
Environment Status Estimation of the Forest Communities Based on Floristic Surveys in the Mordovia State Nature Reserve, Russia


Abstract
:1. Introduction
2. Materials and Methods
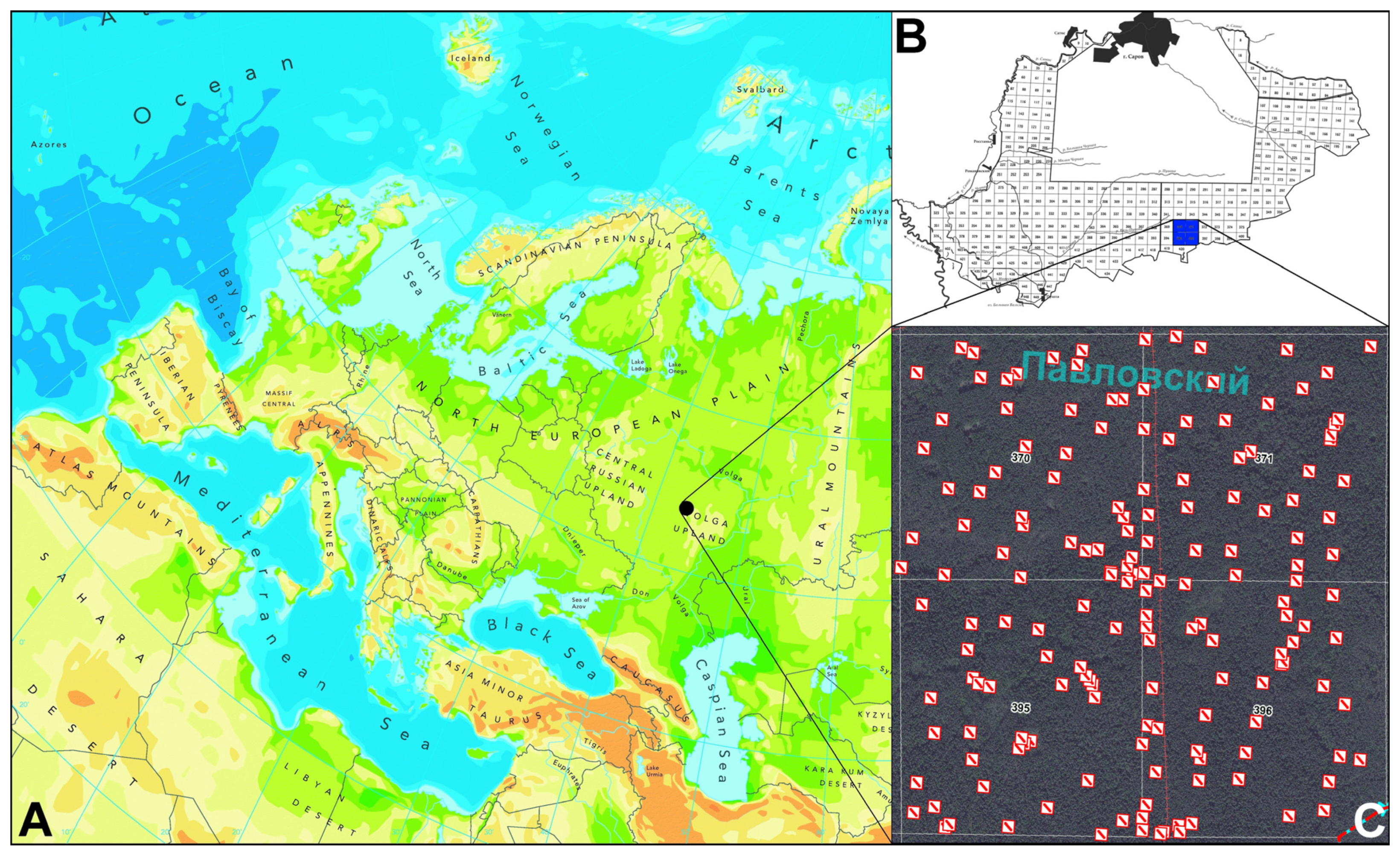
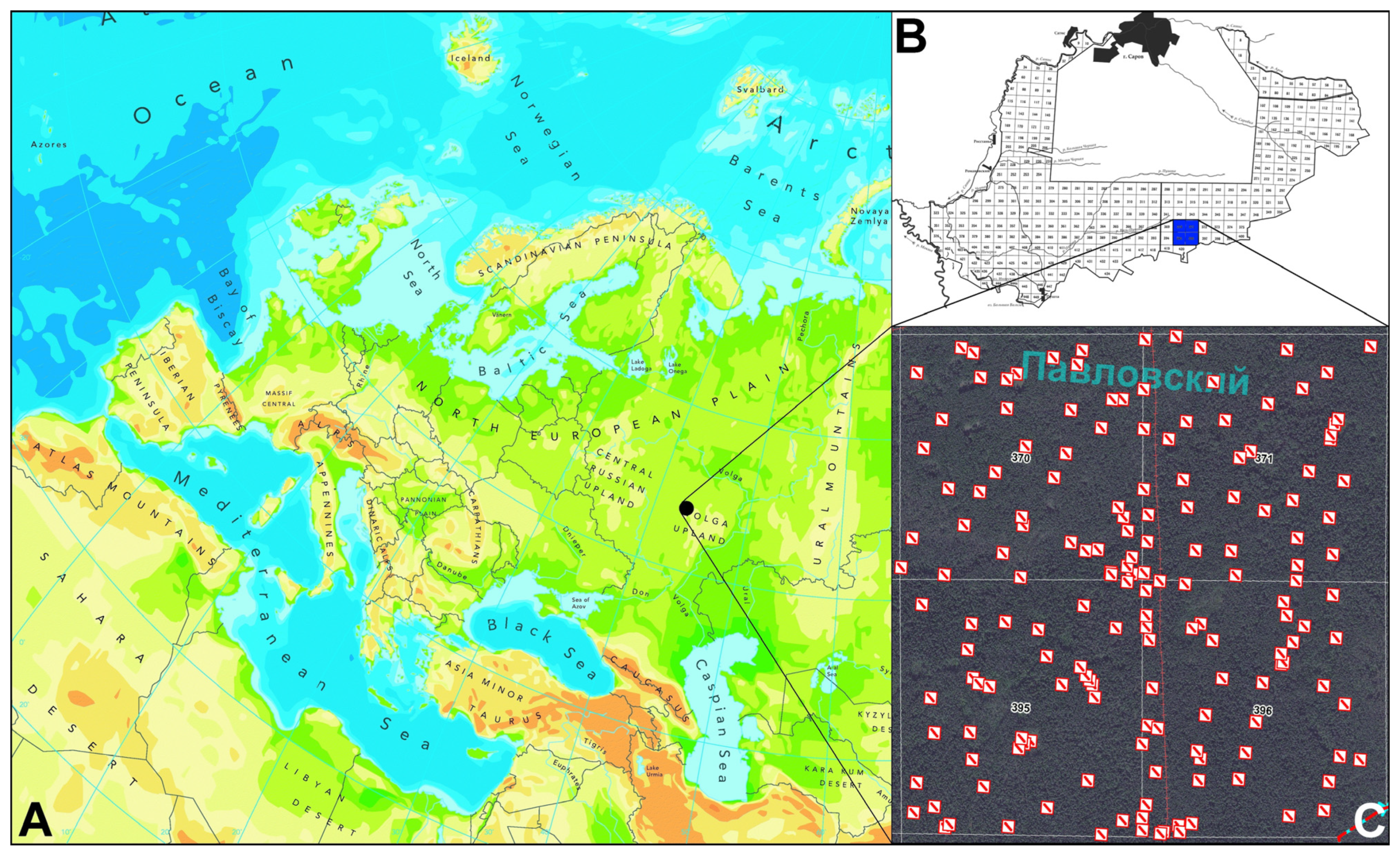
2.1. Study Area
2.2. Field Sampling
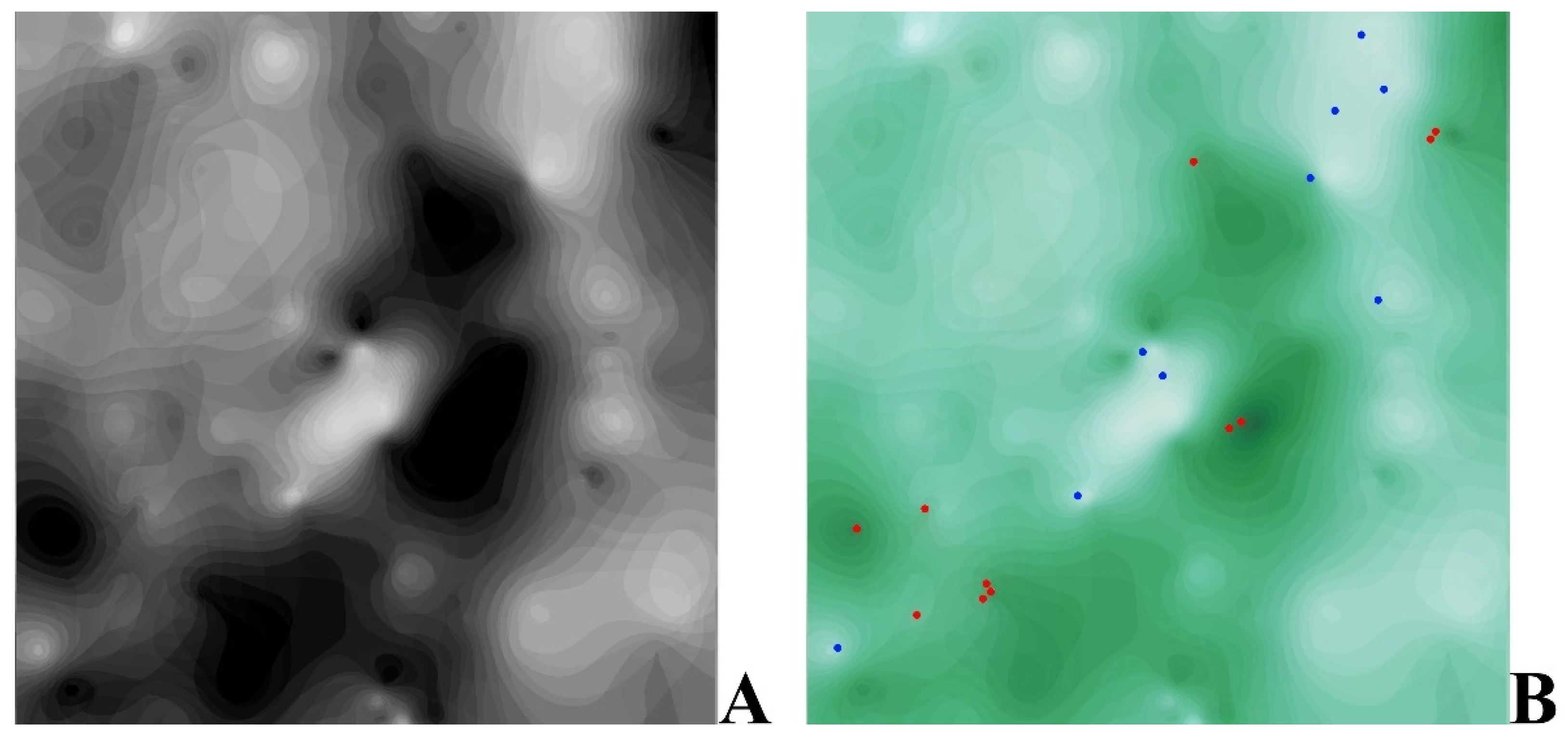
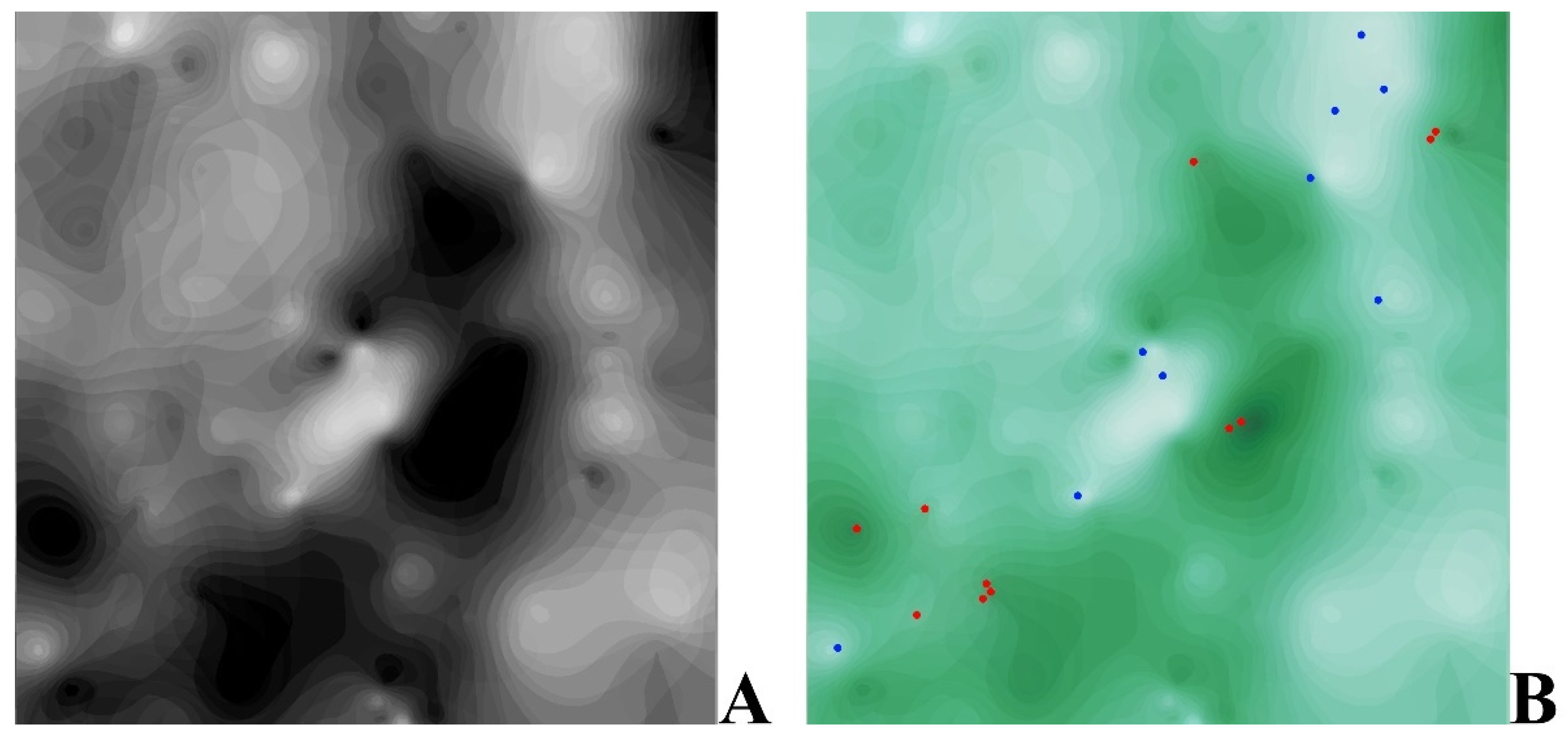
2.3. Data Analysis and Visualisation
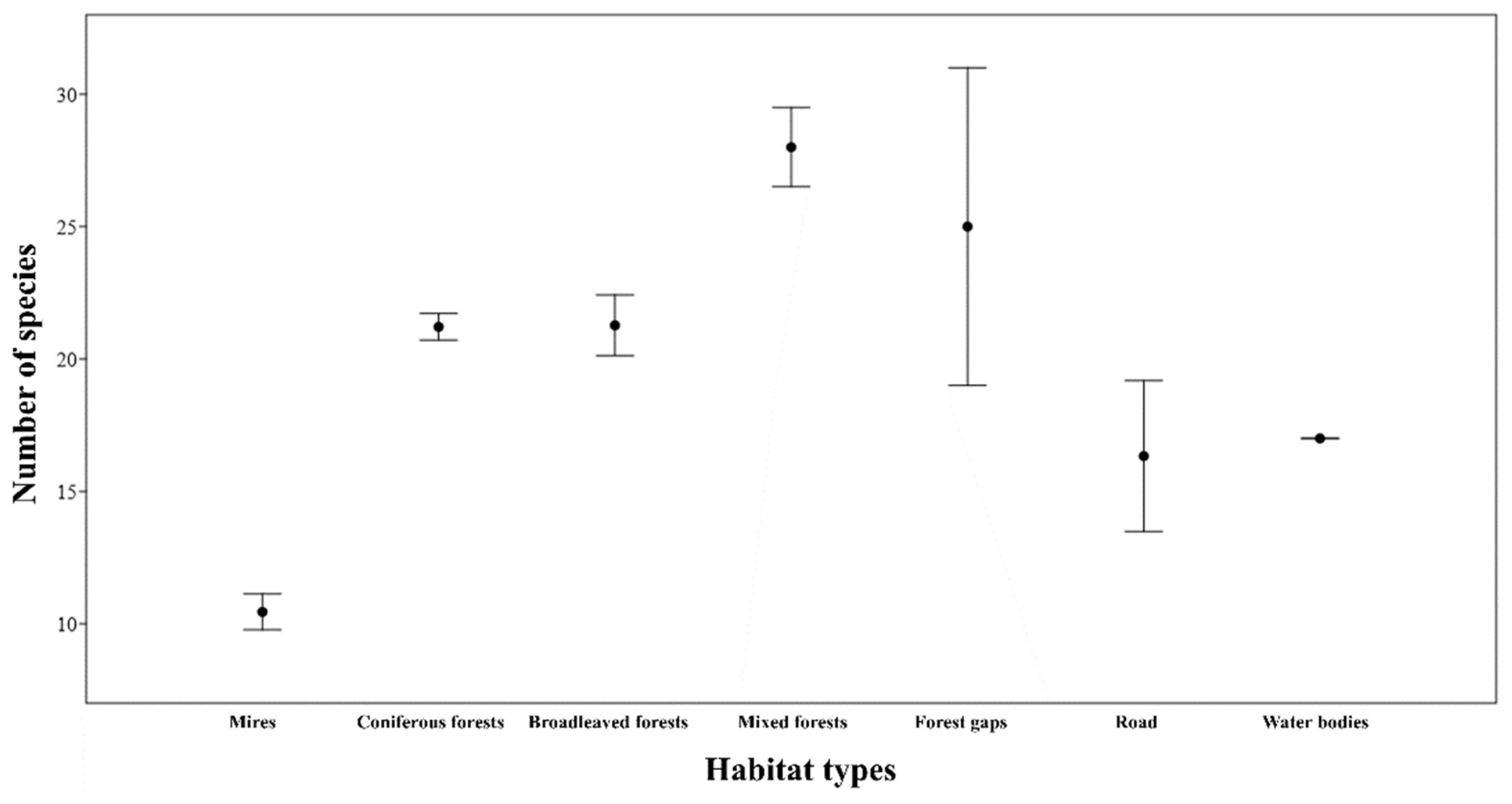
3. Results
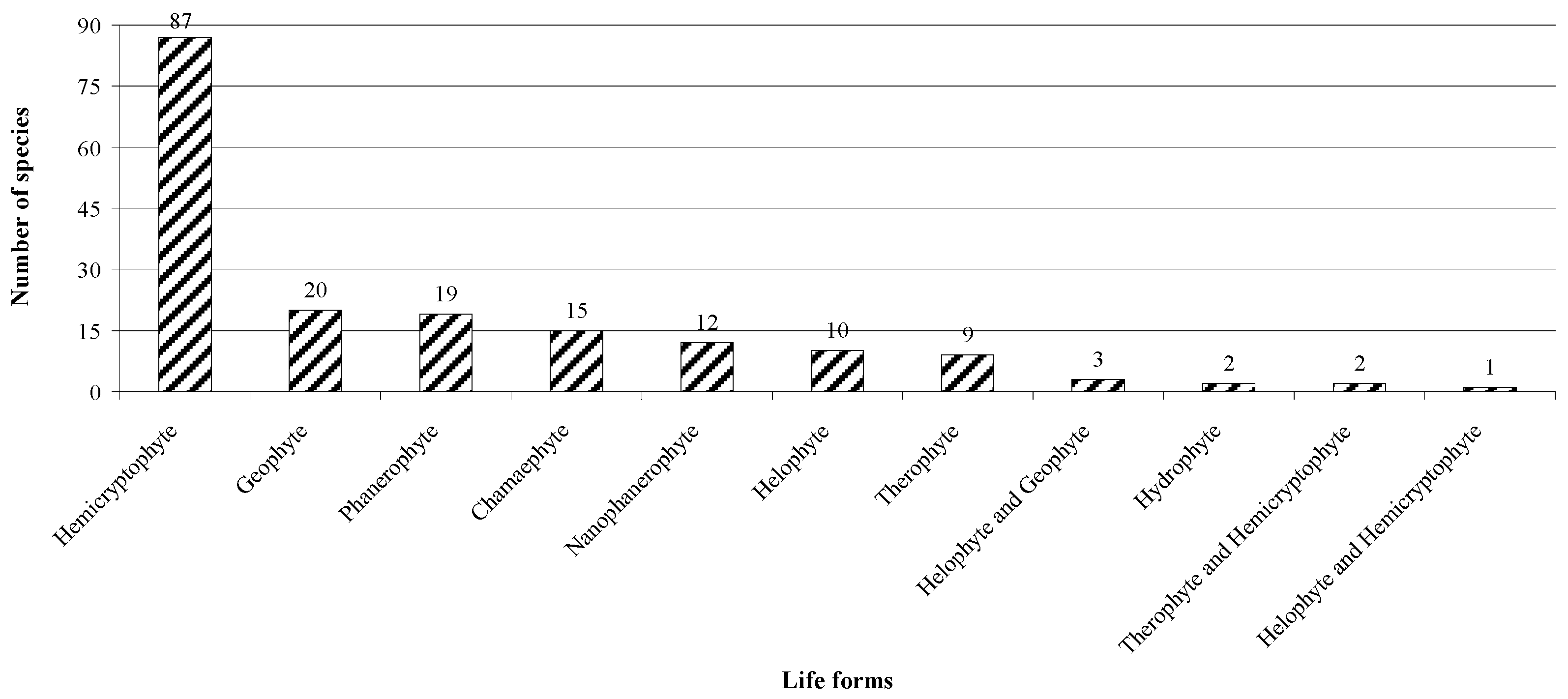
3.1. Vascular Plant Flora of the Study Area
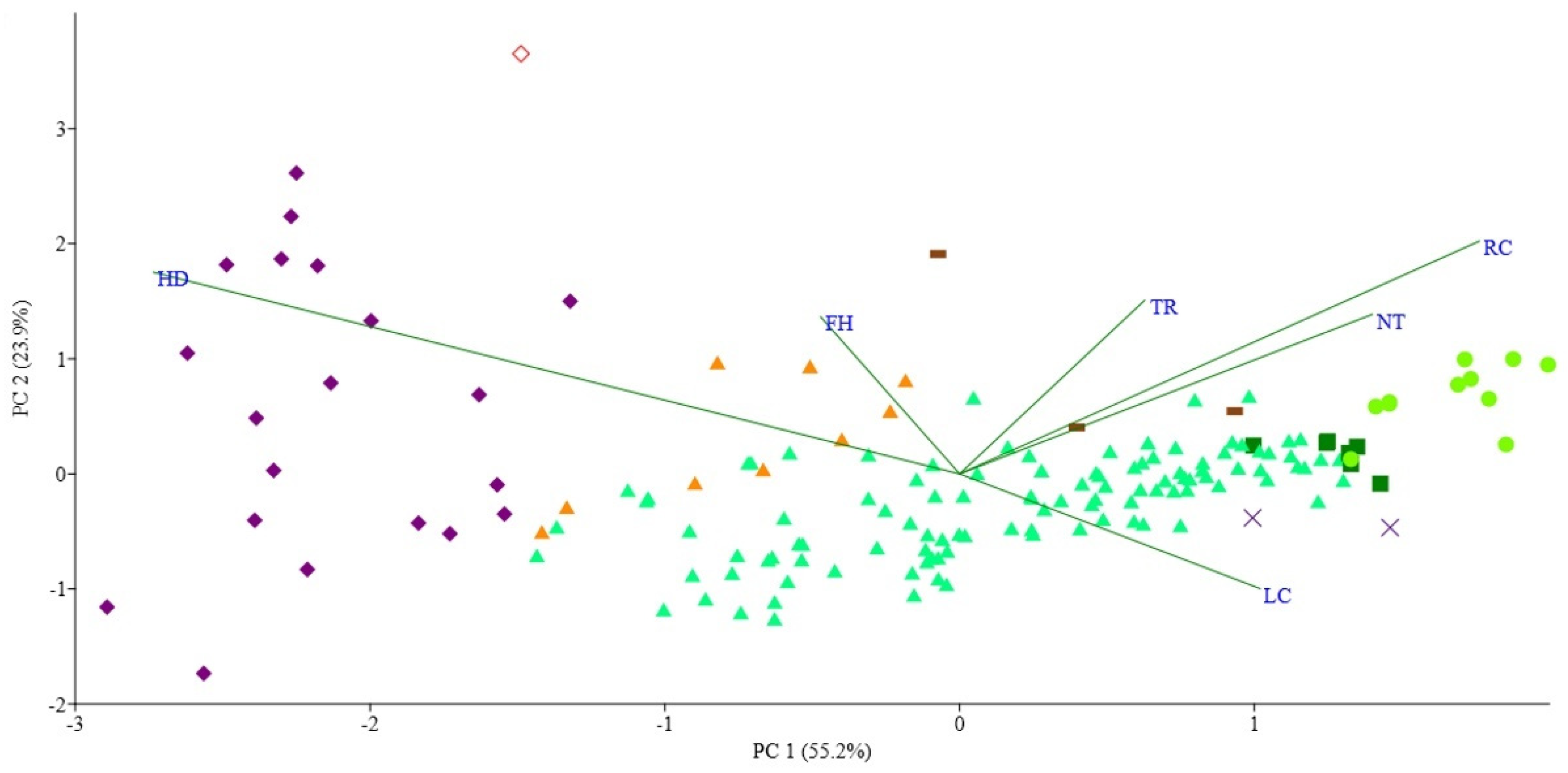
3.2. Plant-Environment Relationships Based on the Tsyganov Scale
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pottier, J.; Dubuis, A.; Pellissier, L.; Maiorano, L.; Rossier, L.; Randin, C.F.; Vittoz, P.; Guisan, A. Climate and species assembly predictions. Glob. Ecol. Biogeogr. 2013, 22, 52–63. [Google Scholar] [CrossRef]
- John, R.; Dalling, J.W.; Harms, K.E.; Yavitt, J.B.; Stallard, R.F.; Mirabello, M.; Hubbell, S.P.; Valencia, R.; Navarrete, H.; Vallejo, M.; et al. Soil nutrients influence spatial distributions of tropical tree species. Proc. Natl. Acad. Sci. USA 2007, 104, 864–869. [Google Scholar] [CrossRef] [Green Version]
- Khapugin, A.A.; Vargot, E.V.; Chugunov, G.G. Vegetation recovery in fire-damaged forests: A case study at the southern boundary of the taiga zone. For. Stud. 2016, 64, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Lennon, J.J.; Beale, C.M.; Reid, C.L.; Kent, M.; Pakeman, R.J. Are richness patterns of common and rare species equally well explained by environmental variables? Ecography 2011, 34, 529–539. [Google Scholar] [CrossRef]
- Popov, S.Y.; Makukha, Y.A. Distribution patterns of Ptilium crista-castrensis (Bryophyta, Hypnaceae) in the East European Plain and Eastern Fennoscandia. Nat. Conserv. Res. 2019, 4, 93–98. [Google Scholar] [CrossRef]
- Dubuis, A.; Rossier, L.; Pottier, J.; Pellissier, L.; Vittoz, P.; Guisan, A. Predicting current and future spatial community patterns of plant functional traits. Ecography 2013, 36, 1158–1168. [Google Scholar] [CrossRef]
- Gebremedihin, K.M.; Birhane, E.; Tadesse, T.; Gbrewahid, H. Restoration of degraded drylands through exclosures enhancing woody species diversity and soil nutrients in the highlands of Tigray, Northern Ethiopia. Nat. Conserv. Res. 2018, 3, 1–20. [Google Scholar] [CrossRef]
- Khan, W.; Khan, S.M.; Ahmad, H.; Ahmad, Z.; Page, S. Vegetation mapping and multivariate approach to indicator species of a forest ecosystem: A case study from the Thandiani sub Forests Division (TsFD) in the Western Himalayas. Ecol. Indic. 2016, 71, 336–351. [Google Scholar] [CrossRef]
- Pajunen, A.M.; Kaarlejärvi, E.M.; Forbes, B.C.; Virtanen, R. Compositional differentiation, vegetation-environment relationships and classification of willow-characterized vegetation in the western Eurasian Arctic. J. Veg. Sci. 2010, 21, 107–119. [Google Scholar] [CrossRef]
- Jarema, S.I.; Samson, J.; McGill, B.J.; Humphries, M.M. Variation in abundance across a species’ range predicts climate change responses in the range interior will exceed those at the edge: A case study with North American beaver. Glob. Chang. Biol. 2009, 15, 508–522. [Google Scholar] [CrossRef]
- Cui, B.S.; Zhai, H.J.; Dong, S.K.; Chen, B.; Liu, S.L. Multivariate analysis of the effects of edaphic and topographical factors on plant distribution in the Yilong lake basin of Yun-Gui Plateau, China. Can. J. Plant. Sci. 2009, 89, 209–219. [Google Scholar] [CrossRef]
- Potts, M.D.; Ashton, P.S.; Kaufman, L.S.; Plotkin, J.B. Habitat patterns in tropical rain forests: A comparison of 105 plots in northwest Borneo. Ecology 2002, 83, 2782–2797. [Google Scholar] [CrossRef]
- Seregin, A.P. Further east: Eutrophication as a major threat to the flora of Vladimir Oblast, Russia. Environ. Sci. Pollut. Res. 2014, 21, 12883–12897. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, O.V.; Lugovaya, D.V.; Prokazina, T.S. Model Reconstruction of Restored Taiga Forest Cover. Biol. Bull. Rev. 2013, 3, 493–504. [Google Scholar] [CrossRef]
- Ramenskiy, L.G.; Tsatsenkin, I.A.; Chijikov, O.N.; Antipin, N.A. Ecological Evaluation of Natural Grasslands by the Use of Vegetation Cover; Selkhozgiz: Moscow, Russia, 1956. (In Russian) [Google Scholar]
- Tsyganov, D.N. Phytoindication of Ecological Regimes in the Mixed Coniferous-Broad-Leaved Forest Subzone; Nauka: Moscow, Russia, 1983. (In Russian) [Google Scholar]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W. Zeigerwerte von Pflanzen in Mitteleuropa, 3, durch gesehene Aufl. Scr. Geobot. 2001, 18, 1–261. [Google Scholar]
- Landolt, E.; Bäumler, B.; Erhardt, A.; Hegg, O.; Klötzli, F.; Lämmler, W.; Nobis, M.; Rudmann-Maurer, K.; Schweingruber, F.H.; Theurillat, J.P.; et al. Ecological Indicator Values and Biological Attributes of the Flora of Switzerland and the Alps; Haupt: Bern, Switzerland, 2010. [Google Scholar]
- Didukh, Y.P. The Ecological Scales for the Species of Ukrainian Flora and Their Use in Synphytoindication; Phytosociocentre: Kyiv, Ukraine, 2011. [Google Scholar]
- Khapugin, A.A. Benefits from visualization of environmental factor gradients: A case study in a protected area in central Russia. Rev. Chapingo Ser. Cienc. For. Y Del Ambient. 2019, 25, 383–398. [Google Scholar] [CrossRef]
- Priputina, I.; Zubkova, E.; Shanin, V.; Smirnov, V.; Komarov, A. Evidence of plant biodiversity changes as a result of nitrogen deposition in permanent pine forest plots in central Russia. Ecoscience 2014, 21, 286–300. [Google Scholar] [CrossRef]
- Gusev, A.P. Specific features of early stages of progressive succession in an anthropogenic landscape: An example from southeastern Belarus. Russ. J. Ecol. 2009, 40, 160–165. [Google Scholar] [CrossRef]
- Khapugin, A.A. Hieracium sylvularum (Asteraceae) in the Mordovia State Nature Reserve: Invasive plant or historical heritage of the flora? Nat. Conserv. Res. 2017, 2, 40–52. [Google Scholar] [CrossRef] [Green Version]
- Kuznetsov, N.I. The conditions of existence and main features of the vegetation cover structure on the territory of Mordovia State Reserve. 1939 year. Proc. Mordovia State Nat. Reserve 2014, 12, 79–195. (In Russian) [Google Scholar]
- Bayanov, N.G. Climate changes of the northwest of Mordovia during the period of existence of the Mordovia Reserve according to the meteorological observations in Temnikov. Proc. Mordovia State Nat. Reserve 2015, 14, 212–219. [Google Scholar]
- Tereshkin, I.S.; Tereshkina, L.V. Vegetation of the Mordovia Reserve. Successive series of the successions. Proc. Mordovia State Nat. Reserve 2006, 7, 186–287. (In Russian) [Google Scholar]
- Raunkiaer, C. The Life Forms of Plant and Statistical Plant Geography; Clarendon Press: Oxford, UK, 1934. [Google Scholar]
- Marinšek, A.; Čarni, A.; Šilc, U.; Manthey, M. What makes a plant species specialist in mixed broad-leaved deciduous forests? Plant Ecol. 2015, 216, 1469–1479. [Google Scholar] [CrossRef]
- Golden Software Inc. Surfer Mapping System. v. 11.1.719 Software. USA, 2012. Available online: http://www.goldensoftware.com/products/surfer/ (accessed on 1 August 2021).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Ivanova, A.V.; Kostina, N.V.; Rozenberg, G.S.; Saksonov, S.V. Family ranges of the flora of the Volga River Basin territory. Bot. Zhurnal 2016, 101, 1042–1055. (In Russian) [Google Scholar]
- Khokhryakov, A.P. Taxonomic spectra and their role in comparative floristics. Bot. Zhurnal 2000, 85, 1–11. (In Russian) [Google Scholar]
- Talbot, S.S.; Talbot, S.L.; Schofield, W.B. Vascular flora of Izembek National Wildlife Refuge, Westernmost Alaska Peninsula, Alaska. Rhodora 2006, 108, 249–293. [Google Scholar] [CrossRef]
- Demina, G.V.; Zakirov, A.R. Floristic characterization of the territory adjacent to the “Gray Heron Colony” protected area. EurAsian J. BioSci. 2018, 12, 227–231. [Google Scholar]
- Ingerpuu, N.; Vellak, K.; Kukk, T.; Pärtel, M. Bryophyte and vascular plant species richness in boreo-nemoral moist forests and mires. Biodivers. Conserv. 2001, 10, 2153–2166. [Google Scholar] [CrossRef]
- Hájek, M.; Hájková, P.; Apostolova, I.; Horsák, M.; Plášek, V.; Shaw, B.; Lazarova, M. Disjunct Occurrences of Plant Species in the Refugial Mires of Bulgaria. Folia Geobot. 2009, 44, 365–386. [Google Scholar] [CrossRef]
- Khapugin, A.A.; Ruchin, A.B. Red Data Book vascular plants in the Mordovia State Nature Reserve, a Protected Area in European Russia. Wulfenia 2019, 26, 53–71. [Google Scholar]
- Mazei, Y.A.; Tsyganov, A.N.; Bobrovsky, M.V.; Mazei, N.G.; Kupriyanov, D.A.; Gałka, M.; Rostanets, D.V.; Khazanova, K.P.; Stoiko, T.G.; Pastukhova, Y.A.; et al. Peatland Development, Vegetation History, Climate Change and Human Activity in the Valdai Uplands (Central European Russia) during the Holocene: A Multi-Proxy Palaeoecological Study. Diversity 2020, 12, 462. [Google Scholar] [CrossRef]
- Jobidon, R.; Cyr, G.; Thiffault, N. Plant species diversity and composition along an experimental gradient of northern hardwood abundance in Picea mariana plantations. For. Ecol. Manag. 2004, 198, 209–221. [Google Scholar] [CrossRef]
- Barbier, S.; Gosselin, F.; Balandier, P. Influence of tree species on understory vegetation diversity and mechanisms involved—A critical review for temperate and boreal forests. For. Ecol. Manag. 2008, 254, 1–15. [Google Scholar] [CrossRef]
- Hammond, M.E.; Pokorny, R.; Dobrovolný, L.; Friedl, M.; Hiitola, N. Effect of gap size on tree species diversity of natural regeneration–case study from Masaryk training forest school enterprise Křtiny. J. For. Sci. 2020, 66, 1–14. [Google Scholar] [CrossRef]
- Hackshaw, A. Small studies: Strengths and limitations. Eur. Respir. J. 2008, 32, 1141–1143. [Google Scholar] [CrossRef]
- Ajay, S.; Micah, B. Sampling techniques and determination of sample size in applied statistics research: An overview. Int. J. Econ. Commer. Manag. 2014, 2, 1–22. [Google Scholar]
- Vahdati, F.B.; Mehrvarz, S.S.; Dey, D.C.; Naqinezhad, A. Environmental factors–ecological species group relationships in the Surash lowland-mountain forests in northern Iran. Nord. J. Bot. 2017, 35, 240–250. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat | Number of Studied Plots | Proportion, % |
|---|---|---|
| Coniferous forests | 117 | 72.7 |
| Mires | 20 | 12.4 |
| Broadleaved forests | 11 | 6.8 |
| Mixed forests | 7 | 4.3 |
| Roads | 3 | 1.9 |
| Forest gaps | 2 | 1.2 |
| Water bodies | 1 | 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khapugin, A.A. Environment Status Estimation of the Forest Communities Based on Floristic Surveys in the Mordovia State Nature Reserve, Russia. Forests 2021, 12, 1475. https://doi.org/10.3390/f12111475
Khapugin AA. Environment Status Estimation of the Forest Communities Based on Floristic Surveys in the Mordovia State Nature Reserve, Russia. Forests. 2021; 12(11):1475. https://doi.org/10.3390/f12111475
Chicago/Turabian StyleKhapugin, Anatoliy A. 2021. "Environment Status Estimation of the Forest Communities Based on Floristic Surveys in the Mordovia State Nature Reserve, Russia" Forests 12, no. 11: 1475. https://doi.org/10.3390/f12111475
APA StyleKhapugin, A. A. (2021). Environment Status Estimation of the Forest Communities Based on Floristic Surveys in the Mordovia State Nature Reserve, Russia. Forests, 12(11), 1475. https://doi.org/10.3390/f12111475