Possibilities of Speciation in the Central Sandy Steppe, Woody Steppe Area of the Carpathian Basin through the Example of Festuca Taxa


 ,
, 
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Inflorescence Measurements
2.2. Leaf Micromorphological Investigations
2.3. Flow Cytometric Analyses
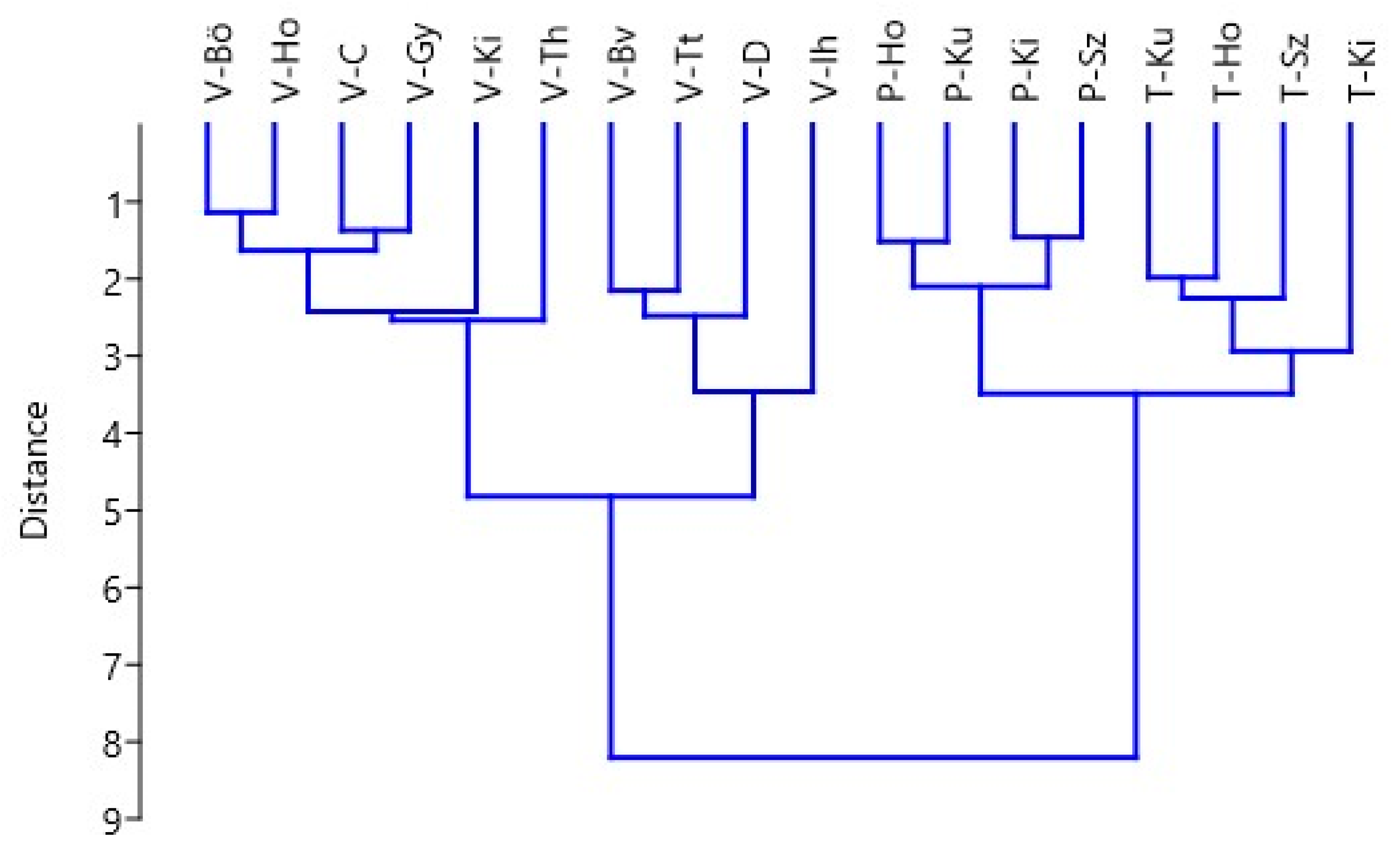
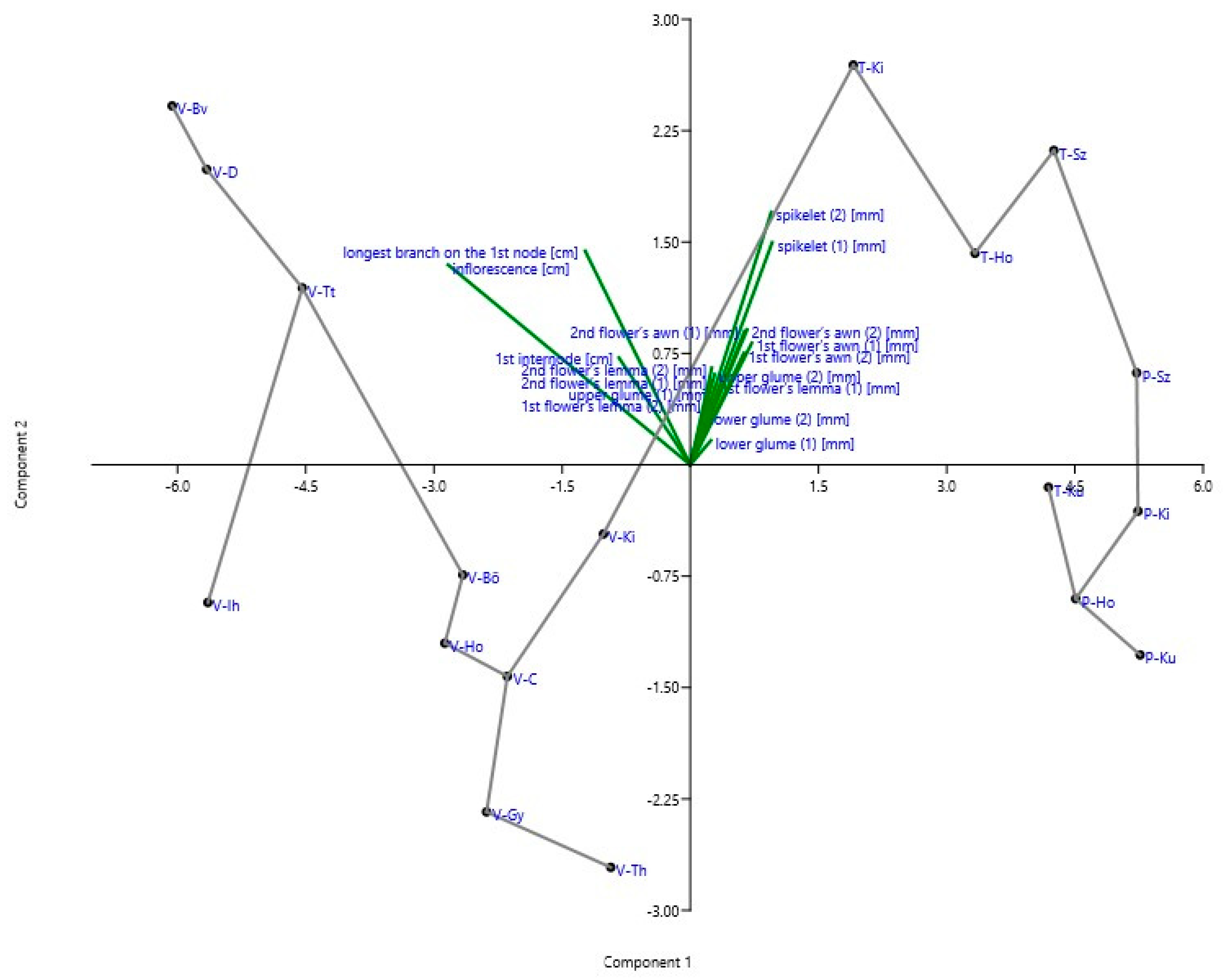
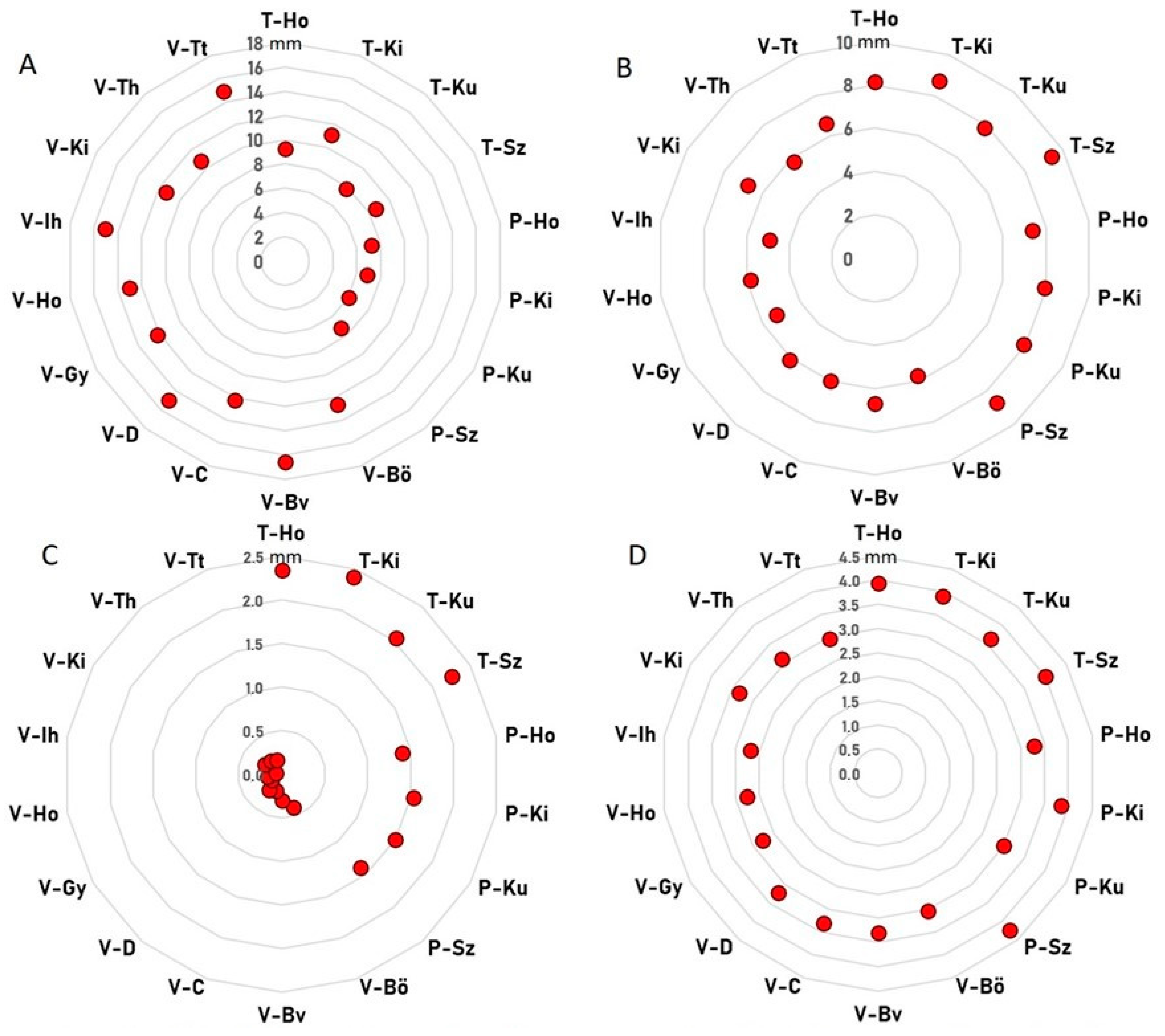
3. Results
3.1. Characters of Inflorescences
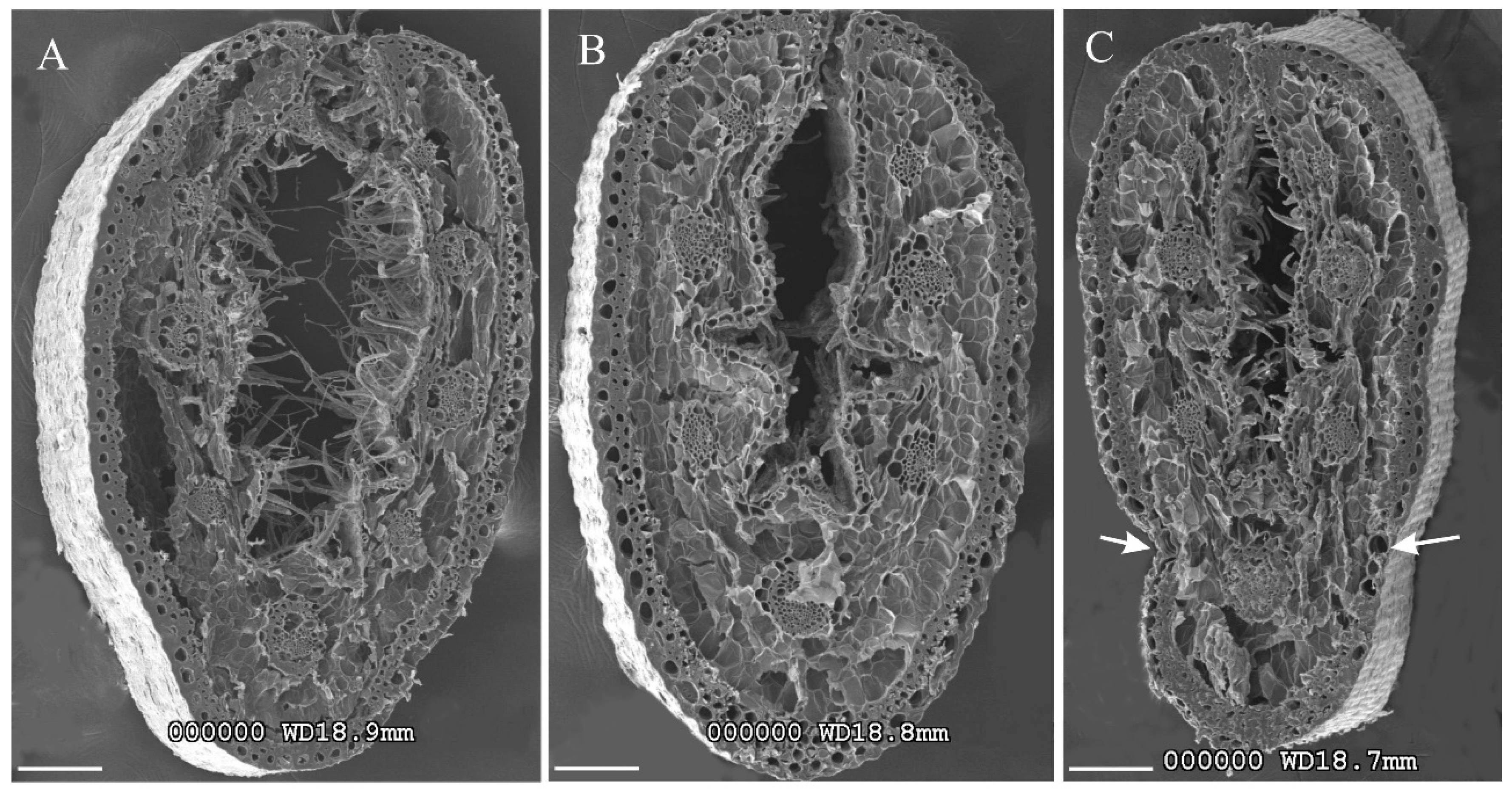
3.2. Leaf Micromorphology
3.2.1. Anatomy
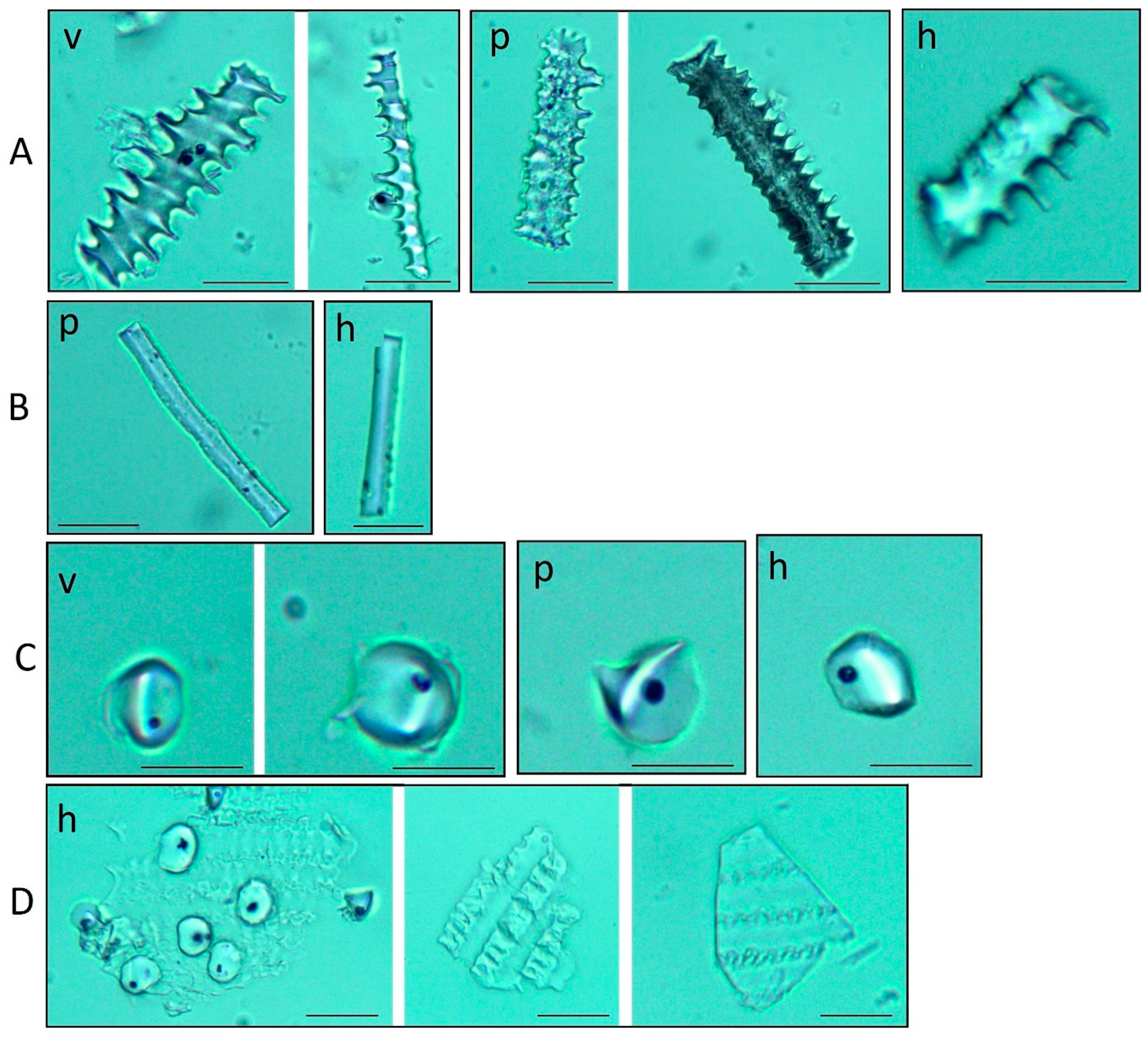
3.2.2. Phytoliths of the Leaves
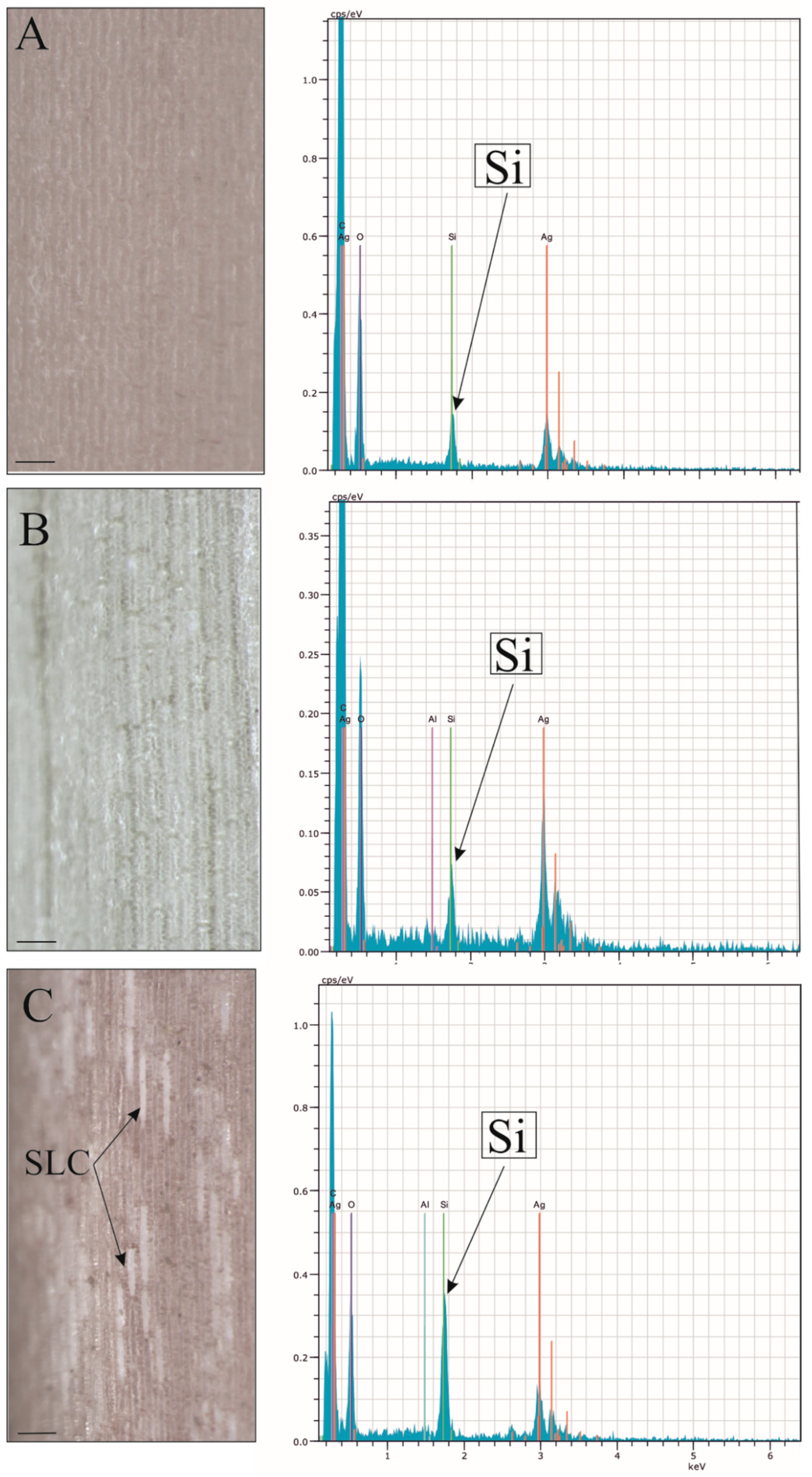
3.2.3. Micromorphological Characters of the Epidermis
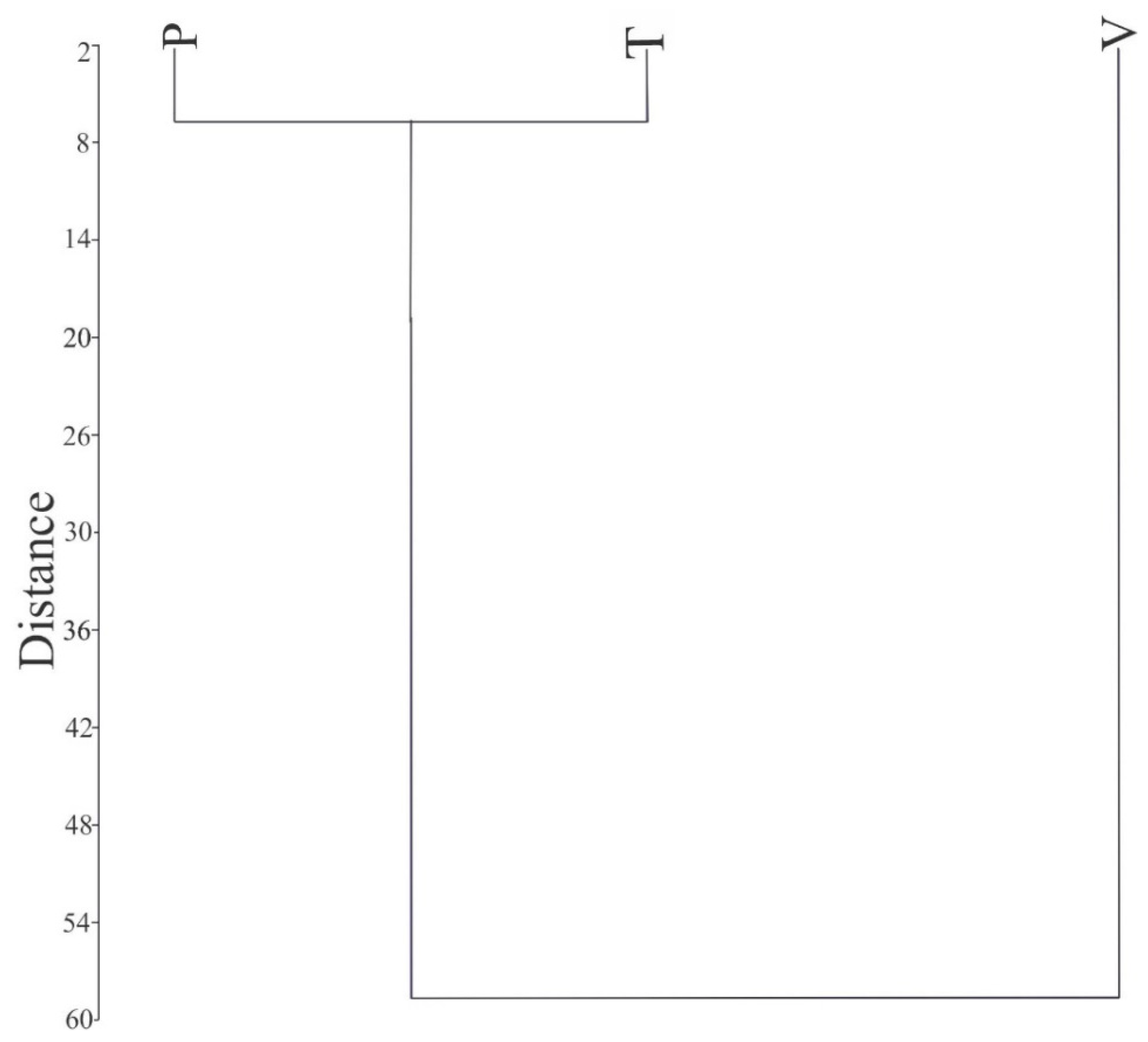
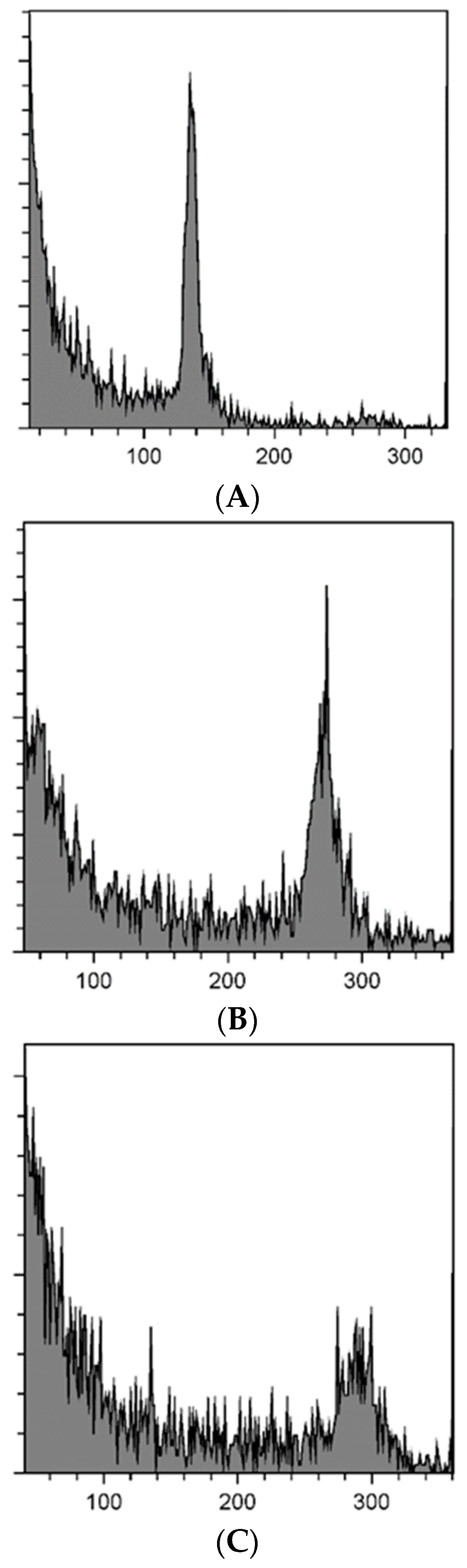
3.3. Determination of Ploidy Level
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Adler, W.; Oswald, K.; Fischer, R. Exkursionflora von Österreich; Excursion Flora of Austria: Ulmer, Austria, 1994. [Google Scholar]
- Dostal, J. Nová Květena ČSSR [New Flora of CSSR] (Vol. 2); Academia Praha: Praha, Czech Republic, 1989. [Google Scholar]
- Király, G. Új Magyar Füvészkönyv. Magyarország Hajtásos Növényei. Határozókulcsok; Aggteleki Nemzeti Park Igazgatóság: Tengerszem, Hungary, 2009.
- Horánszky, A.; Jankó, B.; Vida, G. Zur Biosystematik der Festuca ovina-gruppe in Ungarn [Biosystematics of Festuca ovina-group in Hungary]. Ann. Univ. Sci. Bp. Sect. Biol. 1971, 13, 95–101. [Google Scholar]
- Májovszký, J. Adnotationes ad species gen. Festuca florae Slovakiae additamentum I. Acta Fac. Rer. Nat. Univ. Comen. 1962, 7, 317–355. [Google Scholar]
- Nyárády, E.I.; Nyárády, A. Studie über die Arten der Sektion Ovinae Fr. der Gattung Festuca in der RVR [Study on species of Festuca ovina group in RVR]. Revue Roum. Biol. Sér. Botan. 1964, 9, 99–172. [Google Scholar]
- Patzke, E. Vorschlag zur Gliederung der Festuca ovina L.-Gruppe in Mitteleuropa [Recommendation for classification of the Festuca ovina L. group in Central-Europe]. Österreichische Bot. Z. 1961, 108, 505–507. [Google Scholar] [CrossRef]
- Patzke, E. Zur Kenntnis der Sammelart Festuca ovina L. im südlichen Niedersaschen [Study on Festuca ovina L. in Lower Saxony]. Götting. Flor. Rundbr. 1968, 4, 14–17. [Google Scholar]
- Pils, G. Systematik, Karyologie und Verbreitung der Festuca uaJesiaca-Gruppe (Poaceae) in Österreich und Südtirol. Syst. Karyology Distrib. Festuca Val. Group Austria South Tyrol 1984, 24, 35–77. [Google Scholar]
- Săvulescu, T. Flora Republicii Socialiste România; Editura Academiei Republicii Socialiste România: București, Romania, 1972. [Google Scholar]
- Schwarzová, T. Beitrag zur Lösung taxonomischer Probleme der Festuca vaginata W. et K. und F. psammophila Hack. Acta Fac. Rer. Nat. Univ. Comen. Bot. 1967, 14, 381–414. [Google Scholar]
- Soó, R. Festuca Studien. Acta Bot. Acad. Sci. Hung. 1955, 2, 187–221. [Google Scholar]
- Soó, R. Zeitgemässe Taxonomie der Festuca ovina-gruppe [Recent taxonomy of Festuca ovina-group]. Acta Bot. Sci. Hung. 1973, 18, 363–377. [Google Scholar]
- Soó, R. A Magyar Flóra és Vegetáció Rendszertani-Növényföldrajzi Kézikönyve. II [Taxonomical and Phytogeographical Manual of the Hungarian Flora and Vegetation]; Akadémiai Kiadó: Budapest, Hungary, 1973. [Google Scholar]
- Hackel, E. Monographia Festucarum europaearum. Auctore, Eduardo Hackel. In Monographia Festucarum europaearum. Auctore, Eduardo Hackel.; Smithsonian Institution: Washington, DC, USA, 1882. [Google Scholar]
- Isobe, S.N. Sustainable use of genetic diversity in forage and turf breeding. Ann. Bot. 2013, 111, 41–45. [Google Scholar] [CrossRef][Green Version]
- Bednarska, I.; Kostikov, I.; Tarieiev, A.; Stukonis, V. Morphological, Karyological and Molecular Characteristics of Festuca arietina Klok.—A Neglected Psammophilous Species of the Festuca valesiaca agg. from Eastern Europe. Acta Biol. Crac. Bot. 2017, 59, 35–53. [Google Scholar] [CrossRef]
- Galli, Z.; Penksza, K.; Kiss, E.; Bucherna, N.; Heszky, L. Festuca fajok molekuláris taxonómiai vizsgálata a F. ovina csoport RAPD és AP-PCR analízise [Molecular taxonomic analysis of Festuca species: RAPD and AP-PCR analysis of the F-ovina group]. Növénytermelés 2001, 50, 375–384. [Google Scholar]
- Galli, Z.; Penksza, K.; Kiss, E.; Sági, L.; Heszky, L.E. Low variability of internal transcribed spacer rDNA and trnL (UAA) intron sequences of several taxa in the Festuca ovina aggregate (Poaceae). Acta Biol. Hung. 2006, 57, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Gaut, B.S.; Tredway, L.P.; Kubík, C.; Gaut, R.L.; Meyer, W. Phylogenetic relationships and genetic diversity among members of the Festuca-Lolium complex (Poaceae) based on ITS sequence data. Plant Syst. Evol. 2000, 224, 33–53. [Google Scholar] [CrossRef]
- Loureiro, J.; Kopecký, D.; Castro, S.; Santos, C.; Silveira, P. Flow cytometric and cytogenetic analyses of Iberian Peninsula Festuca spp. Plant Syst. Evol. 2007, 269, 89–105. [Google Scholar] [CrossRef]
- Šmarda, P. DNA ploidy levels and intraspecific DNA content variability in Romanian fescues (Festuca, Poaceae) measured in fresh and herbarium material. Folia Geobot. Phytotaxon. 2006, 41, 417–432. [Google Scholar] [CrossRef]
- Šmarda, P. DNA ploidy level variability of some fescues (Festuca subg. Festuca, Poaceae) from Central and Southern Europe measured in fresh plants and herbarium specimens. Biology 2008, 63, 349–367. [Google Scholar] [CrossRef]
- Šmarda, P.; Kočí, K. Chromosome number variability in central european members of the Festuca ovina and F. pallens groups (sect. Festuca). Folia Geobot. Phytotaxon. 2003, 38, 65–95. [Google Scholar] [CrossRef]
- Šmarda, P.; Šmerda, J.; Knoll, A.; Bureš, P.; Danihelka, J. Revision of Central European taxa of Festuca ser. Psammophilae Pawlus: Morphometrical, karyological and AFLP analysis. Plant Syst. Evol. 2007, 266, 197–232. [Google Scholar] [CrossRef]
- Šmarda, P.; Bureš, P.; Horová, L.; Foggi, B.; Rossi, G. Genome Size and GC Content Evolution of Festuca: Ancestral Expansion and Subsequent Reduction. Ann. Bot. 2008, 101, 421–433. [Google Scholar] [CrossRef]
- Stukonis, V.; Armonienė, R.; Lemežienė, N.; Kemešytė, V.; Statkevičiūtė, G. Identification of fine-leaved species of genus Festuca by molecular methods. Pak. J. Bot. 2015, 47, 1137–1142. [Google Scholar]
- Pawlus, M. Systematyka i rozmieszczenie gatunków grupy Festuca ovina L. w. Polsce [Classification and distribution of Festuca ovina L. in Poland]. Fragm. Florist. Geobot. 1985, 29, 219–295. [Google Scholar]
- Borhidi, A.; Kevey, B.; Lendvai, G.; Seregélyes, T. Plant Communities of Hungary; Akadémiai Kiadó: Budapest, Hungary, 2012. [Google Scholar]
- Krajina, V. Adnotationes ad species generis Festuca in Flora Cechoslovenika exsiccata. Acta Bot. Bohem. 1930, 9, 186–220. [Google Scholar]
- Soó, R.; Jávorka, S. A magyar növényvilág kézikönyve. In Handbook of Hungarian Flora; Akadémiai Kiadó: Budapest, Hungary, 1951. [Google Scholar]
- The Plant List. Available online: www.theplantlist.org (accessed on 8 September 2020).
- Penksza, K.; Szabó, G.; Zimmermann, Z.; Lisztes-Szabó, Z.; Pápay, G.; Járdi, I.; Fűrész, A.; S-Falusi, E. A Festuca vaginata alakkör taxonómiai problematikája és ennek cönoszisztematikai vonatkozásai [The taxonomic problems of the Festuca vaginata agg. and their coenosystematic aspects]. Georg. Agric. 2019, 23, 63–76. [Google Scholar]
- Penksza, K. Kiegészítések a hazai Festuca taxonok ismeretéhez I.: A Festuca psammophila series Festuca vaginata alakköre. Bot. Közlemények 2019, 106, 65–70. [Google Scholar] [CrossRef]
- Wilkinson, M.J. Stace, The taxonomic relationship and typification of Festuca brevipila Tracey and Festuca lemanii Bastard (Poaceae). Watsonia 1989, 17, 289–299. [Google Scholar]
- Budak, H.; Shearman, R.; Gaussoin, R.; Dweikat, I. Application of Sequence-related Amplified Polymorphism Markers for Characterization of Turfgrass Species. HortScience 2004, 39, 955–958. [Google Scholar] [CrossRef]
- Lonati, M.; Lonati, S. Le praterie xerofile a Festuca trachyphylla (Hackel) Krajina della bassa Valsesia (Piemonte, Italia) [The xerophilous Festuca trachyphylla (Hackel) Krajina grasslands in the lower Valsesia (Piedmont, Italy)]. Fitosociologia 2007, 44, 109–118. [Google Scholar]
- Stukonis, V.; Lemežienė, N.; Kanapeckas, J. Suitability of narow-leaved Festuca species for turf. Agron. Res. 2010, 8, 729–734. [Google Scholar]
- Dąbrowska, A. Morpho-anatomical structure of the leaves of Festuca trahyphylla (Hack.) Krajina in the ecological aspect. Mod. Phytomorphol. 2012, 1, 19–22. [Google Scholar] [CrossRef]
- Gugnacka-Fiedor, W.; Adamska, E. The preservation state of the flora and vegetation of the artillery range near the city of Toruń. Ecol. Quest. 2010, 12, 75–86. [Google Scholar] [CrossRef][Green Version]
- Löbel, S.; Dengler, J. Dry grassland communities on southern Öland: Phytosociology, ecology, and diversity. Acta Phytogeogr. Suec. 2008, 88, 13–31. [Google Scholar] [CrossRef][Green Version]
- Rūsiņa, S. Nelku aira Aira Caryophyllea L. Latvija [Aira caryophyllea L. in Latvia]. Latv. Veģetācija 2003, 7, 33–43. [Google Scholar]
- Kaczmarek, Z.; Gajewski, P.; Mocek, A.; Grzelak, M.; Knioła, A.; Glina, B. Geobotanical conditions of ecological grasslands on light river alluvial soils. J. Res. Appl. Agric. Eng. 2015, 60, 131–135. [Google Scholar]
- Böhnert, W.; Reichhoff, L. Die Pflanzengesellschaften des Naturschutzgebietes “Steckby-Lödderitzer Forst”. Plant Communities Steckby-Lödderitzer Forst Nat. Reserve 1978, 15, 106–114. [Google Scholar]
- Fischer, W.; Kummer, V.; Pötsch, J. Zur Vegetation des Feuchtgebietes Internationaler Bedeutung (FIB) untere Havel [Vegetation of lower Havel, an internationally important wetland area]. Nat. Landsch. Brandenbg. 1995, 4, 12–18. [Google Scholar]
- Nienartowicz, A.; Kaminski, D.; Kunz, M.; Deptuła, M.; Adamska, E. Changes in the plant cover of the dune hill in Folusz near Szubin (NW Poland) between 1959 and 2012: The problem of preservation of xerothermic grasslands in the agricultural landscape. Ecol. Quest. 2015, 20, 23. [Google Scholar] [CrossRef][Green Version]
- Kovář, P. Contribution to the syntaxonomy of the Festuca trachyphylla-grass-lands. Preslia 1980, 52, 217–226. [Google Scholar]
- Di Pietro, R. New Dry Grassland Associations from the Ausoni-Aurunci Mountains (Central Italy)-Syntaxonomical Updating and Discussion on the Higher Rank Syntaxa. Hacquetia 2011, 10, 183–231. [Google Scholar] [CrossRef]
- Stace, C.A.; Al-Bermani, A.-K.K.A.; Wilkinson, M.J. The distinction between the Festuca ovina L. and Festuca rubra L. aggregates in the British Isles. Watsonia 1992, 19, 107–112. [Google Scholar]
- Blackman, E. Opaline silica bodies in the range grasses of southern Alberta. Can. J. Bot. 1971, 49, 769–781. [Google Scholar] [CrossRef]
- Brown, W.V. Leaf Anatomy in Grass Systematics. Int. J. Plant Sci. 1958, 119, 170–178. [Google Scholar] [CrossRef]
- Metcalfe, C.R. Anatomy of the Monocotyledons: I. Gramineae; Clarendon Press: Oxford, UK, 1960. [Google Scholar]
- Palmer, P.G.; Tucker, A.E. A Scanning Electron Microscope Survey of the Epidermis of East African Grasses, I. Smithson. Contrib. Bot. 1981, 49, 1–84. [Google Scholar] [CrossRef]
- Tateoka, T.; Inoue, S.; Kawano, S. Notes on Some Grasses. IX. Systematic Significance of Bicellular Microhairs of Leaf Epidermis. Int. J. Plant Sci. 1959, 121, 80–91. [Google Scholar] [CrossRef]
- Brown, D.A. Prospects and limits of a phytolith key for grasses in the central United States. J. Archaeol. Sci. 1984, 11, 345–368. [Google Scholar] [CrossRef]
- Fredlund, G.G.; Tieszen, L.T. Modern Phytolith Assemblages from the North American Great Plains. J. Biogeogr. 1994, 21, 321. [Google Scholar] [CrossRef]
- Krishnan, S.; Samson, N.P.; Ravichandran, P.; Narasimhan, D.; Dayanandan, P. Phytoliths of Indian grasses and their potential use in identification. Bot. J. Linn. Soc. 2000, 132, 241–252. [Google Scholar] [CrossRef]
- Lu, H.; Liu, K.-B. Morphological variations of lobate phytoliths from grasses in China and the south-eastern United States. Divers. Distrib. 2002, 9, 73–87. [Google Scholar] [CrossRef]
- Mulholland, S.C. Phytolith shape frequencies in North Dakota grasses: A comparison to general patterns. J. Archaeol. Sci. 1989, 16, 489–511. [Google Scholar] [CrossRef]
- Mulholland, S.C.; Rapp, G. A Morphological Classification of Grass Silica-Bodies. In Phytolith Systematics; Springer Science and Business Media LLC: Berlin, Germany, 1992; pp. 65–89. [Google Scholar]
- Prychid, C.J.; Rudall, P.J.; Gregory, M. Systematics and Biology of Silica Bodies in Monocotyledons. Bot. Rev. 2003, 69, 377–440. [Google Scholar] [CrossRef]
- Twiss, P.C.; Suess, E.; Smith, R.M. Morphological Classification of Grass Phytoliths. Soil Sci. Soc. Am. J. 1969, 33, 109–115. [Google Scholar] [CrossRef]
- Pepi, M.F.; Zucol, A.F.; Arriaga, M. Comparative phytolith analysis of Festuca (Pooideae: Poaceae) species native to Tierra del Fuego, Argentina. Botany 2012, 90, 1113–1124. [Google Scholar] [CrossRef]
- Ortúñez, E.; De La Fuente, V. Epidermal micromorphology of the genus Festuca L. (Poaceae) in the Iberian Peninsula. Plant Syst. Evol. 2010, 284, 201–218. [Google Scholar] [CrossRef]
- Zarinkamar, F.; Jouyandeh, N.E. Foliar anatomy and micromorphology of Festuca L. and its taxonomic applications. Taxon. Biosyst. 2011, 8, 55–63. [Google Scholar]
- Korneck, D.; Gregor, T. Festuca tomanii sp. nov., ein Dünen-Schwingel des nördlichen oberrhein-, des mittleren main- und des böhmischen Elbetales. Kochia 2015, 9, 37–58. [Google Scholar]
- Bajor, Z. Effect of conservation management practices on sand grassland vegetation in budapest, hungary. Appl. Ecol. Environ. Res. 2016, 14, 233–247. [Google Scholar] [CrossRef]
- Csány-Kovács, C.; Horánszky, A. Charakterisierung der Festuca-Populationen aufgrund der Merkmale der Rispe. Ann. Univ. Sci. Bp. Rolando Eötvös Nomin. Sect. Biol. 1973, 15, 59–74. [Google Scholar]
- Hammer, O.; Harper, D.; Ryan, P. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Harper, D.A.T. Numerical Palaeobiology: Computer-Based Modelling and Analysis of Fossils and Their Distributions; John Wiley & Sons: Hoboken, NJ, USA, 1999. [Google Scholar]
- Weiner, S.; Albert, R. Study of Phytoliths in Prehistoric Ash Layers from Kebara and Tabun Caves Using a Quantitative Approach. In Phytoliths-Applications in Earth Science and Human History; Informa UK Limited: Colchester, UK, 2001; pp. 251–266. [Google Scholar]
- Mercader, J.; Bennett, T.; Esselmont, C.; Simpson, S.; Walde, D. Phytoliths in woody plants from the Miombo woodlands of Mozambique. Ann. Bot. 2009, 104, 91–113. [Google Scholar] [CrossRef]
- Mercader, J.; Astudillo, F.; Barkworth, M.; Bennett, T.; Esselmont, C.; Kinyanjui, R.; Grossman, D.L.; Simpson, S.; Walde, D. Poaceae phytoliths from the Niassa Rift, Mozambique. J. Archaeol. Sci. 2010, 37, 1953–1967. [Google Scholar] [CrossRef]
- Neumann, K.; Strömberg, C.A.E.; Ball, T.; Albert, R.M.; Vrydaghs, L.; Cummings, L.S.; International Committee for Phytolith Taxonomy (ICPT). International Code for Phytolith Nomenclature (ICPN) 2.0. Ann. Bot. 2019, 124, 189–199. [Google Scholar] [CrossRef]
- Galbraith, D.W.; Harkins, K.R.; Maddox, J.M.; Ayres, N.M.; Sharma, D.P.; Firoozabady, E. Rapid flow cytometric analysis of the cell cycle in intact plant tissues. Science 1983, 220, 1049–1051. [Google Scholar] [CrossRef]
- Penksza, K. Festuca pseudovaginata, a new species from sandy areas of the Carpathian Basin. Acta Bot. Hung. 2003, 45, 365–372. [Google Scholar] [CrossRef]
- Lisztes-Szabó, Z.; Kovács, S.; Balogh, P.; Daróczi, L.; Penksza, K.; Pető, Á. Quantifiable differences between phytolith assemblages detected at species level: Analysis of the leaves of nine Poa species (Poaceae). Acta Soc. Bot. Pol. 2015, 84, 369–383. [Google Scholar] [CrossRef]
- Horánszky, A. Festuca-tanulmányok I. [Festuca-studies I.]. Bot. Közlemények 1969, 56, 149–154. [Google Scholar]
- Horánszky, A. Festuca-tanulmányok II. [Festuca-studies II.]. Bot. Közlemények 1970, 57, 207–215. [Google Scholar]
- Ladányi, Z.; Rakonczai, J.; Kovács, F.; Geiger, J.; Deák, J.Á. The effect of recent climatic change on the Great Hungarian Plain. Cereal Res. Commun. 2009, 37, 477–480. [Google Scholar]
- Ladányi, Z.; Rakonczai, J.; Van Leeuwen, B. Evaluation of precipitation-vegetation interaction on a climate-sensitive landscape using vegetation indices. J. Appl. Remote Sens. 2011, 5, 53519. [Google Scholar] [CrossRef]
- Jones, L.H.P.; Handreck, K.A. Studies of silica in the oat plant. Plant Soil 1965, 23, 79–96. [Google Scholar] [CrossRef]
- Madella, M.; Jones, M.; Echlin, P.; Powers-Jones, A.; Moore, M. Plant water availability and analytical microscopy of phytoliths: Implications for ancient irrigation in arid zones. Quat. Int. 2009, 193, 32–40. [Google Scholar] [CrossRef]
- Raven, J.A. The transport and function of silicon in plants. Biol. Rev. 1983, 58, 179–207. [Google Scholar] [CrossRef]
- Sangster, A.G.; Parry, D.W. Silica Deposition in the Grass Leaf in Relation to Transpiration and the Effect of Dinitrophenol. Ann. Bot. 1971, 35, 667–677. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GSSCP (%) | Elongate Entire (%) | Elongate Sinuate (%) | Elongate Dentate (%) | Elongate (%) | Acute Bulbosus (%) | |
|---|---|---|---|---|---|---|
| F. vaginata | 88.3 | 81.2 | 11.9 | 6.9 | 10.7 | 1.0 |
| F. pseudovaginata | 81.6 | 39.2 | 52.5 | 8.3 | 17.7 | 0.7 |
| F. tomanii | 84.5 | 39.6 | 51.4 | 9.0 | 14.5 | 1.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Penksza, K.; Csík, A.; Filep, A.F.; Saláta, D.; Pápay, G.; Kovács, L.; Varga, K.; Pauk, J.; Lantos, C.; Lisztes-Szabó, Z. Possibilities of Speciation in the Central Sandy Steppe, Woody Steppe Area of the Carpathian Basin through the Example of Festuca Taxa. Forests 2020, 11, 1325. https://doi.org/10.3390/f11121325
Penksza K, Csík A, Filep AF, Saláta D, Pápay G, Kovács L, Varga K, Pauk J, Lantos C, Lisztes-Szabó Z. Possibilities of Speciation in the Central Sandy Steppe, Woody Steppe Area of the Carpathian Basin through the Example of Festuca Taxa. Forests. 2020; 11(12):1325. https://doi.org/10.3390/f11121325
Chicago/Turabian StylePenksza, Károly, Attila Csík, Anna Fruzsina Filep, Dénes Saláta, Gergely Pápay, László Kovács, Kristina Varga, János Pauk, Csaba Lantos, and Zsuzsa Lisztes-Szabó. 2020. "Possibilities of Speciation in the Central Sandy Steppe, Woody Steppe Area of the Carpathian Basin through the Example of Festuca Taxa" Forests 11, no. 12: 1325. https://doi.org/10.3390/f11121325
APA StylePenksza, K., Csík, A., Filep, A. F., Saláta, D., Pápay, G., Kovács, L., Varga, K., Pauk, J., Lantos, C., & Lisztes-Szabó, Z. (2020). Possibilities of Speciation in the Central Sandy Steppe, Woody Steppe Area of the Carpathian Basin through the Example of Festuca Taxa. Forests, 11(12), 1325. https://doi.org/10.3390/f11121325