
SilvAdapt.Net: A Site-Based Network of Adaptive Forest Management Related to Climate Change in Spain

 , ,
, ,  ,
,  , , , , , , , and add
Show full author list
, , , , , , , and add
Show full author list
Abstract
:1. Introduction
1.1. Forests and Climate Change
1.2. The Need for Adaptive Forest Management
1.3. The Need of Coordinating Efforts among Research Groups Addressing AFM in Spain
1.4. Objectives
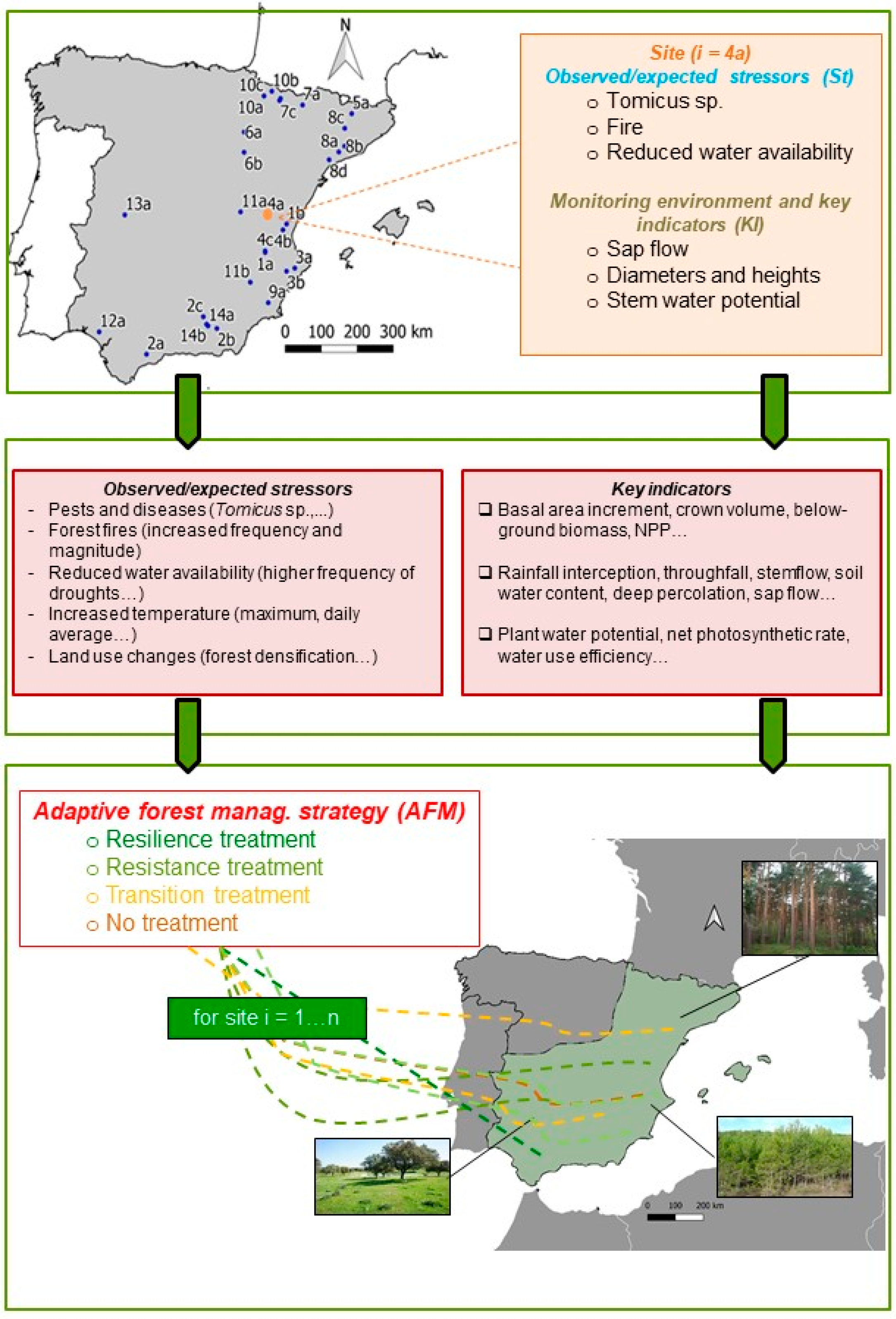
2. Implementing Adaptive Forest Management in Spain
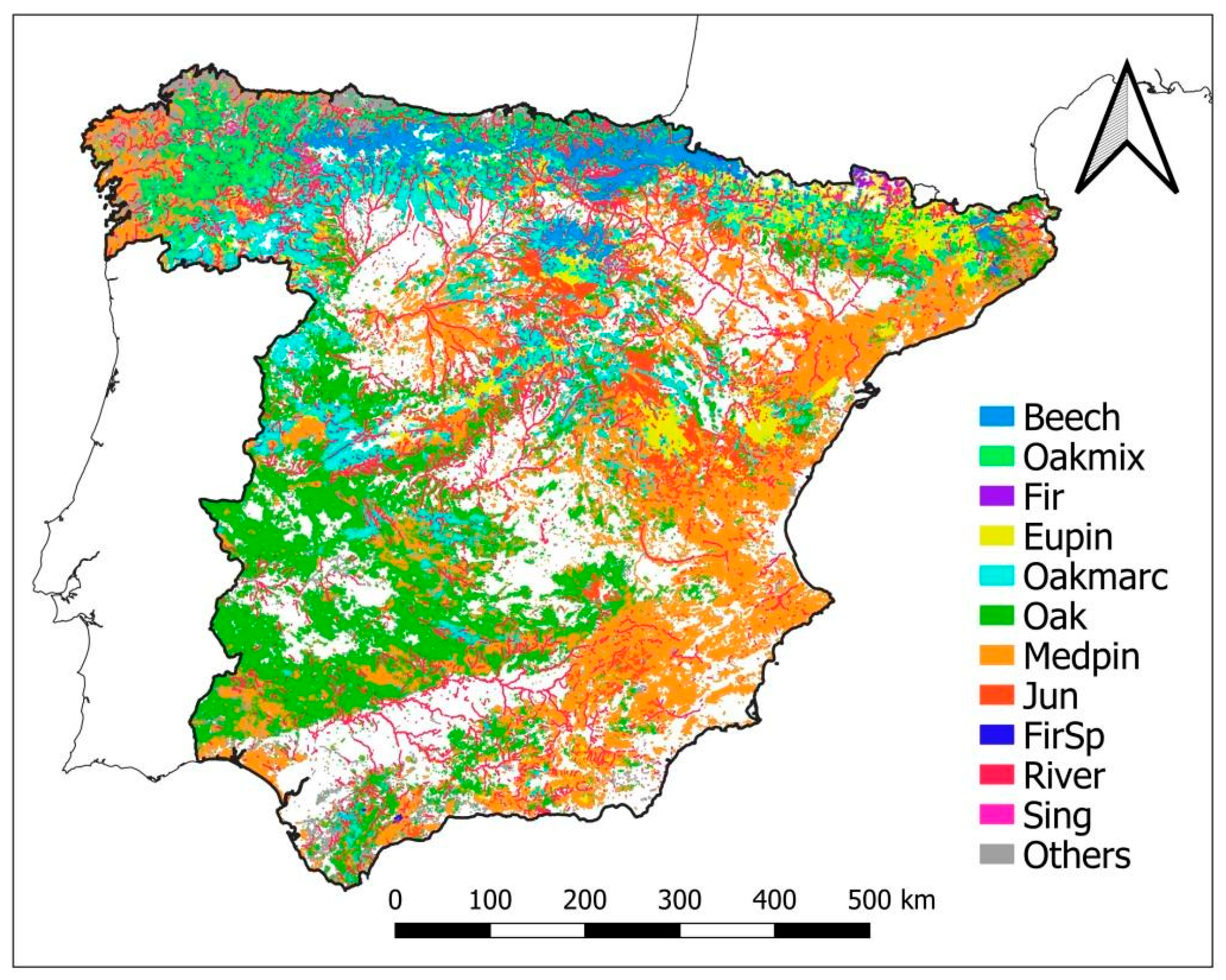
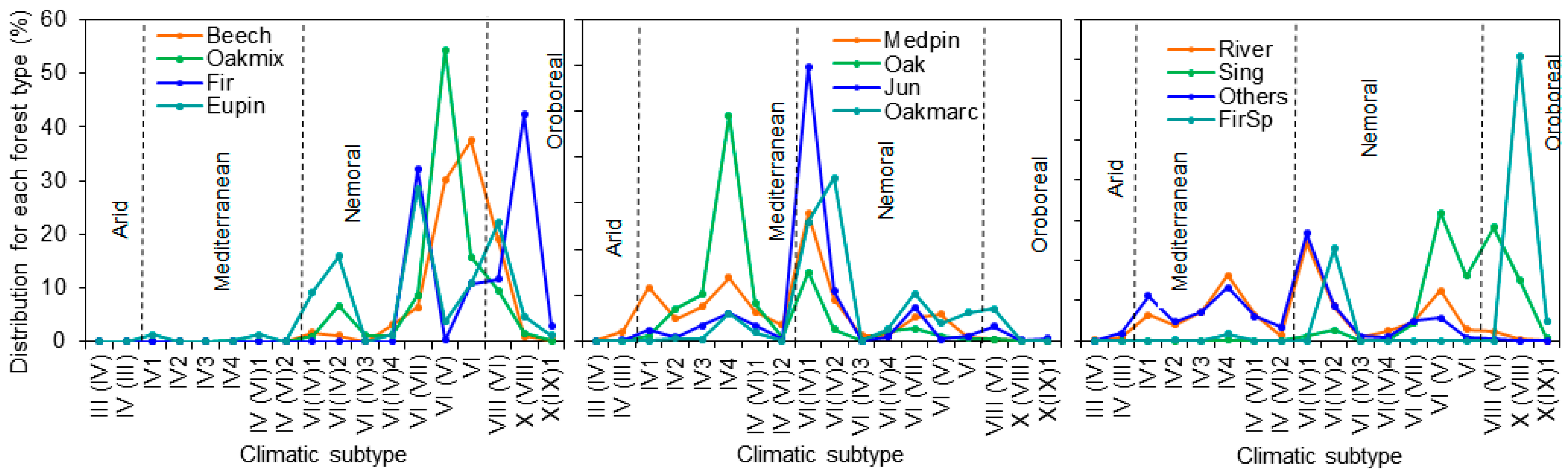
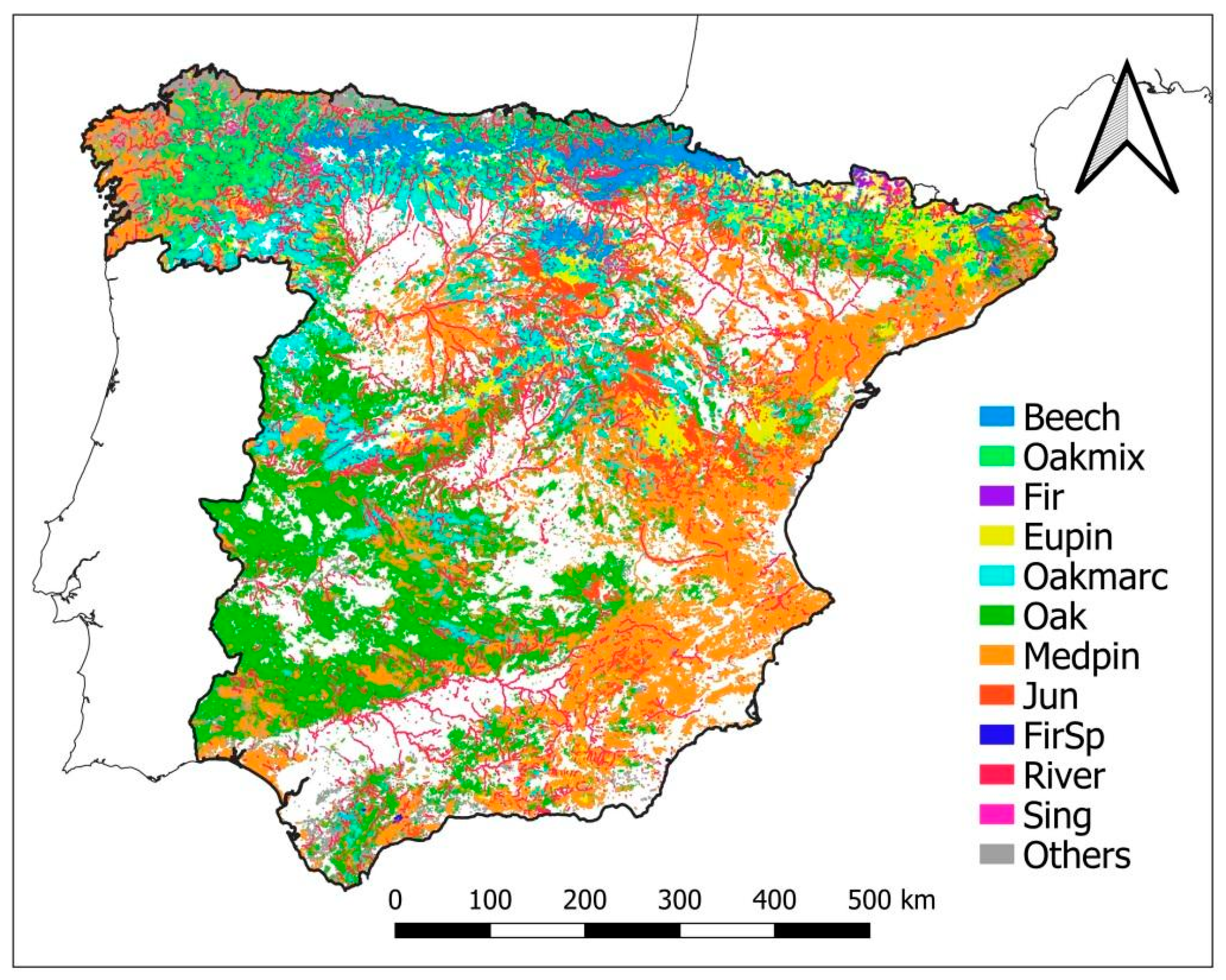
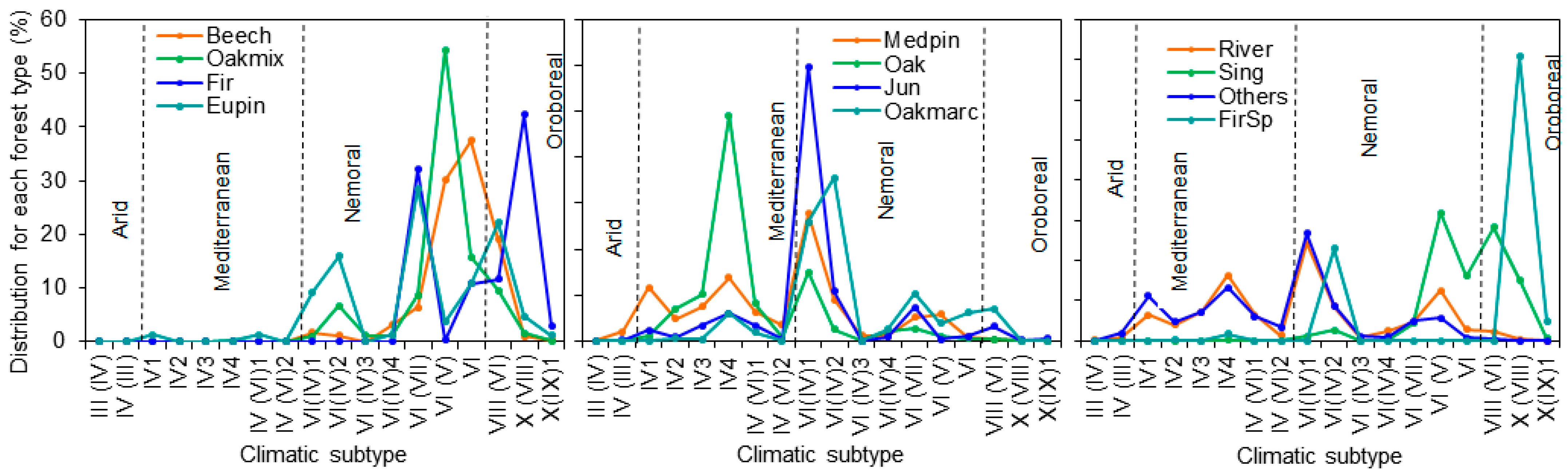
2.1. Framing Different Climates and Forest Types into a Nation-Wide Approach of AFM
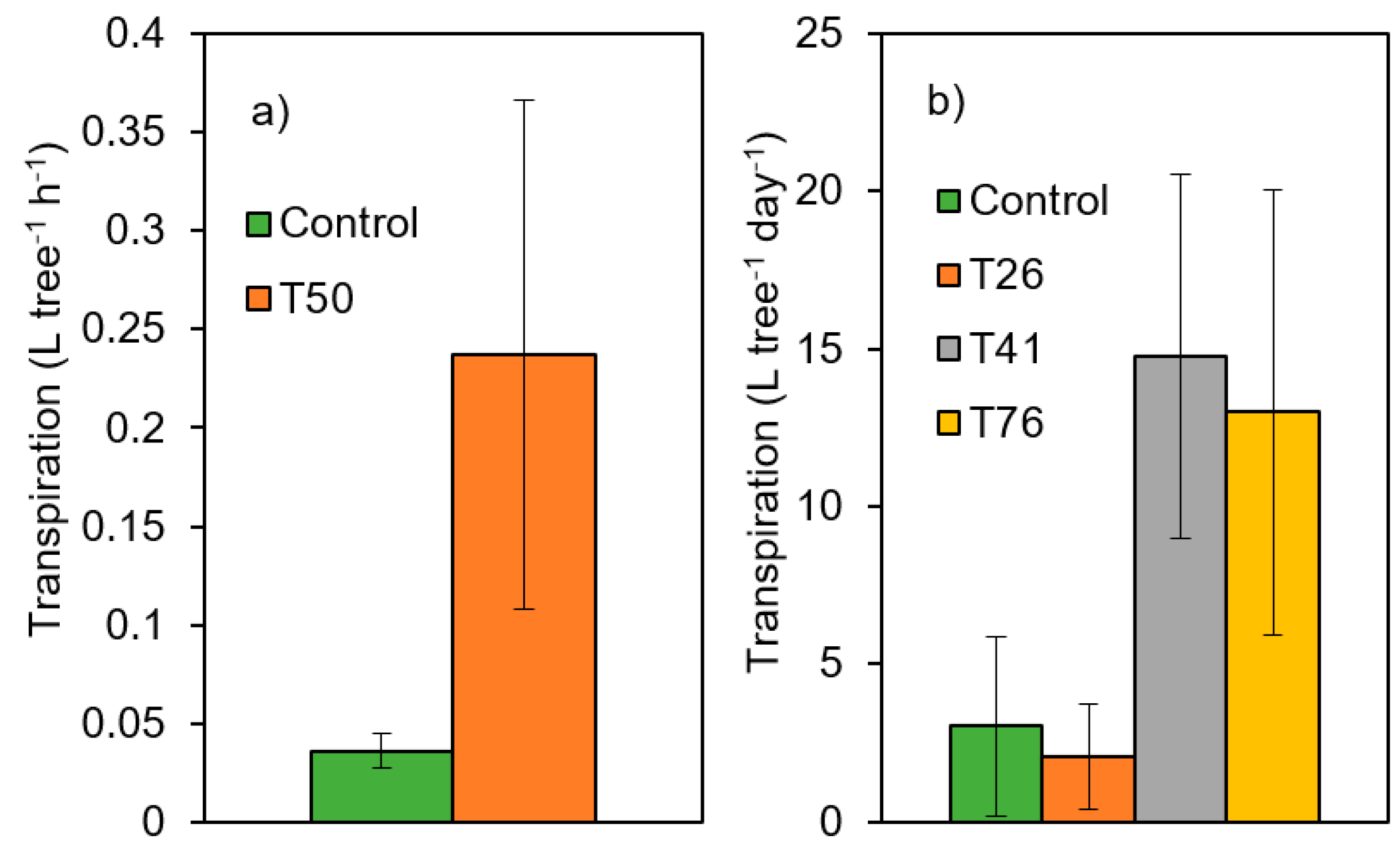
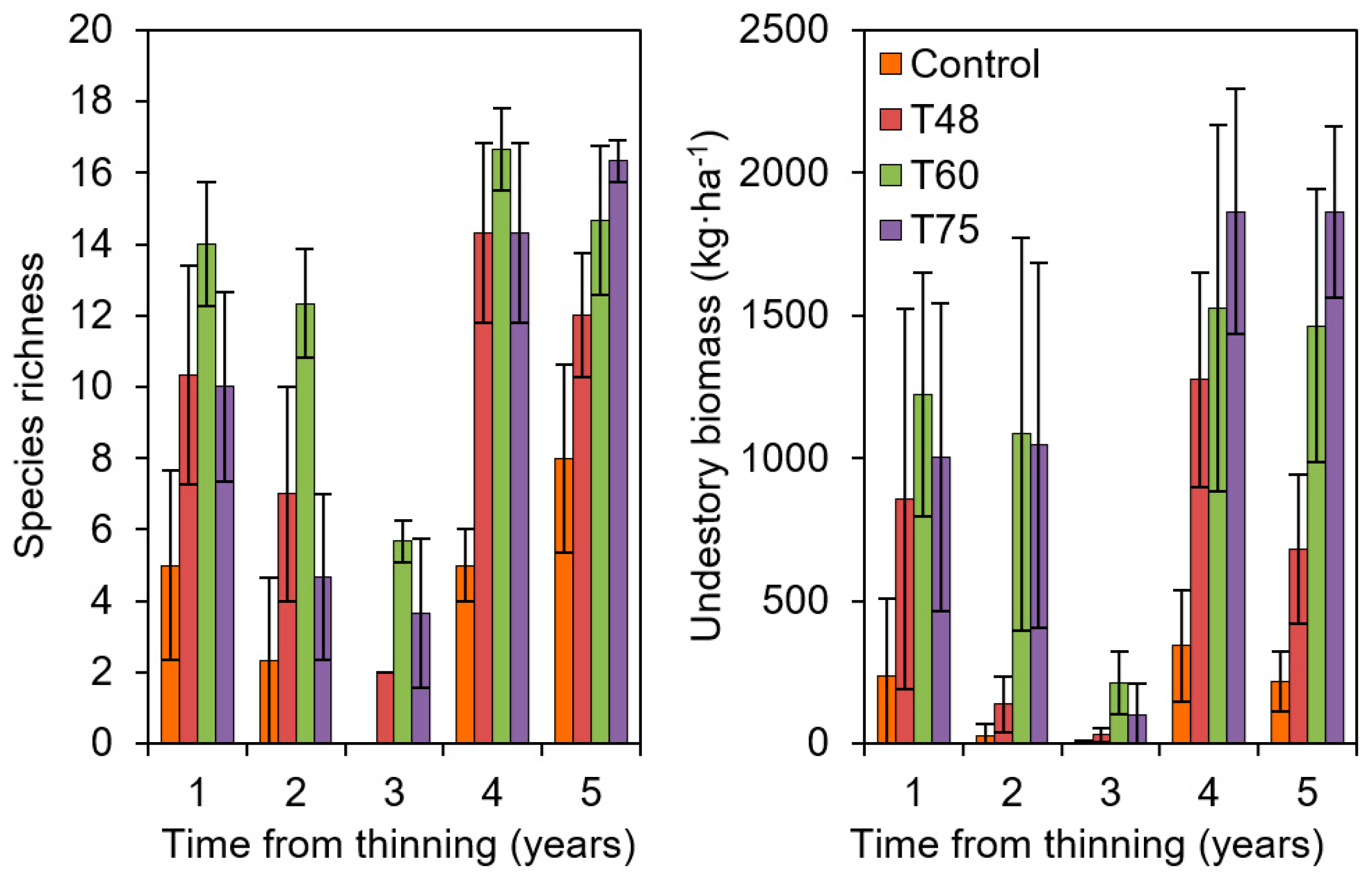
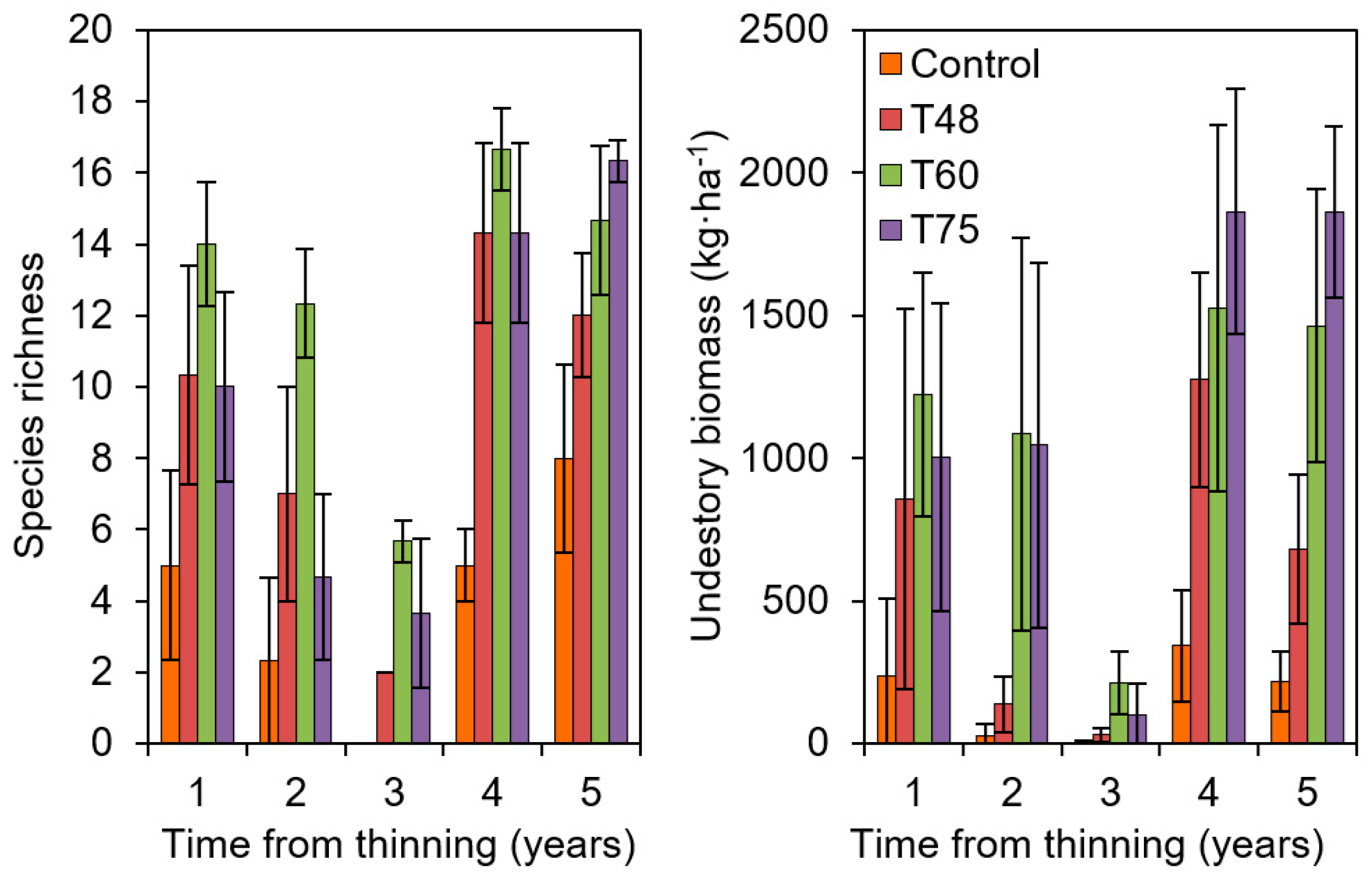
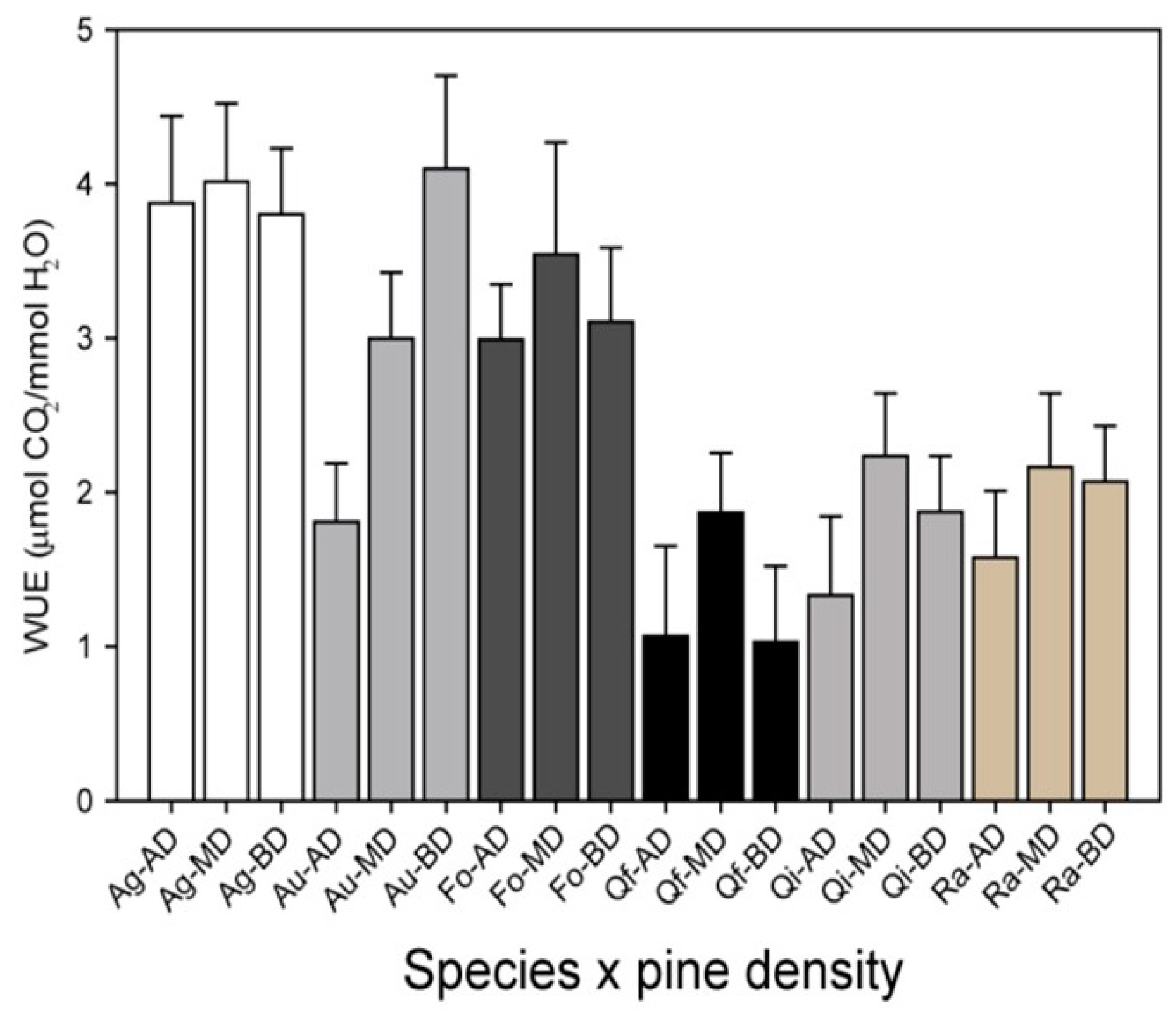
2.2. Observed Effects of AFM in the Network: The Case of Thinning Intensity in Mediterranean Pines
2.3. Upscaling Site Observations into a More Meaningful Framework. Open Questions and Next Steps
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Variables Measured in the Experimental Sites
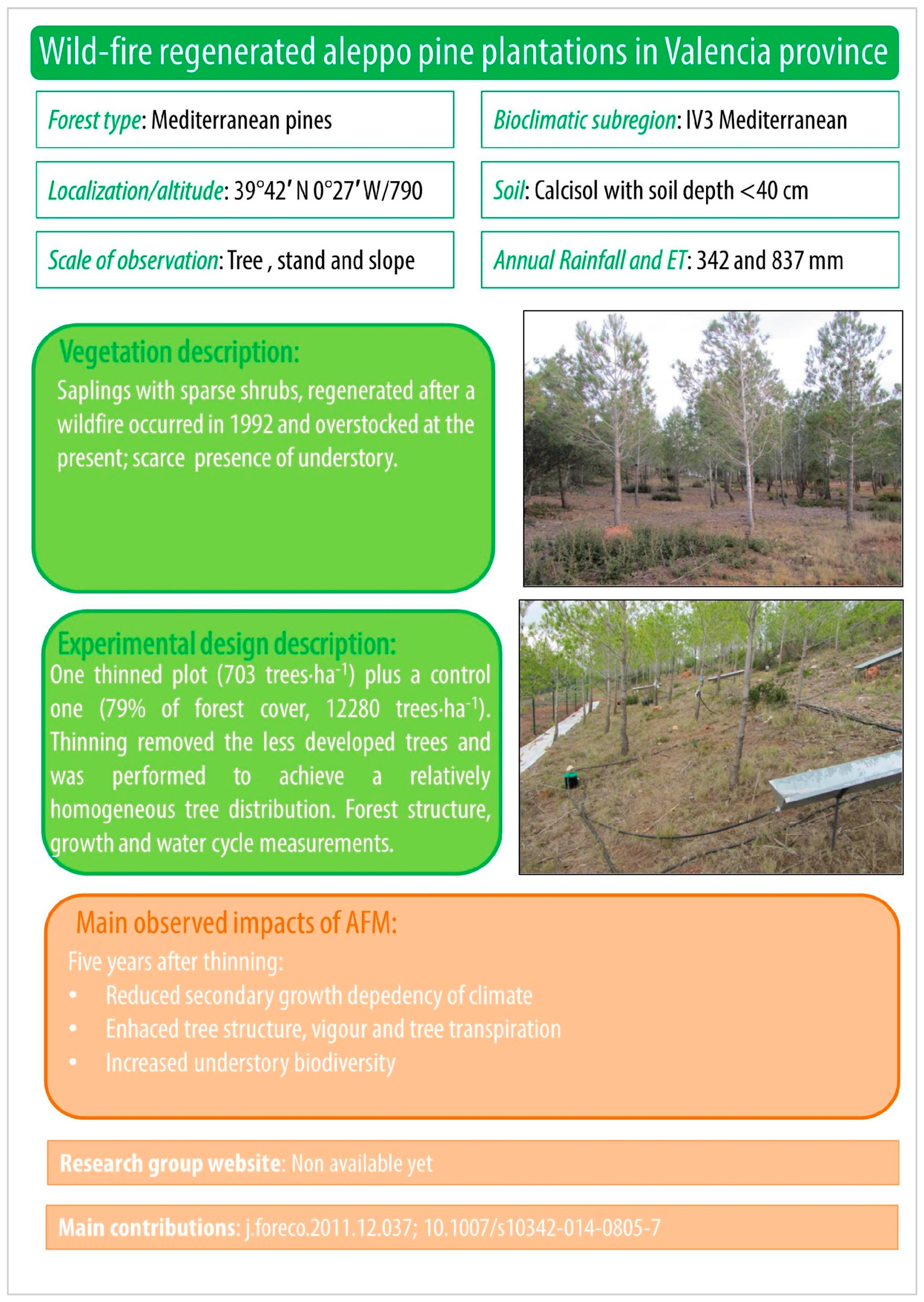
Appendix B. Collection of Cards Summarizing Vegetation Description, Experimental Approach and Main AFM Impacts for Some Sites with Mediterranean Pines within the Silvadapt.Net Network





References
- Vayreda, J.; Martínez-Vilalta, J.; Gracia, M.; Retana, J. Recent climate changes interact with stand structure and management to determine changes in tree carbon stocks in Spanish forests. Glob. Chang. Biol. 2012, 18, 1028–1041. [Google Scholar] [CrossRef]
- Lindner, M.; Fitzgerald, J.B.; Zimmermann, N.E.; Reyer, C.; Delzon, S.; van der Maaten, E.; Schelhaas, M.; Lasch, P.; Eggers, J.; der Maaten-Theunissen, M. Climate change and European forests: What do we know, what are the uncertainties, and what are the implications for forest management? J. Environ. Manag. 2014, 146, 69–83. [Google Scholar] [CrossRef] [Green Version]
- Amblar-Francés, M.P.; Casado Calle, M.J.; Pastor Saavedra, M.A.; Ramos Calzado, P.; Rodríguez Camino, E. Guía de Escenarios Regionalizados de Cambio Climático Sobre España a Partir de los Resultados del IPCC-AR5; AEMET: Madrid, Spain, 2017. [Google Scholar]
- Serrano-Notivoli, R.; Beguería, S.; Saz, M.Á.; de Luis, M. Recent trends reveal decreasing intensity of daily precipitation in Spain. Int. J. Climatol. 2018, 38, 4211–4224. [Google Scholar] [CrossRef]
- Anderegg, W.R. Spatial and temporal variation in plant hydraulic traits and their relevance for climate change impacts on vegetation. New Phytol. 2015, 205, 1008–1014. [Google Scholar] [CrossRef]
- Mastrotheodoros, T.; Pappas, C.; Molnar, P.; Burlando, P.; Keenan, T.F.; Gentine, P.; Gough, C.M.; Fatichi, S. Linking plant functional trait plasticity and the large increase in forest water use efficiency. J. Geophys. Res. Biogeosci. 2017, 122, 2393–2408. [Google Scholar] [CrossRef]
- Metzger, M.J.; Bunce, R.G.H.; Leemans, R.; Viner, D. Projected environmental shifts under climate change: European trends and regional impacts. Environ. Conserv. 2018, 35, 64–75. [Google Scholar] [CrossRef] [Green Version]
- Soteriades, A.D.; Murray-Rust, D.; Trabucco, A.; Metzger, M.J. Understanding global climate change scenarios through bioclimate stratification. Environ. Res. Lett. 2017, 12, 084002. [Google Scholar] [CrossRef]
- Puettmann, K.J. Silvicultural challenges and options in the context of global change: “Simple” fixes and opportunities for new management approaches. J. For. 2011, 109, 321–331. [Google Scholar]
- Keenan, T.; Maria Serra, J.; Lloret, F.; Ninyerola, M.; Sabate, S. Predicting the future of forests in the Mediterranean under climate change, with niche-and process-based models: CO2 matters! Glob. Chang. Biol. 2011, 17, 565–579. [Google Scholar] [CrossRef]
- Reyer, C.P.; Leuzinger, S.; Rammig, A.; Wolf, A.; Bartholomeus, R.P.; Bonfante, A.; de Lorenzi, F.; Dury, M.; Gloning, P.; Jaoudé, R.A.; et al. A plant’s perspective of extremes: Terrestrial plant responses to changing climatic variability. Glob. Chang. Biol. 2013, 19, 75–89. [Google Scholar] [CrossRef]
- Roselló, E. Biogeoclimatic Classification of Peninsular and Balearic Spain; Ministerio de Agricultura, Pesca y Alimentacion: Madrid, Spain, 1997. [Google Scholar]
- Lenoir, J.; Gégout, J.C.; Marquet, P.A.; De Ruffray, P.; Brisse, H. A significant upward shift in plant species optimum elevation during the 20th century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Kearney, M.; Porter, W. Mechanistic niche modelling: Combining physiological and spatial data to predict species’ ranges. Ecol. Lett. 2009, 12, 334–350. [Google Scholar] [CrossRef]
- Seidl, R.; Spies, T.A.; Peterson, D.L.; Stephens, S.L.; Hicke, J.A. Searching for resilience: Addressing the impacts of changing disturbance regimes on forest ecosystem services. J. Appl. Ecol. 2016, 53, 120–129. [Google Scholar] [CrossRef] [Green Version]
- Moriondo, M.; Good, P.; Durao, R.; Bindi, M.; Giannakopoulos, C.; Corte-Real, J. Potential impact of climate change on fire risk in the Mediterranean area. Clim. Res. 2006, 31, 85–95. [Google Scholar] [CrossRef]
- de la Cueva, A.V.; Quintana, J.R.; Cañellas, I. Fire activity projections in the SRES A2 and B2 climatic scenarios in peninsular Spain. Int. J. Wildland Fire 2012, 21, 653–665. [Google Scholar] [CrossRef]
- Vázquez, A.; Climent, J.M.; Casais, L.; Nieto, J.R.Q. Current and future estimates for the fire frequency and the fire rotation period in the main woodland types of peninsular Spain: A case-study approach. For. Syst. 2015, 24, 10. [Google Scholar] [CrossRef] [Green Version]
- Doblas-Miranda, E.; Alonso, R.; Arnan, X.; Bermejo, V.; Brotons, L.; De las Heras, J.; Estiarte, M.; Hódar, J.A.; Llorens, P.; Lloret, F.; et al. A review of the combination among global change factors in forests, shrublands and pastures of the Mediterranean Region: Beyond drought effects. Glob. Planet. Chang. 2017, 148, 42–54. [Google Scholar] [CrossRef] [Green Version]
- Peñuelas, J.; Lloret, F.; Montoya, R. Severe drought effects on Mediterranean woody flora in Spain. For. Sci. 2001, 47, 214–218. [Google Scholar]
- Pasho, E.; Camarero, J.J.; de Luis, M.; Vicente-Serrano, S.M. Impacts of drought at different time scales on forest growth across a wide climatic gradient in north-eastern Spain. Agric. For. Meteorol. 2011, 151, 1800–1811. [Google Scholar] [CrossRef]
- de la Serrana, R.G.; Vilagrosa, A.; Alloza, J.A. Pine mortality in southeast Spain after an extreme dry and warm year: Interactions among drought stress, carbohydrates and bark beetle attack. Trees 2015, 29, 1791–1804. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Cantero, A.; Sánchez-Salguero, R.; Sánchez-Miranda, A.; Granda, E.; Serra-Maluquer, X.; Ibáñez, R. Forest growth responses to drought at short- and long-term scales in Spain: Squeezing the stress memory from tree rings. Front. Ecol. Evol. 2018, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Peña-Gallardo, M.; Vicente-Serrano, S.; Camarero, J.J.; Gazol, A.; Sánchez-Salguero, R.; Domínguez-Castro, F.; el Kenawy, A.; Beguería-Portugés, S.; Gutiérrez, E.; de Luis, M.; et al. Drought sensitiveness on forest growth in peninsular Spain and the Balearic Islands. Forests 2018, 9, 524. [Google Scholar] [CrossRef] [Green Version]
- Morcillo, L.; Gallego, D.; González, E.; Vilagrosa, A. Forest decline triggered by phloem parasitism-related biotic factors in Aleppo pine (Pinus halepensis). Forests 2019, 10, 608. [Google Scholar] [CrossRef] [Green Version]
- De Dios, V.R.; Fischer, C.; Colinas, C. Climate change effects on Mediterranean forests and preventive measures. New For. 2007, 33, 29–40. [Google Scholar] [CrossRef]
- Prieto-Recio, C.; Martín-García, J.; Bravo, F.; Diez, J.J. Unravelling the associations between climate, soil properties and forest management in Pinus pinaster decline in the Iberian Peninsula. For. Ecol. Manag. 2015, 356, 74–83. [Google Scholar] [CrossRef]
- Pautasso, M.; Dehnen-Schmutz, K.; Holdenrieder, O.; Pietravalle, S.; Salama, N.; Jeger, M.J.; Lange, E.; Hehl-Lange, S. Plant health and global change–some implications for landscape management. Biol. Rev. 2010, 85, 729–755. [Google Scholar] [CrossRef] [PubMed]
- Sturrock, R.N.; Frankel, S.J.; Brown, A.V.; Hennon, P.E.; Kliejunas, J.T.; Lewis, K.J.; .Worrall, J.J.; Woods, A.J. Climate change and forest diseases. Plant Pathol. 2011, 60, 133–149. [Google Scholar] [CrossRef]
- MEA. Millennium Ecosystem Assessment: Ecosystems and Human Well-Being 5; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- Millar, C.I.; Stephenson, N.L.; Stephens, S.L. Climate change and forests of the future: Managing in the face of uncertainty. Ecol. Appl. 2007, 17, 2145–2151. [Google Scholar] [CrossRef]
- Janowiak, M.K.; Swanston, C.W.; Nagel, L.M.; Brandt, L.A.; Butler, P.R.; Handler, S.D.; Shannon, P.D.; Iverson, L.R.; Matthews, S.N.; Prasad, A.; et al. A practical approach for translating climate change adaptation principles into forest management actions. J. For. 2014, 112, 424–433. [Google Scholar] [CrossRef] [Green Version]
- Nagel, L.M.; Palik, B.J.; Battaglia, M.A.; D’Amato, A.W.; Guldin, J.M.; Swanston, C.W.; Janowiak, M.K.; Powers, M.P.; Joyce, L.A.; Millar, C.I.; et al. Adaptive silviculture for climate change: A national experiment in manager-scientist partnerships to apply an adaptation framework. J. For. 2017, 115, 167–178. [Google Scholar] [CrossRef]
- del Campo, A.D.; Fernandes, T.J.; Molina, A.J. Hydrology-Oriented (adaptive) silviculture in a semiarid pine plantation: How much can be modified the water cycle through forest management? Eur. J. For. Res. 2014, 133, 879–894. [Google Scholar] [CrossRef]
- Navarro-Cerrillo, R.M.; Sánchez-Salguero, R.; Herrera, R.; Ruiz, C.C.; Moreno-Rojas, J.M.; Manzanedo, R.D.; López-Quintanilla, J. Contrasting growth and water use efficiency after thinning in mixed Abies pinsapo–Pinus pinaster–Pinus sylvestris forests. J. For. Sci. 2016, 62, 53–64. [Google Scholar] [CrossRef]
- Navarro-Cerrillo, R.M.; Sánchez-Salguero, R.; Rodriguez, C.; Lazo, J.D.; Moreno-Rojas, J.M.; Palacios-Rodriguez, G.; Camarero, J.J. Is thinning an alternative when trees could die in response to drought? The case of planted Pinus nigra and P. sylvestris stands in southern Spain. For. Ecol. Manag. 2019, 433, 313–324. [Google Scholar] [CrossRef]
- Felicísimo, A.M.; Muñoz, J.; Mateo, R.G.; Villalba, C.J. Vulnerabilidad de la flora y vegetación españolas ante el cambio climático. Ecosistemas 2012, 21, 1–6. [Google Scholar] [CrossRef]
- Fernández-Cancio, Á.; Sánchez Salguero, R.; Gil Hernández, P.M.; Manrique Menéndez, E.; Fernández Fernández, R.; Navarro Cerrillo, R.M. Efectos del cambio climático sobre la distribución de los alcornocales españoles: Una aproximación fitoclimática para la futura gestión. Ecosistemas 2012, 21, 50–62. [Google Scholar] [CrossRef] [Green Version]
- García-Güemes, C.; Calama, R. La Práctica de la Selvicultura Para la Adaptación al Cambio Climático. Los Bosques y la Biodiversidad Frente al Cambio Climático: Impactos, Vulnerabilidad y Adaptación en España, 12; Miteco: Madrid, Spain, 2014. [Google Scholar]
- Ruiz-Benito, P.; Herrero, A.; Zavala, M.Á. Vulnerabilidad de los bosques Ibéricos frente al Cambio Climático: Evaluación mediante modelos. Ecosistemas 2014, 22, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Tsamir, M.; Gottlieb, S.; Preisler, Y.; Rotenberg, E.; Tatarinov, F.; Yakir, D.; Tague, C.; Klein, T. Stand density effects on carbon and water fluxes in a semi-arid forest, from leaf to stand-scale. For. Ecol. Manag. 2019, 453, 117573. [Google Scholar] [CrossRef]
- Molina, A.J.; González-Sanchis, M.; Biel, C.; del Campo, A.D. Ecohydrological turnover in overstocked Aleppo pine plantations: Does the effect of thinning, in relation to water, persist at the mid-term? For. Ecol. Manag. 2021, 483, 118781. [Google Scholar] [CrossRef]
- Agee, J.K.; Skinner, C.N. Basic principles of forest fuel reduction treatments. For. Ecol. Manag. 2005, 211, 83–96. [Google Scholar] [CrossRef]
- Forestry Commission. Building Wildfire Resilience into Forest Management Planning. Building Wildfire Resilience into Forest Management Planning; Forestry Commission: Edinburgh, UK, 2014. [Google Scholar]
- Vilà-Cabrera, A.; Coll, L.; Martínez-Vilalta, J.; Retana, J. Forest management for adaptation to climate change in the Mediterranean basin: A synthesis of evidence. For. Ecol. Manag. 2018, 407, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H.; del Río, M.; Biber, P.; Arcangeli, C.; Bielak, K.; Brang, P.; Dudzinska, M.; Forrester, D.I.; Klädtke, J.; Kohnle, U.; et al. Maintenance of long-term experiments for unique insights into forest growth dynamics and trends: Review and perspectives. Eur. J. For. Res. 2019, 138, 165–185. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Benito, P.; Vacchiano, G.; Lines, E.R.; Reyer, C.P.; Ratcliffe, S.; Morin, X.; Hartig, F.; Mäkelä, A.; Yousefpour, R.; Chaves, J.E.; et al. Available and missing data to model impact of climate change on European forests. Ecol. Model. 2020, 416, 108870. [Google Scholar] [CrossRef]
- Michel, A.; Seidling, W. Forest Condition in Europe: 2016 Technical Report of ICP Forests: Report under the UNECE Convention on Long-Range Transboundary Air Pollution (CLRTAP); International Cooperative Programme on Assessment and Monitoring of Air Pollution Effects on Forests: Vienna, Austria, 2016; ISBN 978-3-902762-65-8. [Google Scholar]
- Alberdi, I.; Sandoval, V.; Condes, S.; Cañellas, I.; Vallejo, R. El Inventario Forestal Nacional español, una herramienta para el conocimiento, la gestión y la conservación de los ecosistemas forestales arbolados. Ecosistemas 2016, 25, 88–97. [Google Scholar] [CrossRef]
- Astigarraga, J.; Andivia, E.; Zavala, M.A.; Gazol, A.; Cruz-Alonso, V.; Vicente-Serrano, S.M.; Ruiz-Benito, P. Evidence of non-stationary relationships between climate and forest responses: Increased sensitivity to climate change in Iberian forests. Glob. Chang. Biol. 2020, 26, 5063–5076. [Google Scholar] [CrossRef] [PubMed]
- Tramblay, Y.; Koutroulis, A.; Samaniego, L.; Vicente-Serrano, S.M.; Volaire, F.; Boone, A.; Le Page, M.; Llasat, M.C.; Albergel, C.; Burak, S.; et al. Challenges for drought assessment in the Mediterranean region under future climate scenarios. Earth-Sci. Rev. 2020, 210, 103348. [Google Scholar] [CrossRef]
- Allué Andrade, J.L. Atlas Fitoclimático de España Taxonomías, 1st ed.; Instituto Nacional de Investigaciones Agrarias: Madrid, Spain, 1990. [Google Scholar]
- Blanco, E.; Casado, M.A.; Costa, M.; Escribano, R.; García, M.; Génova, M.; Regato, P. Los Bosques ibéricos. Una Interpretación Geobotánica; Planeta: Barcelona, Spain, 1997; p. 598. [Google Scholar]
- Hartmann, H.; Moura, C.F.; Anderegg, W.R.; Ruehr, N.K.; Salmon, Y.; Allen, C.D.; Arndt, S.K.; Breshears, D.D.; Davi, H.; Galbraith, D.; et al. Research frontiers for improving our understanding of drought-induced tree and forest mortality. New Phytol. 2018, 218, 15–28. [Google Scholar] [CrossRef] [Green Version]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Oliva, J.; Vicente-Serrano, S.M. To die or not to die: Early warnings of tree dieback in response to a severe drought. J. Ecol. 2015, 103, 44–57. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Sanz, V. Site-Scale ecological marginality: Evaluation model and application to a case study. Ecol. Model. 2019, 408, 108739. [Google Scholar] [CrossRef]
- Navarro-Cerrillo, R.M.; Rodriguez-Vallejo, C.; Silveiro, E.; Hortal, A.; Palacios-Rodríguez, G.; Duque-Lazo, J.; Camarero, J.J. Cumulative drought stress leads to a loss of growth resilience and explains higher mortality in planted than in naturally regenerated Pinus pinaster stands. Forests 2018, 9, 358. [Google Scholar] [CrossRef] [Green Version]
- Bose, A.K.; Gessler, A.; Bolte, A.; Bottero, A.; Buras, A.; Cailleret, M.; Camarero, J.J.; Haeni, M.; Hereş, A.; Hevia, A.; et al. Growth and resilience responses of Scots pine to extreme droughts across Europe depend on predrought growth conditions. Glob. Chang. Biol. 2020, 26, 4521–4537. [Google Scholar] [CrossRef]
- Manrique-Alba, À.; Beguería, S.; Molina, A.J.; González-Sanchis, M.; Tomàs-Burguera, M.; Del Campo, A.D.; Colangelo, M.; Camarero, J.J. Long-Term thinning effects on tree growth, drought response and water use efficiency at two Aleppo pine plantations in Spain. Sci. Total Environ. 2020, 728, 138536. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, M.N.; Spotswood, E.N.; Cañadas, E.M.; Navarro, F.B. Stand management to reduce fire risk promotes understorey plant diversity and biomass in a semi-arid Pinus halepensis plantation. Appl. Veg. Sci. 2015, 18, 467–480. [Google Scholar] [CrossRef]
- Granados, M.E.; Chirino Miranda, E.; Gandía Navalón, C.; Vallejo Calzada, V.R.; Vilagrosa Carmona, A. Effect of light and soil moisture on physiological variables in six Mediterranean forest species planted under a pine forest canopy. Revista Chapingo Serie Ciencias Forestales y del Ambiente 2019, 25, 461–476. [Google Scholar] [CrossRef]
- Eastaugh, C.S.; Pötzelsberger, E.; Hasenauer, H. Assessing the impacts of climate change and nitrogen deposition on Norway spruce (Picea abies L. Karst) growth in Austria with BIOME-BGC. Tree Physiol. 2011, 31, 262–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masek, J.G.; Goward, S.N.; Kennedy, R.E.; Cohen, W.B.; Moisen, G.G.; Schleeweis, K.; Huang, C. United States forest disturbance trends observed using Landsat time series. Ecosystems 2013, 16, 1087–1104. [Google Scholar] [CrossRef] [Green Version]
- Moreno-de las Heras, M.; Bochet, E.; Monleón, V.; Espigares, T.; Nicolau, J.M.; Molina, M.J.; García-Fayos, P. Aridity induces nonlinear effects of human disturbance on precipitation-use efficiency of Iberian woodlands. Ecosystems 2018, 21, 1295–1305. [Google Scholar] [CrossRef]
- Turner, D.P.; Cohen, W.B.; Kennedy, R.E.; Fassnacht, K.S.; Briggs, J.M. Relationships between leaf area index and Landsat TM spectral vegetation indices across three temperate zone sites. Remote Sens. Environ. 1999, 70, 52–68. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Main Objective | Main Tree Species | Forest Treat | Experimental Design | A | B | C | D | E | F | G | H | Reference doi |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1a | Forest management effects on tree and stand-water relationships | Quercus ilex L. | C + Mth | 2 plots, measures according to slope gradient | x | x | x | x | x | x | x | 10.1016/j.agrformet.2018.10.016 | |
| 1b | Forest management effects on tree and stand-water relationships | Pinus halepensis Mill. | C + Lth + Mth + Hth | 12 plots, 3 replicates per treatment | x | x | x | x | x | x | x | 10.1016/j.foreco.2019.02.020 | |
| 1c | Forest management effects on tree and stand-water relationships | Pinus halepensis | C + Mth | 2 plots, measures according to slope gradient | x | x | x | x | x | x | x | 10.1007/s10342-014-0805-7 | |
| 1d | Forest restoration in degraded land | Quercus sp., Pinus sp., Arbutus unedo L., Fraxinus ornus L., Juniperus phoenicea L. | Tree plantation | Different sp. combinations | x | x | x | 10.1016/j.foreco.2020.118156 | |||||
| 2a | Forest management effects on forest adaption to climate change | Pinus sylvestris L., Pinus nigra Arn. | C + Mth + Hth | 9 plots, 3 replicates per treatment | x | x | x | 10.1016/j.foreco.2018.11.006 | |||||
| 2b | Forest management effects on forest adaption to climate change | Pinus halepensis | C + Mth + Hth | 9 plots, 3 replicates per treatment | x | x | x | ||||||
| 2c | Forest decline implications on forest management | Quercus ilex | Unmanaged | 1 plot | x | x | x | x | x | x | |||
| 2d | Forest decline implications on forest management | Pinus halepensis | Unmanaged | 1 plot | x | x | x | x | 10.1016/j.foreco.2019.117564 | ||||
| 3a | Water balances and ecohydrological dynamics of iso- and aniso-hydrics in water-deficit conditions | Pinus halepensis | Unmanaged | 2 plots | x | x | x | x | 10.1016/j.scitotenv.2016.09.123 | ||||
| 3b | Ecohydrological dynamics of iso- and aniso-hydrics in water-deficit conditions | Pinus halepensis | Unmanaged | 2 plots | x | x | x | ||||||
| 3c | Water balances and ecohydrological dynamics | Pinus halepensis, Quercus ilex | Unmanaged | 3 plots | x | x | x | x | x | ||||
| 4a | Real evapotranspiration during rainfall in summer conditions | Pinus halepensis | Unmanaged | 3 plots | x | x | x | x | 10.3390/f9070426 | ||||
| 4b | Real evapotranspiration during rainfall in summer conditions | Pinus halepensis | Unmanaged | 3 plots | x | x | x | x | 10.1016/j.foreco.2015.12.020 | ||||
| 4c | Plant functional strategies, resistance to drought and water use efficiency in understory species | Pinus halepensis | C + Mth + Hth; Tree plantation under canopy | 3 plots | x | x | x | x | |||||
| 5a | Forest management effects on stand resilience | Pinus sylvestris | C + Mth | Watershed measurements at outlet and at samples points | x | x | x | x | 10.18172/cig.3432 | ||||
| 6a | Forest management effects on forest growth | Pinus halepensis | C + Mth + Hth | 3 plots | x | x | 10.1016/j.scitotenv.2020.138536 | ||||||
| 6b | Forest management effects on forest growth | Pinus halepensis | C + Mth + Hth | 3 plots | x | x | |||||||
| 7a | Run-off response and dynamics of groundwater at the watershed level | Pinus sylvestris, Fagus sylvatica L., Quercus faginea Lam. | Unmanaged | Watershed measurements at outlet | x | 10.1659/mrd.0876 | |||||||
| 7b | Run-off response, erosion and dynamics of groundwater at the watershed level | Pinus sylvestris, Quercus faginea | Unmanaged | Watershed measurements at outlet | x | 10.18172/cig.3475 | |||||||
| 7c | Run-off response, erosion and dynamics of groundwater | Pinus sylvestris, Pinus nigra | Unmanaged | Watershed measurements at outlet | x | 10.1002/hyp.10820 | |||||||
| 8a | Forest management effects on forest vitality and water resources | Quercus ilex, Arbutus unedo | C + Lth | Watershed measurements at outlet and at sample points | x | x | x | 10.1002/ece3.461 | |||||
| 8b | Forest management effects on forest vitality and water resources | Pinus halepensis | C + Lth | Watershed, measurements at outlet and at sample points | x | x | x | x | |||||
| 8c | Forest management and biodiversity | Pinus nigra | C + Lth | 2 plots | x | x | |||||||
| 8d | Forest growth and carbon sink | Pinus halepensis | C + Lth | 2 plots | x | x | |||||||
| 9a | Response of forest structure and understory diversity to thinning | Pinus halepensis | C + Lth + Mth | 9 plots, 3 replicates per treatment | x | x | x | x | x | 10.1111/j.1365-3040.2011.02300.x | |||
| 10a | Forest management effects on nutrients and biodiversity | Pinus sylvestris, Fagus sylvatica, Quercus ilex | C + Lth + Mth | 9 plots, 3 replicates per treat. | x | x | x | x | x | x | x | 10.1111/gcb.14672 | |
| 10b | Forest management effects on nutrients and biodiversity | Pinus sylvestris, Fagus sylvatica | C + Lth + Mth | 9 plots, 3 replicates per treatment | x | x | x | x | x | x | 10.1016/j.agrformet.2018.01.004 | ||
| 10c | Nutrients cycle in a secondary succession | Pinus sylvestris, Fagus sylvatica | Unmanaged | 1 plot | x | x | x | x | 10.1111/1365-2745.12813 | ||||
| 11a | Changes in the multifunctionality role of forest ecosystems of different ages | Pinus halepensis | Unmanaged | 2 plots | x | x | x | x | x | 10.1016/j.jenvman.2018.12.012 | |||
| 11b | Restoration of slopes after fires | Pinus halepensis | C + 3 post-fire prevention treats. | 12 plots, 3 replicates per treatment | x | x | x | ||||||
| 12a | Forest management effects on forest adaptation to climate change | Pinus pinea | C + Lth + Mth Hth | 12 plots, 3 replicates per treatment | x | x | x | ||||||
| 13a | Carbon and water cycles in a dehesa system | Quercus ilex | Soil vs. non-soil intervention | 9 plots, 3 soil treatments, 3 replicates per treatment | x | x | x | x | x | x | x | x | 10.1016/j.agrformet.2018.07.010 |
| 14a | Forest management effects on forest adaptation to climate change | Pinus halepensis | C + Lth + Mth Hth | 12 plots, 3 replicates per treatment | x | x | x | x | x | 10.1016/j.foreco.2019.03.061 10.1111/avsc.12151 | |||
| 14b | Soil management effects on reforestation of agricultural land | Pinus halepensis, Quercus ilex | Soil vs. non-soil intervention | 36 plots, 12 soil treatments, 3 replicates per treatment | x | x | x | x | 1016/j.foreco.2016.09.035 10.1016/j.foreco.2017.09.049 |
| Climatic Type | Subtype | Beech | Oakmix | Fir | Eupin | Oakmarc | Oak | Medpin | Jun | Sing | FirSp | River | Others |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| III (IV) | 0.4 | 0.1 | 0.2 | ||||||||||
| Arid | IV (III) | <0.1 | 1.7 | 83.9 | 0.4 | 1.5 | 11.2 | ||||||
| IV1 | 23.2 | 0.8 | 49.8 | 489.3 | 8.9 | 14.4 | 81.7 | ||||||
| IV2 | 5.3 | 374.0 | 205.2 | 3.1 | <0.1 | 8.9 | 372.4 | ||||||
| IV3 | 0.1 | 2.8 | 541.8 | 312.9 | 12.3 | 16.0 | 173.8 | ||||||
| IV4 | 1.1 | 3.4 | 71.0 | 2622.5 | 580.6 | 22.0 | 0.1 | 36.6 | 769.9 | ||||
| IV (VI)1 | 23.8 | 21.7 | 434.9 | 263.0 | 12.6 | 14.2 | 154.6 | ||||||
| Mediterranean | IV (VI)2 | 0.1 | 0.2 | 0.3 | 33.9 | 148.0 | 1.2 | 2.7 | 65.3 | ||||
| VI(IV)1 | 6.9 | 6.8 | 173.7 | 304.1 | 799.1 | 1168.6 | 221.5 | 0.5 | 54.6 | 887.6 | |||
| VI(IV)2 | 4.3 | 45.4 | 298.0 | 415.2 | 135.0 | 380.2 | 39.8 | 1.2 | 1.6 | 19.1 | 378.0 | ||
| VI (IV)3 | 7.3 | 0.6 | 0.3 | 48.2 | 1.4 | 45.3 | |||||||
| VI(IV)4 | 12.2 | 7.9 | 24.2 | 28.9 | 101.9 | 38.5 | 2.9 | 0.0 | 5.2 | 91.7 | |||
| VI (VII) | 25.3 | 58.2 | 3.8 | 534.8 | 119.6 | 139.7 | 215.7 | 26.9 | 1.9 | 10.6 | 331.9 | ||
| VI (V) | 120.1 | 369.0 | 67.8 | 45.6 | 53.8 | 242.7 | 1.3 | 13.9 | 0.0 | 27.9 | 789.8 | ||
| Nemoral | VI | 149.3 | 106.6 | 1.3 | 201.5 | 72.1 | 24.8 | 33.3 | 3.5 | 7.0 | 6.4 | 106.6 | |
| VIII (VI) | 75.8 | 64.8 | 1.4 | 414.8 | 82.2 | 15.0 | 11.9 | 11.6 | 12.3 | 4.9 | 91.7 | ||
| X (VIII) | 3.6 | 11.0 | 5.1 | 86.2 | 0.5 | 0.6 | 0.1 | 0.9 | 6.6 | 5.1 | 0.9 | 24.2 | |
| Oroboreal | X(IX)1 | 1.1 | 0.4 | 0.3 | 20.7 | 0.4 | 0.1 | 1.8 | 0.2 | 0.3 | 0.9 | ||
| Total (×1000 ha) | 398.8 | 678.6 | 12.0 | 1872.5 | 1171.0 | 5328.7 | 4222.5 | 370.8 | 43.7 | 7.2 | 225.3 | 4376.8 | |
| % of total | 2.1 | 3.6 | 0.1 | 10.0 | 6.3 | 28.5 | 22.6 | 2.0 | 0.2 | <0.1 | 1.2 | 23.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molina, A.J.; Navarro-Cerrillo, R.M.; Pérez-Romero, J.; Alejano, R.; Bellot, J.F.; Blanco, J.A.; Camarero, J.J.; Carrara, A.; Castillo, V.M.; Cervera, T.; et al. SilvAdapt.Net: A Site-Based Network of Adaptive Forest Management Related to Climate Change in Spain. Forests 2021, 12, 1807. https://doi.org/10.3390/f12121807
Molina AJ, Navarro-Cerrillo RM, Pérez-Romero J, Alejano R, Bellot JF, Blanco JA, Camarero JJ, Carrara A, Castillo VM, Cervera T, et al. SilvAdapt.Net: A Site-Based Network of Adaptive Forest Management Related to Climate Change in Spain. Forests. 2021; 12(12):1807. https://doi.org/10.3390/f12121807
Chicago/Turabian StyleMolina, Antonio J., Rafael M. Navarro-Cerrillo, Javier Pérez-Romero, Reyes Alejano, Juan F. Bellot, Juan A. Blanco, Jesús J. Camarero, Arnaud Carrara, Víctor M. Castillo, Teresa Cervera, and et al. 2021. "SilvAdapt.Net: A Site-Based Network of Adaptive Forest Management Related to Climate Change in Spain" Forests 12, no. 12: 1807. https://doi.org/10.3390/f12121807