Prediction of the Suitable Area of the Chinese White Pines (Pinus subsect. Strobus) under Climate Changes and Implications for Their Conservation


Abstract
1. Introduction
2. Materials and Methods
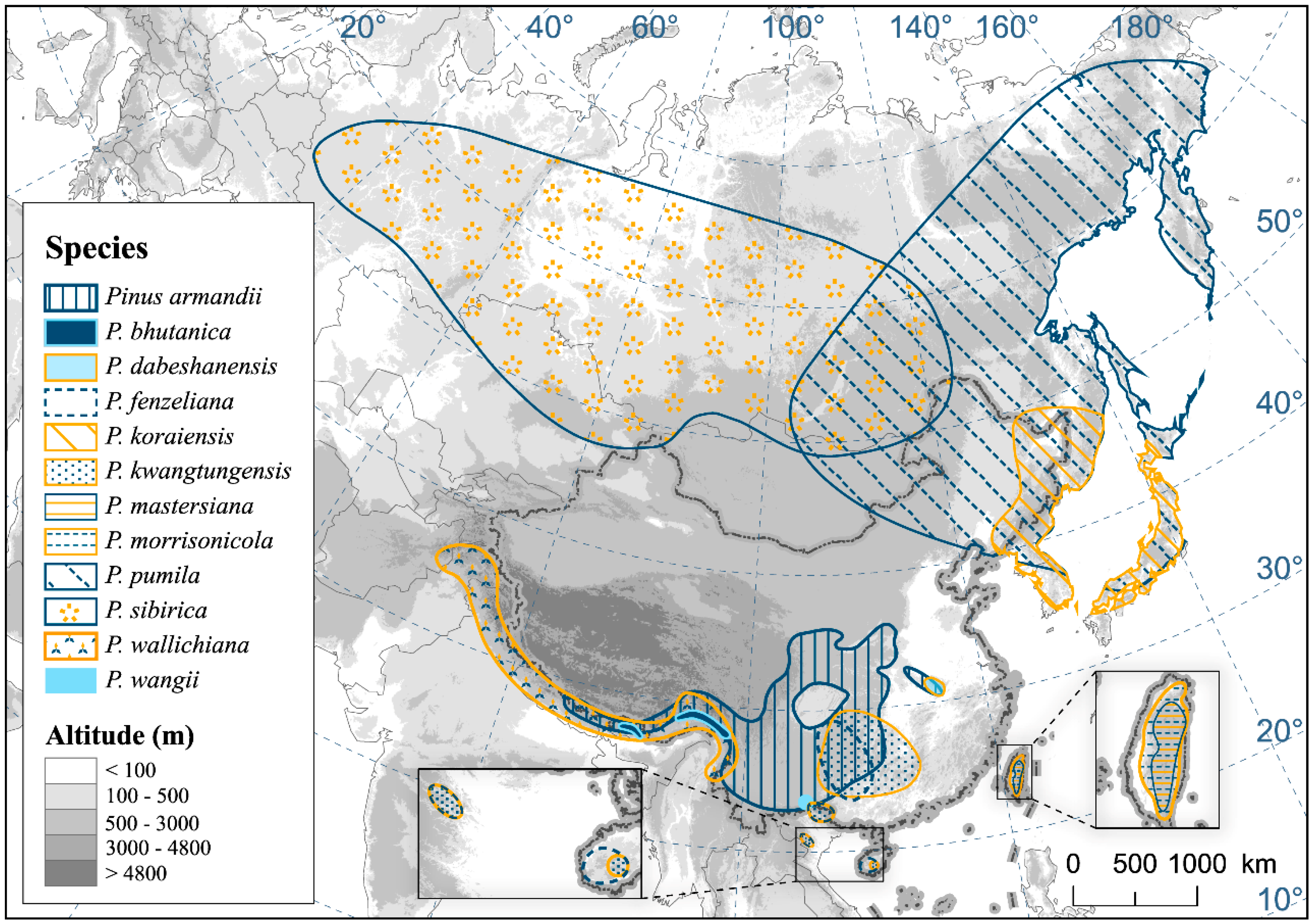
2.1. Study Species Field Survey and Occurrence Data
2.2. Environmental Variables
2.3. Assessment of Important Environmental Factors
2.4. Paleoclimate and Future Climate Scenarios
2.5. Model Simulation and Model Performance Evaluation
2.6. Potential Distribution Areas Classifying, Suitable Habitat Changes in Different Scenarios
3. Results
3.1. Model Accuracy and Key Environmental Variables
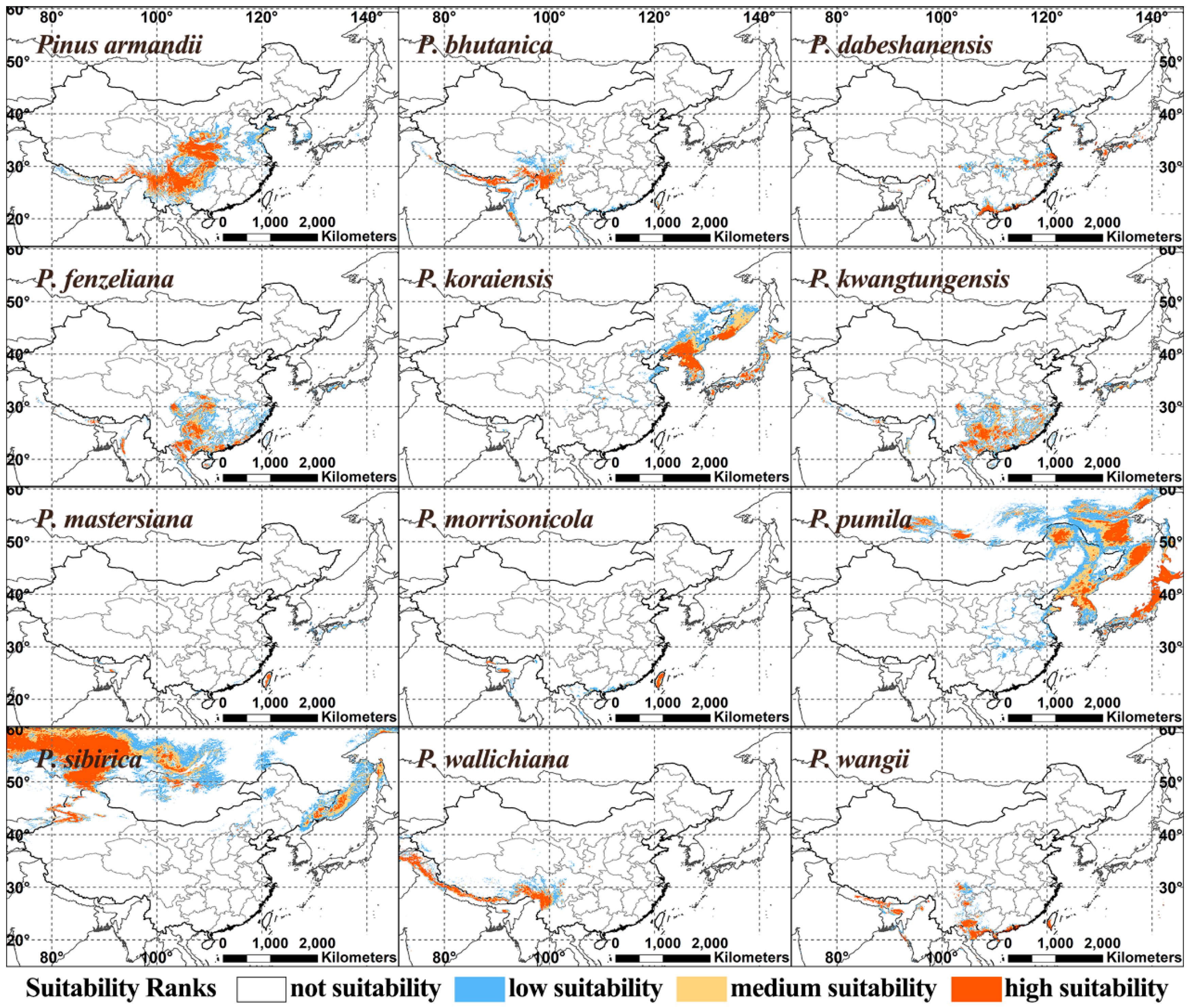
3.2. Suitable Habitats in Current Condition
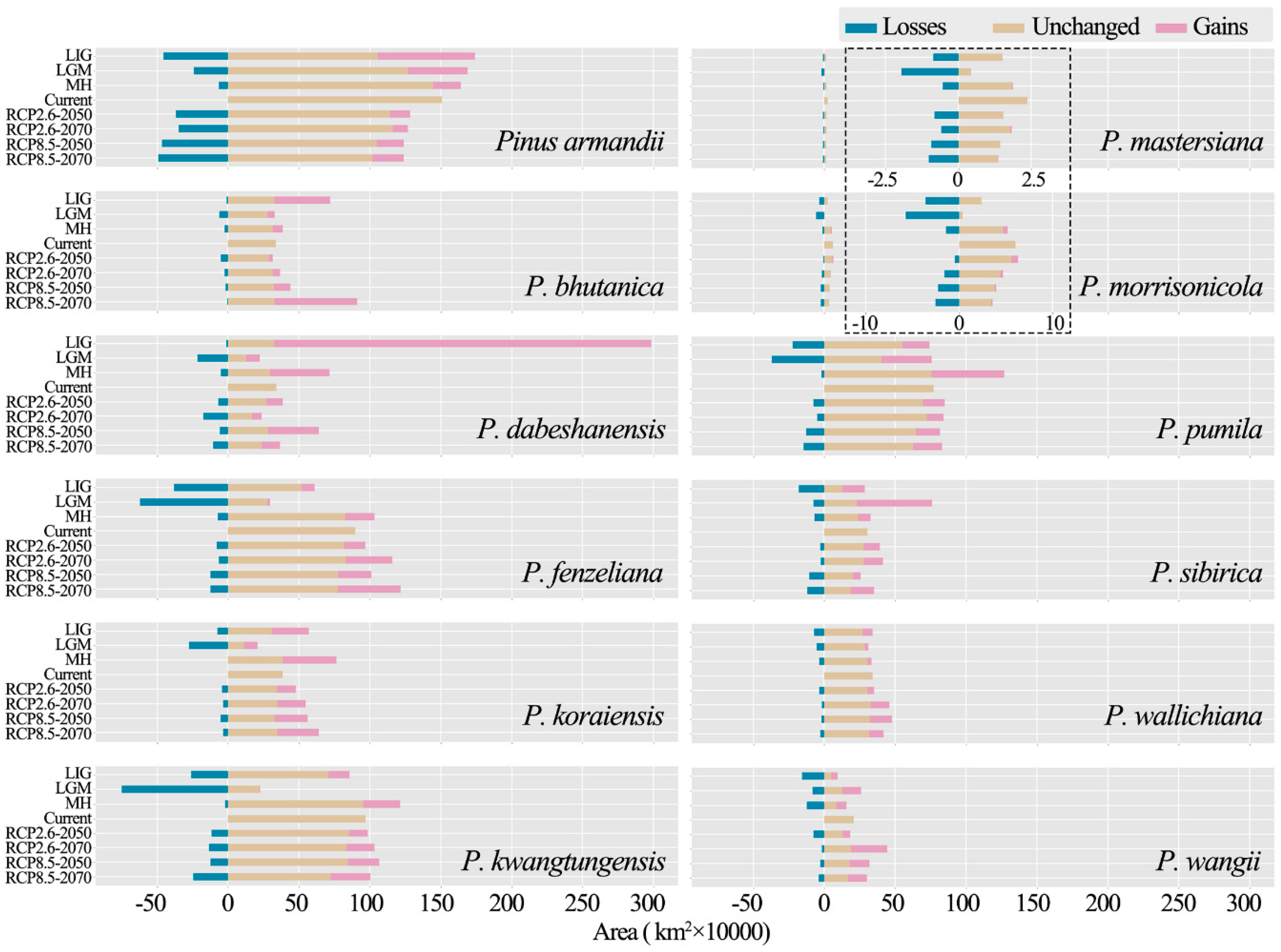
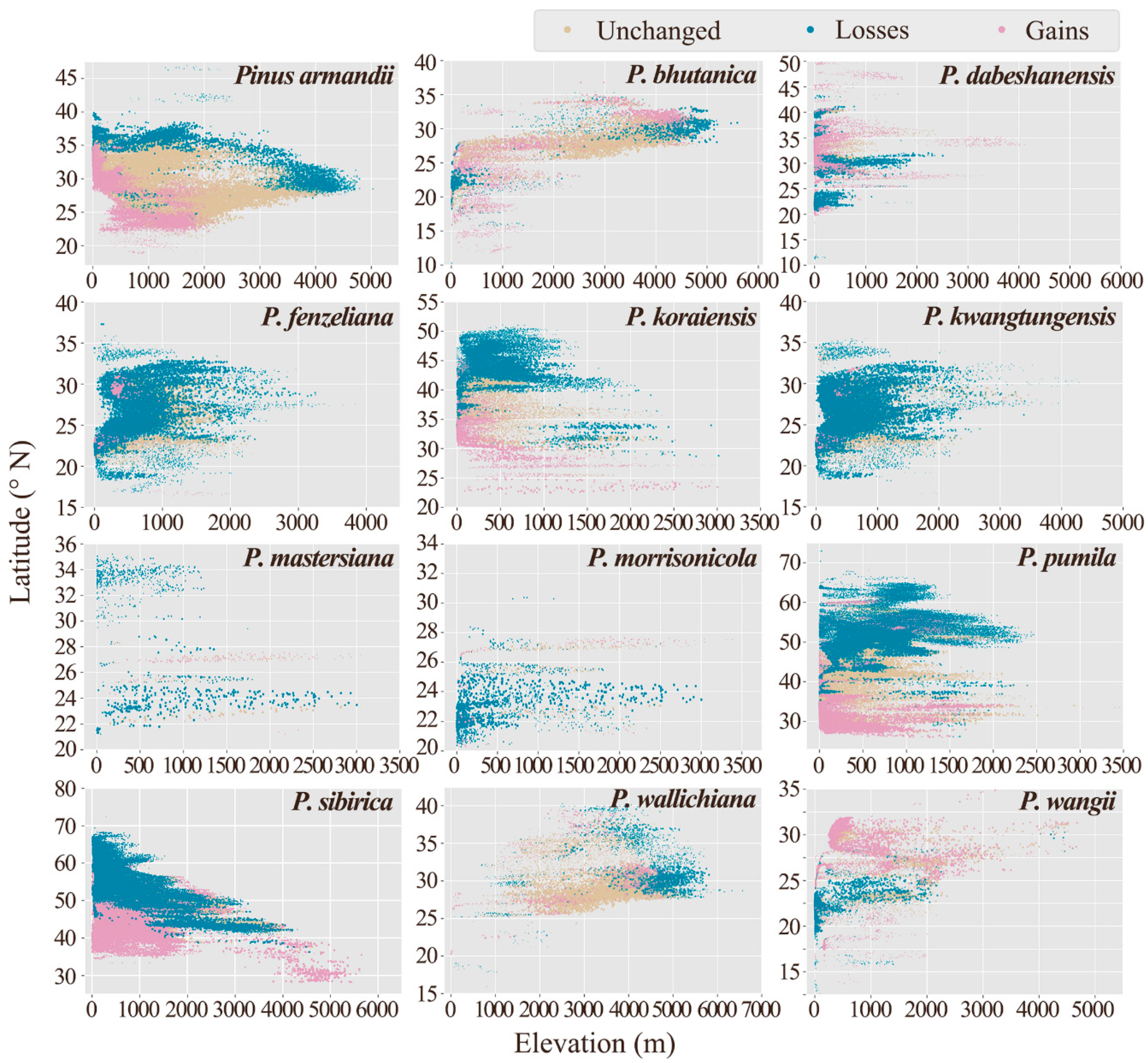
3.3. Distribution Shifts in Different Climate Scenarios
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mittermeier, R.A.; Mittermeier, C.G. Megadiversity: Earth’s Biologically Wealthiest Nations; Agrupación Sierra Madre: Mexico City, Mexico, 1997. [Google Scholar]
- Yu, X. Biodiversity conservation in China: Barriers and future actions. Int. J. Environ. Stud. 2010, 67, 117–126. [Google Scholar] [CrossRef]
- Xu, H.; Wang, S.; Xue, D. Biodiversity conservation in China: Legislation, plans and measures. Biodivers. Conserv. 1999, 8, 819–837. [Google Scholar] [CrossRef]
- Isbell, F.; Gonzalez, A.; Loreau, M.; Cowles, J.; Díaz, S.; Hector, A.; Mace, G.M.; Wardle, D.A.; O’Connor, M.I.; Duffy, J.E. Linking the influence and dependence of people on biodiversity across scales. Nature 2017, 546, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.N.; Balmford, A.; Brook, B.W.; Buettel, J.C.; Galetti, M.; Guangchun, L.; Wilmshurst, J.M. Biodiversity losses and conservation responses in the Anthropocene. Science 2017, 356, 270–275. [Google Scholar] [CrossRef]
- Meng, H.-H.; Zhou, S.-S.; Li, L.; Tan, Y.-H.; Li, J.-W.; Li, J. Conflict between biodiversity conservation and economic growth: Insight into rare plants in tropical China. Biodivers. Conserv. 2019, 28, 523–537. [Google Scholar] [CrossRef]
- You, J.; Qin, X.; Ranjitkar, S.; Lougheed, S.C.; Wang, M.; Zhou, W.; Ouyang, D.; Zhou, Y.; Xu, J.; Zhang, W.; et al. Response to climate change of montane herbaceous plants in the genus Rhodiola predicted by ecological niche modelling. Sci. Rep. 2018, 8, 5879. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; Volume 1535. [Google Scholar]
- Li, Y.; Li, M.; Li, C.; Liu, Z. Optimized maxent model predictions of climate change impacts on the suitable distribution of Cunninghamia lanceolata in China. Forests 2020, 11, 302. [Google Scholar] [CrossRef]
- Wang, D.; Cui, B.; Duan, S.; Chen, J.; Fan, H.; Lu, B.; Zheng, J. Moving north in China: The habitat of Pedicularis kansuensis in the context of climate change. Sci. Total Environ. 2019, 697, 133979. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Tao, J. Predicting the potential distribution of Paeonia veitchii (Paeoniaceae) in China by incorporating climate change into a Maxent model. Forests 2019, 10, 190. [Google Scholar] [CrossRef]
- Bigelow, N.H.; Brubaker, L.B.; Edwards, M.E.; Harrison, S.P.; Prentice, I.C.; Anderson, P.M.; Andreev, A.A.; Bartlein, P.J.; Christensen, T.R.; Cramer, W. Climate change and Arctic ecosystems: 1. Vegetation changes north of 55° N between the last glacial maximum, mid-Holocene, and present. J. Geophys. Res. Atmos. 2003, 108, 8170. [Google Scholar] [CrossRef]
- Harrison, S.; Yu, G.; Takahara, H.; Prentice, I. Palaeovegetation—Diversity of temperate plants in east Asia. Nature 2001, 413, 129–130. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, G. The genetic legacy of the Quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Street-Perrott, F.A.; Metcalfe, S.E.; Brenner, M.; Moreland, M.; Freeman, K.H. Climate change as the dominant control on glacial-interglacial variations in C3 and C4 plant abundance. Science 2001, 293, 1647–1651. [Google Scholar] [CrossRef] [PubMed]
- Prentice, I.C.; Jolly, D.; Afanas’Eva, N.B.; Ager, T.A.; Zheng, Z. Mid-Holocene and glacial-maximum vegetation geography of the northern continents and Africa. J. Biogeogr. 2000, 27, 507–519. [Google Scholar] [CrossRef]
- Shen, L.; Chen, X.Y.; Li, Y.Y. Glacial refugia and postglacial recolonization patterns of organisms. Acta Ecol. Sin. 2002, 22, 1983–1990. [Google Scholar]
- Yu, G.; Chen, X.; Ni, J.; Cheddadi, R.; Guiot, J.; Han, H.; Harrison, S.P.; Huang, C.; Ke, M.; Kong, Z. Palaeovegetation of China: A pollen data-based synthesis for the mid-Holocene and last glacial maximum. J. Biogeogr. 2000, 27, 635–664. [Google Scholar] [CrossRef]
- Stewart, J.R.; Lister, A.M.; Barnes, I.; Dalén, L. Refugia revisited: Individualistic responses of species in space and time. Proc. Biol. Sci. 2010, 277, 661–671. [Google Scholar] [CrossRef]
- Feng, G.; Ma, Z.; Sandel, B.; Mao, L.; Normand, S.; Ordonez, A.; Svenning, J.C. Species and phylogenetic endemism in angiosperm trees across the Northern Hemisphere are jointly shaped by modern climate and glacial–interglacial climate change. Glob. Ecol. Biogeogr. 2019, 28, 1393–1402. [Google Scholar] [CrossRef]
- Chen, I.C.; Hill, J.K.; Ohlemuller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Nogués-Bravo, D.; Araújo, M.B.; Errea, M.P.; Martínez-Rica, J.P. Exposure of global mountain systems to climate warming during the 21st Century. Glob. Environ. Chang. 2007, 17, 420–428. [Google Scholar] [CrossRef]
- Broennimann, O.; Thuiller, W.; Hughes, G.; Midgley, G.F.; Alkemade, J.R.; Guisan, A. Do geographic distribution, niche property and life form explain plants’ vulnerability to global change? Glob. Chang. Biol. 2006, 12, 1079–1093. [Google Scholar] [CrossRef]
- Malcolm, J.R.; Liu, C.; Neilson, R.P.; Hansen, L.; Hannah, L. Global warming and extinctions of endemic species from biodiversity hotspots. Conserv. Biol. 2006, 20, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.D. Translocation of species, climate change, and the end of trying to recreate past ecological communities. Trends Ecol. Evol. 2011, 26, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Gibson, L.; Lee, T.M.; Koh, L.P.; Brook, B.W.; Gardner, T.A.; Barlow, J.; Peres, C.A.; Bradshaw, C.J.; Laurance, W.F.; Lovejoy, T.E. Primary forests are irreplaceable for sustaining tropical biodiversity. Nature 2011, 478, 378–381. [Google Scholar] [CrossRef] [PubMed]
- Watson, J.E.; Evans, T.; Venter, O.; Williams, B.; Tulloch, A.; Stewart, C.; Thompson, I.; Ray, J.C.; Murray, K.; Salazar, A. The exceptional value of intact forest ecosystems. Nat. Ecol. Evol. 2018, 2, 599–610. [Google Scholar] [CrossRef]
- Crausbay, S.D.; Higuera, P.E.; Sprugel, D.G.; Brubaker, L.B. Fire catalyzed rapid ecological change in lowland coniferous forests of the Pacific Northwest over the past 14,000 years. Ecology 2017, 98, 1–14. [Google Scholar] [CrossRef]
- Gernandt, D.S.; López, G.G.; García, S.O.; Liston, A. Phylogeny and classification of Pinus. Taxon 2005, 54, 29–42. [Google Scholar] [CrossRef]
- Hao, Z.Z.; Liu, Y.Y.; Nazaire, M.; Wei, X.X.; Wang, X.Q. Molecular phylogenetics and evolutionary history of sect. Quinquefoliae (Pinus): Implications for Northern Hemisphere biogeography. Mol. Phylogenet. Evol. 2015, 87, 65–79. [Google Scholar] [CrossRef]
- Cheng, W.C.; Fu, L.K. Pinus Linn. In Flora Reipublicae Popularis Sinicae; Science Press: Beijing, China, 1978; Volume 7, pp. 204–281. [Google Scholar]
- Fu, L.G.; Li, N.; Mill, R.R. Pinaceae. In Flora of China; Science Press and Missouri Botanical Garden Press: Beijing, China; St. Louis, MO, USA, 1999; Volume 4, pp. 11–52. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Version 2020-1. Available online: https://www.iucnredlist.org (accessed on 16 April 2020).
- National Forestry Administration. List of National Key Protected Wild Plants (First Group). Available online: http://www.forestry.gov.cn/main/3954/20180925/143410933280757.html (accessed on 23 August 2020).
- Dou, J.J.; Zhou, R.C.; Tang, A.J.; Ge, X.J.; Wu, W. Development and characterization of nine microsatellites for an endangered tree, Pinus wangii (Pinaceae). Appl. Plant Sci. 2013, 1, 1200134. [Google Scholar] [CrossRef]
- Li, Z.-H.; Yang, C.; Mao, K.-S.; Ma, Y.-Z.; Liu, J.; Liu, Z.-L.; Deng, T.-T.; Zhao, G.-F. Molecular identification and allopatric divergence of the white pine species in China based on the cytoplasmic DNA variation. Biochem. Syst. Ecol. 2015, 61, 161–168. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Jin, W.T.; Wei, X.X.; Wang, X.Q. Cryptic speciation in the Chinese white pine (Pinus armandii): Implications for the high species diversity of conifers in the Hengduan Mountains, a global biodiversity hotspot. Mol. Phylogenet. Evol. 2019, 138, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Shuang, T. Molecular evidence for glacial expansion and interglacial retreat during Quaternary climatic changes in a montane temperate pine (Pinus kwangtungensis Chun ex Tsiang) in southern China. Plant Syst. Evol. 2010, 284, 219–229. [Google Scholar] [CrossRef]
- Tao, C.; Li, X.; Wang, Q.; Cui, G. Relationships between geographical distribution of endangered Pinus kwangtungensis and climate in China. Plant Sci. J. 2012, 30, 577–583. [Google Scholar] [CrossRef]
- Tong, Y.; Durka, W.; Zhou, W.; Zhou, L.; Yu, D.; Dai, L. Ex situ conservation of Pinus koraiensis can preserve genetic diversity but homogenizes population structure. For. Ecol. Manag. 2020, 465, 117820. [Google Scholar] [CrossRef]
- Tong, Y.W.; Lewis, B.J.; Zhou, W.M.; Mao, C.R.; Wang, Y.; Zhou, L.; Yu, D.P.; Dai, L.M.; Qi, L. Genetic diversity and population structure of natural Pinus koraiensis populations. Forests 2020, 11, 39. [Google Scholar] [CrossRef]
- Wang, F.; Liang, D.; Pei, X.; Zhang, Q.; Zhang, P.; Zhang, J.; Lu, Z.; Yang, Y.; Liu, G.; Zhao, X. Study on the physiological indices of Pinus sibirica and Pinus koraiensis seedlings under cold stress. J. For. Res. 2019, 30, 1255–1265. [Google Scholar] [CrossRef]
- Xiang, X.-Y.; Zhang, Z.-X.; Duan, R.-Y.; Zhang, X.-P.; Wu, G.-L. Genetic diversity and structure of Pinus dabeshanensis revealed by expressed sequence tag-simple sequence repeat (EST-SSR) markers. Biochem. Syst. Ecol. 2015, 61, 70–77. [Google Scholar] [CrossRef]
- You, J.; Lin, L.L.; Xie, L.; Cui, G.F.; University, B.F. Interaspecific and intraspecific competitions of Pinus fenzeliana in Yinggeling, Hainan Province. Guihaia 2017, 37, 776–782. [Google Scholar] [CrossRef]
- Zhou, Y.; Jiang, H.; Yang, W.Z.; Zhang, S.S.; Xiang, Z.Y. Study on Stock of Pinus wangii,an Extremely Small Population Species. J. West China For. Sci. 2012, 41, 80–83. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Ackerly, D.D.; Loarie, S.R.; Cornwell, W.K.; Weiss, S.B.; Hamilton, H.; Branciforte, R.; Kraft, N.J.B. The geography of climate change: Implications for conservation biogeography. Divers. Distrib. 2010, 16, 476–487. [Google Scholar] [CrossRef]
- Adhikari, D.; Tiwary, R.; Singh, P.P.; Upadhaya, K.; Singh, B.; Haridasan, K.E.; Bhatt, B.B.; Chettri, A.; Barik, S.K. Ecological niche modeling as a cumulative environmental impact assessment tool for biodiversity assessment and conservation planning: A case study of critically endangered plant Lagerstroemia minuticarpa in the Indian Eastern Himalaya. J. Environ. Manag. 2019, 243, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Chucholl, C. Niche-based species distribution models and conservation planning for endangered freshwater crayfish in south-western Germany. Aquat. Conserv. 2017, 27, 698–705. [Google Scholar] [CrossRef]
- Qin, A.; Liu, B.; Guo, Q.; Bussmann, R.W.; Ma, F.; Jian, Z.; Xu, G.; Pei, S. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch., an extremely endangered conifer from southwestern China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Sarania, B.; Devi, A.; Kumar, A.; Sarma, K.; Gupta, A.K. Predictive distribution modeling and population status of the endangered Macaca munzala in Arunachal Pradesh, India. Am. J. Primatol. 2017, 79, 1–10. [Google Scholar] [CrossRef]
- Wang, W.; Gao, S.; Wang, S. Predictive studies of potential invasive areas for four poisonous weeds in Gansu Grassland. Acta Ecol. Sin. 2019, 39, 4301–5307. [Google Scholar] [CrossRef]
- Zeng, Y.; Low, B.W.; Yeo, D.C. Novel methods to select environmental variables in MaxEnt: A case study using invasive crayfish. Ecol. Modell. 2016, 341, 5–13. [Google Scholar] [CrossRef]
- Chen, F.; Du, Y.; Niu, S.; Zhao, J. Modeling forest lightning fire occurrence in the Daxinganling Mountains of Northeastern China with MAXENT. Forests 2015, 6, 1422–1438. [Google Scholar] [CrossRef]
- Duan, H.; Xia, S.; Hou, X.; Liu, Y.; Yu, X. Conservation planning following reclamation of intertidal areas throughout the Yellow and Bohai Seas, China. Biodivers. Conserv. 2019, 28, 3787–3801. [Google Scholar] [CrossRef]
- Raney, P.A.; Leopold, D.J. Fantastic wetlands and where to find them: Modeling rich fen distribution in New York State with Maxent. Wetlands 2018, 38, 81–93. [Google Scholar] [CrossRef]
- Roces-Díaz, J.V.; Jiménez-Alfaro, B.; Chytrý, M.; Díaz-Varela, E.R.; Álvarez-Álvarez, P. Glacial refugia and mid-Holocene expansion delineate the current distribution of Castanea sativa in Europe. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 491, 152–160. [Google Scholar] [CrossRef]
- Shitara, T.; Nakamura, Y.; Matsui, T.; Tsuyama, I.; Ohashi, H.; Kamijo, T. Formation of disjunct plant distributions in Northeast Asia: A case study of Betula davurica using a species distribution model. Plant Ecol. 2018, 219, 1105–1115. [Google Scholar] [CrossRef]
- Wang, S.; Xu, X.; Shrestha, N.; Zimmermann, N.E.; Tang, Z.; Wang, Z. Response of spatial vegetation distribution in China to climate changes since the Last Glacial Maximum (LGM). PLoS ONE 2017, 12, e0175742. [Google Scholar] [CrossRef]
- Songer, M.; Delion, M.; Biggs, A.; Huang, Q. Modeling impacts of climate change on giant panda habitat. Int. J. Ecol. 2012, 2012, 108752. [Google Scholar] [CrossRef]
- Tang, C.Q.; Dong, Y.-F.; Herrando-Moraira, S.; Matsui, T.; Ohashi, H.; He, L.-Y.; Nakao, K.; Tanaka, N.; Tomita, M.; Li, X.-S. Potential effects of climate change on geographic distribution of the Tertiary relict tree species Davidia involucrata in China. Sci. Rep. 2017, 7, 43822. [Google Scholar] [CrossRef] [PubMed]
- Vieilledent, G.; Cornu, C.; Sanchez, A.C.; Pock-Tsy, J.-M.L.; Danthu, P. Vulnerability of baobab species to climate change and effectiveness of the protected area network in Madagascar: Towards new conservation priorities. Biol. Conserv. 2013, 166, 11–22. [Google Scholar] [CrossRef]
- Bai, Y.; Wei, X.; Li, X. Distributional dynamics of a vulnerable species in response to past and future climate change: A window for conservation prospects. PeerJ 2018, 6, e4287. [Google Scholar] [CrossRef]
- Hayata, B. Pinus armandii var. mastersiana (Hayata) Hayata. J. Coll. Sci. Imp. Univ. Tokyo 1908, XXV, 217. [Google Scholar]
- Li, J.; Fan, G.; He, Y. Predicting the current and future distribution of three Coptis herbs in China under climate change conditions, using the MaxEnt model and chemical analysis. Sci. Total Environ. 2020, 698, 134141. [Google Scholar] [CrossRef]
- Li, M.; He, J.; Zhao, Z.; Lyu, R.; Yao, M.; Cheng, J.; Xie, L. Predictive modelling of the distribution of Clematis sect. Fruticella s. str. under climate change reveals a range expansion during the Last Glacial Maximum. PeerJ 2020, 8, e8729. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J.; Hijmans, M.R.J. Dismo: Species Distribution Modeling. Available online: https://cran.r-project.org/package=dismo (accessed on 24 October 2019).
- Grierson, A.J.C.; Long, D.G.; Page, C.N. Notes relating to the flora of Bhutan:(III). Pinus bhutanica: A new 5-needle pine from Bhutan and India. Notes R. Bot. Gard. Edinb. 1980, 38, 297–310. [Google Scholar]
- Komarov, V.L.e. Flora of the USSR; Izdatel’stvo Akademii Nauk SSSR: Leningrad, Russia, 1934; Volume I. [Google Scholar]
- Syfert, M.M.; Smith, M.J.; Coomes, D.A. The effects of sampling bias and model complexity on the predictive performance of MaxEnt species distribution models. PLoS ONE 2013, 8, e55158. [Google Scholar] [CrossRef]
- Brown, J.L. SDM toolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Beckmann, M.; Václavík, T.; Manceur, A.M.; Šprtová, L.; von Wehrden, H.; Welk, E.; Cord, A.F. gl UV: A global UV-B radiation data set for macroecological studies. Methods Ecol. Evol. 2014, 5, 372–383. [Google Scholar] [CrossRef]
- IIASA; FAO. Global Agro-cological Zones (GAEZ v3.0); IIASA: Laxenburg, Austria; FAO: Rome, Italy, 2012. [Google Scholar]
- Baldwin, R.A. Use of maximum entropy modeling in wildlife research. Entropy 2009, 11, 854–866. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Fourcade, Y.; Engler, J.O.; Rödder, D.; Secondi, J. Mapping species distributions with MAXENT using a geographically biased sample of presence data: A performance assessment of methods for correcting sampling bias. PLoS ONE 2014, 9, e97122. [Google Scholar] [CrossRef]
- Phillips, S.J. A Brief Tutorial on Maxent. Available online: https://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 13 November 2019).
- Gent, P.R.; Danabasoglu, G.; Donner, L.J.; Holland, M.M.; Hunke, E.C.; Jayne, S.R.; Lawrence, D.M.; Neale, R.B.; Rasch, P.J.; Vertenstein, M.; et al. The Community Climate System Model Version 4. J Clim. 2011, 24, 4973–4991. [Google Scholar] [CrossRef]
- Duffy, K.J.; Jacquemyn, H. Climate change increases ecogeographical isolation between closely related plants. J. Ecol. 2019, 107, 167–177. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A. Insights into the area under the receiver operating characteristic curve (AUC) as a discrimination measure in species distribution modelling. Glob. Ecol. Biogeogr. 2012, 21, 498–507. [Google Scholar] [CrossRef]
- Liu, C.; White, M.; Newell, G. Measuring and comparing the accuracy of species distribution models with presence-absence data. Ecography 2011, 34, 232–243. [Google Scholar] [CrossRef]
- Lei, Z.; Wang, L.; Liu, S.; Sun, P.; Zhang, X. An evaluation of four threshold selection methods in species occurrence modelling with random forest: Case studies with Davidia involucrata and Cunninghamia lanceolata. Chin. J. Plant. Ecol. 2017, 41, 387–395. [Google Scholar] [CrossRef]
- Thuiller, W.; Georges, D.; Engler, R.; Breiner, F.; Georges, M.D.; Thuiller, C.W. Package ‘Biomod2′: Ensemble Platform for Species Distribution Modeling. Available online: https://cran.r-project.org/web/packages/biomod2/biomod2.pdf (accessed on 6 March 2020).
- Wang, Y.; Xie, B.; Wan, F.; Xiao, Q.; Dai, L. Application of ROC curve analysis in evaluating the performance of alien species potential distribution models. Biodiv. Sci. 2007, 15, 365–372. [Google Scholar] [CrossRef]
- Luo, M.; Wang, H.; Lyu, Z. Evaluating the performance of species distribution models Biomod2 and MaxEnt using the giant panda distribution data. Chin. J. Appl. Ecol. 2017, 28, 4001–4006. [Google Scholar] [CrossRef]
- Yang, X.-Q.; Kushwaha, S.; Saran, S.; Xu, J.; Roy, P. Maxent modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda L. in Lesser Himalayan foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Janská, V.; Jiménez-Alfaro, B.; Chytrý, M.; Divíšek, J.; Anenkhonov, O.; Korolyuk, A.; Lashchinskyi, N.; Culek, M. Palaeodistribution modelling of European vegetation types at the Last Glacial Maximum using modern analogues from Siberia: Prospects and limitations. Quat. Sci. Rev. 2017, 159, 103–115. [Google Scholar] [CrossRef]
- Zhuang, H.; Zhang, Y.; Wang, W.; Ren, Y.; Liu, F.; Du, J.; Zhou, Y. Optimized hot spot analysis for probability of species distribution under different spatial scales based on MaxEnt model: Manglietia insignis case. Biodiv. Sci. 2018, 26, 931–940. [Google Scholar] [CrossRef]
- Diekmann, O.E.; Coyer, J.A.; Ferreira, J.; Olsen, J.L.; Stam, W.T.; Pearson, G.A.; Serrão, E.A. Population genetics of Zostera noltii along the west Iberian coast: Consequences of small population size, habitat discontinuity and near-shore currents. Mar. Ecol. Prog. Ser. 2005, 290, 89–96. [Google Scholar] [CrossRef]
- Kobayashi, T.; Sota, T. Contrasting effects of habitat discontinuity on three closely related fungivorous beetle species with diverging host-use patterns and dispersal ability. Ecol. Evol. 2019, 9, 2475–2486. [Google Scholar] [CrossRef] [PubMed]
- Bush, A.; Mokany, K.; Catullo, R.; Hoffmann, A.; Kellermann, V.; Sgrò, C.; McEvey, S.; Ferrier, S. Incorporating evolutionary adaptation in species distribution modelling reduces projected vulnerability to climate change. Ecol. Lett. 2016, 19, 1468–1478. [Google Scholar] [CrossRef]
- Chen, M. A preliminary study on climate re-gionalization in China. Sci. Geogr. Sin. 1990, 010, 308–315. [Google Scholar]
- Li, G.; Du, S.; Guo, K. Evaluation of limiting climatic factors and simulation of a climatically suitable habitat for Chinese sea buckthorn. PLoS ONE 2015, 10, e0131659. [Google Scholar] [CrossRef]
- Gao, L.; Wang, X. Response of Pinus koraiensis and Picea jezoensis var. komarovii to climate in the transition zone of Changbai Mountain, China. Chin. J. Plant Ecol. 2011, 35, 27–34. [Google Scholar] [CrossRef]
- Yin, X.; Wu, X. Modelling analysis of Huashan pine growth response to climate. Q. J. Appl. Meteorol. 1995, 006, 257–264. [Google Scholar]
- Feng, J. Spatial patterns of species diversity of seed plants in China and their climatic explanation. Biodiv. Sci. 2008, 16, 470–476. [Google Scholar] [CrossRef]
- Yang, Q.S.; Xing, Y.W.; Zhou, Z.K. Modern geographical distribution of Tsuga and its climatic conditions in the Asian monsoon region. Acta Bot. Yunnanica 2009, 31, 389–398. [Google Scholar] [CrossRef]
- Chong, K.; Yong, W.; Tan, K. Advances on research of vernalization in higher plants. Chin. Bull. Bot. 1999, 16, 481–487. [Google Scholar]
- Ma, C.; Zhang, Y.; An, D. Geographic variation of cold hardiness of Pinus armandii. For. Res. 1990, 3, 113–118. [Google Scholar]
- Day, T.A.; Neale, P.J. Effects of UV-B radiation on terrestrial and aquatic primary producers. Annu. Rev. Ecol. Syst. 2002, 33, 371–396. [Google Scholar] [CrossRef]
- Ren, J.; Li, C. Research advances in response of seed plants to enhanced ultraviolet-B radiation. Chin. J. Ecol. 2005, 24, 315–320. [Google Scholar]
- Fu, G.; Shen, Z.-X. Effects of enhanced UV-B radiation on plant physiology and growth on the Tibetan Plateau: A meta-analysis. Acta Physiol. Plant. 2017, 39, 85. [Google Scholar] [CrossRef]
- Laakso, K.; Sullivan, J.H.; Huttunen, S. The effects of UV-B radiation on epidermal anatomy in loblolly pine (Pinus taeda L.) and Scots pine (Pinus sylvestris L.). Plant Cell Environ. 2000, 23, 461–472. [Google Scholar] [CrossRef]
- Matías, L.; Linares, J.C.; Sánchez-Miranda, Á.; Jump, A.S. Contrasting growth forecasts across the geographical range of Scots pine due to altitudinal and latitudinal differences in climatic sensitivity. Glob. Chang. Biol. 2017, 23, 4106–4116. [Google Scholar] [CrossRef]
- Yang, S.; Ding, Z.; Li, Y.; Wang, X.; Jiang, W.; Huang, X. Warming-induced northwestward migration of the East Asian monsoon rain belt from the Last Glacial Maximum to the mid-Holocene. PNAS 2015, 112, 13178–13183. [Google Scholar] [CrossRef]
- Xiao, J.; Shang, Z.; Shu, Q.; Yin, J.; Wu, X. The vegetation feature and palaeoenvironment significance in the mountainous interior of southern China from the Last Glacial Maximum. Sci. China Earth Sci. 2017, 61, 71–81. [Google Scholar] [CrossRef]
- Qiu, Y.-X.; Fu, C.-X.; Comes, H.P. Plant molecular phylogeography in China and adjacent regions: Tracing the genetic imprints of Quaternary climate and environmental change in the world’s most diverse temperate flora. Mol. Phylogenet. Evol. 2011, 59, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gao, P.; Kang, M.; Lowe, A.J.; Huang, H. Refugia within refugia: The case study of a canopy tree (Eurycorymbus cavaleriei) in subtropical China. J. Biogeogr. 2009, 36, 2156–2164. [Google Scholar] [CrossRef]
- Zheng, Z. Response of altitudinal belts of vegetation to the Late Quaternary climatic changes in tropical Asia. Geogr. Res. 1999, 18, 96–104. [Google Scholar]
- Cheng, Y.; Jiang, W. Vegetation and climate changes since the Last Glacial Maximum in the Northern Loess Plateau. Quat. Sci. 2011, 31, 982–989. [Google Scholar] [CrossRef]
- Hou, K.; Qian, H.; Zhang, Y.; Zhang, Q. Influence of tectonic uplift of the Qinling Mountains on the paleoclimatic environment of surrounding areas: Insights from loess–paleosol sequences, Weihe Basin, central China. Catena 2020, 187, 104336. [Google Scholar] [CrossRef]
- Xu, X.X.; Cheng, F.Y.; Peng, L.P.; Sun, Y.Q.; Hu, X.G.; Li, S.Y.; Xian, H.L.; Jia, K.H.; Abbott, R.J.; Mao, J.F. Late Pleistocene speciation of three closely related tree peonies endemic to the Qinling–Daba Mountains, a major glacial refugium in Central China. Ecol. Evol. 2019, 9, 7528–7548. [Google Scholar] [CrossRef]
- Zhang, H.; Lu, H.; Jiang, S.-Y.; Vandenberghe, J.; Wang, S.; Cosgrove, R. Provenance of loess deposits in the Eastern Qinling Mountains (central China) and their implications for the paleoenvironment. Quat. Sci. Rev. 2012, 43, 94–102. [Google Scholar] [CrossRef]
- Cong, M.; Xu, Y.; Tang, L.; Yang, W.; Jian, M. Predicting the dynamic distribution of Sphagnum bogs in China under climate change since the last interglacial period. PLoS ONE 2020, 15, e0230969. [Google Scholar] [CrossRef]
- Büchi, L.; Vuilleumier, S. Coexistence of specialist and generalist species is shaped by dispersal and environmental factors. Am. Nat. 2014, 183, 612–624. [Google Scholar] [CrossRef]
- Liang, Y.; Duveneck, M.J.; Gustafson, E.J.; Serra-Diaz, J.M.; Thompson, J.R. How disturbance, competition, and dispersal interact to prevent tree range boundaries from keeping pace with climate change. Glob. Chang. Biol. 2018, 24, e335–e351. [Google Scholar] [CrossRef]
- Sun, H.; Zhou, Z.; Yu, H. The vegetation of the Big Bend Gorge of Yalu Tsangpo River, SE Tibet, E. Himalayas. Acta Bot. Yunnanica 1997, 19, 57–66. [Google Scholar]
- Xiang, X.; Zhang, Z.; Wang, Z.; Zhang, X.; Wu, G. Transcriptome sequencing and development of EST-SSR markers in Pinus dabeshanensis, an endangered conifer endemic to China. Mol. Breed. 2015, 35, 158. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, H.; Chen, W.; Pang, X.; Li, Y. Genetic diversity and structure of native and non-native populations of the endangered plant Pinus dabeshanensis. Genet. Mol. Res. 2003, 15, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fordham, D.A.; Resit Akçakaya, H.; Araújo, M.B.; Elith, J.; Keith, D.A.; Pearson, R.; Auld, T.D.; Mellin, C.; Morgan, J.W.; Regan, T.J. Plant extinction risk under climate change: Are forecast range shifts alone a good indicator of species vulnerability to global warming? Glob. Chang. Biol. 2012, 18, 1357–1371. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
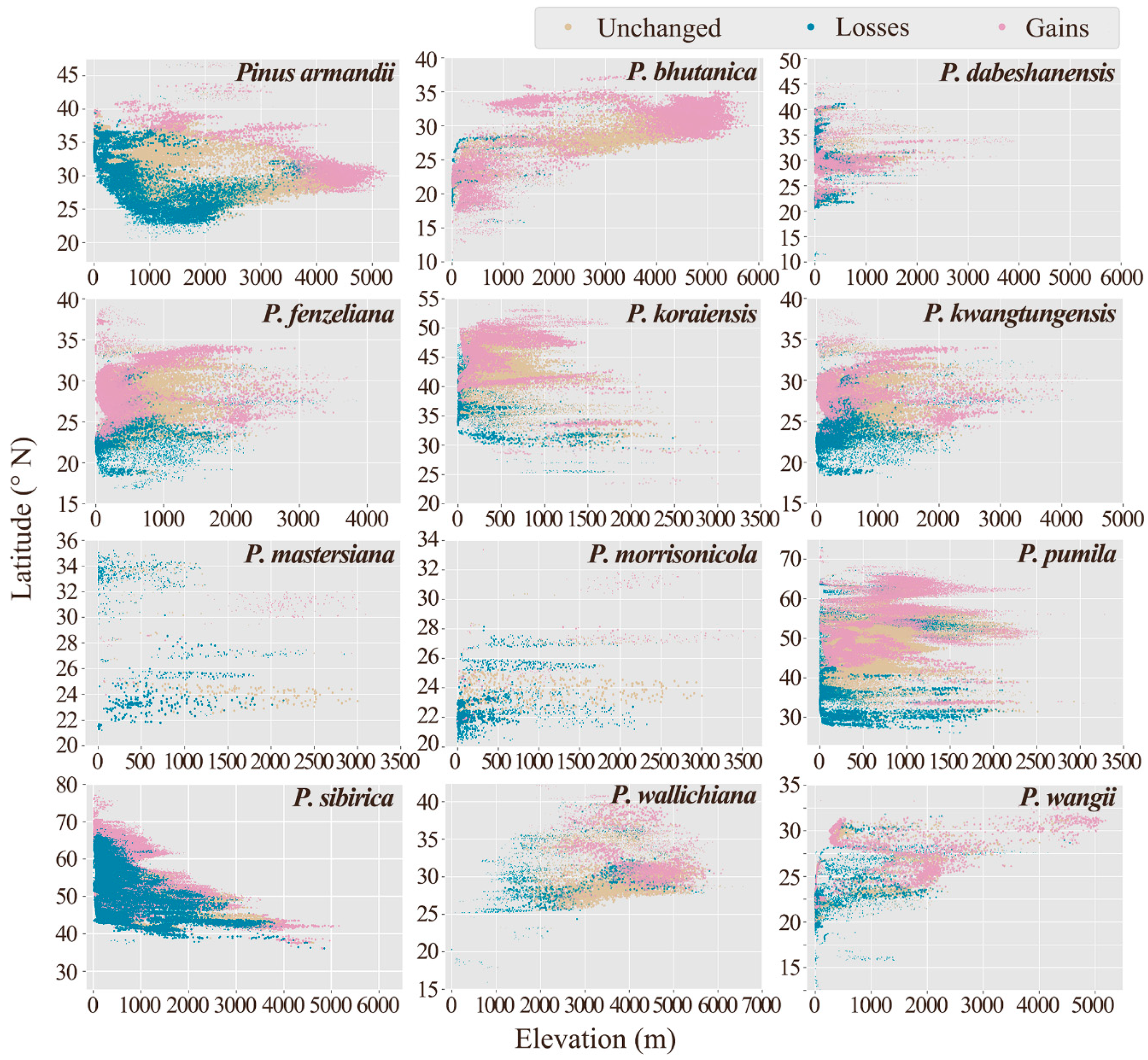{kind=link}
| Environmental Variable | Ecological Meaning | Unit | Data Source |
|---|---|---|---|
| Bio1 | Annual mean temperature | °C × 10 | WorldClim |
| Bio2 | mean diurnal range (mean of monthly (max temp—min temp)) | °C × 10 | WorldClim |
| Bio3 | Isothermality (Bio2/(max temperature of warmest month—min temperature of coldest month)) (×100) | ×100 | WorldClim |
| Bio4 | Temperature seasonality (standard deviation ×100) | ×100 | WorldClim |
| Bio6 | Min temperature of coldest month | °C × 10 | WorldClim |
| Bio8 | Mean temperature of wettest quarter | °C × 10 | WorldClim |
| Bio9 | Mean temperature of driest quarter | °C × 10 | WorldClim |
| Bio12 | Annual precipitation | mm | WorldClim |
| Bio13 | Precipitation of wettest month | mm | WorldClim |
| Bio16 | Precipitation of wettest quarter | mm | WorldClim |
| Bio18 | Precipitation of warmest quarter | mm | WorldClim |
| Bio19 | Precipitation of coldest quarter | mm | WorldClim |
| FOR | Forest land | % | FAO |
| GRS | Grass/scrub/woodland | % | FAO |
| NVG | Barren/very sparsely vegetated land | % | FAO |
| UVB1 | Annual mean ultraviolet B (UVB) | J·m−2·day−1 | glUV |
| UVB3 | Mean UVB of highest month | J·m−2·day−1 | glUV |
| WAT | Mapped water Bodies | % | FAO |
| Species | AUC | TSS |
|---|---|---|
| Pinus armandii | 0.978 | 0.954 |
| P. bhutanica | 0.999 | 0.997 |
| P. dabeshanensis | 0.997 | 0.995 |
| P. fenzeliana | 0.920 | 0.742 |
| P. koraiensis | 0.985 | 0.954 |
| P. kwangtungensis | 0.933 | 0.867 |
| P. mastersiana | 0.986 | 0.971 |
| P. morrisonicola | 0.986 | 0.971 |
| P. pumila | 0.921 | 0.746 |
| P. sibirica | 0.904 | 0.776 |
| P. wallichiana | 0.917 | 0.833 |
| P. wangii | 0.997 | 0.994 |
| Species | Bio1 | Bio2 | Bio3 | Bio4 | Bio6 | Bio9 | Bio12 | Bio13 | Bio16 | Bio18 | Bio19 | UVB1 | UVB3 | FOR | GRS | NVG | Sum | |
| Pinus armandii | PC 2 | 8.9 | 15.9 | 28.0 | 22.8 | 75.6 | ||||||||||||
| PI 3 | 22.4 | 5.6 | 58.1 | 2.8 | 88.9 | |||||||||||||
| JR 4 | Ⅲ | Ⅰ | Ⅱ | Ⅳ | ||||||||||||||
| P. bhutanica | PC | 6.2 | 29.9 | 41.2 | 8.8 | 86.1 | ||||||||||||
| PI | 1.1 | 6.9 | 29.4 | 44.8 | 82.2 | |||||||||||||
| JR | Ⅳ | Ⅰ | Ⅲ | Ⅱ | ||||||||||||||
| P. dabeshanensis | PC | 9.4 | 29.5 | 22.0 | 12.8 | 73.7 | ||||||||||||
| PI | 39.2 | 25.3 | 21.5 | 7.4 | 93.4 | |||||||||||||
| JR | Ⅲ | Ⅳ | Ⅰ | Ⅱ | ||||||||||||||
| P. fenzeliana | PC | 15.8 | 11.7 | 50.2 | 5.5 | 83.2 | ||||||||||||
| PI | 56.9 | 3.1 | 25.3 | 3.3 | 88.6 | |||||||||||||
| JR | Ⅳ | Ⅱ | Ⅰ | Ⅲ | ||||||||||||||
| P. koraiensis | PC | 23 | 50.6 | 6.1 | 7.7 | 87.4 | ||||||||||||
| PI | 12.1 | 14.6 | 36.5 | 16.9 | 80.1 | |||||||||||||
| JR | Ⅳ | Ⅰ | Ⅱ | Ⅲ | ||||||||||||||
| P. kwangtungensis | PC | 11.8 | 17.9 | 8.0 | 47.5 | 85.2 | ||||||||||||
| PI | 48.0 | 18.5 | 8.0 | 15.4 | 89.9 | |||||||||||||
| JR | Ⅳ | Ⅱ | Ⅰ | Ⅲ | ||||||||||||||
| P. mastersiana | PC | 7.2 | 68.8 | 8.3 | 5.7 | 90.0 | ||||||||||||
| PI | 27.2 | 42.2 | 2.1 | 6.9 | 78.4 | |||||||||||||
| JR | Ⅱ | Ⅳ | Ⅲ | Ⅰ | ||||||||||||||
| P. morrisonicola | PC | 3.2 | 9.6 | 73.7 | 8.2 | 94.7 | ||||||||||||
| PI | 69.7 | 24.5 | 2.2 | 1.4 | 97.8 | |||||||||||||
| JR | Ⅳ | Ⅲ | Ⅱ | Ⅰ | ||||||||||||||
| P. pumila | PC | 27.9 | 6.7 | 32.7 | 5.6 | 72.9 | ||||||||||||
| PI | 7.7 | 26.1 | 40.7 | 6.1 | 80.6 | |||||||||||||
| JR | Ⅱ | Ⅲ | Ⅳ | Ⅰ | ||||||||||||||
| P. sibirica | PC | 26.0 | 9.8 | 17.5 | 21.3 | 74.6 | ||||||||||||
| PI | 33.3 | 19.6 | 19.2 | 6.5 | 78.6 | |||||||||||||
| JR | Ⅰ | Ⅳ | Ⅲ | Ⅱ | ||||||||||||||
| P. wallichiana | PC | 9.2 | 11.0 | 47.8 | 13.9 | 81.9 | ||||||||||||
| PI | 8.5 | 3.6 | 46.1 | 30.0 | 88.2 | |||||||||||||
| JR | Ⅳ | Ⅱ | Ⅲ | Ⅰ | ||||||||||||||
| P. wangii | PC | 11.9 | 29.2 | 8.9 | 9.7 | 59.7 | ||||||||||||
| PI | 21.7 | 16.3 | 12.5 | 22.2 | 72.7 | |||||||||||||
| JR | Ⅳ | Ⅲ | Ⅰ | Ⅱ |
| Species | LIG 1 | LGM 2 | Middle Holocene | Current | 3 RCP2.6-2050 | RCP2.6-2070 | RCP8.5-2050 | RCP8.5-2070 |
|---|---|---|---|---|---|---|---|---|
| Pinus armandii | 1,602,309 | 1,573,767 | 1,537,343 | 1,419,288 | 1,211,267 | 1,194,878 | 1,171,996 | 1,174,323 |
| P. bhutanica | 671,962 | 305,382 | 358,941 | 313,594 | 290,312 | 341,180 | 409,861 | 855,833 |
| P. dabeshanensis | 2,827,499 | 214,861 | 679,427 | 321,788 | 361,667 | 224,635 | 603,663 | 343,420 |
| P. fenzeliana | 544,757 | 270,364 | 940,659 | 814,809 | 882,309 | 1,059,409 | 923,541 | 1,116,198 |
| P. koraiensis | 626,996 | 205,833 | 820,139 | 417,257 | 528,854 | 605,434 | 629,930 | 710,104 |
| P. kwangtungensis | 767,725 | 203,646 | 1,101,770 | 878,107 | 892,812 | 943,802 | 969,809 | 919,878 |
| P. mastersiana | 13,212 | 3767 | 16,441 | 20,903 | 13,490 | 16,042 | 12,760 | 12,222 |
| P. morrisonicola | 21,389 | 3767 | 46,354 | 54,306 | 56,372 | 42,118 | 35,486 | 32,066 |
| P. pumila | 830,642 | 758,993 | 1,369,757 | 878,524 | 979,444 | 959,705 | 949,496 | 975,416 |
| P. sibirica | 318,229 | 845,121 | 364,184 | 347,743 | 448,055 | 473,351 | 296,337 | 417,187 |
| P. wallichiana | 314,271 | 288,472 | 309,323 | 316,753 | 326,649 | 429,444 | 450,156 | 392,292 |
| P. wangii | 86,198 | 239,392 | 140,382 | 189,010 | 166,302 | 408,368 | 288,750 | 271,910 |
| Species | Suitability ranks | Current | 1 RCP2.6-2050 | RCP2.6-2070 | RCP8.5-2050 | RCP8.5-2070 |
|---|---|---|---|---|---|---|
| Pinus armandii | Not | 5.72 | 13.68 | 11.44 | 15.42 | 19.40 |
| Low | 12.44 | 17.16 | 16.92 | 18.41 | 20.15 | |
| Medium | 19.90 | 17.91 | 20.65 | 23.63 | 20.15 | |
| High | 61.94 | 51.24 | 51.00 | 42.54 | 40.30 | |
| P. bhutanica | Not | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Low | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Medium | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| High | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | |
| P. dabeshanensis | Not | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Low | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Medium | 20.00 | 20.00 | 20.00 | 0.00 | 10.00 | |
| High | 80.00 | 80.00 | 80.00 | 100.00 | 90.00 | |
| P. fenzeliana | Not | 3.85 | 0.00 | 0.00 | 3.85 | 3.85 |
| Low | 19.23 | 26.92 | 15.38 | 19.23 | 19.23 | |
| Medium | 15.38 | 7.69 | 19.23 | 15.38 | 15.38 | |
| High | 61.54 | 65.38 | 65.38 | 61.54 | 61.54 | |
| P. koraiensis | Not | 6.25 | 0.00 | 0.00 | 0.00 | 0.00 |
| Low | 37.50 | 18.75 | 0.00 | 0.00 | 6.25 | |
| Medium | 12.50 | 31.25 | 18.75 | 31.25 | 6.25 | |
| High | 43.75 | 50.00 | 81.25 | 68.75 | 87.50 | |
| P. kwangtungensis | Not | 1.61 | 1.61 | 1.61 | 1.61 | 9.68 |
| Low | 6.45 | 9.68 | 4.84 | 4.84 | 8.06 | |
| Medium | 16.13 | 12.90 | 19.35 | 16.13 | 6.45 | |
| High | 75.81 | 75.81 | 74.19 | 77.42 | 75.81 | |
| P. mastersiana | Not | 0.98 | 0.98 | 0.98 | 2.94 | 1.96 |
| Low | 1.96 | 4.90 | 4.90 | 11.76 | 15.69 | |
| Medium | 9.80 | 14.71 | 12.75 | 24.51 | 4.90 | |
| High | 87.25 | 79.41 | 81.37 | 60.78 | 77.45 | |
| P. morrisonicola | Not | 0.00 | 0.00 | 1.41 | 1.41 | 1.41 |
| Low | 1.41 | 4.23 | 1.41 | 0.00 | 7.04 | |
| Medium | 15.49 | 14.08 | 9.86 | 15.49 | 19.72 | |
| High | 83.10 | 81.69 | 87.32 | 83.10 | 71.83 | |
| P. pumila | Not | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Low | 3.85 | 11.54 | 11.54 | 15.38 | 19.23 | |
| Medium | 23.08 | 19.23 | 19.23 | 30.77 | 7.69 | |
| High | 73.08 | 69.23 | 69.23 | 53.85 | 73.08 | |
| P. sibirica | Not | 42.86 | 57.14 | 42.86 | 42.86 | 57.14 |
| Low | 0.00 | 14.29 | 0.00 | 28.57 | 14.29 | |
| Medium | 14.29 | 14.29 | 14.29 | 28.57 | 0.00 | |
| High | 42.86 | 14.29 | 42.86 | 0.00 | 28.57 | |
| P. wallichiana | Not | 5.95 | 13.10 | 8.33 | 14.29 | 11.90 |
| Low | 8.33 | 2.38 | 9.52 | 4.76 | 11.90 | |
| Medium | 13.10 | 7.14 | 8.33 | 16.67 | 8.33 | |
| High | 72.62 | 77.38 | 73.81 | 64.29 | 67.86 | |
| P. wangii | Not | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Low | 0.00 | 0.00 | 0.00 | 50.00 | 25.00 | |
| Medium | 0.00 | 50.00 | 25.00 | 25.00 | 50.00 | |
| High | 100.00 | 50.00 | 75.00 | 25.00 | 25.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, L.; He, J.; Xie, L.; Cui, G. Prediction of the Suitable Area of the Chinese White Pines (Pinus subsect. Strobus) under Climate Changes and Implications for Their Conservation. Forests 2020, 11, 996. https://doi.org/10.3390/f11090996
Lin L, He J, Xie L, Cui G. Prediction of the Suitable Area of the Chinese White Pines (Pinus subsect. Strobus) under Climate Changes and Implications for Their Conservation. Forests. 2020; 11(9):996. https://doi.org/10.3390/f11090996
Chicago/Turabian StyleLin, Lele, Jian He, Lei Xie, and Guofa Cui. 2020. "Prediction of the Suitable Area of the Chinese White Pines (Pinus subsect. Strobus) under Climate Changes and Implications for Their Conservation" Forests 11, no. 9: 996. https://doi.org/10.3390/f11090996
APA StyleLin, L., He, J., Xie, L., & Cui, G. (2020). Prediction of the Suitable Area of the Chinese White Pines (Pinus subsect. Strobus) under Climate Changes and Implications for Their Conservation. Forests, 11(9), 996. https://doi.org/10.3390/f11090996