Aspergillus tubingensis Is a Pre-Emergent Pathogen of Date Palm Seedlings



Abstract
1. Introduction
2. Materials and Methods
2.1. Seed Germination and Seedling Emergence
2.2. Identification of Seedborne Fungi
2.3. Koch’s Postulates/Initial Aspergillus Experiment
2.4. Experiment with Aspergillus tubingensis
2.5. Statistical Analyses
3. Results
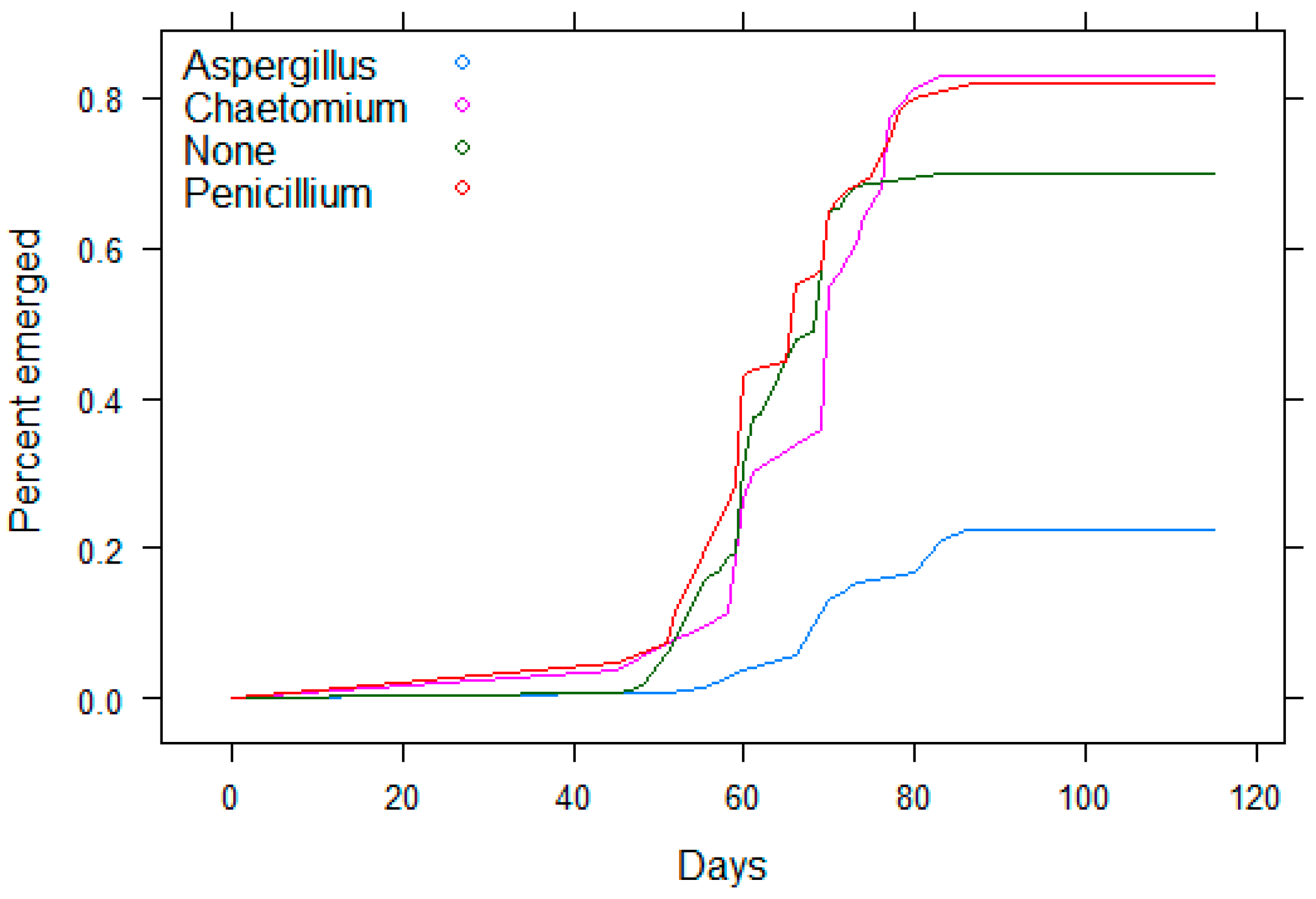
3.1. Koch’s Postulates/Initial Aspergillus Experiment
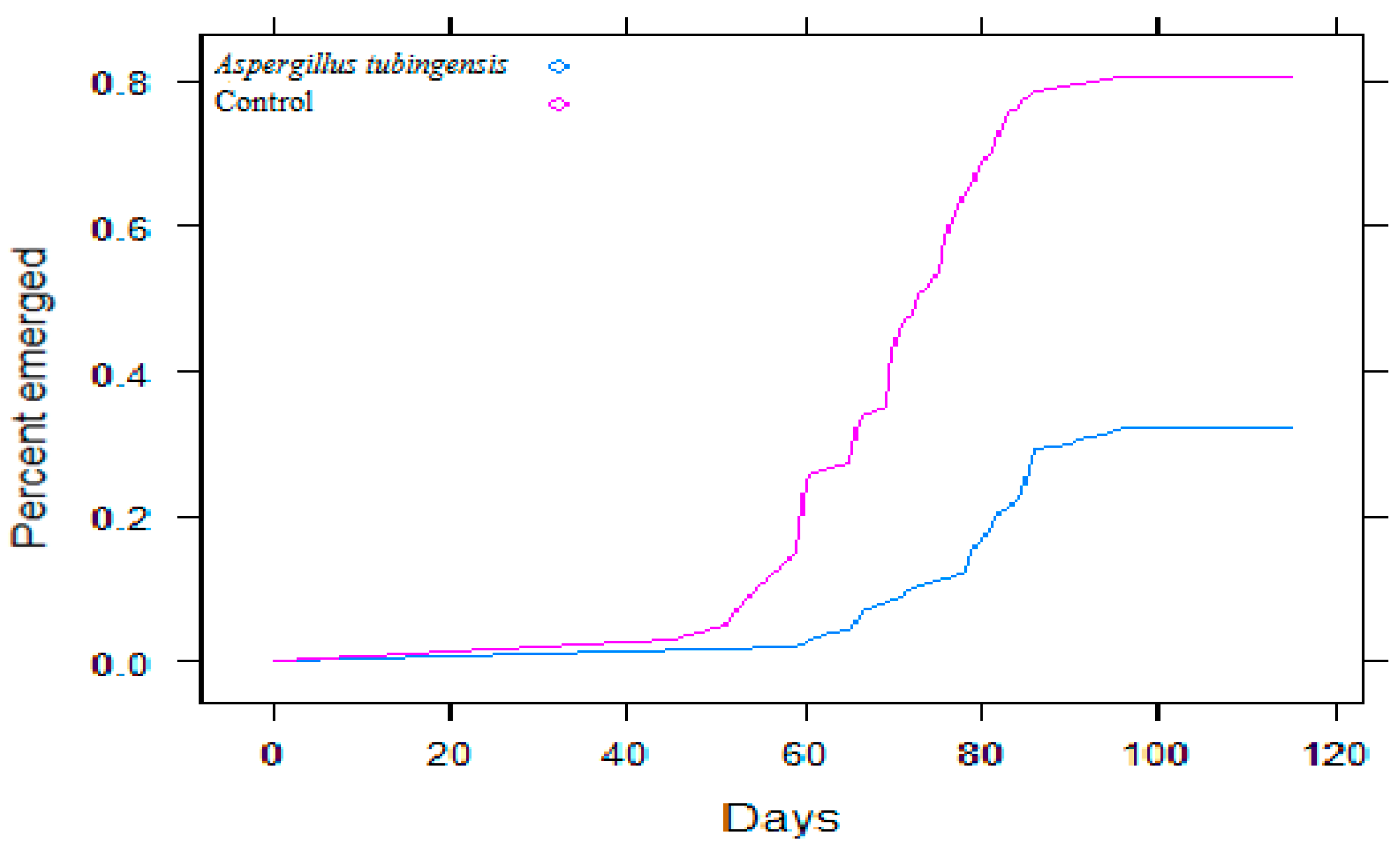
3.2. Experiment with Aspergillus tubingensis
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abdelmonem, A.M.; Rasmy, M.R. Major diseases of date palm and their control. Commun. Inst. For. Bohem. 2007, 23, 9–23. [Google Scholar]
- Chao, C.T.; Krueger, R.R. The Date Palm (Phoenix dactylifera L.): Overview of Biology, Uses, and Cultivation. HortScience 2007, 42, 1077–1082. [Google Scholar] [CrossRef]
- Fairchild, D. Seeds and Plants Imported during the Period from July 1, 1906 to December 1, 1907; Inventory No. 13: Nos. 19058 to 21730; U.S. Government Publishing Office: Washington, DC, USA, 1908.
- McCarthy, M.A. Date Palms in the Desert: Reimagining and Cooperating with Nature in Arid Arizona. Ariz. J. Interdiscip. Stud. 2012, 1, 39. [Google Scholar]
- Mitchell, C.E.; Power, A.G. Release of invasive plants from fungal and viral pathogens. Nat. Cell Biol. 2003, 421, 625–627. [Google Scholar] [CrossRef] [PubMed]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, Systematic Mycology and Microbiology Laboratory, ARS, USDA. Available online: http://nt.ars-grin.gov/fungaldatabases/ (accessed on 12 October 2020).
- Zaid, A.; Arias-Jimenez, A.J. Date Palm Cultivation; FAO Plant Production and Protection Paper 156. 2017. Available online: http://www.fao.org/docrep/006/Y4360E/y4360e0g.htm#bm16 (accessed on 12 October 2020).
- Sallon, S.; Solowey, E.; Cohen, Y.; Korchinsky, R.; Egli, M.; Woodhatch, I.; Simchoni, O.; Kislev, M. Germination, Genetics, and Growth of an Ancient Date Seed. Science 2008, 320, 1464. [Google Scholar] [CrossRef] [PubMed]
- Al-Sheikh, H. Date-Palm Fruit Spoilage and Seed-Borne Fungi of Saudi Arabia. Res. J. Microbiol. 2009, 4, 208–213. [Google Scholar] [CrossRef]
- Bokhary, H. Seed-borne fungi of date-palm, Phoenix dactylifera L. from Saudi Arabia. Saudi J. Biol. Sci. 2010, 17, 327–329. [Google Scholar] [CrossRef]
- Houbraken, J.; Kocsubé, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.-C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef]
- Samson, R.A.; Visagie, C.; Houbraken, J.; Hong, S.-B.; Hubka, V.; Klaassen, C.; Perrone, G.; Seifert, K.; Susca, A.; Tanney, J.; et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef]
- Simmons, E.G. Alternaria: An Identification Manual; Series No.6, Utrecht; CBS Fungal Biodiversity Centre: Utrecht, The Netherlands, 2007. [Google Scholar]
- Haq, I.U.; Khan, N.A. Fungal Diseases of Date Palm (Phoenix dactylifera): Etiology and Management. In Sustainability in Plant and Crop Protection; Springer: Cham, Switzerland, 2020; pp. 169–196. [Google Scholar]
- Varga, J.; Frisvad, J.; Kocsubé, S.; Brankovics, B.; Tóth, B.; Szigeti, G.; Samson, R. New and revisited species in Aspergillus section Nigri. Stud. Mycol. 2011, 69, 1–17. [Google Scholar] [CrossRef]
- Abdullah, S.K.; Monfort, E.; Asensio, L.; Salinas, J.; Llorca, L.V.L.; Jansson, H.B. Soil mycobiota of date palm plantations in Elche, SE Spain. Czech Mycol. 2010, 61, 149–162. [Google Scholar] [CrossRef]
- Palou, L.; Rosales, R.; Taberner, V.; Vilella-Espla, J. Incidence and etiology of postharvest diseases of fresh fruit of date palm (Phoenix dactylifera L.) in the grove of Elx (Spain). Phytopathol. Mediterr. 2017, 55, 391–400. [Google Scholar]
- Andersen, B.; Thrane, U. Food-borne fungi in fruit and cereals and their production of mycotoxins. In Advances in Experimental Medicine and Biology; Springer: Boston, MA, USA, 2006; Volume 571, pp. 137–152. [Google Scholar]
- Piombo, E.; Abdelfattah, A.; Danino, Y.; Salim, S.; Feygenberg, O.; Spadaro, D.; Wisniewski, M.; Droby, S. Characterizing the Fungal Microbiome in Date (Phoenix dactylifera) Fruit Pulp and Peel from Early Development to Harvest. Microorganisms 2020, 8, 641. [Google Scholar] [CrossRef] [PubMed]
- Quaglia, M.; Santinelli, M.; Sulyok, M.; Onofri, A.; Covarelli, L.; Beccari, G. Aspergillus, Penicillium and Cladosporium species associated with dried date fruits collected in the Perugia (Umbria, Central Italy) market. Int. J. Food Microbiol. 2020, 322, 108585. [Google Scholar] [CrossRef]
- Guo, J.-W.; Gao, Y.; Li, C.-Y.; Yang, L.-F.; Tian, X.-J.; Hong, L.; Kong, Q.; Zhang, Y.-G.; Li, W.-J. First Report of Leaf Spot Disease Caused by Aspergillus tubingensis on Jatropha curcas in Yunnan, China. Plant Dis. 2017, 101, 505. [Google Scholar] [CrossRef]
- Ridout, M.; Newcombe, G. Sydowia polyspora is both a foliar endophyte and a pre-emergent seed pathogen in Pinus ponderosa. Plant Dis. 2018, 102, 640–644. [Google Scholar] [CrossRef]
- Palou, L.; Montesinos-Herrero, C.; Taberner, V.; Vilella-Esplá, J. First Report of Alternaria alternata Causing Postharvest Black Spot of Fresh Date Palm Fruit in Spain. Plant Dis. 2013, 97, 286. [Google Scholar] [CrossRef]
- Metoui, M.; Essid, A.; Bouzoumita, A.; Ferchichi, A. Chemical Composition, Antioxidant and Antibacterial Activity of Tunisian Date Palm Seed. Pol. J. Environ. Stud. 2018, 28, 267–274. [Google Scholar] [CrossRef]
- AbdelGawad, H.; Saleh, A.M.; Al Jaouni, S.; Selim, S.; Hassan, M.O.; Wadaan, M.A.; Shuikan, A.M.; Mohamed, H.S.; Hozzein, W.N. Utilization of actinobacteria to enhance the production and quality of date palm (Phoenix dactylifera L.) fruits in a semi-arid environment. Sci. Total Environ. 2019, 665, 690–697. [Google Scholar] [CrossRef]
- Boutheina, Z.-E.; Aya, H.; Naima, B.; Ahmed, N. Responses of date Palm Seedling to co-Inoculation with Phosphate Solubilizing Bacteria and Mycorrhizal Arbuscular Fungi. Int. J. Environ. Agric. Biotechnol. 2019, 4, 581–588. [Google Scholar] [CrossRef]
- Shipunov, A.; Newcombe, G.; Raghavendra, A.K.H.; Anderson, C.L. Hidden diversity of endophytic fungi in an invasive plant. Am. J. Bot. 2008, 95, 1096–1108. [Google Scholar] [CrossRef] [PubMed]
- Baynes, M.; Newcombe, G.; Dixon, L.; Castlebury, L.; O’Donnell, K. A novel plant–fungal mutualism associated with fire. Fungal Biol. 2012, 116, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Newcombe, G.; Harding, A.; Ridout, M.; Busby, P.E. A Hypothetical Bottleneck in the Plant Microbiome. Front. Microbiol. 2018, 9, 1645. [Google Scholar] [CrossRef] [PubMed]
- Raghavendra, A.K.H.; Newcombe, G.; Shipunov, A.; Baynes, M.; Tank, D. Exclusionary interactions among diverse fungi infecting developing seeds of Centaurea stoebe. FEMS Microbiol. Ecol. 2013, 84, 143–153. [Google Scholar] [CrossRef] [PubMed][Green Version]




{kind=link}
{kind=link}
{kind=link}
| Alternaria | Aspergillus | Chaetomium | Penicillium | Unknown | Total | |
|---|---|---|---|---|---|---|
| Th-SA | 0 | 4 | 14 | 13 | 1 | 32 |
| Th-USA | 0 | 8 | 27 | 73 | 0 | 108 |
| Ha-SA | 0 | 45 | 14 | 19 | 0 | 78 |
| Ha-USA | 0 | 6 | 1 | 2 | 0 | 9 |
| Ba-SA | 0 | 67 | 0 | 5 | 0 | 72 |
| Ba-USA | 34 | 25 | 17 | 22 | 0 | 98 |
| TOTAL | 34 | 155 | 73 | 134 | 1 | 397 |
| Percentages | 8.56% | 39% | 18.38% | 33.75% | 0.25% |
| Status of Seed | (# of Emerged Seedlings) | (#Planted That Did Not Emerge) | Average Number of Days for Emergence | Number of Seeds Germinated in the Lab | Number of Seeds That Did Not Germinate in the Lab |
|---|---|---|---|---|---|
| Fungus-free | 166 | 72 | 62.55 | 160 | 78 |
| Penicillium | 86 | 19 | 63.31 | 81 | 24 |
| Penicillium with Aspergillus | 5 | 7 | 64.2 | 9 | 3 |
| Alternaria | 21 | 10 | 62.3 | 25 | 6 |
| Alternaria with Penicillium | 1 | 1 | 63 | 1 | 1 |
| Alternaria with Aspergillus | 0 | 1 | - | 0 | 1 |
| Penicillium with Chaetomium | 12 | 3 | 66 | 10 | 5 |
| Chaetomium | 44 | 9 | 67 | 18 | 35 |
| Aspergillus with Chaetomium | 4 | 1 | 72.75 | 2 | 3 |
| Aspergillus | 31 | 106 | 71.7 | 58 | 79 |
| Unknown fungi | 1 | 0 | 70 | 1 | 0 |
| Total | 371 | 229 | 365 | 235 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alomran, M.; Houbraken, J.; Newcombe, G. Aspergillus tubingensis Is a Pre-Emergent Pathogen of Date Palm Seedlings. Forests 2020, 11, 1327. https://doi.org/10.3390/f11121327
Alomran M, Houbraken J, Newcombe G. Aspergillus tubingensis Is a Pre-Emergent Pathogen of Date Palm Seedlings. Forests. 2020; 11(12):1327. https://doi.org/10.3390/f11121327
Chicago/Turabian StyleAlomran, Maryam, Jos Houbraken, and George Newcombe. 2020. "Aspergillus tubingensis Is a Pre-Emergent Pathogen of Date Palm Seedlings" Forests 11, no. 12: 1327. https://doi.org/10.3390/f11121327
APA StyleAlomran, M., Houbraken, J., & Newcombe, G. (2020). Aspergillus tubingensis Is a Pre-Emergent Pathogen of Date Palm Seedlings. Forests, 11(12), 1327. https://doi.org/10.3390/f11121327