
Enzymatic Activity of Soils Forming on an Afforested Heap from an Opencast Sulphur Mine




Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling for Analyses
2.2. Laboratory Analyses
2.2.1. Total Organic Carbon (TOC), Total Nitrogen (Nt), C/N Ratio
2.2.2. Soil Enzyme Activity
2.3. Statistical Analyses
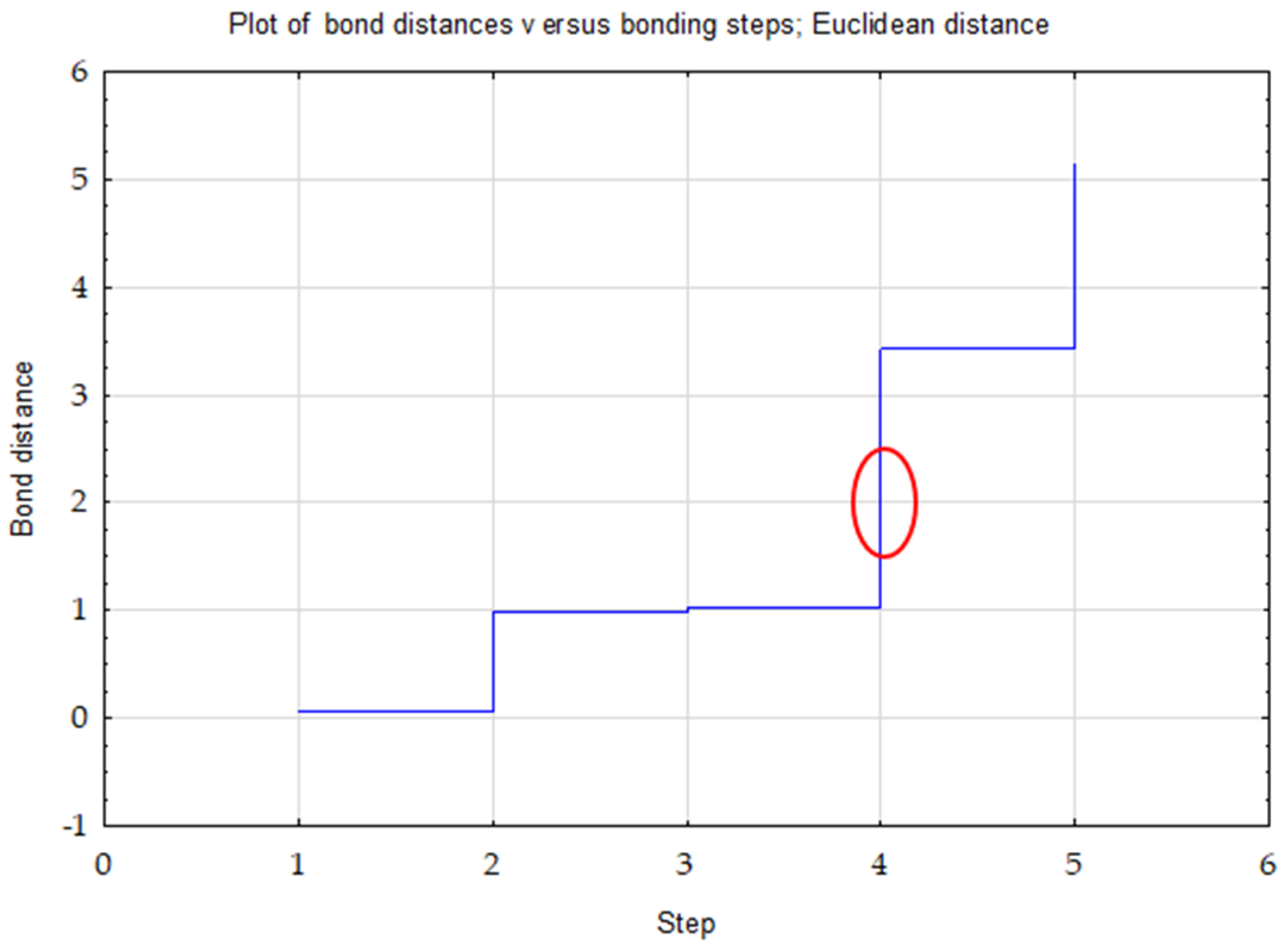
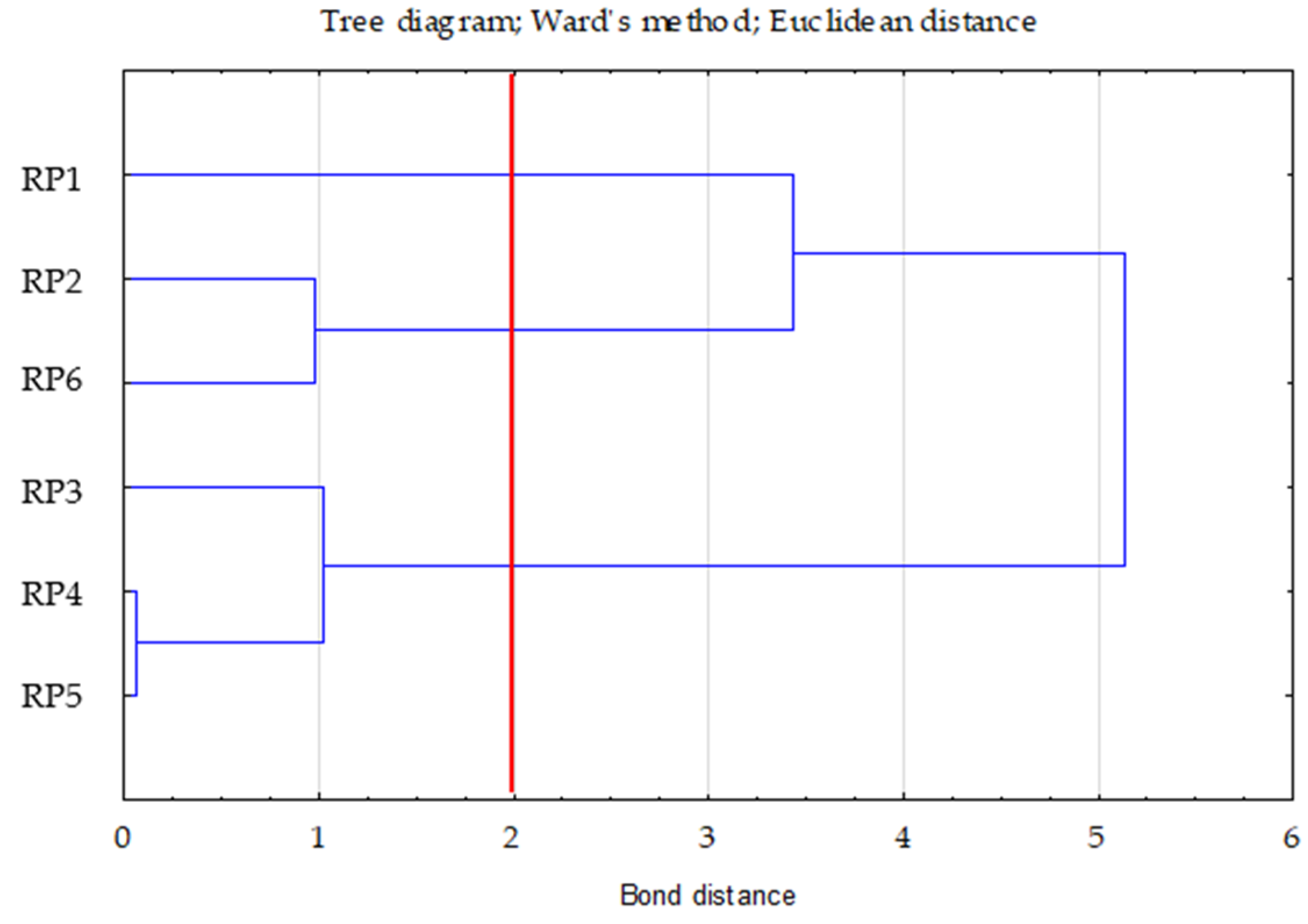
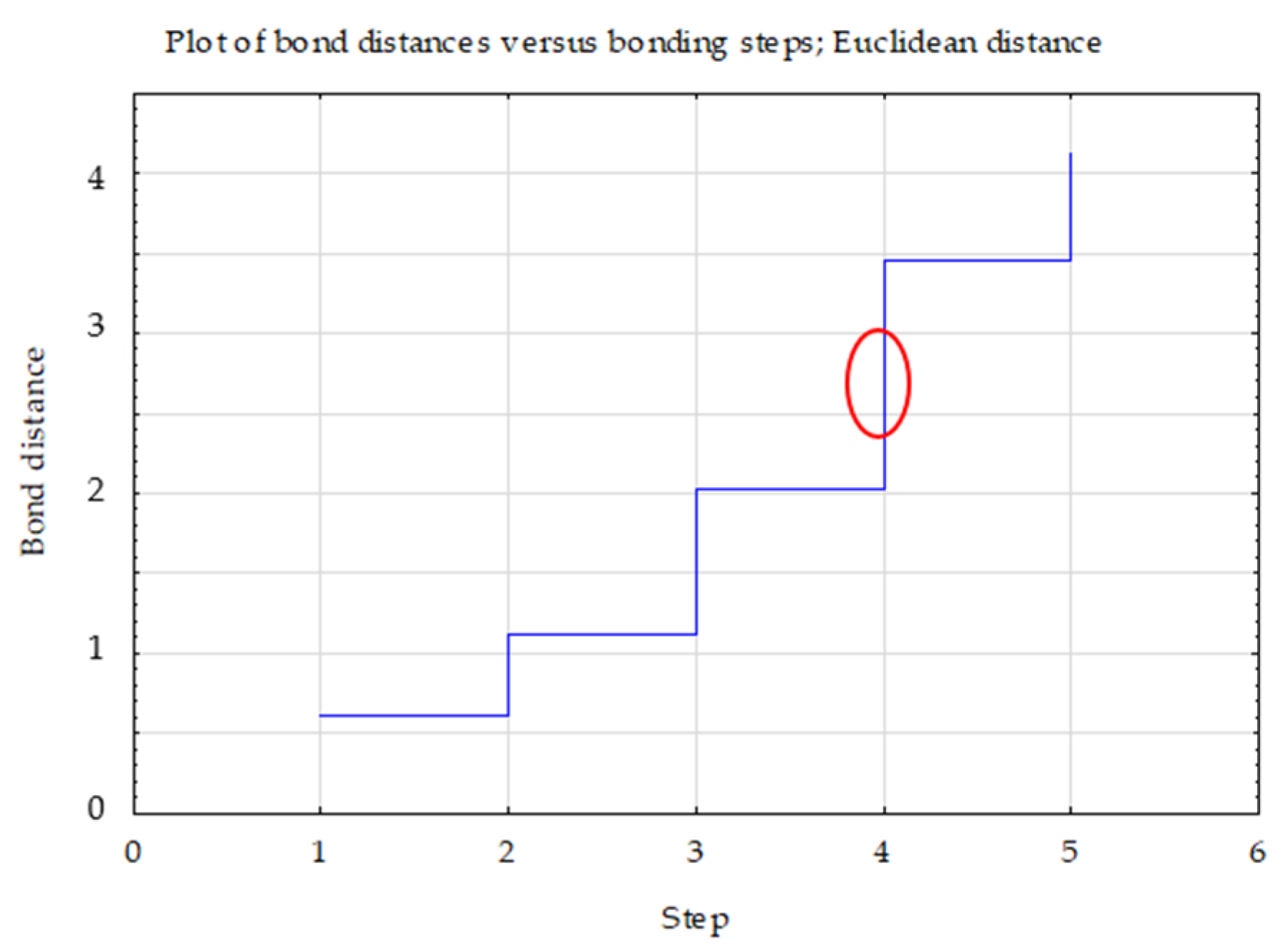
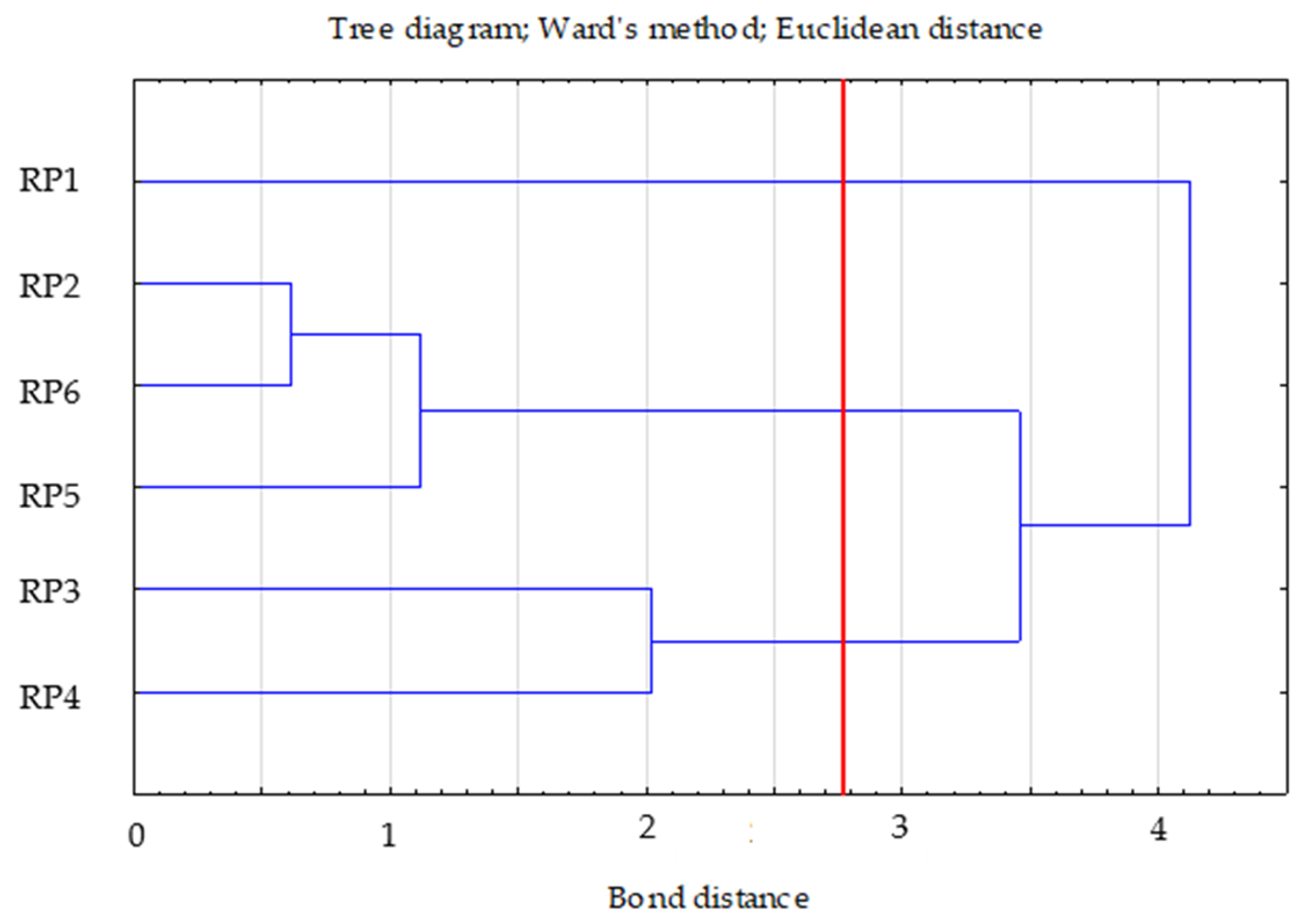
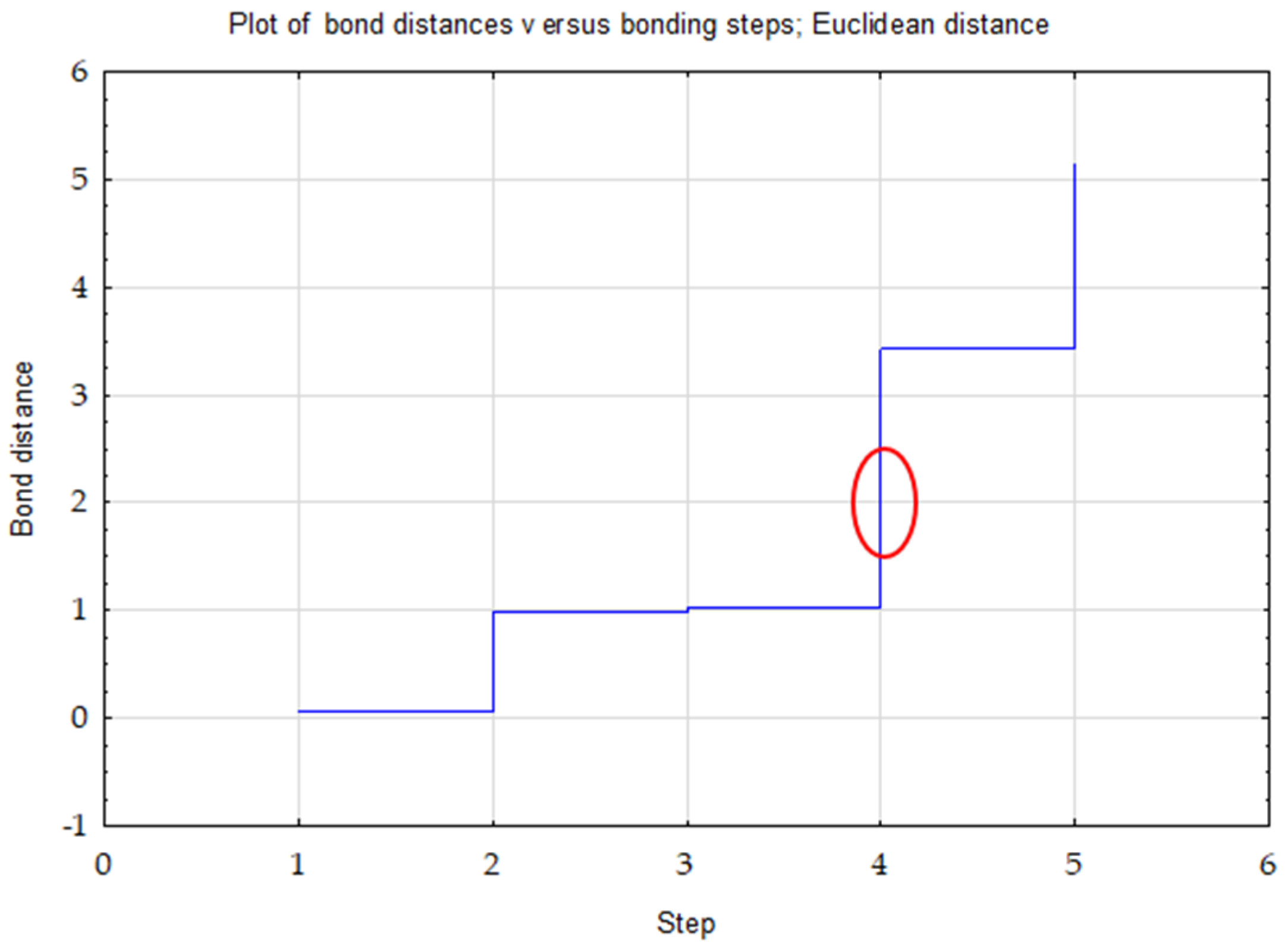
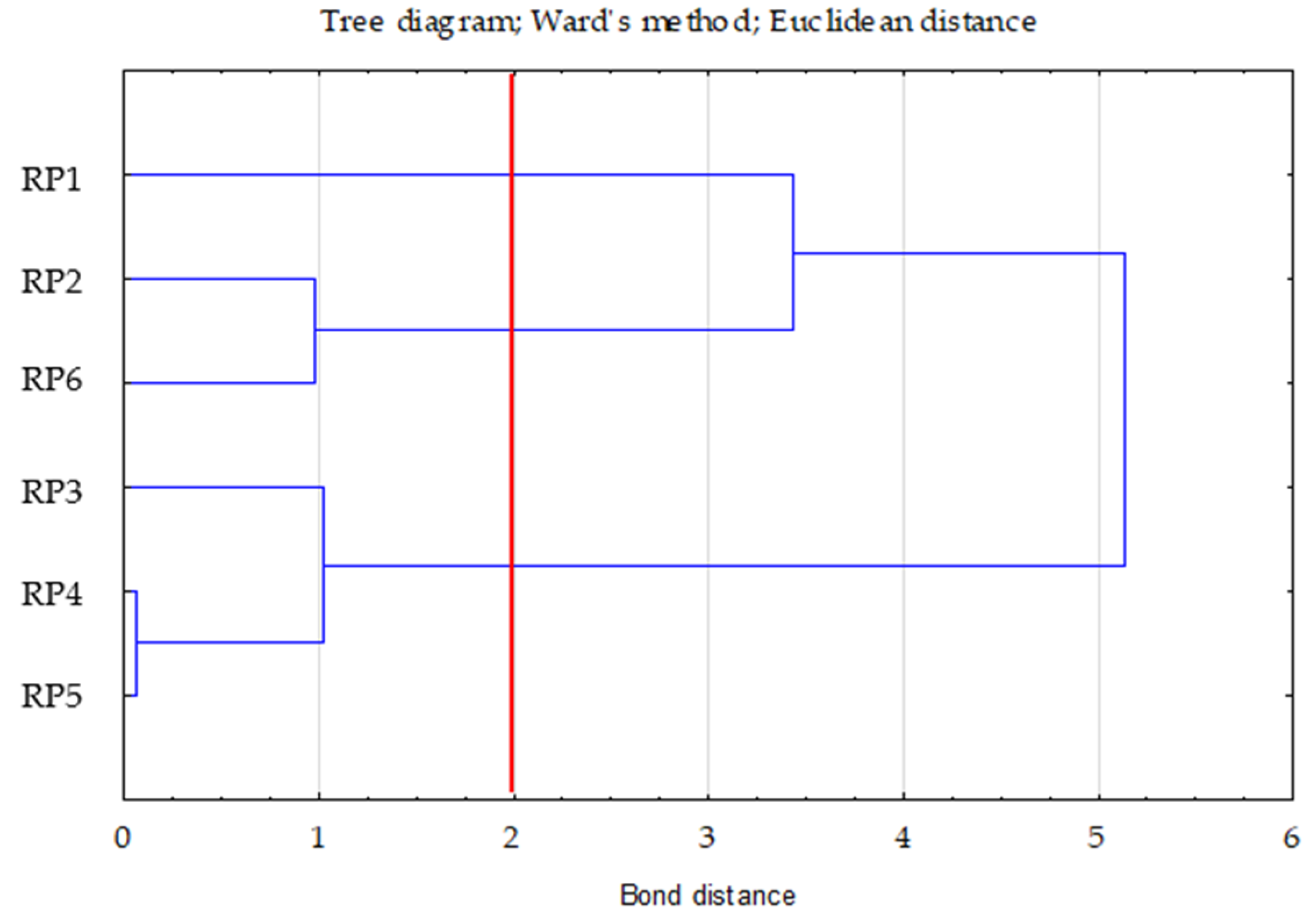
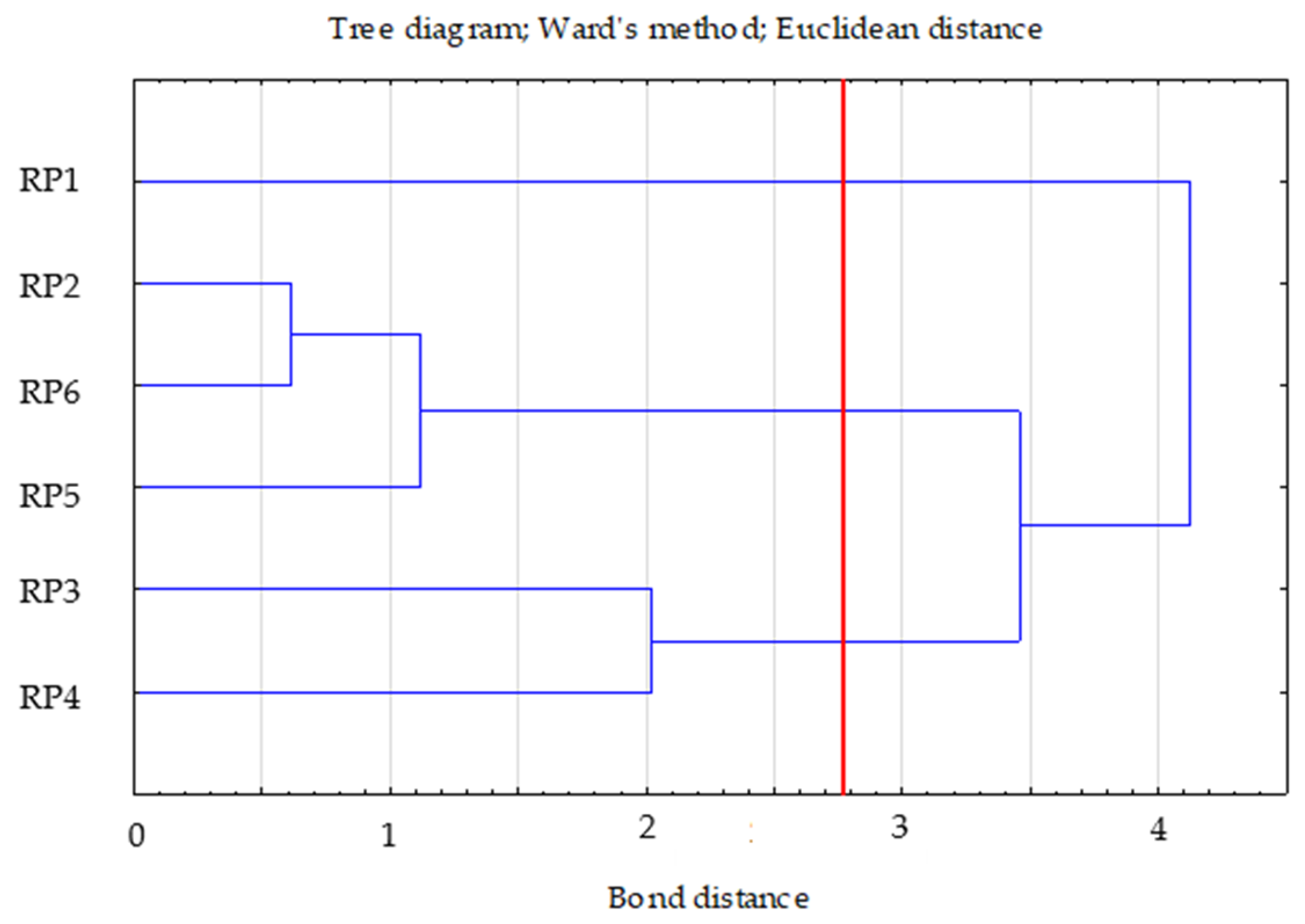
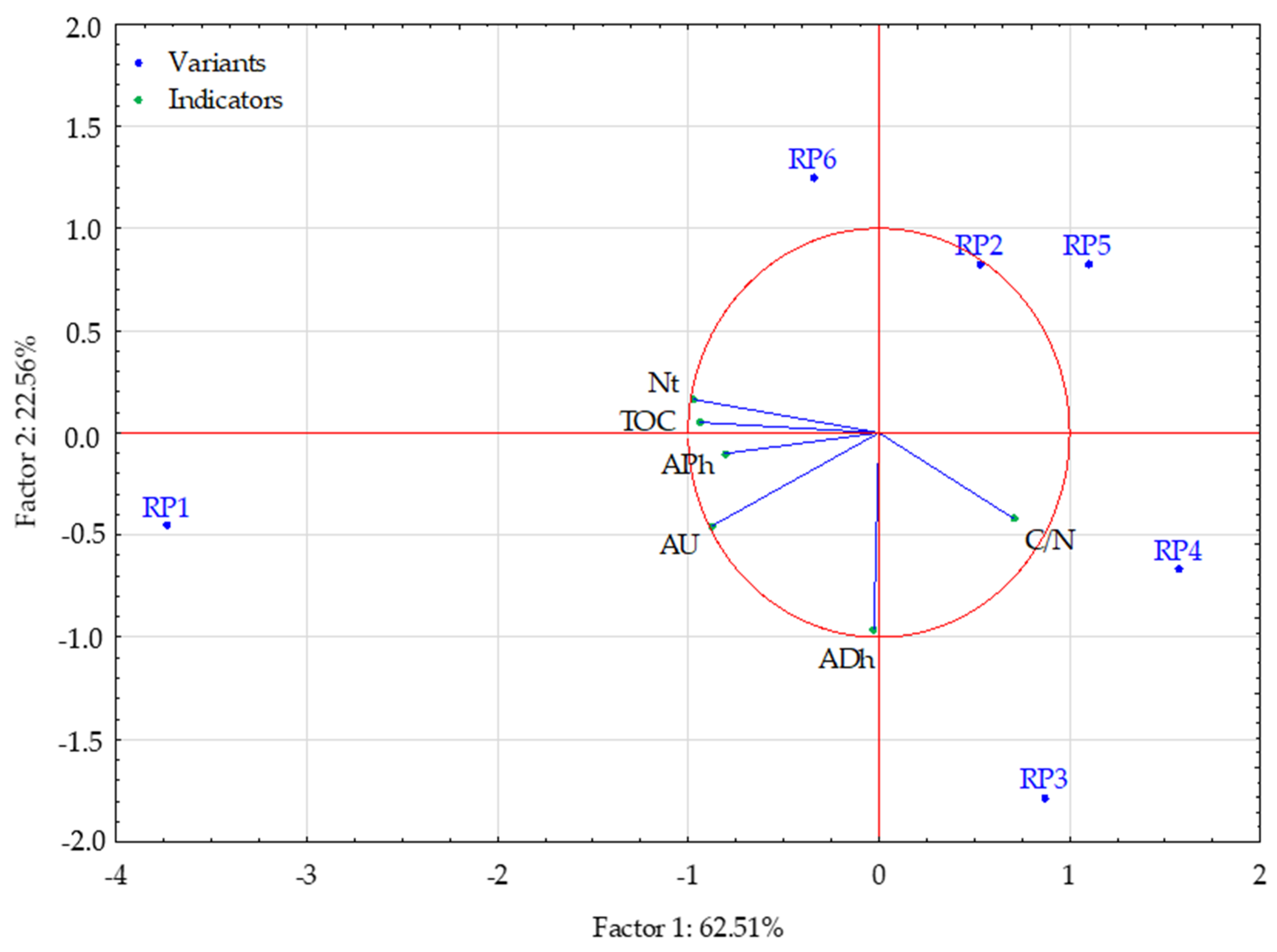
- Ward’s method—to determine the number of clusters (distance measure—Euclidean) based on visual analysis of the graph of distance changes in successive agglomeration stages (significant increase in agglomeration distance on the graph) and the connection tree (Ward’s dendrogram);
- The k-means method—to determine the composition of clusters. The analysis of variance was also performed to determine the significance of cluster differentiation.
- Internal homogeneity (homogeneity) of the clusters;
- The extent to which the obtained clusters differed from each other;
- Which sub-variables determined the differentiation of the internal structure of the reclamation variants that form the clusters.
3. Results
Enzymatic Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hüttl, R.F.; Weber, E. Forest ecosystem development in post-mining landscapes: A case study of the Lusatian lignite district. Naturwissenschaften 2001, 88, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Sheoran, V.; Sheoran, A.S.; Poonia, P. Soil reclamation of abandoned mine land by revegetation. Int. J. Soil Sediment Water 2010, 3, 13. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, S.E.; Landhausser, S.M.; Skousen, J.; Franklin, J.; Frouz, J.; Hall, S.; Jacobs, D.; Quideau, S. Forest restoration following surface mining disturbance: Challenges and solutions. New Forest 2015, 46, 703–732. [Google Scholar] [CrossRef] [Green Version]
- Pietrzykowski, M. Tree species selection and reaction to mine soil reconstructed at reforested post-mine sites: Central and eastern European experiences. Ecol. Eng. 2019, 3, 100012. [Google Scholar] [CrossRef]
- Lal, R. Restoring soil quality to mitigate soil degradation. Sustainability 2015, 7, 5875–5895. [Google Scholar] [CrossRef] [Green Version]
- Gomiero, T. Soil degradation, land scarcity and food security: Reviewing a complex challenge. Sustainability 2016, 8, 281. [Google Scholar] [CrossRef] [Green Version]
- DeLong, C.; Cruse, R.; Wieneret, J. The Soil Degradation Paradox: Compromising Our Resources When We Need Them the Most. Sustainability 2015, 7, 866–879. [Google Scholar] [CrossRef] [Green Version]
- FAO. Status of the World’s Soil Resources. 2015. Available online: http://www.fao.org/3/i5228e/i5228e.pdf (accessed on 19 September 2021).
- Northey, S.; Haque, N.; Mudd, G. Using sustainability reporting to assess the environmental footprint of copper mining. J. Clean. Prod. 2013, 40, 118–128. [Google Scholar] [CrossRef]
- Miao, Z.; Marrs, R. Ecological restoration and land reclamation in open-cast mines in Shanxi Province, China. J. Environ. Manag. 2000, 59, 205–215. [Google Scholar] [CrossRef]
- Lin, C.; Tong, X.; Lu, W.; Yan, L.; Wu, Y.; Nie, C.; Chu, C.; Long, J. Environmental impacts of surface mining on mined lands, affected streams and agricultural lands in the Dabaoshan Mine region, southern China. Land Degrad. Dev. 2005, 16, 463–474. [Google Scholar] [CrossRef]
- Cooke, J.A.; Johnson, M.S. Ecological restoration of land with particular reference to the mining of metals and industrial minerals. Environ. Rev. 2002, 10, 41–71. [Google Scholar] [CrossRef] [Green Version]
- Mensah, A.K. Role of revegetation in restoring fertility of degraded mined soils in Ghana: A review. Int. J. Biodivers. Conserv. 2015, 7, 57–80. [Google Scholar] [CrossRef] [Green Version]
- Vela-Almeida, D.; Brooks, G.; Kosoy, N. Setting the limits to extraction: A biophysical approach to mining activities. Ecol. Econ. 2015, 119, 189–196. [Google Scholar] [CrossRef]
- Franks, D.M.; Boger, D.V.; Côte, C.M.; Mulligan, D.R. Sustainable development principles for the disposal of mining and mineral processing wastes. Resour. Policy 2011, 36, 114–122. [Google Scholar] [CrossRef]
- Feng, J.; Wang, Z.; Bai, L. Reading Effects of surface coal mining and land reclamation on soil properties: A review. Earth Sci. Rev. 2019, 191, 12–25. [Google Scholar] [CrossRef]
- Wang, J.; Guo, L.; Bai, Z.; Yang, L. Using computed tomography (CT) images and multi-fractal theory to quantify the pore distribution of reconstructed soils during ecological restoration in opencast coal-mine. Ecol. Eng. 2016, 92, 148–157. [Google Scholar] [CrossRef]
- Society for Ecological Restoration Science the SER Premieron Ecological Restoration. 2002. Available online: https://www.ser.org/ (accessed on 19 September 2021).
- Seabrook, L.; McAlpine, C.A.; Bowen, M.E. Restore, repair or reinvent: Options for sustainable landscapes in a changingclimate. Landsc. Urban Plan. 2011, 100, 407–410. [Google Scholar] [CrossRef]
- Lima, A.T.; Mitchell, K.; O’Connell, D.W.; Verhoeven, J.; Van Cappellen, P. The legacy of surface mining: Remediation, restoration, reclamation and rehabilitation. Environ. Sci. Policy 2016, 66, 227–233. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Higgs, E.; Harris, J.A. Novel ecosystems: Implicationsfor conservation and restoration. Trends Ecol. Evol. 2009, 24, 599–605. [Google Scholar] [CrossRef]
- Frouz, J.; Livečková, M.; Albrechtová, J.; Chroňáková, A.; Cajthamlae, T.; Pižl, V.; Háněl, L.; Starý, J.; Baldrian, P.; Lhotáková, Z.; et al. Is the effect of trees on soil properties mediated by soil fauna? A case study from post-mining sites. Forest Ecol. Manag. 2013, 309, 87–95. [Google Scholar] [CrossRef]
- Hüttl, R.F.; Bradshaw, A. Aspect of reclamation ecology. Landsc. Urban Plan. 2000, 51, 73–74. [Google Scholar] [CrossRef]
- Pietrzykowski, M.; Gruba, P.; Sproul, G. The effectiveness of Yellow lupine (Lupinusluteus L.) green manure cropping in sand mine cast reclamation. Ecol. Eng. 2017, 2, 72–79. [Google Scholar] [CrossRef]
- Baumann, K.; Rumpelt, A.; Schneider, B.U.; Marschner, P.; Hüttl, R.F. Seedling biomass and element content of Pinus sylvestris and Pinus nigra grown in sandy substrates with lignite. Geoderma 2006, 136, 573–578. [Google Scholar] [CrossRef]
- Chodak, M.; Niklińska, M. The effect of different tree species on the chemical and microbial properties of reclaimed mine soils. Biol. Fertil. Soils 2010, 46, 555–566. [Google Scholar] [CrossRef]
- Frouz, J.; Voborilova, V.; Janusova, I.; Kodochova, S. Spontaneous establishement of late successional tree species English oak (Quercusrobur) and European beech (Fagus sylvatica) at reclaimed alder plantation and unreclaimed post mining sites. Ecol. Eng. 2015, 77, 1–8. [Google Scholar] [CrossRef]
- Pietrzykowski, M.; Socha, J.; van Doorn, N.S. Scots pine (Pinus sylvestris L.) site index in relation to physico-chemical and biological properties in reclaimed mine soils. New Forest 2014, 46, 247–266. [Google Scholar] [CrossRef] [Green Version]
- Cline, L.C.; Zak, D.R. Soil microbial communities are shaped by plant-driven changes in resource availability during secondary succession. Ecology 2015, 96, 3374–3385. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Ogdahl, M.; Chorover, J.; Chadwick, O.A.; Oleksyn, J.; Zytkowiak, R.; Reich, P.B. Tree Species Effects on Soil Organic Matter Dynamics: The Role of Soil Cation Composition. Ecosystems 2007, 10, 999–1018. [Google Scholar] [CrossRef]
- Doran, J.W.; Parkin, T.B. Quantitative indicators of soil quality: A minimum data set. In Methods for Assessing Soil Quality. Soil Science Society of America; Doran, J.W., Jones, A.J., Eds.; Special Publication 49; Soil Science Society of America: Madison, WI, USA, 1996; pp. 25–37. [Google Scholar] [CrossRef]
- Wolters, V.; Schaefer, M. Effects of acid deposition on soil organisms and decomposition processes. In Effects of Acid Rain on Forest Processes; Benckiser, G., Ed.; Wiley: New York, NY, USA, 1994; pp. 83–127. [Google Scholar]
- Schloter, M.; Dilly, O.; Munch, J.C. Indicators for evaluating soil quality. Agric. Ecosyst. Environ. 2003, 98, 255–262. [Google Scholar] [CrossRef]
- Bielińska, E.J.; Futa, B.; Mocek-Płóciniak, A. Soil Enzymes as Bioindicators of Soil Quality and Health; Scientific Monograph; Society of Scientific Publishers “LIBROPOLIS”: Lublin, Poland, 2014. [Google Scholar]
- De la Paz Jimenez, M.; de la Horra, A.M.; Pruzzo, L.; Palma, R.M. Soil quality: A new index based on microbiological and biochemical parameters. Biol. Fertil. Soils 2002, 35, 302–306. [Google Scholar] [CrossRef]
- Bastida, F.; Zsolnay, A.; Hernamdez, T.; Garcia, C. Past, present and future of soil quality indices: A biological perspective. Geoderma 2008, 147, 159–171. [Google Scholar] [CrossRef]
- Trasar-Cepada, C.; Leirós, M.C.; Seoane, S.; Gil-Sotres, F. Limitations of soil enzymes as indicators of soil pollution. Soil Biol. Biochem. 2000, 32, 867–875. [Google Scholar] [CrossRef]
- Schimel, J.P.; Bennett, J. Nitrogen mineraliztion: Challenges of a changing paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Gelsomino, A.; Badalucco, L.; Landi, L.C.G. Soil carbon, nitrogen and phosphorus dynamics as affected by solarization alone or combined with organic amendment. Plant Soil 2006, 279, 307–325. [Google Scholar] [CrossRef]
- Topac, F.O.; Dindar, E.; Ucaroglu, S.; Baskaya, H.S. Effect of a sulfonated azo dye and sulfanilic acid on nitrogen transformation processes in soil. J. Hazard. Mater. 2009, 170, 1006–1013. [Google Scholar] [CrossRef]
- Fernández, D.A.; Roldán, A.; Azcón, R.; Caravaca, F.; Bååth, E. Effects of water stress, organic amendment and mycorrhizal inoculation on soil microbial community structure and activity during the establishment of two heavy metal-tolerant native plant species. Microb. Ecol. 2012, 63, 794–803. [Google Scholar] [CrossRef]
- Kaczyńska, G.; Borowik, A.; Wyszkowska, J. Soil dehydrogenases as an indicator of contamination of the environment with petroleum products. Water Air Soil Pollut. 2015, 226, 372. [Google Scholar] [CrossRef] [Green Version]
- Gil-Sotres, F.; Trasar-Cepeda, C.; Leiros, M.C.; Seoane, S. Different approaches to evaluating soil quality using biochemical properties. Soil Biol. Biochem. 2005, 37, 877–887. [Google Scholar] [CrossRef]
- Błońska, E.; Lasota, J.; Zwydak, M. The relationship between soil properties, enzyme activity and land use. For. Res. Pap. 2017, 78, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Luo, K.; Li, X.M.; Wang, D.B.; Zheng, W.; Zeng, G.M.; Liu, J.J. Enhanced efficiency of biological excess sludge hydrolysis under anaerobic digestion by additional enzymes. Bioresour. Technol. 2010, 101, 2924–2930. [Google Scholar] [CrossRef]
- Skawina, T. Characteristics of Reclamation Activities on the External Dump of the „Piaseczno” Sulfur Mine; Institute of Environmental Development and Protection: Kraków, Poland, 1974. [Google Scholar]
- Ziemnicki, S. Reclamation of an Opencast Mine Dump (on the Example of Piaseczno). Problems of Industrialized Regions. PAN, Research Commission of Industrialized Regions; PWN: Warsaw, Poland, 1980. [Google Scholar]
- Węgorek, T. Changes in some properties of the earth material and development of phytocoenoses in the external dump of sulfur mine as a result of target forest reclamation. Sci. Diss. AR Lub. 2003, 275, 1–140. [Google Scholar]
- Classification of Forest Soils in Poland. Warsaw. 2000. Available online: https://www.worldcat.org/title/klasyfikacja-gleb-lesnych-polski/oclc/749272043 (accessed on 19 September 2021).
- International Organization for Standardization. Soil Quality. Sampling; ISO 18400; ISO: Geneva, Switzerland, 2018; Volume 13, p. 131. [Google Scholar]
- PN-EN 15936:2013:02. Available online: https://sklep.pkn.pl/pn-en-15936-2013-02p.html (accessed on 19 September 2021).
- Schinner, F.; Ohlinger, R.; Kandeler, E.; Margesin, R. Methods in Soil Biology; Springer: Berlin/Heidelberg, Germany, 1995. [Google Scholar]
- PN-ISO10381_6:1998. Available online: https://sklep.pkn.pl/pn-iso-10381-6-1998p.html (accessed on 19 September 2021).
- Malina, A. Multidimensional analysis of the Spatial Differentiation of the Structure of the Polish Economy by Voivodship. Publishing House of the Cracow University of Economics; Wydawnictwo Akademii Ekonomicznej: Kraków, Poland, 2004; p. 162. [Google Scholar]
- Leirós, M.C.; Gil-Sotres, F.; Ceccanti, B.; Trasar-Cepeda, M.C.; González-Sangregorio, M.V. Humification processes in reclaimed open-cast lignite mine soils. Soil Biol. Biochem. 1993, 25, 1391–1397. [Google Scholar] [CrossRef]
- Żukowska, G.; Myszura, M.; Zdeb, M.; Pawłowska, M. Carbon Sequestration in Soil as a Sustainable Way of Greenhouse Effect Mitigation—Problems of Sustainable Development. J. Eur. Acad. Sci. Arts 2020, 15, 195–205. [Google Scholar] [CrossRef]
- Brożek, S.; Lasota, J.; Zwydak, M. An attempt to use the forest soil trophism index for the diagnosis of lowland and upland habitats. Acta Agrar. Silvestria Ser. Silvestris 2001, 39, 35–46. [Google Scholar]
- Paradelo, R.; Barral, M.T. Effect of moisture and dis-aggregation on the microbial activity of soil. Soil Till. Res. 2009, 104, 317–319. [Google Scholar] [CrossRef]
- Notaro, K.A.; Medeiros, E.V.; Duda, G.P.; Silva, A.O.; Moura, P.M. Agroforestry systems, nutrients in litter and microbial activity in soils cultivated with coffee at high altitude. Sci Agric. 2014, 71, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Pandey, D.; Agrawal, M.; Bohra, J.S. Effects of conventional tillage and no tillage permutations on extracellular soil enzyme activities and microbial biomass under rice cultivation. Soil Tillage Res. 2014, 136, 51–60. [Google Scholar] [CrossRef]
- de Medeiros, E.V.; Notaro, K.A.; de Barros, J.A.; Moraes, W.S.; Silva, A.O.; Moreira, K.A. Absolute and specific enzymatic activities of sandy entisol from tropical dry forest, monoculture and intercropping areas. Soil Tillage Res. 2015, 145, 208–215. [Google Scholar] [CrossRef]
- Moghimian, N.; Hosseini, S.M.; Kooch, Y.; Darki, B.Z. Impacts of changes in land use/cover on soil microbial and enzyme activity. Catena 2017, 157, 407–414. [Google Scholar] [CrossRef]
- Ahirwal, J.; Kumar, A.; Pietrzykowski, M.; Maiti, S.K. Reclamation of coal mine spoil and its effect on Technosol quality and carbon sequestration: A case study from India. Environ. Sci. Pollut. Res. 2018, 25, 27992–28003. [Google Scholar] [CrossRef]
- Ahirwal, J.; Kumar, A.; Maiti, S.K. Effect of fast-growing trees on soil properties and carbon storage in an afforested coal mine land (India). Minerals 2020, 10, 840. [Google Scholar] [CrossRef]
- Ahirwal, J.; Maiti, S.K. Assessment of soil properties of different land uses generated due to surface coal mining activities in tropical Sal (Shorea robusta) forest, India. Catena 2016, 140, 155–163. [Google Scholar] [CrossRef]
- Simmons, J.A.; Currie, W.S.; Eshleman, K.N.; Kuers, K.; Monteleone, S.; Negley, T.L.; Pohlad, B.R.; Thomas, C.L. Forest to reclaimed mine land use change leads to altered ecosystem structure and function. Ecol. Appl. 2008, 18, 104–118. [Google Scholar] [CrossRef]
- Jing, C.L.; Xu, Z.C.; Zou, P.; Tang, Q.; Li, Y.Q.; You, X.W.; Zhang, C.S. Coastal halophytes alter properties and microbial community structure of the saline soils in the Yellow River Delta China. Appl. Soil Ecol. 2019, 134, 1–7. [Google Scholar] [CrossRef]
- Yu, P.J.; Tang, X.G.; Zhang, A.C.; Fan, G.H.; Liu, S.W. Responses of soil specific enzyme activities to short-term land use conversions in a salt-affected region, northeastern China. Sci. Total Environ. 2019, 15, 687:939–945. [Google Scholar] [CrossRef]
- Bielińska, E.J.; Mocek, A. Aktywność enzymatyczna gleby użytkowanej sadowniczo jako wskaźnik stanu środowiska wywołany stosowaniem ściółek z tworzyw sztucznych. Zesz. Probl. Post. Nauk Rol. 2003, 492, 25–37. [Google Scholar]
- Kao-Knifn, J.T.; Balser, T.C. Elevated CO2 diferentially alters belowground plant and soil microbial community structure in reed canary grass-invaded experimental wetlands. Soil Biol. Biochem. 2007, 39, 517–525. [Google Scholar] [CrossRef]
- Błońska, E.; Lasota, J.; Gruba, P. Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation to Rother properties of soil derived from less and galciofluvial sand. Ecol. Res. 2016, 31, 655–664. [Google Scholar] [CrossRef] [Green Version]
- Kotroczó, Z.; Veres, Z.; Fekete, J.; Krakomperger, Z.; Tóth, J.A.; Lajtha, K.; Tóthmérisz, B. Soil enzyme activity in response to long-term organic matter manipulation. Soil Biol. Biochem. 2014, 70, 237–243. [Google Scholar] [CrossRef]
- Janssens, J.A.; Sampson, D.A.; Curiel-Yuste, J.; Carrara, A.; Cenlemans, R. The carbon cost of fine root turnover in a Scots pine forest. For. Ecol. Manag. 2002, 168, 231–240. [Google Scholar] [CrossRef]
- Holubík, O.; Podrázský, V.; Vopravil, J.; Khel, T.; Remeš, J. Effect of agricultural lands afforestation and tree species composition on the soil reaction, total organic carbon and nitrogen content in the uppermost mineral soil profile. Soil Water Res. 2014, 9, 192–200. [Google Scholar] [CrossRef] [Green Version]
- Kätterer, T.; Bolinder, M.A.; Andrén, O.; Kirchmann, H.; Menichetti, L. Roots contribute more to refractory soil organic matter than above-ground crop residues, as revealed by a long-term field experiment. Agric. Ecosyst. Environ. 2011, 141, 184–192. [Google Scholar] [CrossRef] [Green Version]
- Deng, L.; Shangguang, Z. Afforestation drives soil carbon and nitrogen changes in China. Land Degrad. Dev. 2016, 21, 151–165. [Google Scholar] [CrossRef] [Green Version]
- Kara, O.; Babur, E.; Altun, L.; Seyis, M. Effects of afforestation on microbial biomass C and respiration in eroded soils of Turkey. J. Sustain. For. 2016, 35, 385–396. [Google Scholar] [CrossRef]
- Kang, H.; Gao, H.; Yu, W.; Yi, Y.; Wang, Y.; Ning, M. Changes in soil microbial community structure and function after afforestation depend on species and age: Case study in a subtropical alluvial Island. Sci. Total Environ. 2018, 625, 1423–1432. [Google Scholar] [CrossRef]
- Baldrian, P. Distribution of extracellular enzymes in soils: Spatial heterogeneity and determining factors AT various scales. Soil Sci. Soc. Am. J. 2014, 78, 11–18. [Google Scholar] [CrossRef]
- Józefowska, A.; Woś, B.; Pietrzykowski, M. Tree species and soil substrate effects on soil biota during early soil forming stages at afforested mine sites. Appl. Soil Ecol. 2016, 102, 70–79. [Google Scholar] [CrossRef]
- Gałązka, A.; Gawryjołek, K.; Perzyński, A.; Gałązka, R.; Księżak, J. Changes in Enzymatic Activities and Microbial Communities in Soil under Long-Term Maize Monoculture and Crop Rotation. Pol. J. Environ. Stud. 2017, 26, 39–46. [Google Scholar] [CrossRef]
- Brzezińska, M.; Włodarczyk, T. Enzymes of intracellular redox trasformations (Oxidoreductases). Acta Agrophysica Rozpr. Monogr. 2005, 3, 11–26. [Google Scholar]
- Li, J.J.; Zheng, Y.M.; Yan, J.X.; Li, H.J.; He, J.Z. Succession of plant and soil microbial communities with restoration of abandoned land in the Loess Plateau, China. J. Soil. Sediment 2013, 13, 760–769. [Google Scholar] [CrossRef]
- Ciarkowska, K.; Solek-Podwika, K.; Wieczorek, J. Enzyme activity as an indicator of soil-rehabilitation processes at a zinc and lead ore mining and processing area. J. Environ. Manag. 2014, 132, 250–256. [Google Scholar] [CrossRef]
- Blonska, E.; Jarosław, L.; Zwydak, M. Relacje pomiędzy aktywnością enzymatyczną a właściwościami gleb i sposobem użytkowania. Leśne Prace Badawcze 2017, 78, 1. [Google Scholar] [CrossRef] [Green Version]
- Steiner, C.; Teixeira, W.G.; Lehman, J.; Nehls, T.; de Macêdo, J.L.U.; Blum, W.E.M.; Zech, W. Long term effects of manure, charcoal and mineral fertilization on crop production and fertility on a highly weathered Central Amazonian upland soil. Plant Soil 2007, 291, 275–290. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, R.; Prakash, V.; Kundu, S.; Srisignificantva, A.K.; Gupta, S.M. Long term effects of fertilization on carbon and nitrogen sequestration and aggregate associated carbon and nitrogen in the Indian sub-Himalayas. Nutr. Cycl. Agroecosyst. 2010, 86, 1–16. [Google Scholar] [CrossRef]
- Bueis, T.; Turrión, M.B.; Bravo, F.; Pando, V.; Muscolo, A. Factors determining enzyme activities in soils under Pinus halepensis and Pinus sylvestris plantations in Spain: A basis for establishing sustainable forest management strategies. Ann. For. Sci. 2018, 75, 1–13. [Google Scholar] [CrossRef] [Green Version]
- García, C.; Hernández, T.; Costa, F. Microbial activity in soils under Mediterranean environmental conditions. Soil Biol. Biochem. 1994, 26, 1185–1191. [Google Scholar] [CrossRef]
- Muscolo, A.; Settineri, G.; Attina, E. Early warning indicators of changes in soil ecosystem functioning. Ecol. Indic. 2015, 48, 542–549. [Google Scholar] [CrossRef]
- Zhang, Q.M.; Zhu, L.; Wang, J.; Xie, H.; Wang, J.; Wang, F.; Sun, F. Efects of fomesafen on soil enzyme activity, microbial population, and bacterial community composition. Environ. Monit. Assess. 2014, 186, 2801–2812. [Google Scholar] [CrossRef]
- Fernández-Calviño, D.; Soler-Rovira, P.; Polo, A.; Díaz-Raviña, M.; Arias-Estévez, M.; Plaza, C. Enzyme activities in vineyard soils long-term treated with copper-based fungicides. Soil Biol. Biochem. 2010, 42, 2119–2127. [Google Scholar] [CrossRef]
- Zhang, W.Y.; Yao, D.X.; Zhang, Z.G.; Yang, Q.; An, S.K. Characteristics of soil enzymes and the dominant species of repair trees in the reclamation of coal mine area. J. Coal Sci. Eng. 2013, 19, 256–261. [Google Scholar] [CrossRef]
- Margalef, O.; Sardans, J.; Maspons, J.; Molowny-Horas, R.; Fernández-Martínez, M.; Janssens, I.A.; Peñuelas, J. The effect of global change on soil phosphatase activity. Glob. Chang. Biol. 2021, 27, 5989–6003. [Google Scholar] [CrossRef]
- Margalef, O.; Sardans, J.; Fernández-Martínez, M.; Molowny-Horas, R.; Janssens, I.A.; Ciais, P.; Goll, D.; Richter, A.; Obersteiner, M.; Asensio, D.; et al. Global patterns of phosphatase activity in natural soils. Sci. Rep. 2017, 7, 1337. [Google Scholar] [CrossRef] [Green Version]
- Olander, L.; Vitousek, P. Regulation of soil phosphatase and chitinase activity by N and P availability. Biogeochemistry 2000, 49, 175–191. [Google Scholar] [CrossRef]
- Marklein, A.R.; Houlton, B.Z. Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems. New Phytol. 2012, 193, 696–704. [Google Scholar] [CrossRef]
- Li, Y.T.; Rouland, C.; Benedetti, M.; Li, F.B.; Pando, A.; Lavelle, P.; Dai, J. Microbial biomass, enzyme and mineralization activity in relation to soil organic C, N and P turnover influenced by acid metal stress. Soil Biol. Biochem. 2009, 41, 969–977. [Google Scholar] [CrossRef]
- Dick, R.P.; Burns, R.G. A brief history of soil enzymology research. In Methods of Soil Enzymology; Dick, R.P., Ed.; Soil Science Society of America: Madison, WI, USA, 2011; pp. 1–34. [Google Scholar] [CrossRef]
- Vahed, H.S.; Shahinrokhsar, P.; Rezaei, M. Influence of some soil properties and temperature on urease activity in Wetland Rice soils. Am. Eurasian J. Agric. Environ. Sci. 2011, 11, 310–313. [Google Scholar]
- Ostrowska, A.; Porębska, G. Assessment of the C/N ratio as as indicato of the decomposability of organic matter in forest soils. Ecol. Indic. 2015, 49, 104–109. [Google Scholar] [CrossRef]
- Bielińska, E.J.; Mocek−Płóciniak, A.; Kaczmarek, Z. Indices of the eco−chemical condition of forest soils on a large−area forest fire. Pol. J. Environ. Stud. 2008, 17, 665–671. [Google Scholar]
- Eisenhauer, N.; Lanoue, A.; Strecker, T.; Scheu, S.; Steinauer, K.; Thakur, M.P.; Mommer, L. Root biomass and exudates link plant diversity with soil bacterial and fungal biomass. Sci. Rep. 2017, 7, 44641. [Google Scholar] [CrossRef] [Green Version]
- Vesterdal, L.; Schmidt, I.; Callesen, I.; Nilsson, L.; Gundersen, P. Carbon and nitrogen in forest floor and mineral soil under six common European tree species. For. Ecol. Manag. 2008, 255, 35–48. [Google Scholar] [CrossRef]
- Guckland, A.; Jacob, M.; Flessa, H.; Thomas, F.M.; Leuschner, C. Acidity, nutrient stocks, and organic-matter content in soils of a temperate deciduous forest with different abundance of European beech (Fagus sylvatica L.). J. Plant Nutr. Soil Sci. 2009, 172, 500–511. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Research Point | Soil Texture | Tree stand |
|---|---|---|
| RP 1 | clay | beech dominant stand (Fagus sylvatica L.) |
| RP 2 | bearded birch stand (betula pendula roth) | |
| RP 3 | weak clayey sands | European larch stand (larix decidua mill) |
| RP 4 | red oak stand (Quercus rubra L.) | |
| RP 5 | sands loose | predominantly hornbeam stand (Carpinus betulus) |
| RP 6 | acacia robinia stand (tree stand from the upper part of the slope) (Robinia pseudacacia L.) |
| Research Point | pH | H | S | T | V |
|---|---|---|---|---|---|
| (1 mol KCl) | (cmol(+)·kg−1) | (%) | |||
| RP 1 | 6.90 ± 0.02 | 2.00 ± 0.24 | 20.19 ± 0.05 | 20.19 ± 0.06 | 90.94 ± 0.15 |
| RP 2 | 6.90 ± 0.03 | 1.93 ± 0.13 | 17.89 ± 0.04 | 19.86 ± 0.12 | 89.81 ± 0.14 |
| RP 3 | 5.60 ± 0.02 | 5.10 ± 0.05 | 10.05 ± 0.09 | 15.30 ± 0.03 | 65.56 ± 0.02 |
| RP 4 | 5.80 ± 0.06 | 6.23 ± 0.04 | 8.54 ± 0.03 | 14.92 ± 0.03 | 57.17 ± 0.38 |
| RP 5 | 4.80 ± 0.09 | 14.02 ± 0.15 | 4.85 ± 0.05 | 18.94 ± 0.47 | 26.94 ± 0.20 |
| RP 6 | 4.70 ± 0.05 | 8.02 ± 0.02 | 2.95 ± 0.03 | 10.94 ± 0.02 | 25.76 ± 0.01 |
| Research Point (RP) | TOC | Nt | C/N |
|---|---|---|---|
| g·kg−1 | |||
| RP1 | 31.30 ± 0.10 e | 2.04 ± 0.02 e | 15.34 |
| RP2 | 19.84 ± 0.03 c | 1.23 ± 0.03 c | 16.13 |
| RP3 | 17.48 ± 0.08 b | 0.96 ± 0.02 b | 18.21 |
| RP4 | 20.38 ± 0.07 a | 1.07 ± 0.02 a | 19.05 |
| RP5 | 20.46 ± 0.45 a | 1.08 ± 0.01 a | 18.94 |
| RP6 | 21.67 ± 0.06 d | 1.46 ± 0.01 d | 14.84 |
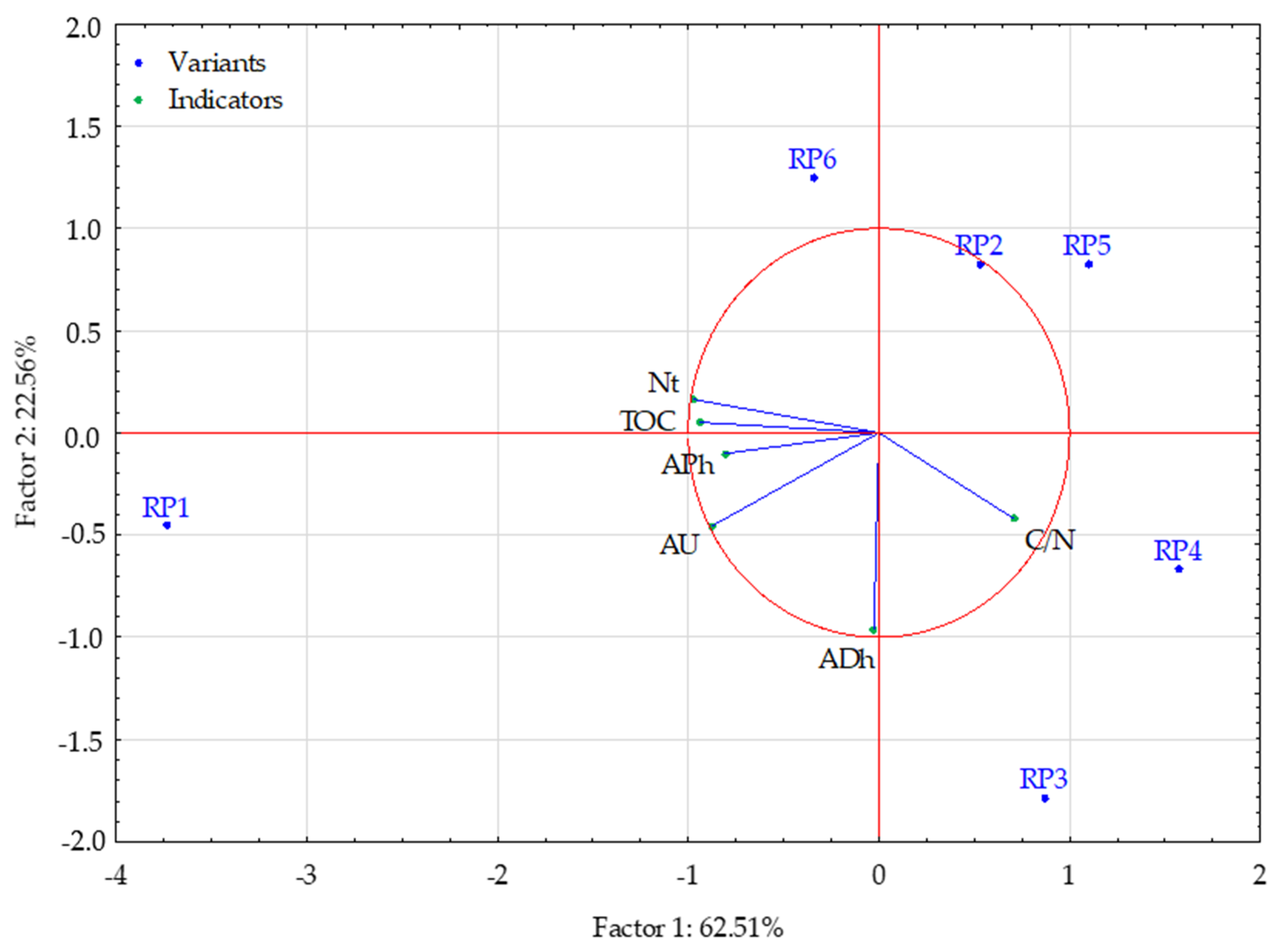
| Variable | between SS | df | within SS | df | F | p * |
|---|---|---|---|---|---|---|
| TOC | 4.68 | 2 | 0.32 | 3 | 22.02 | 0.016 * |
| Nt | 4.78 | 2 | 0.22 | 3 | 32.31 | 0.009 * |
| C/N | 4.64 | 2 | 0.36 | 3 | 19.60 | 0.019 * |
| Clusters | 1 | 2 | 3 |
|---|---|---|---|
| 1 | 0.00 | 2.28 | 1.16 |
| 2 | 0.00 | 1.11 | |
| 3 | 0.00 |
| Variable | Mean Values of the Output Variables in the Clusters | ||
|---|---|---|---|
| Clusters 1 (RP1) | Clusters 2 (RP3, RP4, RP5) | Clusters 3 (RP2, RP6) | |
| TOC | 31.30 | 19.44 | 20.75 |
| Nt | 2.04 | 1.04 | 1.34 |
| C/N | 15.34 | 18.73 | 15.49 |
| Research Point (RP) | ADh | APh | AU |
|---|---|---|---|
| mg TPF∙kg−1∙d−1 | mmol PNP∙kg−1∙d−1 | mg NH4+∙kg−1∙d−1 | |
| RP 1 | 14.33 ± 0.12 d | 167.40 ± 0.10 f | 28.67 ± 0.58 e |
| RP 2 | 6.44 ± 0.04 c | 99.49 ± 0.09 b | 11.24 ± 0.05 c |
| RP 3 | 25.07 ± 0.06 f | 126.87 ± 0.15 d | 15.30 ± 0.10 d |
| RP 4 | 16.37 ± 0.06 e | 76.43 ± 0.15 a | 10.73 ± 0.15 c |
| RP 5 | 2.08 ± 0.07 a | 128.89 ± 0.09 e | 6.28 ± 0.07 a |
| RP 6 | 6.00 ± 0.10 b | 117.10 ± 0.10 c | 9.59 ± 0.09 b |
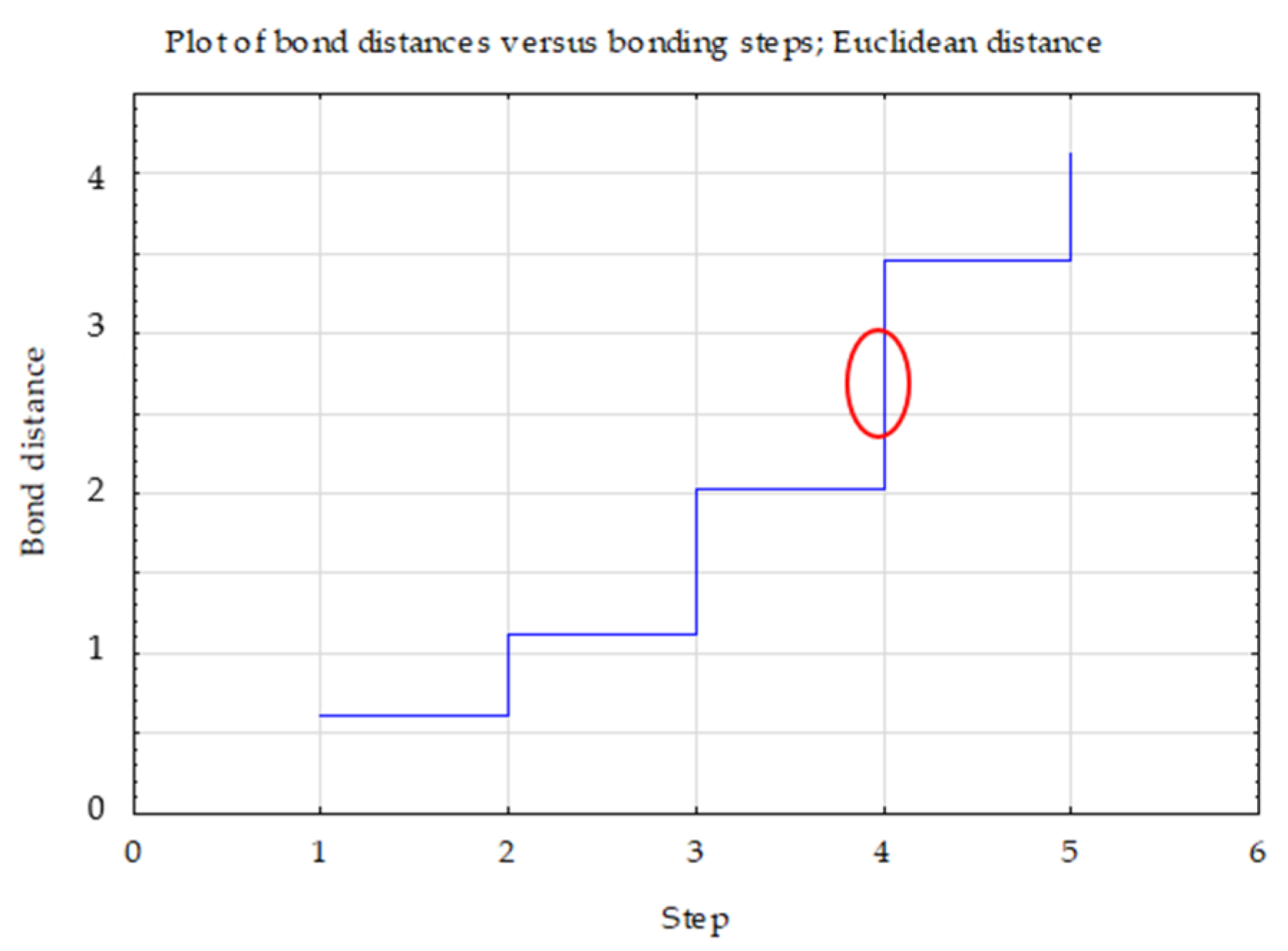
| Variable | between SS | df | within SS | df | F | Istotn. p |
|---|---|---|---|---|---|---|
| dehydrogenases | 4.68 | 2 | 0.32 | 3 | 22.02 | 0.016 * |
| phosphatase | 4.78 | 2 | 0.22 | 3 | 32.31 | 0.009 * |
| urease | 4.64 | 2 | 0.36 | 3 | 19.60 | 0.019 * |
| Clusters | 1 | 2 | 3 |
|---|---|---|---|
| 1 | 0.00 | 1.15 | 1.74 |
| 2 | 0.00 | 1.85 | |
| 3 | 0.00 |
| Variable | Mean Values of the Output Variables in the Clusters | ||
|---|---|---|---|
| Clusters 1 (RP3, RP4) | Clusters 2 (RP2, RP5, RP6) | Clusters 3 (RP1) | |
| dehydrogenases | 20.72 | 4.84 | 14.33 |
| urease | 13.02 | 9.04 | 28.67 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Myszura, M.; Żukowska, G.; Kobyłka, A.; Mazurkiewicz, J. Enzymatic Activity of Soils Forming on an Afforested Heap from an Opencast Sulphur Mine. Forests 2021, 12, 1469. https://doi.org/10.3390/f12111469
Myszura M, Żukowska G, Kobyłka A, Mazurkiewicz J. Enzymatic Activity of Soils Forming on an Afforested Heap from an Opencast Sulphur Mine. Forests. 2021; 12(11):1469. https://doi.org/10.3390/f12111469
Chicago/Turabian StyleMyszura, Magdalena, Grażyna Żukowska, Agata Kobyłka, and Jakub Mazurkiewicz. 2021. "Enzymatic Activity of Soils Forming on an Afforested Heap from an Opencast Sulphur Mine" Forests 12, no. 11: 1469. https://doi.org/10.3390/f12111469
APA StyleMyszura, M., Żukowska, G., Kobyłka, A., & Mazurkiewicz, J. (2021). Enzymatic Activity of Soils Forming on an Afforested Heap from an Opencast Sulphur Mine. Forests, 12(11), 1469. https://doi.org/10.3390/f12111469