Transformation of American Chestnut (Castanea dentata (Marsh.) Borkh) Using RITA® Temporary Immersion Bioreactors and We Vitro Containers

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
Embryos for Transformation
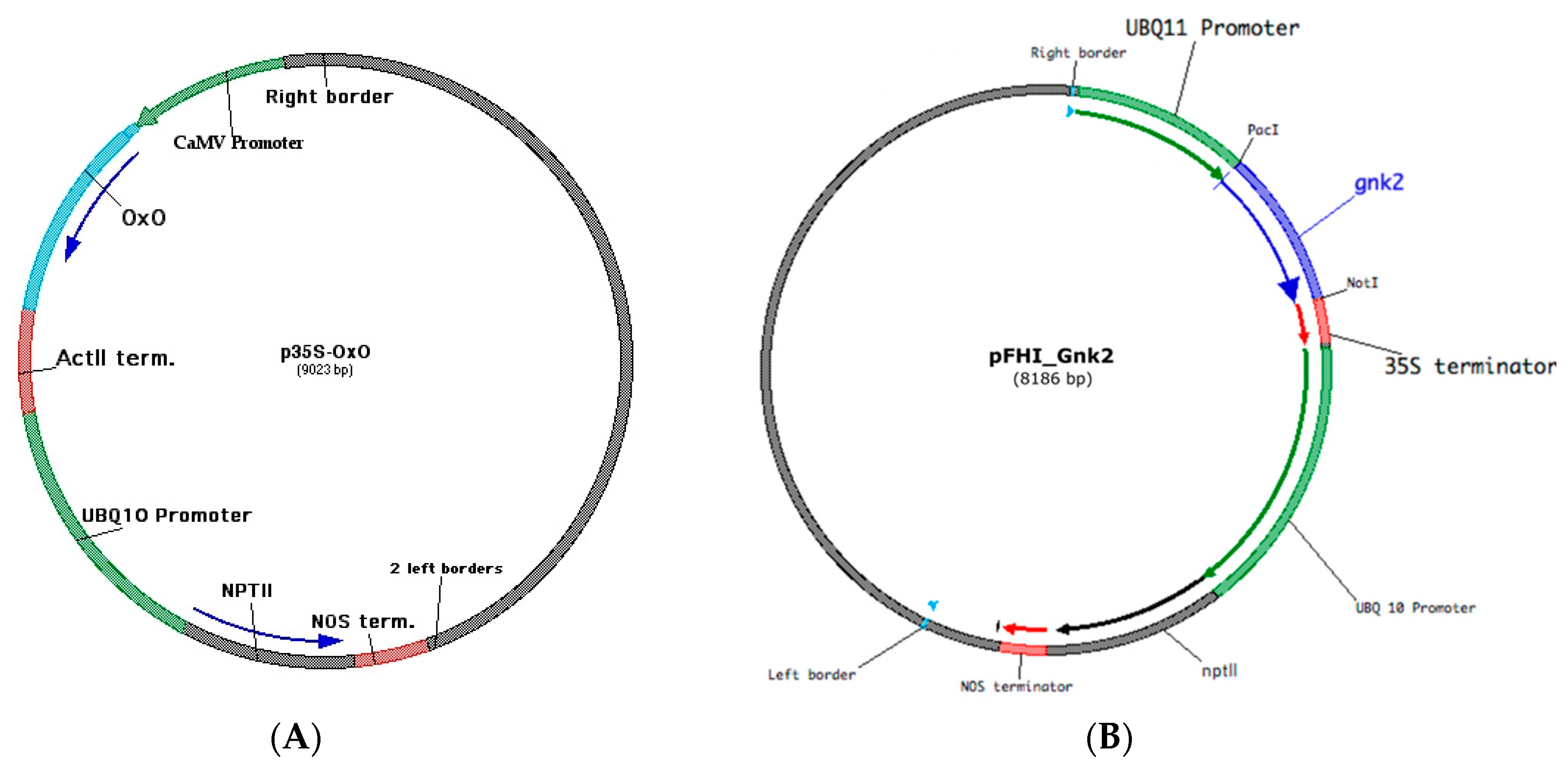
2.2. Plasmid DNA
2.2.1. p35S-OxO for Blight Tolerance
2.2.2. pFHI_Gnk2 for Putative Phytophthora-Tolerance
2.3. Transformation Protocol
2.3.1. Transformation of American Chestnut for Blight Tolerance
2.3.2. Transformation of American Chestnut for Putative Phytophthora-Tolerance
2.4. Multiplication, Rooting, and Acclimatization of Regenerated Shoots Expressing OxO
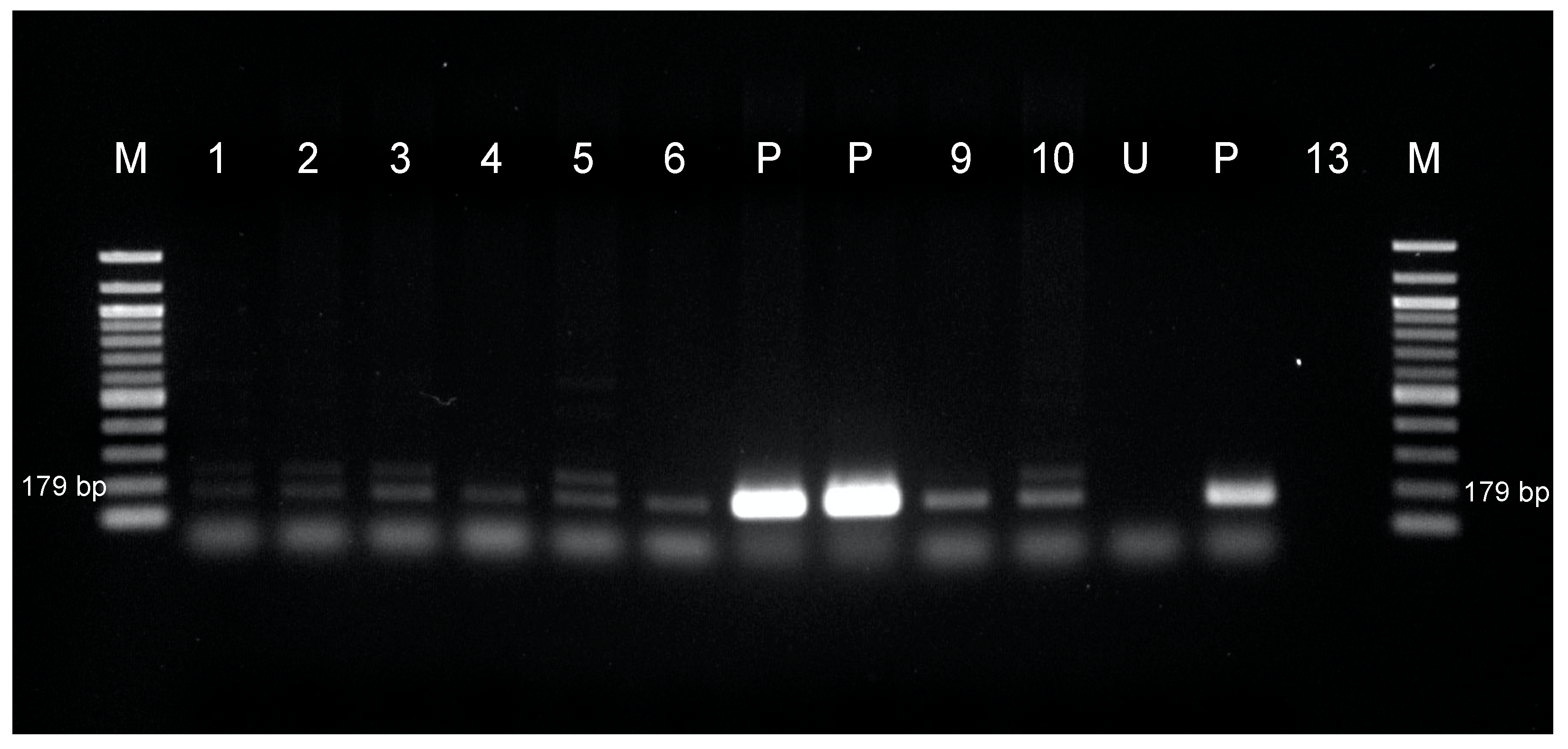
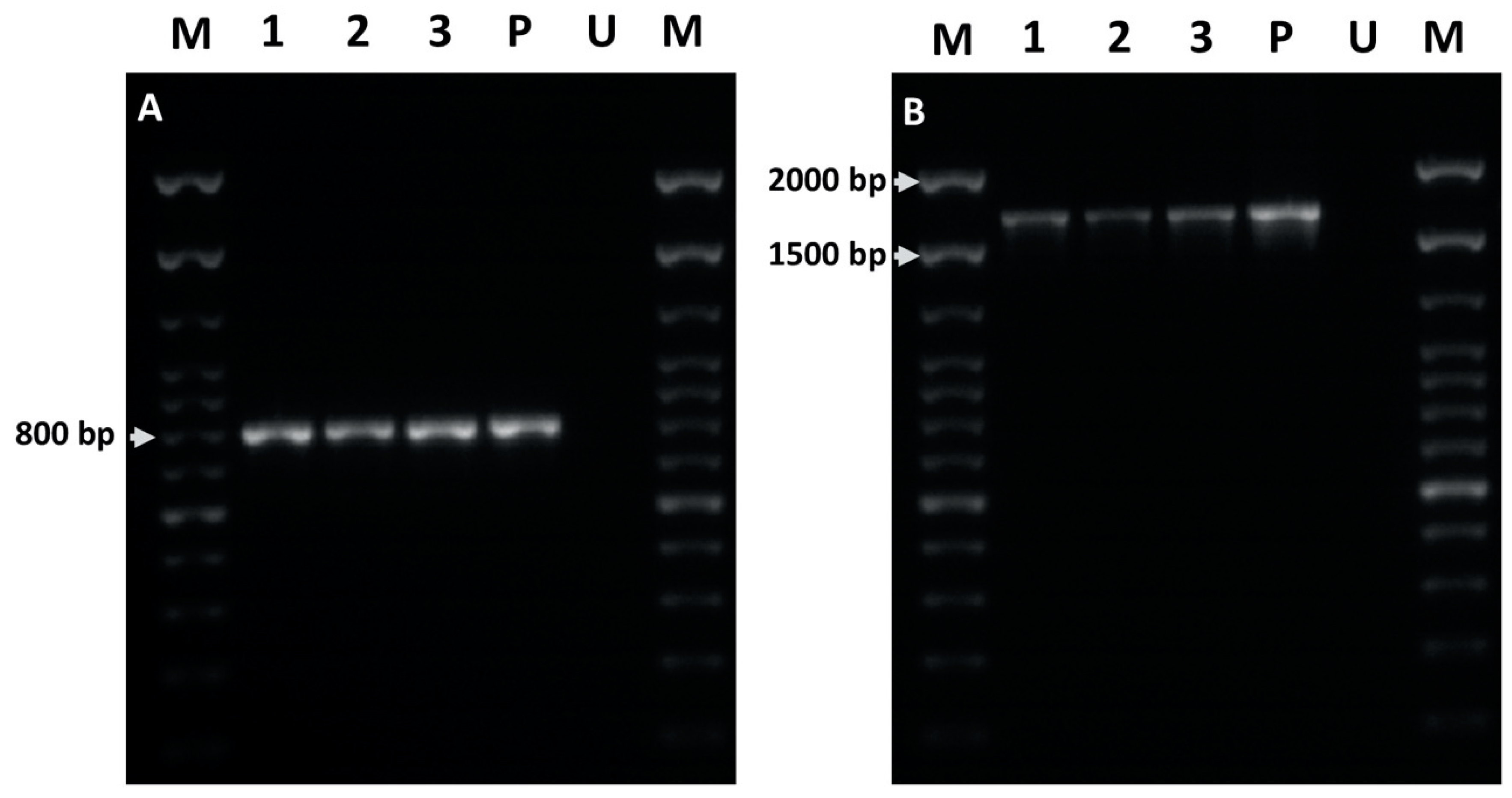
2.5. PCR Analysis
2.6. Data Analysis
3. Results
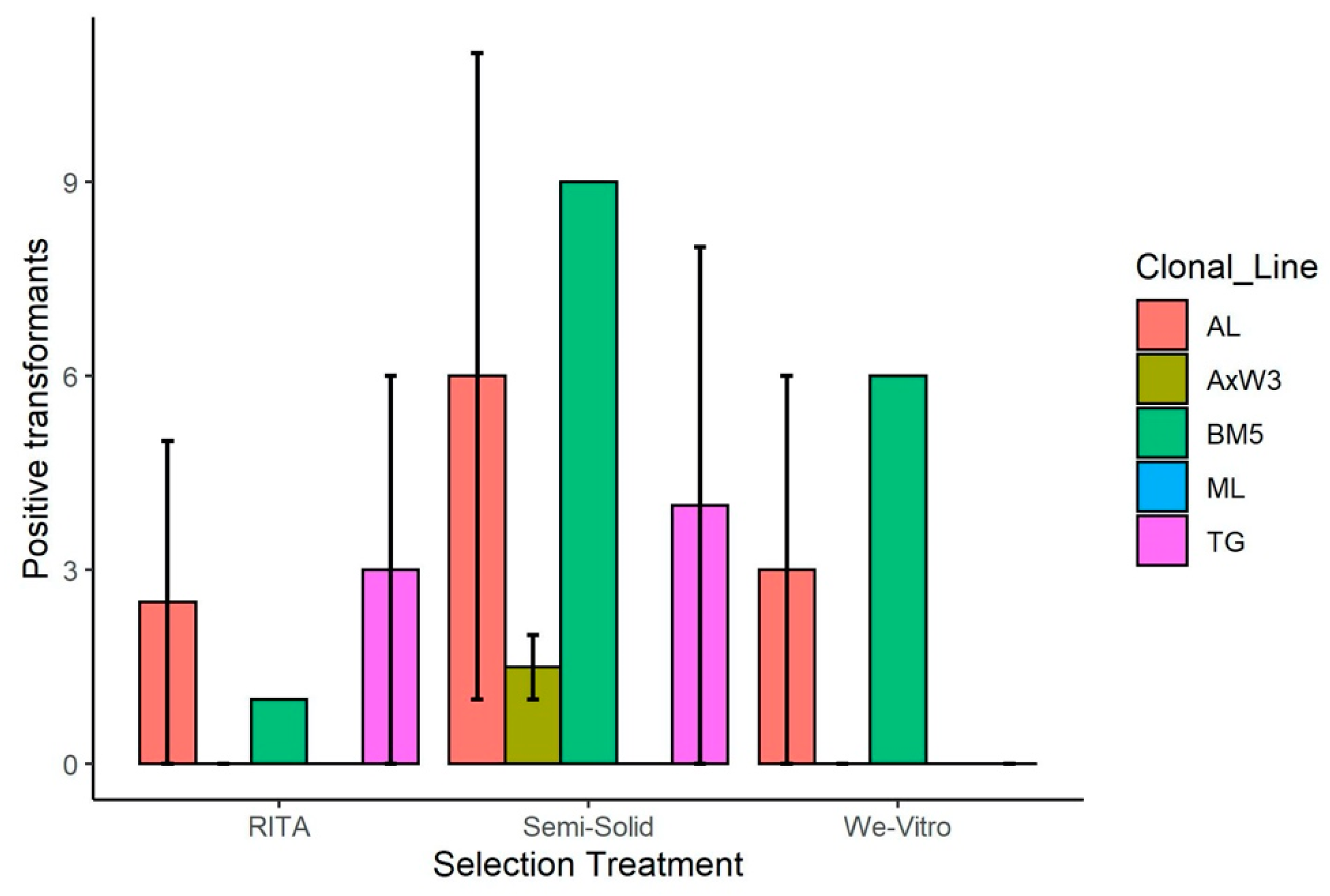
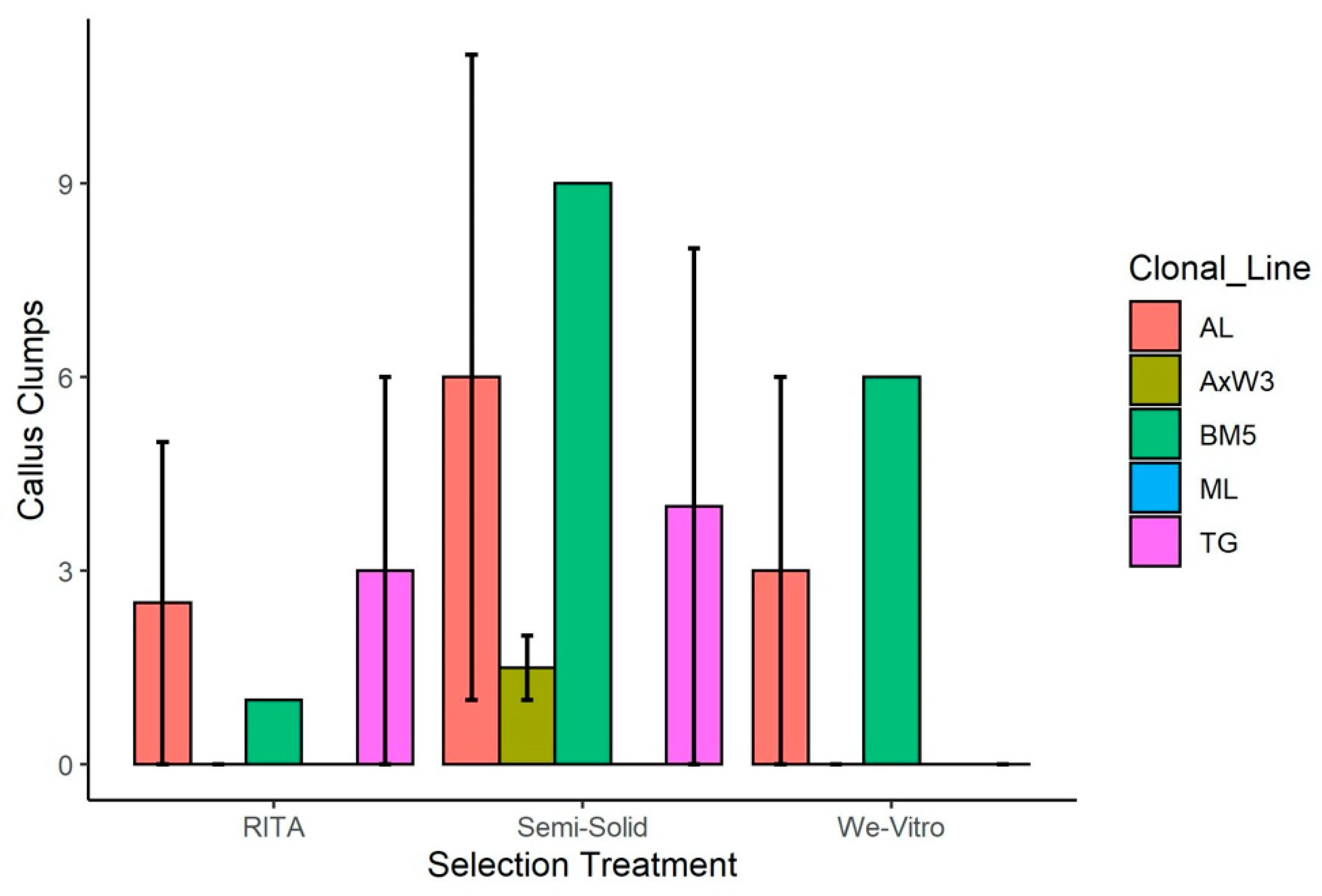
3.1. Transformations
3.1.1. Transformation of American Chestnut for Blight Tolerance
3.1.2. Transformation of American Chestnut for Putative Phytophthora-Tolerance
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rigling, D.; Prospero, S. Cryphonectria parasitica, the Causal Agent of Chestnut Blight: Invasion History, Population Biology and Disease Control. Mol. Plant Pathol. 2018, 19, 7–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anagnostakis, S.L. Chestnut Breeding in the United States for Disease and Insect Resistance. Plant Dis. 2012, 96, 1392–1403. [Google Scholar] [CrossRef] [PubMed]
- Steiner, K.C.; Westbrook, J.W.; Hebard, F.V.; Georgi, L.L.; Powell, W.A.; Fitzsimmons, S.F. Rescue of American Chestnut with Extraspecific Genes Following Its Destruction by a Naturalized Pathogen. New For. 2017, 48, 317–336. [Google Scholar] [CrossRef] [Green Version]
- Westbrook, J.W.; Zhang, Q.; Mandal, M.K.; Jenkins, E.V.; Barth, L.E.; Jenkins, J.W.; Grimwood, J.; Schmutz, J.; Holliday, J.A. Optimizing Genomic Selection for Blight Resistance in American Chestnut Backcross Populations: A Trade-off with American Chestnut Ancestry Implies Resistance Is Polygenic. Evol. Appl. 2020, 13, 31–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newhouse, A.E.; Powell, W.A. Intentional Introgression of a Blight Tolerance Transgene to Rescue the Remnant Population of American Chestnut. Conserv. Sci. Pract. 2020, in press. [Google Scholar]
- Powell, W.A.; Newhouse, A.E.; Coffey, V. Developing Blight-Tolerant American Chestnut Trees. Cold Spring Harb. Perspect. Biol. 2019, 11, a034587. [Google Scholar] [CrossRef] [Green Version]
- Carraway, D.T.; Merkle, S.A. Plantlet Regeneration from Somatic Embryos of American Chestnut. Can. J. For. Res. 1997, 27, 1805–1812. [Google Scholar] [CrossRef]
- Maynard, C.A.; McGuigan, L.D.; Oakes, A.D.; Zhang, B.; Newhouse, A.E.; Northern, L.C.; Chartrand, A.M.; Will, L.R.; Baier, K.M.; Powell, W.A. Chestnut, American (Castanea dentata (Marsh.) Borkh.). In Agrobacterium Protocols; Methods in Mol Biol; Springer: New York, NY, USA, 2015; pp. 143–161. [Google Scholar] [CrossRef]
- Lloyd, G.; McCown, B. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. Comb. Proc. Int. Plant Propagators’ Soc. 1980, 30, 421–427. [Google Scholar]
- Nitsch, J.P.; Nitsch, C. Haploid Plants from Pollen Grains. Science 1969, 163, 85–87. [Google Scholar] [CrossRef]
- Merkle, S.A.; Wiecko, A.T.; Watson-Pauley, B.A. Somatic Embryogenesis in American Chestnut. Can. J. For. Res. 1991, 21, 1698–1701. [Google Scholar] [CrossRef]
- Zhang, B.; Oakes, A.D.; Newhouse, A.E.; Baier, K.M.; Maynard, C.A.; Powell, W.A. A Threshold Level of Oxalate Oxidase Transgene Expression Reduces Cryphonectria parasitica-induced Necrosis in a Transgenic American Chestnut (Castanea dentata) Leaf Bioassay. Transgenic Res. 2013, 22, 973–982. [Google Scholar] [CrossRef] [Green Version]
- Guilley, H.; Dudley, R.K.; Jonard, G.; Balàzs, E.; Richards, K.E. Transcription of Cauliflower Mosaic Virus DNA: Detection of Promoter Sequences, and Characterization of Transcripts. Cell 1982, 30, 763–773. [Google Scholar] [CrossRef]
- Fraley, R.T.; Rogers, S.G.; Horsch, R.B.; Gelvin, S.B. Genetic Transformation in Higher Plants. Crit. Rev. Plant Sci. 1986, 4, 1–46. [Google Scholar] [CrossRef]
- Velten, J.; Schell, J. Selection-expression Plasmid Vectors for Use in Genetic Transformation of Higher Plants. Nucleic Acid Res. 1985, 13, 6981–6998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newhouse, A.; Coffey, V.; McGuigan, L.; Oakes, A.; Breda, K.; Matthews, D.; Drake, J.; Dougherty, J.; French, J.; Braverman, M.; et al. Petition for Determination of Nonregulated Status for Blight-Tolerant Darling 58 American Chestnut (Castanea dentata). 2020. Available online: https://beta.regulations.gov/document/APHIS-2020-0030-0002 (accessed on 28 September 2020).
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning. In A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar] [CrossRef]
- Oakes, A.D.; Pilkey, H.C.; Powell, W.A. Improving Ex Vitro Rooting and Acclimatization Techniques for Micropropagated American Chestnut. J. Environ. Hortic. 2020, in press. [Google Scholar]
- Oakes, A.D.; Desmarais, T.; Powell, W.A.; Maynard, C.A. Improving Rooting and Shoot Tip Survival of Micropropagated Transgenic American Chestnut Shoots. HortScience 2016, 51, 171–176. [Google Scholar] [CrossRef]
- Oakes, A.D.; Desmarais, T.R.; Powell, W.A.; Maynard, C.A. Ex Vitro Rooting of American Chestnut Improves Acclimatization Survival and Plantlet Quality. J. Environ. Hortic. 2016, 34, 75–79. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://www.R-project.org/ (accessed on 5 October 2020).
- Ogle, D.H.; Wheeler, P.; Dinno, A. FSA: Fisheries Stock Analysis. R Package Version 0.8.30. Available online: https://github.com/droglenc/FSA (accessed on 5 October 2020).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer: New York, NY, USA, 2016; Available online: https://ggplot2.tidyverse.org (accessed on 5 October 2020).
- Barakat, A.; DiLoreto, D.S.; Zhang, Y.; Smith, C.; Baier, K.; Powell, W.A.; Wheeler, N.; Sederoff, R.; Carlson, J.E. Comparison of the Transcriptomes of American Chestnut (Castanea dentata) and Chinese Chestnut (Castanea mollissima) in Response to the Chestnut Blight Infection. BMC Plant Biol. 2009, 9, 51. [Google Scholar] [CrossRef] [Green Version]
- Barakat, A.; Staton, M.; Cheng, C.-H.; Park, J.; Yassin, N.B.M.; Ficklin, S.; Yeh, C.-C.; Hebard, F.; Baier, K.; Powell, W.; et al. Chestnut Resistance to the Blight Disease: Insights from Transcriptome Analysis. BMC Plant Biol. 2012, 12, 38. [Google Scholar] [CrossRef] [Green Version]
- Serrazina, S.; Santos, C.; Machado, H.; Pesquita, C.; Vicentini, R.; Pais, M.S.; Sebastiana, M.; Costa, R. Castanea Root Transcriptome in Response to Phytophthora cinnamomi Challenge. Tree Genet. Genomes 2015, 11, 6. [Google Scholar] [CrossRef]
- Kubisiak, T.; Nelson, C.; Staton, M.; Zhebentyayeva, T.; Smith, C.; Olukolu, B.; Fang, G.-C.; Hebard, F.; Anagnostakis, S.; Wheeler, N.; et al. A Transcriptome-based Genetic Map of Chinese Chestnut (Castanea mollissima) and Identification of Regions of Segmental Homology with Peach (Prunus persica). Tree Genet. Genomes 2013, 9, 557–571. [Google Scholar] [CrossRef] [Green Version]
- Santos, C.; Nelson, C.D.; Zhebentyayeva, T.; Machado, H.; Gomes-Laranjo, J.; Costa, R.L. First Interspecific Genetic Linkage Map for Castanea sativa × Castanea crenata Revealed QTLs for Resistance to Phytophthora cinnamomi. PLoS ONE 2017, 12, e0184381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhebentyayeva, T.N.; Sisco, P.H.; Georgi, L.L.; Jeffers, S.N.; Perkins, M.T.; James, J.B.; Hebard, F.V.; Saski, C.; Nelson, C.D.; Abbott, A.G. Dissecting Resistance to Phytophthora cinnamomi in Interspecific Hybrid Chestnut Crosses Using Sequence-based Genotyping and QTL Mapping. Phytopathology 2019, 109, 1594–1604. [Google Scholar] [CrossRef] [PubMed]
- Polin, L.D.; Liang, H.; Rothrock, R.E.; Nishii, M.; Diehl, D.L.; Newhouse, A.E.; Joseph Nairn, C.; Powell, W.A.; Maynard, C.A. Agrobacterium-mediated Transformation of American Chestnut (Castanea Dentata (Marsh.) Borkh.) Somatic Embryos. Plant Cell Tissue Organ Cult. 2006, 84, 69–79. [Google Scholar] [CrossRef]
- Rothrock, R.E.; Polin-McGuigan, L.D.; Newhouse, A.E.; Powell, W.A.; Maynard, C.A. Plate Flooding as an Alternative Agrobacterium-mediated Transformation Method for American Chestnut Somatic Embryos. Plant Cell Tissue Organ Cult. 2007, 88, 93–99. [Google Scholar] [CrossRef]
- Andrade, G.M.; Nairn, C.J.; Le, H.T.; Merkle, S.A. Sexually Mature Transgenic American Chestnut Trees via Embryogenic Suspension-based Transformation. Plant Cell Rep. 2009, 28, 1385–1397. [Google Scholar] [CrossRef]
- Corredoira, E.; José, M.C.S.; Vieitez, A.M.; Allona, I.; Aragoncillo, C.; Ballester, A. Agrobacterium -mediated Transformation of European Chestnut Somatic Embryos with a Castanea sativa (Mill.) Endochitinase Gene. New For. 2016, 47, 669–684. [Google Scholar] [CrossRef]
- Mallón, R.; Vieitez, A.M.; Vidal, N. High-efficiency Agrobacterium-mediated Transformation in Quercus robur: Selection by Use of a Temporary Immersion System and Assessment by Quantitative PCR. Plant Cell Tissue Organ Cult. 2013, 114, 171–185. [Google Scholar] [CrossRef] [Green Version]
- Sawano, Y.; Miyakawa, T.; Yamazaki, H.; Tanokura, M.; Hatano, K. Purification, Characterization, and Molecular Gene Cloning of an Antifungal Protein from Ginkgo biloba Seeds. Biol. Chem. 2007, 388. [Google Scholar] [CrossRef]
- Wang, H.; Ng, T.B. Ginkbilobin, a Novel Antifungal Protein from Ginkgo biloba Seeds with Sequence Similarity to Embryo-Abundant Protein. Biochem. Biophys. Res. Commun. 2000, 279, 407–411. [Google Scholar] [CrossRef]
- Gao, N.; Wadhwani, P.; Mühlhäuser, P.; Liu, Q.; Riemann, M.; Ulrich, A.S.; Nick, P. An Antifungal Protein from Ginkgo biloba Binds Actin and Can Trigger Cell Death. Protoplasma 2016, 253, 1159–1174. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.; Duarte, S.; Tedesco, S.; Fevereiro, P.; Costa, R.L. Expression Profiling of Castanea Genes During Resistant and Susceptible Interactions with the Oomycete Pathogen Phytophthora cinnamomi Reveal Possible Mechanisms of Immunity. Front. Plant Sci. 2017, 8, 515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, P.; Machado, M.H.; Silva, M.C.; Costa, R.L. Histopathological Study Reveals New Insights into Responses of Chestnut (Castanea spp.) to Root Infection by Phytophthora cinnamomi. Phytopathology 2020. [Google Scholar] [CrossRef] [PubMed]
- Xing, Z.; Satchwell, M.F.; Powell, W.A.; Maynard, C.A. Micropropagation of American Chestnut: Increasing Rooting Rate and Preventing Shoot-Tip Necrosis. In Vitro Cell. Dev. Biol. Plant 1997, 33, 43–48. [Google Scholar] [CrossRef]
- Lapierre, S. Studies in American Chestnut (Castanea Dentata Marsh. (Borhk.)) Micropropagation and Acclimatization. Master’s Thesis, State University of New York College of Environmental Science and Forestry, Syracuse, NY, USA, January 2003. [Google Scholar]
- Oakes, A.D.; Powell, W.A.; Maynard, C.A. Doubling Acclimatization Survival of Micropropagated American Chestnuts with Darkness and Shortened Rooting Induction Time. J. Environ. Hortic. 2013, 31, 77–83. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial | Clonal Line | Treatment | Starting Mass (mg) | Positive Events | Escapes |
|---|---|---|---|---|---|
| 1 | AL | RITA | 50 | 3 | 5 |
| 2 | AL | RITA | 110 | 10 | 0 |
| 1 | AxW3 | RITA | 37 | 6 | 0 |
| 2 | AxW3 | RITA | 113 | 1 | 0 |
| 1 | BM5 | RITA | 98 | 0 | 1 |
| 2 | ML | RITA | 141 | 4 | 0 |
| 1 | TG | RITA | 86 | 8 | 6 |
| 2 | TG | RITA | 133 | 8 | 0 |
| 1 | AL | Semisolid | 86 | 8 | 11 |
| 2 | AL | Semisolid | 120 | 6 | 1 |
| 1 | AxW3 | Semisolid | 108 | 6 | 2 |
| 2 | AxW3 | Semisolid | 135 | 5 | 1 |
| 1 | BM5 | Semisolid | 65 | 8 | 9 |
| 2 | ML | Semisolid | 136 | 6 | 0 |
| 1 | TG | Semisolid | 59 | 8 | 8 |
| 2 | TG | Semisolid | 92 | 3 | 0 |
| 1 | AL | We Vitro | 65 | 3 | 6 |
| 2 | AL | We Vitro | 121 | 9 | 0 |
| 1 | AxW3 | We Vitro | 36 | 0 | 0 |
| 2 | AxW3 | We Vitro | 98 | 6 | 0 |
| 1 | BM5 | We Vitro | 94 | 6 | 6 |
| 2 | ML | We Vitro | 157 | 3 | 0 |
| 1 | TG | We Vitro | 63 | 4 | 0 |
| 2 | TG | We Vitro | 113 | 2 | 0 |
| Clonal Line | Vector | Starting Mass (mg) | Putative Events |
|---|---|---|---|
| AxW3 | pFHI_Gnk2 | 144 | 5 |
| SH4 | pFHI_Gnk2 | 103 | 0 |
| SH11 | pFHI_Gnk2 | 113 | 2 |
| Ellis #1 | pFHI_Gnk2 | 168 | 2 |
| pFHI_Gnk2 | 180 | 5 | |
| pFHI_GFP | 168 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McGuigan, L.; Fernandes, P.; Oakes, A.; Stewart, K.; Powell, W. Transformation of American Chestnut (Castanea dentata (Marsh.) Borkh) Using RITA® Temporary Immersion Bioreactors and We Vitro Containers. Forests 2020, 11, 1196. https://doi.org/10.3390/f11111196
McGuigan L, Fernandes P, Oakes A, Stewart K, Powell W. Transformation of American Chestnut (Castanea dentata (Marsh.) Borkh) Using RITA® Temporary Immersion Bioreactors and We Vitro Containers. Forests. 2020; 11(11):1196. https://doi.org/10.3390/f11111196
Chicago/Turabian StyleMcGuigan, Linda, Patrícia Fernandes, Allison Oakes, Kristen Stewart, and William Powell. 2020. "Transformation of American Chestnut (Castanea dentata (Marsh.) Borkh) Using RITA® Temporary Immersion Bioreactors and We Vitro Containers" Forests 11, no. 11: 1196. https://doi.org/10.3390/f11111196
APA StyleMcGuigan, L., Fernandes, P., Oakes, A., Stewart, K., & Powell, W. (2020). Transformation of American Chestnut (Castanea dentata (Marsh.) Borkh) Using RITA® Temporary Immersion Bioreactors and We Vitro Containers. Forests, 11(11), 1196. https://doi.org/10.3390/f11111196