
Hybridization Assays in Strawberry Tree toward the Identification of Plants Displaying Increased Drought Tolerance



Abstract
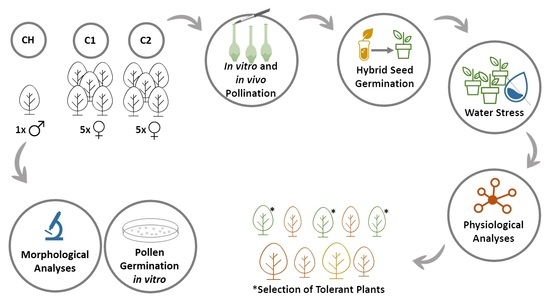
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Reproductive Phenology and Anatomy
2.3. Pollen Germination
2.4. In Vitro and In Situ Pollination Assays
2.5. Seed Germination and Plant Development
2.6. Drought Stress Assays
2.7. Statistical Analysis
3. Results
3.1. Reproductive Phenology and Anatomy
3.2. Pollen Germination
3.3. In Vitro and In Situ Pollination Assays
3.4. Seed Germination and Plant Development
3.5. Plant Water Status and Gas Exchange
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Torres, J.A.; Valle, F.; Pinto, C.; García-Fuentes, A.; Salazar, C.; Cano, E. Arbutus unedo L. communities in southern Iberian Peninsula mountains. Plant Ecol. 2002, 160, 207–223. [Google Scholar] [CrossRef]
- Morgado, S.; Morgado, M.; Plácido, A.I.; Roque, F.; Duarte, A.P. Arbutus unedo L.: From traditional medicine to potential uses in modern pharmacotherapy. J. Ethnopharmacol. 2018, 225, 90–102. [Google Scholar] [CrossRef]
- Alarcão-e-Silva, M.L.C.M.M.; Leitão, A.E.B.; Azinheira, H.G.; Leitão, M.C.A. The Arbutus Berry: Studies on its color and chemical characteristics at two mature stages. J. Food Compos. Anal. 2001, 14, 27–35. [Google Scholar] [CrossRef]
- Bussotti, F.; Ferrini, F.; Pollastrini, M.; Fini, A. The challenge of Mediterranean sclerophyllous vegetation under climate change: From acclimation to adaptation. Environ. Exp. Bot. 2014, 103, 80–98. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Peñuelas, J. Drought-induced oxidative stress in strawberry tree (Arbutus unedo L.) growing in Mediterranean field conditions. Plant Sci. 2004, 166, 1105–1110. [Google Scholar] [CrossRef]
- Martins, J.F.; Correia, S.I.; Canhoto, J.M. Somatic embryogenesis induction and plant regeneration in strawberry tree (Arbutus unedo L.). In Methods in Molecular Biology; Germana, M., Lambardi, M., Eds.; Humana Press: New York, NY, USA, 2016; Volume 1359, pp. 329–339. [Google Scholar]
- Martins, J.F.; Santos, T.; Correia, S.I.; Canhoto, J.M. Somatic embryogenesis in Arbutus unedo L. and other Ericaceae. In Vegetative Propagation of Forest Trees; Park, Y.S., Bonga, J., Moon, H.K., Eds.; National Institute of Forest Science: Seoul, Korea, 2016; pp. 565–590. [Google Scholar]
- Gomes, F.; Canhoto, J.M. Micropropagation of strawberry tree (Arbutus unedo L.) from adult plants. Vitr. Cell. Dev. Biol. Plant 2009, 45, 72–82. [Google Scholar] [CrossRef]
- Martins, J.F.; Correia, S.; Correia, B.; Pinto, G.; Canhoto, J.M. Shoot proliferation and organogenesis on Arbutus unedo: Physiological analysis under water stress. Biol. Plant. 2019, 63, 278–286. [Google Scholar] [CrossRef]
- Villa, R.S. Ricerche sulla biologia di Arbutus unedo L. (Ericaceae): Ciclo di sviluppo. Boll. Soc. Sarda Sci. Nat. 1982, 21, 309–317. [Google Scholar]
- Mateus, J. Pollen morphology of Portuguese Ericales. Rev. Biol. 1989, 14, 135–208. [Google Scholar]
- Doorenbos, J. Shortening the breeding cycle of rhododendron. Euphytica 1955, 4, 141–146. [Google Scholar] [CrossRef]
- Escaravage, N.; Pornon, A.; Doche, B.; Till-Bottraud, I. Breeding system in an alpine species: Rhododendron ferrugineum L. (Ericaceae) in the French northern Alps. Can. J. Bot. 1997, 75, 736–743. [Google Scholar] [CrossRef]
- Lyrene, P.M. Value of various taxa in breeding tetraploid blueberries in Florida. Euphytica 1997, 94, 15–22. [Google Scholar] [CrossRef]
- Usui, M.; Kevan, P.G.; Obbard, M. Pollination and breeding system of lowbush blueberries, Vaccinium angustifolium Ait. and V. myrtilloides Michx. (Ericacaeae), in the boreal forest. Can. Field Nat. 2005, 119, 48–57. [Google Scholar] [CrossRef]
- Igarashi, M.; Hatsuyama, Y.; Harada, T.; Fukasawa-Akada, T. Biotechnology and apple breeding in Japan. Breed. Sci. 2016, 66, 18–33. [Google Scholar] [CrossRef] [PubMed]
- Aravanopoulos, F.A. Breeding of fast growing forest tree species for biomass production in Greece. Biomass Bioenergy 2010, 34, 1531–1537. [Google Scholar] [CrossRef]
- Fryxell, P. Mode of reproduction of higher plants. Bot. Rev. 1957, 23, 135–233. [Google Scholar] [CrossRef]
- Erdtman, G. The acetolysis method in a revised description. Sven. Bot. Tidskr. Lund 1960, 54, 561–564. [Google Scholar]
- Punt, W.; Hoen, P.P.; Blackmore, S.; Nilsson†, S.; Le Thomas, A. Glossary of pollen and spore terminology. Rev. Palaeobot. Palynol. 2007, 143, 1–81. [Google Scholar] [CrossRef]
- Spurr, A.R. A low-viscosity epoxy resin embedding medium for electron microscopy. J. Ultrastruct. Res. 1969, 26, 31–43. [Google Scholar] [CrossRef]
- Roland, J.C. General preparation and staining of thin sections. In Electron Microscopy and Cytochemistry of Plant Cells; Hall, J.L., Ed.; Elsevier/North-Holland Biomedical Press: Amsterdam, The Netherlands, 1978; pp. 1–62. [Google Scholar]
- Jahier, J.; Chevre, A.M.; Eber, F.; Delourme, R.; Tanguy, A.M. Techniques de Cytogenetique Vegetale; INRA: Paris, France, 1992; ISBN 9782738003966. [Google Scholar]
- Martin, F.W. Staining and observing pollen tubes in the style by means of fluorescence. Stain Technol. 1959, 34, 125–128. [Google Scholar] [CrossRef]
- Yang, J.; Lovett-Doust, J.; Lovett-Doust, L. Seed germination patterns in green dragon (Arisaema dracontium, Araceae). Am. J. Bot. 1999, 86, 1160–1167. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/2017 (accessed on 10 October 2020).
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [PubMed]
- Vu, V.Q. Ggbiplot: A Ggplot2 Based Biplot. R Package Version 0.55. Available online: http://github.com/vqv/ggbiplot (accessed on 10 October 2020).
- Hermann, P.M.; Palser, B.F. Stamen development in the Ericaceae. I. Anther wall, microsporogenesis, inversion, and appendages. Am. J. Bot. 2000, 87, 934–957. [Google Scholar] [CrossRef] [PubMed]
- Ćalić, D.; Devrnja, N.; Kostić, I.; Kostić, M. Pollen morphology, viability, and germination of Prunus domestica cv. PoŽegača. Sci. Hortic. 2013, 155, 118–122. [Google Scholar] [CrossRef]
- Acar, I.; Gopal, V. The effects of temperature on in vitro pollen germination and pollen tube growth of Pistacia spp. Sci. Hortic. 2010, 125, 569–572. [Google Scholar] [CrossRef]
- Rosell, P.; Herrero, M.; Galán Saúco, V. Pollen germination of cherimoya (Annona cherimola Mill.). Sci. Hortic. 1999, 81, 251–265. [Google Scholar] [CrossRef]
- Vuletin Selak, G.; Perica, S.; Goreta Ban, S.; Poljak, M. The effect of temperature and genotype on pollen performance in olive (Olea europaea L.). Sci. Hortic. 2013, 156, 38–46. [Google Scholar] [CrossRef]
- Cane, J.H. Pollen viability and pollen tube attrition in cranberry (Vaccinium macrocarpon Aiton). Acta Hortic. 2009, 810, 563–566. [Google Scholar] [CrossRef]
- Okusaka, K.; Hiratsuka, S. Fructose inhibits pear pollen germination on agar medium without loss of viability. Sci. Hortic. 2009, 122, 51–55. [Google Scholar] [CrossRef]
- Nátrová, Z.; Hlaváč, M. The effect of gametocides on microsporogenesis of winter rye. Biol. Plant. 1975, 17, 256–262. [Google Scholar] [CrossRef]
- Verma, M.M.; Kumar, J. Ethrel-a male gametocide that can replace the male sterility genes in barley. Euphytica 1978, 27, 865–868. [Google Scholar] [CrossRef]
- Saimbhi, M.S.; Brar, J.S. A review of the practical use of gametocides on vegetable crops. Sci. Hortic. 1978, 8, 11–17. [Google Scholar] [CrossRef]
- Chakraborty, K.; Devakumar, C. Ethyloxanilates as specific male gametocides for wheat (Triticum aestivum L.). Plant Breed. 2006, 125, 441–447. [Google Scholar] [CrossRef]
- Voyiatzis, D.G.; Paraskevopoulou-Paroussi, G. Factors affecting the quality and in vitro germination capacity of strawberry pollen. J. Hortic. Sci. Biotechnol. 2002, 77, 200–203. [Google Scholar] [CrossRef]
- Cano-Medrano, R.; Darnell, R.L. Effect of GA3 and pollination on fruit set and development in rabbiteye blueberry. Hortic. Sci. 1998, 33, 632–635. [Google Scholar] [CrossRef]
- Horsley, T.N.; Johnson, S.D. Is Eucalyptus cryptically self-incompatible? Ann. Bot. 2007, 100, 1373–1378. [Google Scholar] [CrossRef]
- Kaufmane, E.; Rumpunen, K. Pollination, pollen tube growth and fertilization in Chaenomeles japonica (Japanese quince). Sci. Hortic. 2002, 94, 257–271. [Google Scholar] [CrossRef]
- Demirsoy, L.; Demirsoy, H.; Celikel, G.; Macit, I.; Ersoy, B. Seed treatment with GA3 or stratification enhances emergence of some strawberry tree genotypes—Short communication. Hortic. Sci. 2010, 37, 34–37. [Google Scholar] [CrossRef]
- Ertekin, M.; Kirdar, E. Breaking seed dormancy of strawberry tree (Arbutus unedo). Int. J. Agric. Biol. 2010, 12, 57–60. [Google Scholar]
- Tilki, F. Improvement in seed germination of Arbutus unedo L. Pak. J. Biol. Sci. 2004, 7, 1640–1642. [Google Scholar] [CrossRef]
- Castell, C.; Terradas, J. Effects of water and nutrient availability on water relations, gas exchange and growth rate of mature plants and resprouts of Arbutus unedo L. Ann. Bot. 1994, 73, 595–602. [Google Scholar] [CrossRef]
- Raimondo, F.; Trifilò, P.; Lo Gullo, M.A.; Andri, S.; Savi, T.; Nardini, A. Plant performance on Mediterranean green roofs: Interaction of species-specific hydraulic strategies and substrate water relations. AOB Plants 2015, 7, plv007. [Google Scholar] [CrossRef] [PubMed]
- Vasques, A.; Chirino, E.; Vilagrosa, A.; Vallejo, V.R.; Keizer, J.J. The role of seed provenance in the early development of Arbutus unedo seedlings under contrasting watering conditions. Environ. Exp. Bot. 2013, 96, 11–19. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | ci | E | gs | A | RWC | WP | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| t0 | t3 | t0 | t3 | t0 | t3 | t0 | t3 | |||
| 1 | 181 | 166 | 2.76 | 0.86 | 0.32 | 0.06 | 25.87 | 7.67 | 63.4 | −50 * |
| 2 | 243 | 220 | 1.98 | 0.34 | 0.18 | 0.02 | 11.81 | 2.31 | 72.8 | −50 * |
| 3 | 181 | 159 | 3.5 | 0.64 | 0.51 | 0.04 | 32.36 | 5.49 | 70.5 | −50 * |
| 4 | 215 | 302 | 2.88 | 0.14 | 0.32 | 0.01 | 21.2 | 0.45 | 47.5 | −50 * |
| 5 | 176 | 223 | 3.34 | 0.55 | 0.42 | 0.03 | 29.84 | 3.07 | 70.9 | −50 * |
| 6 | 193 | 258 | 1.98 | 0.53 | 0.16 | 0.03 | 14.7 | 2.21 | 59.4 | −50 * |
| 7 | 186 | 204 | 2.48 | 0.84 | 0.23 | 0.05 | 19.59 | 5.18 | 63.9 | −50 * |
| 8 | 172 | 230 | 2.91 | 0.45 | 0.31 | 0.02 | 25.23 | 2.32 | 53.0 | −50 * |
| 9 | 209 | 364 | 2.82 | 0.19 | 0.28 | 0.01 | 19.85 | 0.15 | 48.6 | −50 * |
| 10 | 205 | 296 | 3.52 | 0.29 | 0.43 | 0.02 | 26.22 | 0.82 | 50.0 | −50 * |
| 11 | 192 | 435 | 2.96 | 0.27 | 0.3 | 0.01 | 22.51 | −0.46 | 45.7 | −50 * |
| 12 + | 196 | 220 | 2.96 | 2.52 | 0.3 | 0.22 | 22.1 | 15.61 | 84.6 | −21 |
| 13 + | 165 | 165 | 3.5 | 1.63 | 0.43 | 0.11 | 31.25 | 12.51 | 81.2 | −32 |
| 14 ++ | 160 | 171 | 2.11 | 2.7 | 0.16 | 0.23 | 16.92 | 20.27 | 84.7 | −24 |
| 15 ++ | 163 | 201 | 2.8 | 3.27 | 0.24 | 0.33 | 22.5 | 22.19 | 79.4 | −14 |
| 16 | 179 | 232 | 2.89 | 0.63 | 0.24 | 0.03 | 20.86 | 2.89 | 59.6 | −50 * |
| 17 + | 199 | 190 | 1.87 | 1.59 | 0.12 | 0.1 | 11.24 | 10.47 | 78.8 | −26 |
| 18 ++ | 139 | 208 | 2.58 | 3.41 | 0.2 | 0.34 | 20.96 | 21.7 | 71.7 | −10.5 |
| 19 | 157 | 305 | 2.85 | 0.91 | 0.22 | 0.05 | 21.26 | 2.01 | 73.3 | −18.5 |
| 20 | 163 | 260 | 2.82 | 0.68 | 0.22 | 0.03 | 20.68 | 2.45 | 44.0 | −50 * |
| 21 | 105 | 302 | 3.28 | 0.63 | 0.26 | 0.03 | 29.13 | 1.24 | 74.0 | −31.25 |
| 22 | 125 | 260 | 2.93 | 0.6 | 0.23 | 0.03 | 24.77 | 2.16 | 42.3 | −50 * |
| 23 | 76 | 188 | 2.64 | 0.64 | 0.18 | 0.03 | 25.25 | 3.73 | 77.6 | −42.5 |
| 24 | 87 | 191 | 2.72 | 0.92 | 0.19 | 0.05 | 25.27 | 5.3 | 58.2 | −48.5 |
| 25 | 191 | 325 | 3.28 | 0.19 | 0.46 | 0.01 | 29.66 | 0.55 | 50.6 | −50 * |
| 26 | 246 | 377 | 2.42 | 0.33 | 0.27 | 0.02 | 16.36 | 0.02 | 50.5 | −50 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins, J.; Monteiro, P.; Pinto, G.; Canhoto, J. Hybridization Assays in Strawberry Tree toward the Identification of Plants Displaying Increased Drought Tolerance. Forests 2021, 12, 148. https://doi.org/10.3390/f12020148
Martins J, Monteiro P, Pinto G, Canhoto J. Hybridization Assays in Strawberry Tree toward the Identification of Plants Displaying Increased Drought Tolerance. Forests. 2021; 12(2):148. https://doi.org/10.3390/f12020148
Chicago/Turabian StyleMartins, João, Pedro Monteiro, Glória Pinto, and Jorge Canhoto. 2021. "Hybridization Assays in Strawberry Tree toward the Identification of Plants Displaying Increased Drought Tolerance" Forests 12, no. 2: 148. https://doi.org/10.3390/f12020148
APA StyleMartins, J., Monteiro, P., Pinto, G., & Canhoto, J. (2021). Hybridization Assays in Strawberry Tree toward the Identification of Plants Displaying Increased Drought Tolerance. Forests, 12(2), 148. https://doi.org/10.3390/f12020148