Effect of Silviculture on Carbon Pools during Development of a Ponderosa Pine Plantation


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Original Design
2.2. Aboveground Carbon for Planted Ponderosa Pine
2.3. Competing Vegetation Carbon
2.4. Forest Floor and Soil Carbon
2.5. Statistical Analysis
3. Results
3.1. 28-Year-Old Stand Carbon
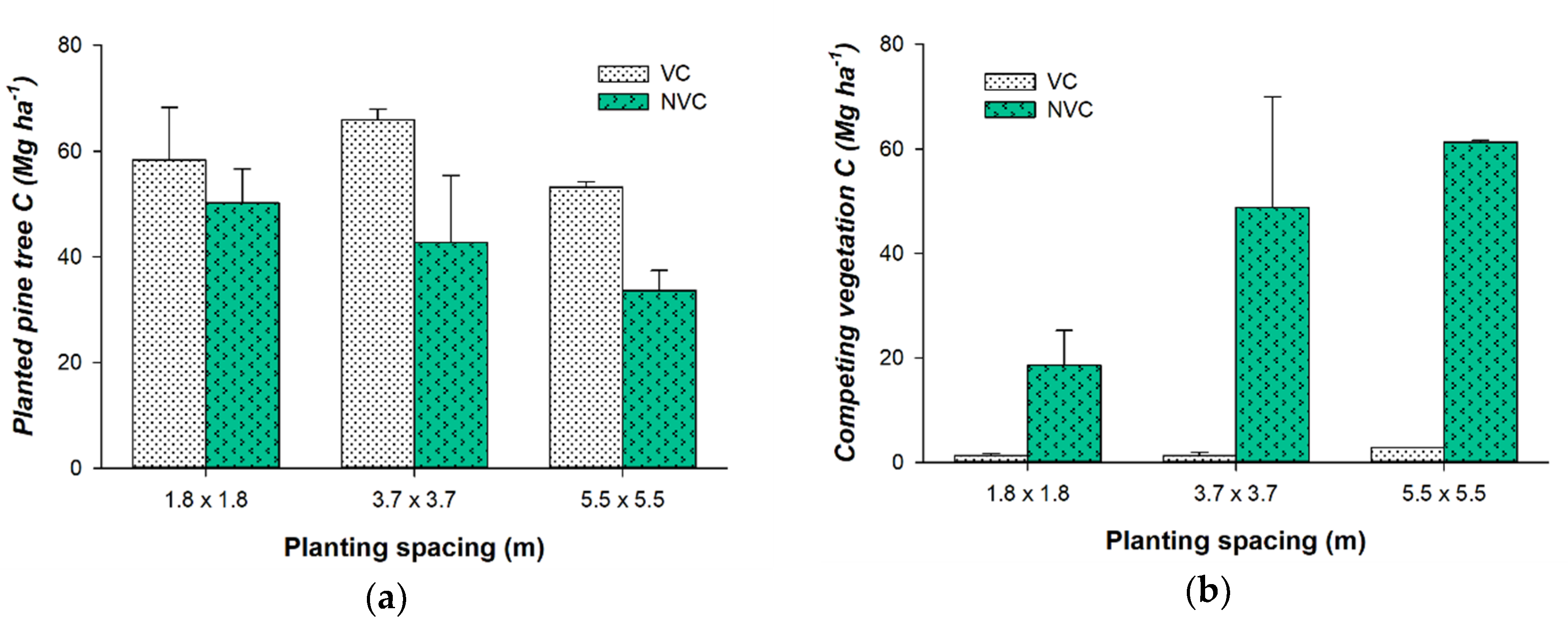
3.1.1. Aboveground Vegetation Carbon
3.1.2. Forest Floor Carbon
3.1.3. Soil Carbon
3.1.4. Summary at Age 28
3.2. 54-Year-Old Stand Carbon
3.2.1. Aboveground Vegetation Carbon
3.2.2. Forest Floor Carbon
3.2.3. Soil Carbon
3.2.4. Summary at Age 54
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef]
- Millar, C.; Stephenson, N. Temperate forest health in an ear of emerging megadisturbance. Science 2015, 349, 823–826. [Google Scholar] [CrossRef]
- Zhang, J.W.; Finley, K.A.; Knapp, E.E. Resilience of a ponderosa pine plantation to a backfiring operation during a mid-summer wildfire. Int. J. Wildland Fire 2019, 28. [Google Scholar] [CrossRef]
- Zhang, J.W.; Powers, R.F.; Skinner, C.N. To Manage or Not to Manage: The Role of Silviculture in Sequestering Carbon in the Specter of Climate Change. In Integrated Management of Carbon Sequestration and Biomass Utilization Opportunities in a Changing Climate, Proceedings of the 2009 National Silviculture Workshop, Boise, ID, USA, 15–18 June 2009; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2009. [Google Scholar]
- Stevens-Rumann, C.S.; Morgan, P. Tree regeneration following wildfires in the western US: A review. Fire Ecol. 2019, 15. [Google Scholar] [CrossRef]
- McDonald, P.M.; Fiddler, G.O. Twenty-Five Years of Managing Vegetation in Conifer Plantations in Northern and Central California: Results, Application, Principles, and challenges; General Technical Report PSW-GTR-231; U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2010; 87p.
- Zhang, J.W.; Powers, R.F.; Oliver, W.W.; Young, D.H. Response of ponderosa pine plantations to competing vegetation control in Northern California, USA: A meta- analysis. Forestry 2013, 86, 3–11. [Google Scholar] [CrossRef]
- Oliver, W.W. Spacing and shrub competition influence 20-year-old development of planted ponderosa pine. West. J. Appl. For. 1990, 5, 79–82. [Google Scholar] [CrossRef]
- McDonald, P.M.; Powers, R.F. Vegetation Trends and Carbon Balance in a Ponderosa Pine Plantation: Long-Term Effects of Different Shrub Densities. In Proceedings of the 24th Annual Forest Vegetation Management Conference, University of California Cooperative Extension (UCCE), Redding, CA, USA, 14–16 January 2003; UCCE: Redding, CA, USA, 2003. [Google Scholar]
- Wagner, R.G. Competition and critical-period thresholds for vegetation management decisions in young conifer stands. For. Chron. 2000, 76, 961–968. [Google Scholar] [CrossRef]
- Zhang, J.W.; Oliver, W.W.; Ritchie, M.W.; Neal, D.L. Overstory and Understory Dynamics in a Ponderosa Pine Plantation Vary with Stand Density in the Sierra Nevada: 40-Year Results. For. Sci. 2013, 59, 670–680. [Google Scholar] [CrossRef]
- Zhang, J.W.; Busse, M.D.; Fiddler, G.O.; Fredrickson, E. Thirteen-year growth response of ponderosa pine plantations to dominant shrubs (Arctostaphylos and Ceanothus). J. For. Res. 2019. [Google Scholar] [CrossRef]
- Justine, M.F.; Yang, W.; Wu, F.; Khan, M.N. Dynamics of biomass and carbon sequestration across a chronosequence of masson pine plantations. J. Geophys. Res. Biogeosci. 2017, 122, 578–591. [Google Scholar] [CrossRef]
- Powers, R.F.; Busse, M.D.; McFarlane, K.J.; Zhang, J.; Young, D.H. Long-term effects of silviculture on soil carbon storage: Does vegetation control make a difference? Forestry 2013, 86, 47–58. [Google Scholar] [CrossRef]
- Peri, P.L.; Gargaglione, V.; Martínez Pastur, G.; Lencinas, M.V. Carbon accumulation along a stand development sequence of Nothofagus antarctica forests across a gradient in site quality in Southern Patagonia. For. Ecol. Manag. 2010, 260, 229–237. [Google Scholar] [CrossRef]
- Zhang, J.W.; Oliver, W.W.; Busse, M.D. Growth and development of ponderosa pine on sites of contrasting productivities: Relative importance of stand density and shrub competition effects. Can. J. For. Res. 2006, 36, 2426–2438. [Google Scholar] [CrossRef]
- Cairns, M.A.; Brown, S.; Helmer, E.H.; Baumgardner, G.A. Root biomass allocation in the world’s upland forests. Oecologia 1997, 111, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Snell, J.A.K.; Little, S.N. Predicting Crown Weight and Bole Volume of Five Western Hardwoods; General Technical Report PNW-151; U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1983; 42p.
- Jenkins, J.C.; Chojnacky, D.C.; Heath, L.S.; Birdsey, R.A. National-scale biomass estimators for United States tree species. For. Sci. 2003, 49, 12–35. [Google Scholar]
- Wiant, H.V. Tables and Procedures for Estimating Weights of Some Appalachian Hardwoods; West. Virginia Univeristy, Agricultural Experimental Station: Morgantown, WV, USA, 1977. [Google Scholar]
- Van Wagner, C.E. The line-intersect method in forest fuel sampling. For. Sci. 1968, 14, 20–26. [Google Scholar]
- Brown, J.K.; Oberheu, R.D.; Johnston, C.M. Handbook for Inventorying Surface Fuels and Biomass in the Interior West; General Technical Report INT-129; U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1981; 48p.
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon and organic matter. In Methods of Soil Analysis Part. 3–Chemical Methods; Sparks, D.L., Ed.; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1996. [Google Scholar]
- Zhang, J.W.; Busse, M.D.; Young, D.H.; Fiddler, G.O.; Sherlock, J.W.; TenPas, J.D. Aboveground biomass responses to organic matter removal, soil compaction, and competing vegetation control on 20-year mixed conifer plantations in California. For. Ecol. Manag. 2017, 401, 341–353. [Google Scholar] [CrossRef]
- Yang, Y.; Luo, Y.; Finzi, A.C. Carbon and nitrogen dynamics during forest stand development: A global synthesis. New Phytol. 2011, 190, 977–989. [Google Scholar] [CrossRef]
- Zhang, J.W.; Young, D.H.; Oliver, W.W.; Fiddler, G.O. Effect of overstorey trees on understorey vegetation in California (USA) ponderosa pine plantations. Forestry 2016, 89, 91–99. [Google Scholar] [CrossRef]
- Odum, E.P. The strategy of ecosystem development. Science 1969, 164, 262–270. [Google Scholar] [CrossRef]
- McFarlance, K.J.; Schoenholtz, S.H.; Powers, R.F. Plantation management intensity affects belowground carbon and nitrogen storage in Northern California. Soil Sci. Soc. Am. J. 2009, 73, 1020–1032. [Google Scholar] [CrossRef]
- Zhang, J.W.; Webster, J.; Young, D.H.; Fiddler, G.O. Effect of thinning and soil treatments on Pinus ponderosa plantations: 15-year results. For. Ecol. Manag. 2016, 368, 123–132. [Google Scholar] [CrossRef]
- Powers, R.F.; Ferrell, G.T. Moisture, nutrient, and insect constraints on plantation growth: The “Garden of Eden” study. N. Z. J. For. Sci. 1996, 26, 126–144. [Google Scholar]
- Campbell, J.; Alberti, G.; Martin, J.; Law, B.E. Carbon dynamics of a ponderosa pine plantation following a thinning treatment in the northern Sierra Nevada. For. Ecol. Manag. 2009, 257, 453–463. [Google Scholar] [CrossRef]
- Busse, M.D.; Zhang, J.W.; Fiddler, G.O.; Young, D.H. Compaction and organic matter retention in mixed-conifer forests of California: 20-year effects on soil physical and chemical health. For. Ecol. Manag. under review.
- Bridgwater, F.E.; William, C.G.; Campbell, R.G. Patterns of leader elongation in loblolly pine families. For. Sci. 1985, 31, 933–944. [Google Scholar]
- Zhang, J.W.; Young, D.H.; Luckow, K.R. Effect of redistributing windrowed topsoil on growth and development of ponderosa pine plantations. For. Ecol. Manag. 2015, 353, 148–155. [Google Scholar] [CrossRef]
- Stephens, S.L.; McIver, J.D.; Boerner, R.E.; Fettig, C.J.; Fontaine, J.B.; Hartsough, B.R.; Kennedy, P.L.; Schwilk, D.W. The effects of forest fuel-reduction treatments in the United States. BioScience 2012, 62, 549–560. [Google Scholar] [CrossRef]
- Oliver, W.W.; Ryker, R.A. Pinus ponderosa Dougl. ex Laws. Ponderosa pine. Silv. N. Am. 1990, 1, 413–424. [Google Scholar]
- Oren, R.; Waring, R.H.; Stafford, S.G.; Barrett, J.W. Twenty-four years of ponderosa pine growth in relation to canopy leaf area and understory competition. For. Sci. 1987, 33, 538–547. [Google Scholar]
- Stephens, S.L.; Collins, B.M.; Fettig, C.J.; Finney, M.A.; Hoffman, C.M.; Knapp, E.E.; North, M.P.; Safford, H.D.; Wayman, R.B. Drought, tree mortality, and wildfire in forests adapted to frequent fire. BioScience 2018, 68, 77–88. [Google Scholar] [CrossRef]
- Zhang, J.W.; Finley, K.A.; Johnson, N.G.; Ritchie, M.W. Lowering stand density enhances resiliency of ponderosa pine forests to disturbances and climate change. For. Sci. 2019, 65, 496–507. [Google Scholar] [CrossRef]
- Finley, K.; Zhang, J. Climate effect on ponderosa pine radial growth varies with tree density and shrub removal. Forests 2019, 10, 477. [Google Scholar] [CrossRef]
- Zhang, J.W.; Ritchie, M.W.; Maguire, D.A.; Oliver, W.W. Thinning ponderosa pine (Pinus ponderosa) stands reduces mortality while maintaining stand productivity. Can. J. For. Res. 2013, 43, 311–320. [Google Scholar] [CrossRef]
- Skog, K.E. Sequestration of carbon in harvested wood products for the United States. For. Prod. J. 2008, 58, 56–72. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Competing Vegetation Treatment | Stand Age | Overstory Planting Spacings | TPH 1 | Height (m) | QMD 2 (cm) | BA 3 (m2 ha−1) |
|---|---|---|---|---|---|---|
| Vegetation Control | Age 28 | 1.8 × 1.8 m | 2621 | 10.2 | 13.7 | 39 |
| 3.7 × 3.7 m | 747 | 14.2 | 24.6 | 35.6 | ||
| 5.5 × 5.5 m | 318 | 15.2 | 32.2 | 25.8 | ||
| Age 54 | 1.8 × 1.8 m | 1248 | 19.9 | 20.9 | 42.4 | |
| 3.7 × 3.7 m | 622 | 23.7 | 32.6 | 52.2 | ||
| 5.5 × 5.5 m | 263 | 26.8 | 45.8 | 43.4 | ||
| No vegetation Control | Age 28 | 1.8 × 1.8 m | 2746 | 9.7 | 12.6 | 34.3 |
| 3.7 × 3.7 m | 747 | 11.3 | 20.2 | 24.4 | ||
| 5.5 × 5.5 m | 332 | 13.3 | 26 | 17.6 | ||
| Age 54 | 1.8 × 1.8 m | 1123 | 19.5 | 21.8 | 43.7 | |
| 3.7 × 3.7 m | 622 | 19.9 | 30.5 | 46.7 | ||
| 5.5 × 5.5 m | 318 | 23.1 | 39.1 | 38.2 |
| Understory Treatment | Layer 1 | Overstory Planting Spacings | ||
|---|---|---|---|---|
| 1.8 × 1.8 m | 3.7 × 3.7 m | 5.5 × 5.5 m | ||
| Vegetation control | Woody debris | 1.4 | 0.4 | 0.3 |
| O horizon | 16.7 | 13.7 | 13.8 | |
| FF Total | 18.1 | 14.1 | 14.2 | |
| Soil (0–10 cm) | 32.5 | 42.1 | 38 | |
| Soil (10–20 cm) | 40.4 | 49.1 | 51.2 | |
| Soil Total | 72.9 | 91.2 | 89.2 | |
| Grand Total | 91.1 | 105.3 | 103.2 | |
| No vegetation control | Woody debris | 1 | 1.3 | 0.6 |
| O horizon | 18.1 | 19.4 | 23.4 | |
| FF Total | 19.1 | 20.7 | 23.7 | |
| Soil (0–10 cm) | 38.8 | 42.3 | 36.9 | |
| Soil (10–20 cm) | 53.4 | 54.3 | 31.1 | |
| Soil Total | 92.2 | 96.6 | 68 | |
| Grand Total | 111.3 | 117.3 | 91.7 | |
| Understory Treatment | Layer 1 | Overstory Planting Spacings | ||
|---|---|---|---|---|
| 1.8 × 1.8 m | 3.7 × 3.7 m | 5.5 × 5.5 m | ||
| Vegetation control | Woody debris | 0.7 | 0.5 | 0.2 |
| O horizon | 32.4 | 23.3 | 10.1 | |
| FF Total | 33.1 | 23.8 | 10.3 | |
| Soil (0–10 cm) | 45.8 | 44.6 | 58.2 | |
| Soil (10–20 cm) | 32.9 | 28.9 | 31.6 | |
| Soil Total | 78.7 | 73.5 | 89.8 | |
| Grand Total | 111.8 | 97.3 | 100.1 | |
| No vegetation control | Woody debris | 0.9 | 1.2 | 1.6 |
| O Horizon | 19.8 | 23.6 | 18.4 | |
| FF Total | 20.7 | 24.8 | 20 | |
| Soil (0–10 cm) | 55.7 | 45.3 | 49.6 | |
| Soil (10–20 cm) | 41.4 | 36.4 | 38 | |
| Soil Total | 97.1 | 81.7 | 87.6 | |
| Grand Total | 117.8 | 106.5 | 107.6 | |
| Age | Understory Treatment | Overstory Planting Spacings | |||
|---|---|---|---|---|---|
| 1.8 × 1.8 m | 3.7 × 3.7 m | 5.5 × 5.5 m | Means | ||
| 28 | VC | 2.7 | 2.8 | 2.5 | 2.7 |
| NVC | 3.1 | 3.9 | 4.1 | 3.7 | |
| Means | 2.9 | 3.4 | 3.3 | 3.2 | |
| 54 | VC | 1.8 | 2.1 | 1.7 | 1.9 |
| NVC | 1.4 | 0.8 | 1.0 | 1.1 | |
| Means | 1.6 | 1.5 | 1.4 | 1.5 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Zhang, J.; Mattson, K.; Finley, K. Effect of Silviculture on Carbon Pools during Development of a Ponderosa Pine Plantation. Forests 2020, 11, 997. https://doi.org/10.3390/f11090997
Zhang J, Zhang J, Mattson K, Finley K. Effect of Silviculture on Carbon Pools during Development of a Ponderosa Pine Plantation. Forests. 2020; 11(9):997. https://doi.org/10.3390/f11090997
Chicago/Turabian StyleZhang, Jie, Jianwei Zhang, Kim Mattson, and Kaelyn Finley. 2020. "Effect of Silviculture on Carbon Pools during Development of a Ponderosa Pine Plantation" Forests 11, no. 9: 997. https://doi.org/10.3390/f11090997
APA StyleZhang, J., Zhang, J., Mattson, K., & Finley, K. (2020). Effect of Silviculture on Carbon Pools during Development of a Ponderosa Pine Plantation. Forests, 11(9), 997. https://doi.org/10.3390/f11090997