Picea abies–Armillaria–Ips: A Strategy or Coincidence?


{kind=link}
Abstract
1. Introduction
2. Hypothesis
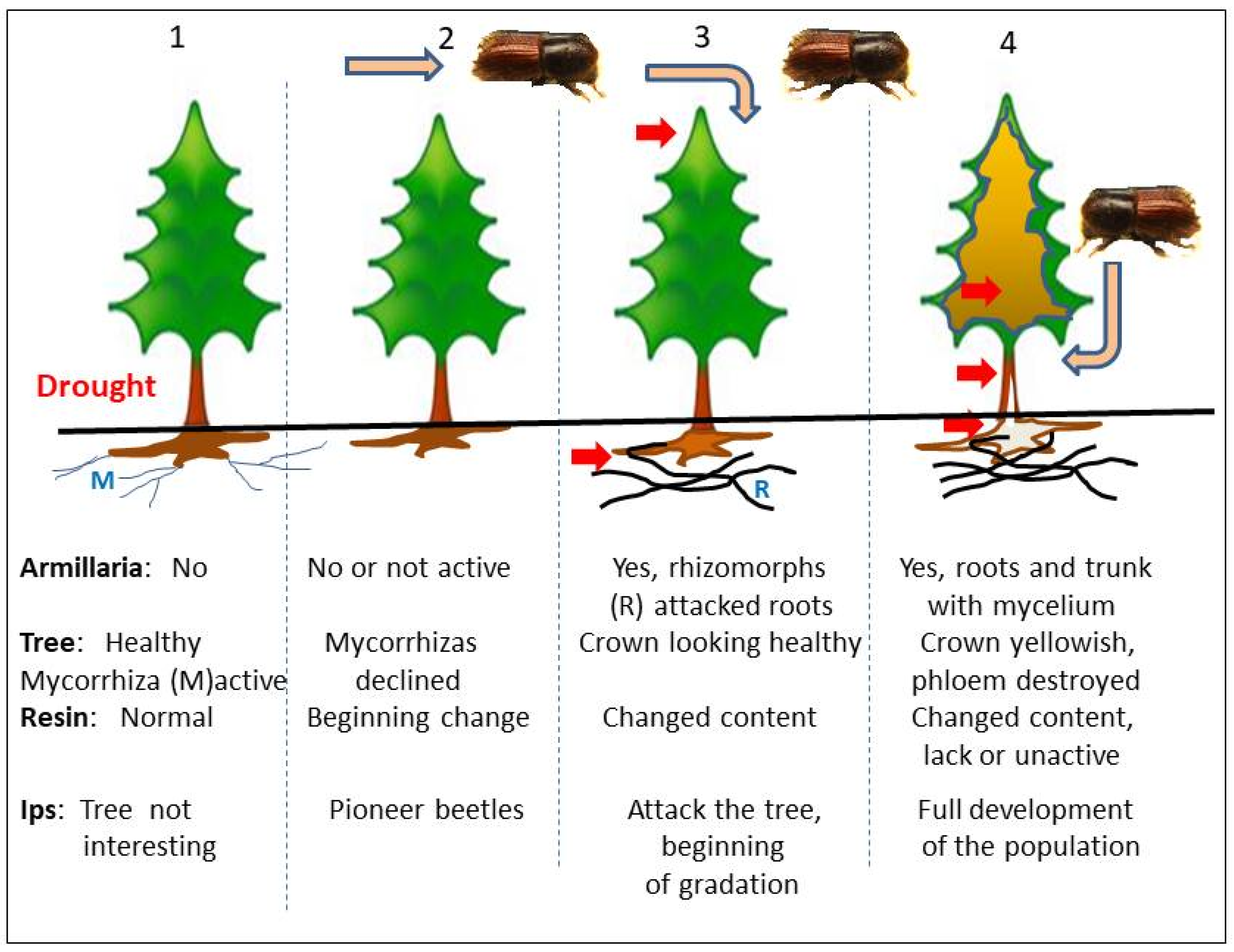
3. Fungal Infection and Pest Invasion
4. Final Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mattheck, M.; Bethge, C. The mechanical survival strategy of trees. Arbor. J. 1998, 22, 369–386. [Google Scholar] [CrossRef]
- Gadagkar, R. Survival Strategies. Cooperation and Conflict in Animal Societies; Harvard University Press: Massachusetts, MA, USA, 2001. [Google Scholar]
- Casadevall, A. Determinants of virulence in the pathogenic fungi. Fungal. Biol. Rev. 2007, 21, 130–132. [Google Scholar] [CrossRef]
- Merrill, W. Mechanisms of Resistance to Fungi in Woody Plants: A Historical Perspective. In Defense Mechanisms of Woody Plants Against Fungi, Springer Series in Wood Science; Blanchette, R.A., Biggs, A.R., Eds.; Springer: Berlin/Heidelberg, Germany, 1992. [Google Scholar]
- Heitman, J.; Sun, S.; James, T.Y. Evolution of fungal sexual reproduction. Mycologia 2013, 105, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.; Stenlid, J. Infection of roots of Norway spruce (Picea abies) by Heterobasidion annosum. Eur. J. For. Path. 1985, 15, 32–45. [Google Scholar] [CrossRef]
- Robinson, R.M.; Sturrock, R.N.; Davidson, J.J. Detection of a chitinase-like protein in the roots of Douglas-fir trees infected with Armillaria ostoyae and Phellinus weirii. Tree Phys. 2000, 20, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Prospero, S.; Holdenrieder, O.; Rigling, D. Comparison of the virulence of Armillaria cepistipes and Armillaria ostoyae on four Norway spruce provenances. For. Pathol. 2004, 34, 1–14. [Google Scholar] [CrossRef]
- Holuša, J.; Lubojacký, J.; Čurn, V.; Tonka, T.; Lukášová, K.; Horák, J. Combined effects of drought stress and Armillaria infection on tree mortality in Norway spruce plantations. For. Ecol. Manag. 2018, 427, 434–445. [Google Scholar] [CrossRef]
- Sipos, G.; Prasanna, A.N.; Walter, M.C.; O’Connor, E.; Bálint, B.; Krizsán, K.; Kiss, B.; Hess, J.; Varga, T.; Slot, J.; et al. Genome expansion and lineage-specific genetic innovations in the forest pathogenic fungi. Armillaria. Nat. Ecol. Evol. 2017, 1, 1931–1941. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, B.A.; Dreisbach, T.A.; Parks, C.G.; Filip, G.M.; Schmitt, C.L. Coarse-scale population structure of pathogenic Armillaria species in a mixed-conifer forest in the Blue Mountains of northeast Oregon. Can. J. For. Res. 2003, 33, 612–623. [Google Scholar] [CrossRef]
- Baumgartner, K.; Coetzee, M.P.; Hoffmeister, D. Secrets of the subterranean pathosystem of Armillaria. Mol. Plant. Pathol. 2011, 12, 515–534. [Google Scholar] [CrossRef]
- Morrison, D.J. Rhizomorph growth habit, saprophytic ability and virulence of 15 Armillaria species. For. Pathol. 2004, 34, 15–26. [Google Scholar] [CrossRef]
- Kubiak, K.; Żółciak, A.; Damszel, M.; Lech, P.; Sierota, Z. Armillaria Pathogenesis under Climate Changes. Forests 2017, 8, 100. [Google Scholar] [CrossRef]
- Heizelmann, R.; Dutech, C.; Tsykun, T.; Labbe, F.; Soularule, J.-P. Latest advances and future perspectives in Armillaria research. Can. J. Plant. Path. 2019, 41, 1–23. [Google Scholar] [CrossRef]
- Legrand, P.; Ghahari, S.; Guillaumin, J.J. Occurrence of genets of Armillaria spp. In four mountain forests in Central France: The colonization strategy of Armillaria ostoyae. New Phytol. 1996, 133, 321–332. [Google Scholar] [CrossRef]
- Schelhaas, M.-J.; Nabuurs, G.-J.; Schuck, A. Natural disturbances in the European forests in the 19th and 20th centuries. Glob. Chang. Biol. 2003, 9, 1620–1633. [Google Scholar] [CrossRef]
- Grégoire, J.-C.; Evans, H. Damage and Control of Bawbilt Organisms, An Overview. In Bark and Wood Boring Insects in Living Trees in Europe, a Synthesis; Lieutier, F., Day, K.R., Battisti, A., Grégoire, J.-C., Evans, H., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 2004; pp. 19–37. [Google Scholar]
- Misiek, M.; Hoffmeister, D. Sesquiterpene aryl ester natural products in North American Armillaria species. Mycol. Progress 2012, 11, 7–15. [Google Scholar] [CrossRef]
- Raffa, K.F.; Klepzig, K.D. Tree Defense Mechanisms Against Fungi Associated with Insects. In Defense Mechanisms of Woody Plants Against Fungi; Blanchette, R.A., Biggs, A.R., Eds.; Springer Series in Wood Science: Berlin/Heidelberg, Germany, 1992. [Google Scholar]
- Six, D.L.; Wingfield, M.J. The role of phytopathogenicity in bark beetle–Fungus Symbioses: A challenge to the classic paradigm. Ann. Rev. Ent. 2011, 56, 255–272. [Google Scholar] [CrossRef]
- Lombardero, M.J.; Solla, A.; Ayres, M.P. Pine defenses against the pitch canker disease are modulated by a native insect newly associated with the invasive fungus. For. Ecol. Manag. 2019, 437, 253–262. [Google Scholar] [CrossRef]
- Fettig, C.J.; Hilszczanski, J. Management strategies for bark beetles in conifer forests. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Springer: London, UK, 2015. [Google Scholar]
- Jankovský, L.; Cudlín, P.; Moravec, I. Root decays as a potential predisposition factor of a bark beetle disaster in the Šumava Mts. J. For. Sci. 2003, 49, 125–132. [Google Scholar] [CrossRef]
- Clegg, C.J.; Mackean, D.G. Advanced Biology: Principles and Applications; John Murray: London, UK, 2006. [Google Scholar]
- Eastwood, D.C.; Floudas, D.; Binder, M.; Majcherczyk, A.; Schneider, P.; Aerts, A.; Baker, S.E.; Barry, K.; Bendiksby, M.; Blumentritt, M.; et al. The Plant Cell Wall–Decomposing Machinery Underlies the Functional Diversity of Forest Fungi. Science 2011, 333, 762. [Google Scholar] [CrossRef]
- Cruickshank, M.G.; Lejour, D.; Morrison, D.J. Traumatic resin canals as markers of infection events in Douglas fir roots infected with Armillaria root disease. For. Pathol. 2006, 36, 372–384. [Google Scholar] [CrossRef]
- Christiansen, E.; Waring, R.H.; Berryman, A.A. Resistance of conifers to bark beetle attack: Searching for general relationships. For. Ecol. Manag. 1987, 22, 89–106. [Google Scholar] [CrossRef]
- Byers, J.A. Chemical ecology of bark beetles in a complex olfactory landscape. In Bark and Wood Boring Insects in Living Trees in Europe, a Synthesis; Lieutier, F., Day, K.R., Battisti, A., Grégoire, J.-C., Evans, H., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 2004; pp. 89–134. [Google Scholar]
- Wermelinger, B. Ecology and management of the spruce bark beetle Ips typographus—A review of recent research. For. Ecol. Manag. 2004, 202, 67–82. [Google Scholar] [CrossRef]
- Donnelly, D.M.X.; Abe, F.; Coveney, D.; Fukuda, N.; O’Reilly, J.; Polonsky, J.; Prange, T. Antibacterial sesquiterpene aryl esters from Armillaria mellea. J. Nat. Prod. 1985, 48, 10–16. [Google Scholar] [CrossRef]
- Zhang, Q.-H.; Schlyter, F. Olfactory recognition and behavioral avoidance of angiosperm non host volatiles by conifer-inhabiting bark beetles. Agric. For. Entom. 2004, 6, 1–19. [Google Scholar] [CrossRef]
- Austarå, O.; Bakke, A.; Midtgaard, F. Response in Ips typographus to logging waste odors and synthetic pheromones. J. Appl. Entom. 1986, 101, 194–198. [Google Scholar] [CrossRef]
- Lindelöw, A.; Risberg, B.; Sjodin, K. Attraction during flight of Scolytids and other bark and wood-dwelling beetles to volatiles from fresh and stored spruce wood. Can. J. For. Res. 1992, 22, 224–228. [Google Scholar] [CrossRef]
- Schlyter, F.; Birgersson, G.; Byers, J.A.; Löfqvist, J.; Bergström, G. Field response of spruce bark beetle, Ips typographus, to aggregation pheromone candidates. J. Chem. Ecol. 1987, 13, 701–716. [Google Scholar] [CrossRef]
- Brignolas, F.; Lieutier, F.; Sauvard, D.; Christiansen, E.; Berrymann, A.A. Phenolic predictors for Norway spruce resistance to bark beetle Ips typographus and an associated fungus Ceratocystis polonica. Can. J. For. Res. 1998, 28, 720–728. [Google Scholar] [CrossRef]
- Lieutier, F.; Brignolas, F.; Sauvard, D.; Yart, A.; Galet, C.; Brunet, M.; Van de Sype, H. Intra- and inter-provenance variability in phloem polyphenols of Picea abies (L.) Karst. and relation with resistance to a bark-beetle-associated fungus. Tree Physiol. 2003, 23, 247–256. [Google Scholar] [CrossRef]
- Tommerås, B.A.; Mustaparta, H.; Grégoire, J.-C. Receptor cells in Ips typographus and Dendroctonus micans specific to pheromones of the reciprocal genus. J. Chem. Ecol. 1984, 10, 759–769. [Google Scholar] [CrossRef]
- Schroeder, L.M. Attraction of the bark beetle Tomicus piniperda to Scots pine trees in relation to tree vigor and attack density. Entom. Exp. Applic. 1987, 44, 53–58. [Google Scholar] [CrossRef]
- Madziara-Borusiewicz, K.; Strzelecka, H. Conditions of spruce (Picea excelsa Lk.) infestation by the engraver beetle (Ips typographus L.) in mountains of Poland. Z. Angew. Entom. 1977, 83, 409–415. [Google Scholar] [CrossRef]
- Nebeker, T.E.; Schmitz, R.F.; Tisdale, R.A.; Hobson, K.R. Chemical and nutritional status of dwarf mistletoe, Armillaria root rot, and comandra blister rust infected trees which may influence tree susceptibility to bark beetle attack. Can. J. Bot. 1995, 73, 360–369. [Google Scholar] [CrossRef]
- Isidorov, V.A.; Lech, P.; Żółciak, A.; Rusak, M.; Szczepaniak, L. Gas chromatographic-mass spectrometric investigation of metabolites from the needles and roots of pine seedlings at early stages of pathogenic fungi Armillaria ostoyae attack. Trees 2008, 22, 531–542. [Google Scholar] [CrossRef]
- Dörfer, M.; Gressler, M.; Hoffmeister, D. Diversity and bioactivity of Armillaria sesquiterpene aryl ester natural products. Mycol. Prog. 2019, 18, 1027–1037. [Google Scholar] [CrossRef]
- Longauerová, V.; Leontovyč, R.; Krajmerová, D.; Vakula, J.; Grodzki, W. Fungal pathogens–hidden agents of decline. In Spruce Forests Decline in the Beskids–Hynutie smrekových porastov v Beskydoch; Hlásny, T., Sitková, Z., Eds.; National Forest Centre–Forest Research Institute Zvolen: Zvolen, Slovakia; Czech University of Life Sciences Prague: Prague, Czech Republic; Forestry and Game Management Research Institute Jíloviště–Strnady: Jíloviště, Czech Republic, 2010; pp. 93–105. [Google Scholar]
- Hertert, H.D.; Miller, D.L.; Partridge, A.D. Interaction of bark beetles (Coleoptera: Scolytidae) and root rot pathogens in grand fir in northern Idaho. Can. Entom. 1975, 107, 899–904. [Google Scholar] [CrossRef]
- Christiansen, E.; Huse, K.J. Infestation ability of Ips typographus in Norway spruce, in relation to butt rot, tree vitality, and increment. Medd. NISK 1980, 35, 469–482. [Google Scholar]
- Krokene, P.; Solheim, H. Fungal associates of five bark beetle species colonizing Norway spruce. Can. J. For. Res. 1997, 26, 2115–2122. [Google Scholar] [CrossRef]
- Jankowiak, R. Fungi associated with Ips typographus on Picea abies in southern Poland and their succession into the phloem and sapwood of beetle-infested trees and logs. For. Path. 2005, 35, 37–55. [Google Scholar] [CrossRef]
- Jankowiak, R.; Hilszczański, J. Ophiostomatoid fungi associated with Ips typographus L. on Picea abies (L.) H. Karst. and Pinus sylvestris L. in North-Eastern Poland. Acta Soc. Bot. Pol. 2005, 74, 345–350. [Google Scholar] [CrossRef]
- Kirisits, T. Fungi isolated from Picea abies infested by the bark beetle Ips typographus in the Białowieża forest in north-eastern Poland. For. Path. 2010, 40, 100–110. [Google Scholar] [CrossRef]
- Linnakoski, R.; de Beer, Z.W.; Niemelä, P.; Wingfield, M.J. Associations of conifer-infesting bark beetles and fungi in fennoscandia. Insects 2012, 3, 200–227. [Google Scholar] [CrossRef]
- Six, D.L. Ecological and evolutionary determinants of bark beetle—Fungus Symbioses. Insects 2012, 3, 339–366. [Google Scholar] [CrossRef] [PubMed]
- Westra, E.R.; van Houte, S.; Oyesiku-Blakemore, S.; Makin, B.; Broniewski, J.M.; Best, A.; Bondy-Denomy, J.; Davidson, A.; Boots, M.; Buckling, A. Parasite exposure drives selective evolution of constitutive versus inducible defense. Curr. Biol. 2015, 25, 1043–1049. [Google Scholar] [CrossRef]
- Raffa, K.F. Mixed messages across multiple trophic levels: The ecology of bark beetle chemical communication systems. Chemoecology 2001, 11, 49–65. [Google Scholar] [CrossRef]
- Harding, S. The Influence of Mutualistic Blue Stain Fungi on Bark Beetle Population Dynamics. Ph.D. Thesis, Department of Zoology, Royal Veterinary & Agricultural University, Copenhagen, Denmark, 1989. [Google Scholar]
- Hammerbacher, A.; Kandasamy, D.; Ullah, C.; Schmidt, A.; Wright, L.; Gershenzon, J. Flavanone-3-hydroxylase plays an important role in the biosynthesis of spruce phenolic defenses against bark beetles and their fungal associates. Front. Plant. Sci. 2019, 10, 208. [Google Scholar] [CrossRef]
- Malloch, D.; Blackwell, M. Dispersal biology of the Ophiostomatid fungi. In Ceratocystis and Ophiostoma: Taxonomy, Ecology, and Pathogenicity; Wingfield, M.J., Seifert, K.A., Webber, J.F., Eds.; APS Press: St. Paul, MN, USA, 1993; pp. 195–206. [Google Scholar]
- Ayres, M.P.; Wilkens, R.T.; Ruel, J.J.; Vallery, E. Fungal relationships and the nitrogen budget of phloem-feeding bark beetles (Coleoptera:Scolytidae). Ecology 2000, 81, 2198–2210. [Google Scholar] [CrossRef]
- Mondy, N.; Corio-Coster, M.F. The response of the grape berry moth (Lobesia botrana) to a dietary phytopathogenic fungus (Botrytis cinerea): The significance of fungus sterols. J. Ins. Physiol. 2000, 46, 1557–1564. [Google Scholar] [CrossRef]
- Clayton, R.B. The utilization of sterols by insects. J. Lipid Res. 1964, 5, 3–19. [Google Scholar]
- Kleipzig, K.D.; Six, D.L. Bark beetle-funga;l symbiosis: Contex dependency in complex associations. Symbiosis 2004, 37, 189–205. [Google Scholar]
- Zang, W.; Song, H.; Ding, Q. Application of methane fermenative residues in control of crop diseases and insect pests. Res. Agric. Modern. 2001, 22, 167–170. [Google Scholar]
- Lenhart, K.; Bunge, M.; Ratering, S.; Neu, T.R.; Schüttmann, I.; Greule, M.; Kammann, C.; Schnell, S.; Müller, C.; Zorn, H. Evidence for methane production by saprotrophic fungi. Nat. Comm. 2012, 3, 1046. [Google Scholar] [CrossRef]
- Weissteiner, S.; Huetteroth, W.; Kollmann, M.; Weißbecker, B.; Romani, R.; Schachtner, J.; Schütz, S. Cockchafer larvae smell host root scents in soil. PLoS ONE 2012, 7, e45827. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, D.; Gershenzon, J.; Andersson, M.N.; Hammerbacher, A. Volatile organic compounds influence the interaction of the Eurasian spruce bark beetle (Ips typographus) with its fungal symbionts. ISME 2019, 13, 1788–1800. [Google Scholar] [CrossRef]
- Zhao, T.; Borg-Karlson, A.-K.; Erbilgin, N.; Krokene, P. Host resistance elicited by methyl jasmonate reduces emission of aggregation pheromones by the spruce bark beetle, Ips typographus. Oecologia 2011, 167, 691–699. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Krekling, T.; Christiansen, E. Picea abies (Pinaceae) stems induces defense-related responses in phloem and xylem. Am. J. Bot. 2002, 89, 602–610. [Google Scholar] [CrossRef]
- Blažytė-Čereškienė, L.; Apšegaitė, V.; Radžiutė, S.; Mozūraitis, R.; Būda, V.; Pečiulytė, D. Electrophysiological and behavioural responses of Ips typographus (L.) to trans-4-thujanol—A host tree volatile compound. Ann. For. Sci. 2016, 73, 247–256. [Google Scholar] [CrossRef]
- Vega, F.E.; Blackwell, M. (Eds.) Insect–Fungal Associations: Ecology and Evolution; Oxford University Press: Oxsford, UK, 2005. [Google Scholar]
- Rosenberg, E.; Zilber-Rosenberg, I. Symbiosis and development: The hologenome concept. Birth Defects Res. (Part C) 2011, 93, 56–66. [Google Scholar] [CrossRef]
- Rosenberg, E.; Zilber-Rosenberg, I. The hologenome concept of evolution after 10 years. Microbiome 2018, 6, 78. [Google Scholar] [CrossRef]
- Pilosof, S.; Porter, M.A.; Pascual, M.; Kéfi, S. The multilayer nature of ecological networks. Nat. Ecol. Evol. 2017, 1, 1–9. [Google Scholar] [CrossRef] [PubMed]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sierota, Z.; Grodzki, W. Picea abies–Armillaria–Ips: A Strategy or Coincidence? Forests 2020, 11, 1023. https://doi.org/10.3390/f11091023
Sierota Z, Grodzki W. Picea abies–Armillaria–Ips: A Strategy or Coincidence? Forests. 2020; 11(9):1023. https://doi.org/10.3390/f11091023
Chicago/Turabian StyleSierota, Zbigniew, and Wojciech Grodzki. 2020. "Picea abies–Armillaria–Ips: A Strategy or Coincidence?" Forests 11, no. 9: 1023. https://doi.org/10.3390/f11091023
APA StyleSierota, Z., & Grodzki, W. (2020). Picea abies–Armillaria–Ips: A Strategy or Coincidence? Forests, 11(9), 1023. https://doi.org/10.3390/f11091023