | ns | ns | ns |
| P SON | ns | ns | ns | ns | ns | ns | ns | ns |
| P TOT | ns | ns | ns | ns | ns | ns | ns | ns |
| iWUE PJ_J | iWUE PJC_J | iWUE PJA_J | iWUE PJE_J | iWUE PJ_P | iWUE PJC_P | iWUE PJA_P | iWUE PJE_P | |
|---|---|---|---|---|---|---|---|---|
| Tmin MAM | ns | ns | ns | ns | ns | ns | ns | ns |
| Tmin JJA | ns | ns | ns | ns | ns | ns | ns | ns |
| Tmin SON | ns | ns | ns | ns | −0.80 * | ns | ns | ns |
| Tmax MAM | ns | ns | ns | 0.86 * | ns | ns | ns | ns |
| Tmax JJA | ns | ns | ns | ns | ns | ns | ns | ns |
| Tmax SON | ns | ns | ns | ns | ns | ns | ns | ns |
| Tmedium MAM | ns | ns | ns | 0.89 * | ns | ns | ns | ns |
| Tmedium JJA | ns | ns | ns | ns | ns | ns | ns | ns |
| Tmedium SON | ns | ns | ns | ns | −0.82 * | ns | ns | ns |
| P MAM | ns | ns | ns | ns | ns | ns | ns | ns |
| P JJA | ns | ns | ns | 0.84 * | ns | ns | ns | 0.95 * |
| P SON | ns | ns | ns | ns | ns | ns | ns | ns |
| P TOT | ns | ns | ns | ns | ns | ns | ns | ns |
| Sand | Silt | Clay | BD | pHCaCl2 | TOC | TIC | TN | C/N | Stable C | |
|---|---|---|---|---|---|---|---|---|---|---|
| ‰ | ‰ | ‰ | g cm3 | g kg−1 | g kg−1 | g kg−1 | % of TOC | |||
| PJC | 59,555 a | 256 ± 79 a | 149 ± 26 a | 1.4 ± 0.1 a | 7.0 ± 0.2 a | 18.19 ± 0.68 a | 18.3 ± 0.5 a | 1.18 ± 0.43 a | 15.7 ± 2.6 a | 2.5 ± 0.7 a |
| PJE | 537 ± 38 abc | 294 ± 14 ab | 169 ± 32 a | 1.5 ± 0.2 a | 7.1 ± 0.2 a | 13.16 ± 2.90 b | 15.9 ± 0.4 a | 1.12 ± 0.34 a | 12.2 ± 2.2 b | 2.9 ± 1.8 a |
| PJA | 375 ± 82 b | 424 ± 67 b | 200 ± 20 a | 1.4 ± 0.2 a | 7.0 ± 0.2 a | 15.19 ± 4.65 ab | 19.1 ± 0.7 a | 1.33 ± 0.39 a | 11.5 ± 1.8 b | 2.5 ± 1.1 a |
| PJ | 437 ± 53 c | 380 ± 43 b | 183 ± 32 a | 1.4 ± 0.1 a | 7.0 ± 0.1 a | 13.41 ± 4.00 b | 21.8 ± 0.6 a | 1.15 ± 0.45 a | 13.0 ± 4.9 b | 8.4 ± 2.4 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niccoli, F.; Danise, T.; Innangi, M.; Pelleri, F.; Manetti, M.C.; Mastrolonardo, G.; Certini, G.; Fioretto, A.; Battipaglia, G. Tree Species Composition in Mixed Plantations Influences Plant Growth, Intrinsic Water Use Efficiency and Soil Carbon Stock. Forests 2021, 12, 1251. https://doi.org/10.3390/f12091251
Niccoli F, Danise T, Innangi M, Pelleri F, Manetti MC, Mastrolonardo G, Certini G, Fioretto A, Battipaglia G. Tree Species Composition in Mixed Plantations Influences Plant Growth, Intrinsic Water Use Efficiency and Soil Carbon Stock. Forests. 2021; 12(9):1251. https://doi.org/10.3390/f12091251
Chicago/Turabian StyleNiccoli, Francesco, Tiziana Danise, Michele Innangi, Francesco Pelleri, Maria Chiara Manetti, Giovanni Mastrolonardo, Giacomo Certini, Antonietta Fioretto, and Giovanna Battipaglia. 2021. "Tree Species Composition in Mixed Plantations Influences Plant Growth, Intrinsic Water Use Efficiency and Soil Carbon Stock" Forests 12, no. 9: 1251. https://doi.org/10.3390/f12091251
APA StyleNiccoli, F., Danise, T., Innangi, M., Pelleri, F., Manetti, M. C., Mastrolonardo, G., Certini, G., Fioretto, A., & Battipaglia, G. (2021). Tree Species Composition in Mixed Plantations Influences Plant Growth, Intrinsic Water Use Efficiency and Soil Carbon Stock. Forests, 12(9), 1251. https://doi.org/10.3390/f12091251