Timing is Not Everything: Assessing the Efficacy of Pre- Versus Post-Harvest Herbicide Applications in Mitigating the Burgeoning Birch Phenomenon in Regenerating Hardwood Stands

Abstract
1. Introduction
2. Methods
2.1. Study Area
2.2. Vegetation Surveys
2.3. Statistical Analyses
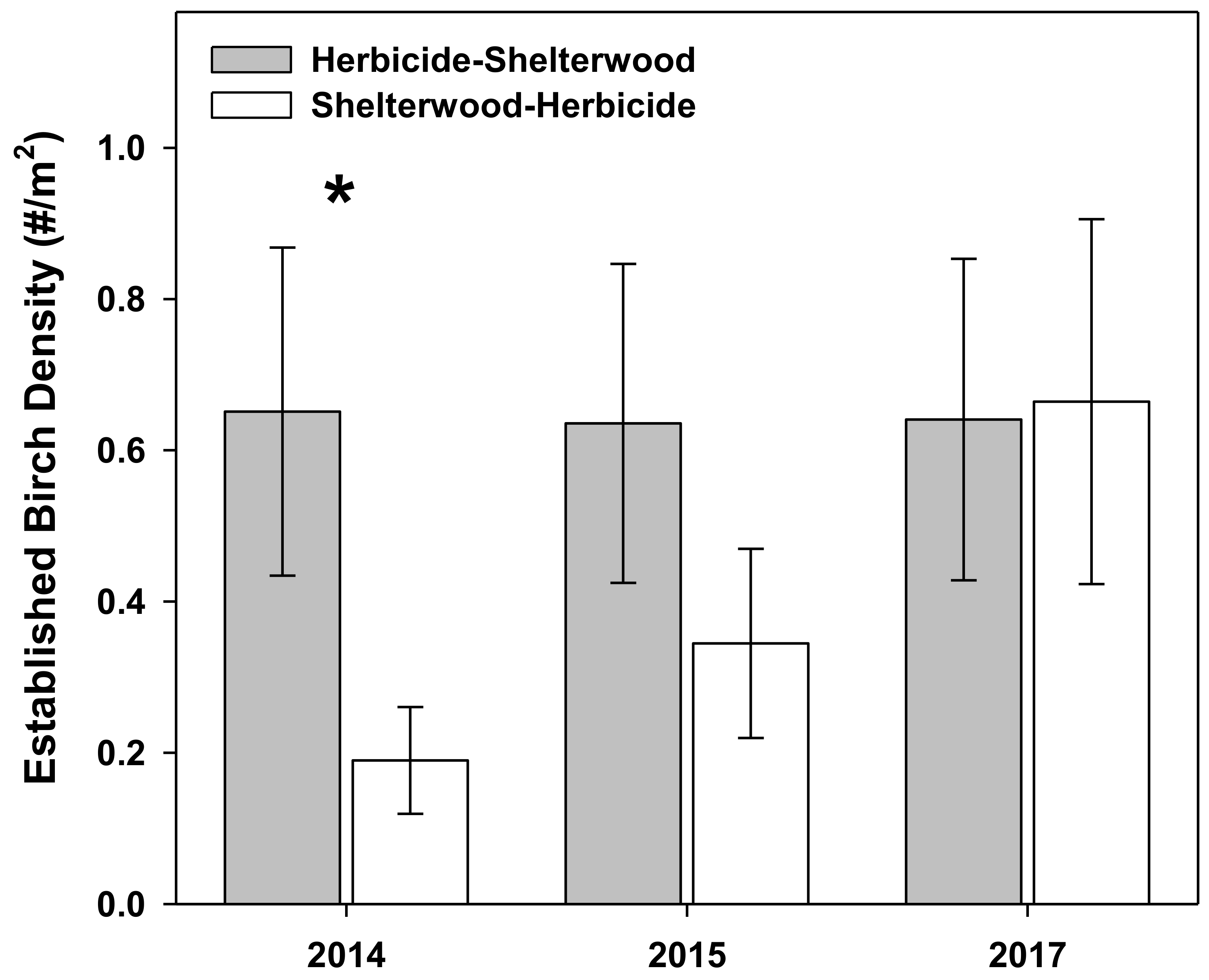
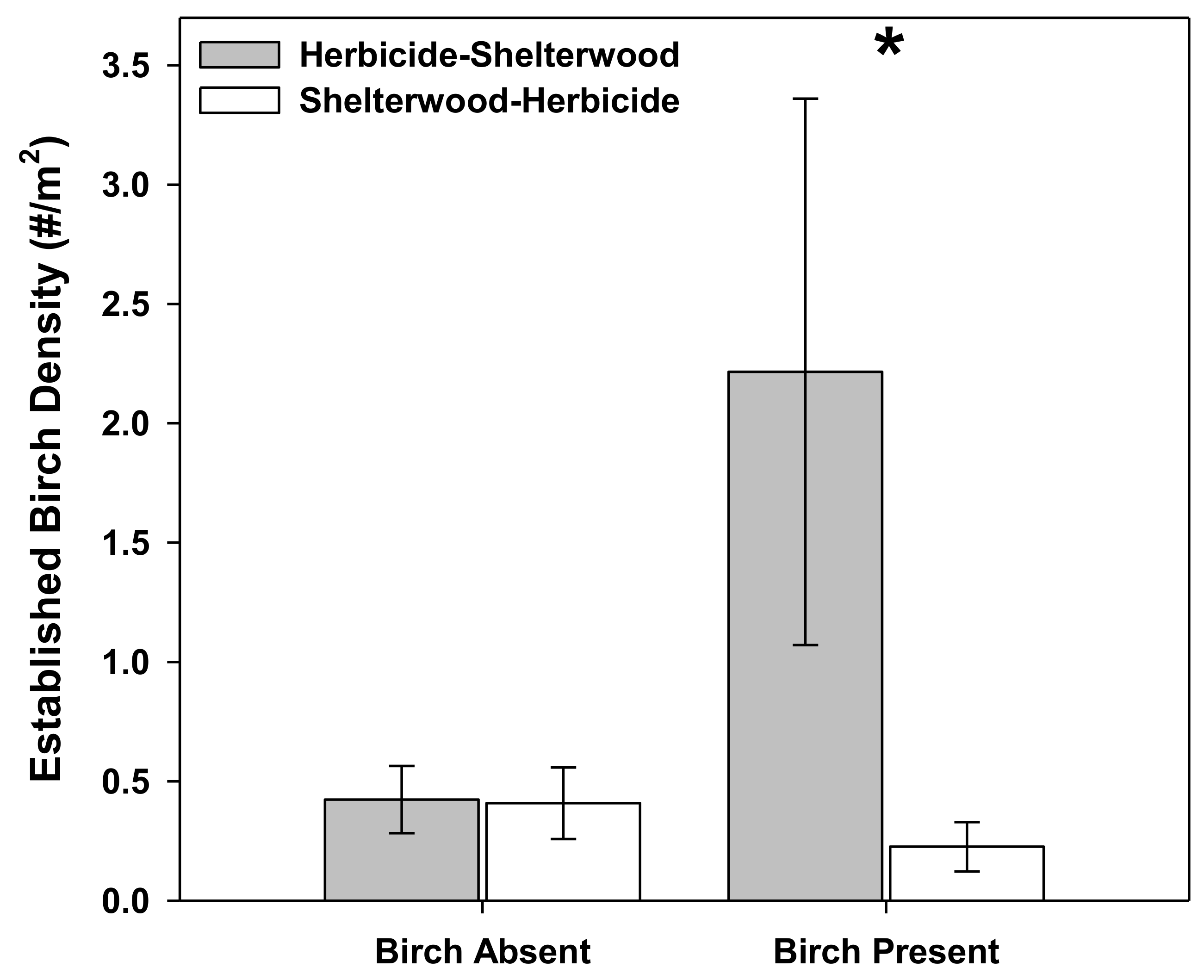
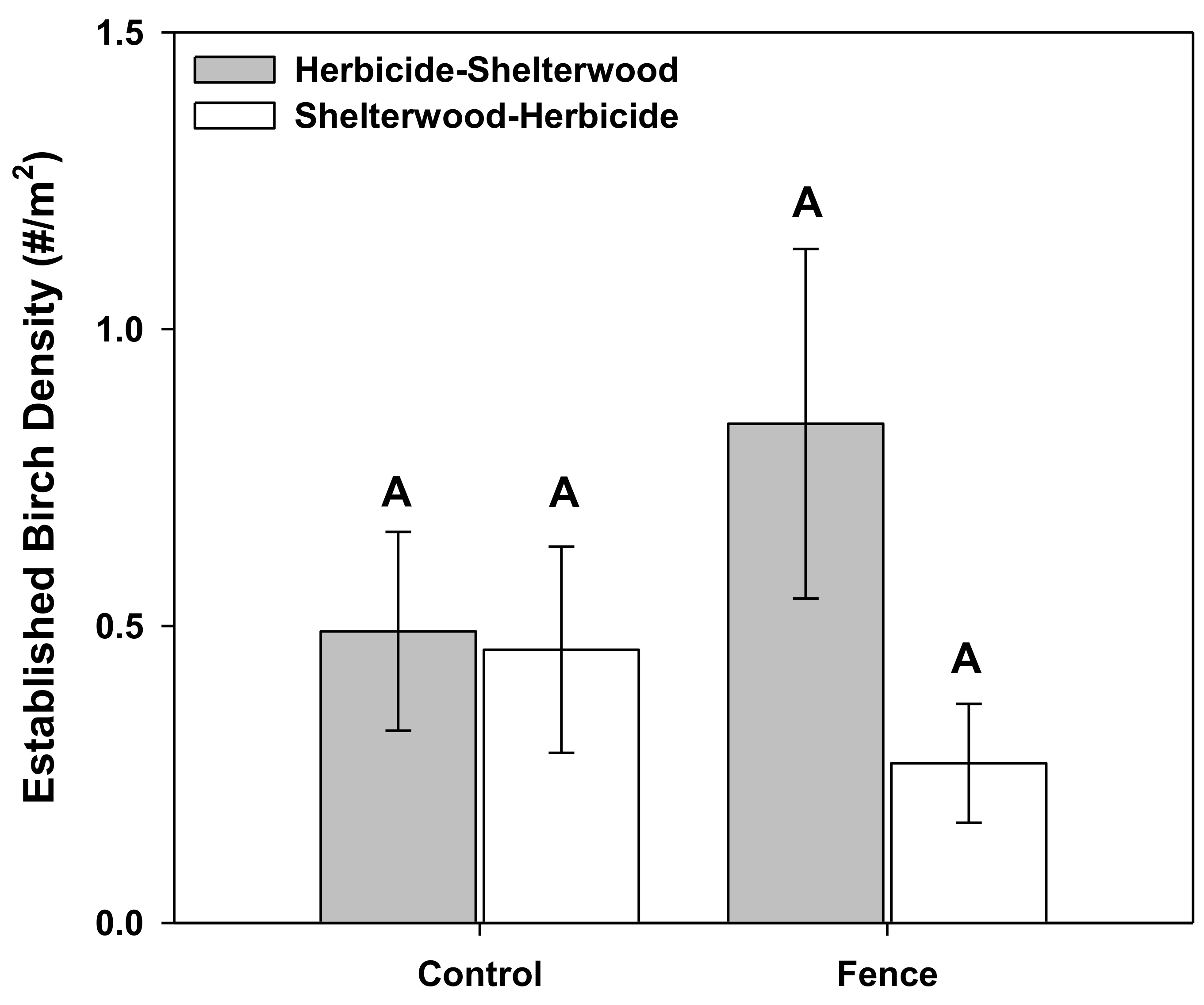
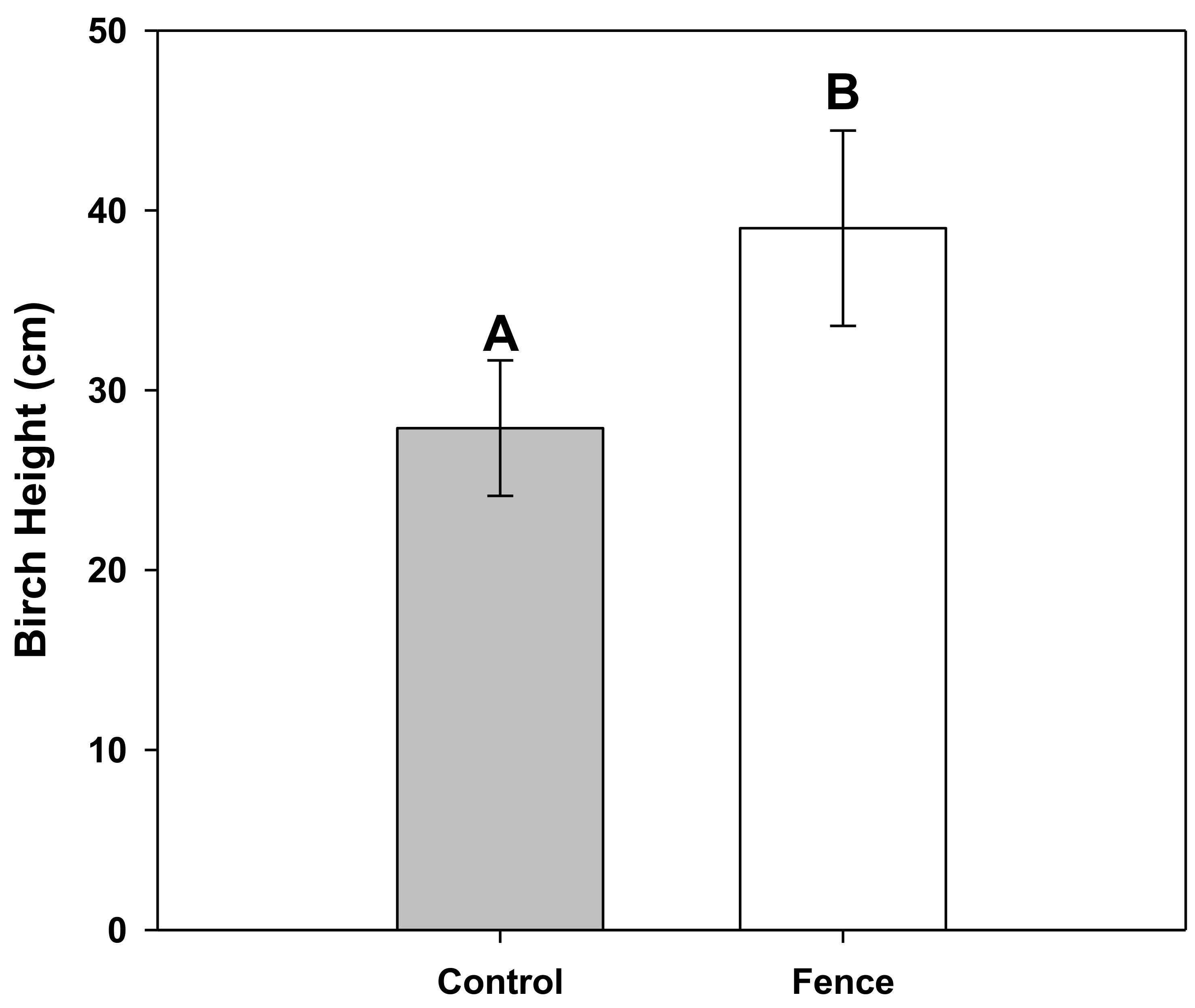
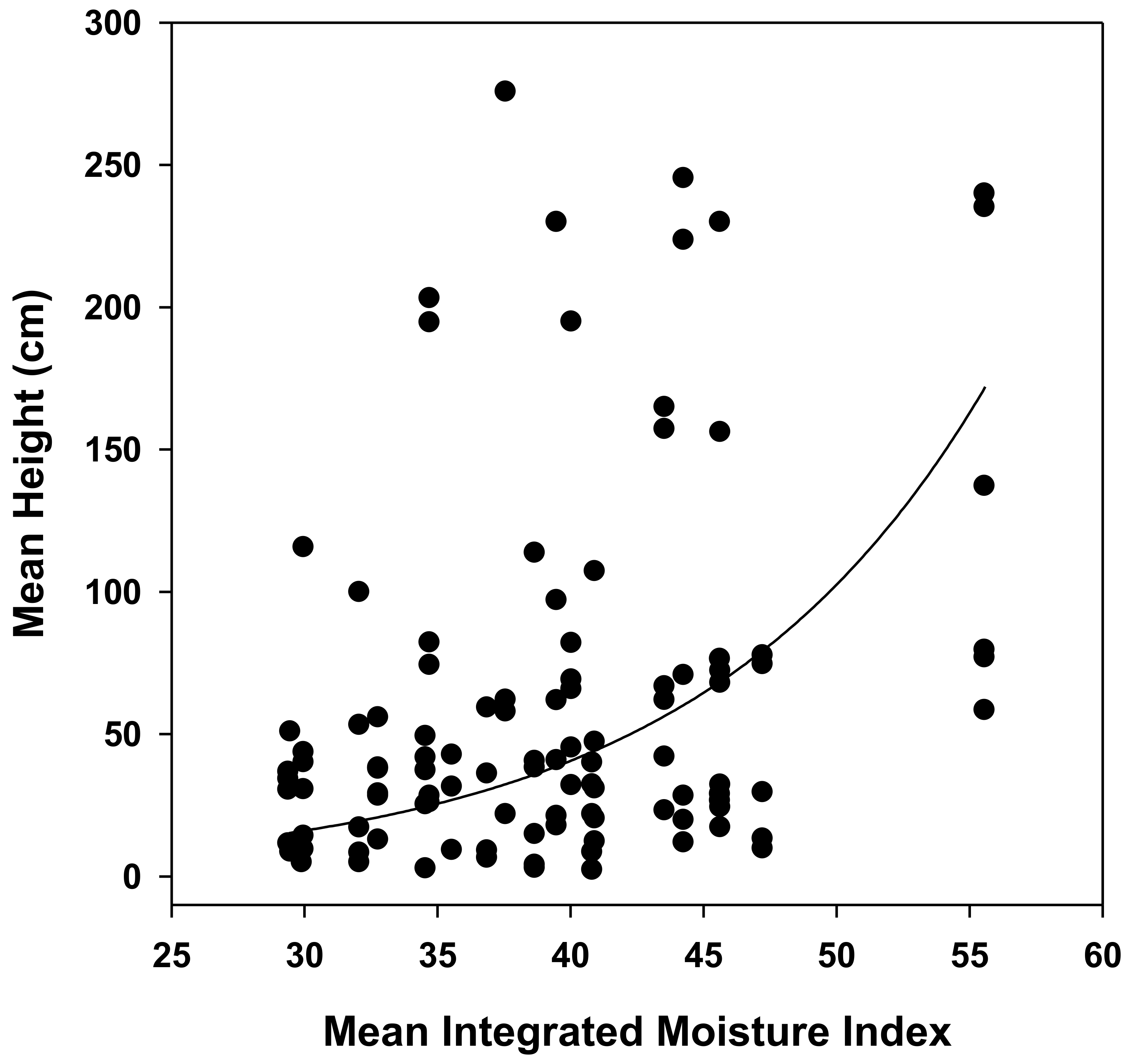
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Sequence | Browsing | IMI | Basal Area | Relative Density | Overstory Birch | Herbicide Year | Harvest Date | Elevation | Slope | Aspect |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Balanton | HS | Exclosure | 29.4 | 17.9 | 42 | Absent | 2009 | 2012 | 640.9 | 5.2 | south |
| Control | 15.7 | 37 | Absent | ||||||||
| Bump Run | HS | Exclosure | 36.9 | 27.2 | 87 | Absent | 2012 | 2013 | 548.1 | 15.5 | west |
| Control | 18.0 | 63 | Present | ||||||||
| BWAC-C9 | HS | Control | 34.6 | 15.6 | 28 | Absent | 2011 | 2012 | 700.2 | 3.3 | west |
| Exclosure | 12.2 | 34 | Absent | ||||||||
| Cash Crop | HS | Control | 45.6 | 9.6 | 41 | Absent | 2009 | 2013 | 640.2 | 6.7 | northeast |
| Exclosure | 20.5 | 30 | Absent | ||||||||
| Close Call | HS | Exclosure | 34.7 | 14.7 | 35 | Absent | 2010 | 2013 | 610.5 | 3.4 | south |
| Control | 22.3 | 52 | Present | ||||||||
| Potter11 | HS | Control | 44.3 | 19.3 | 51 | Absent | 2012 | 2013 | 641.0 | 2.2 | north |
| Exclosure | 21.4 | 52 | Absent | ||||||||
| Rush | HS | Exclosure | 35.6 | 20.2 | 64 | Absent | 2010 | 2014 | 667.6 | 18.2 | southeast |
| Control | 18.9 | 42 | Absent | ||||||||
| ScreamingEagle | HS | Control | 37.6 | 18.4 | 31 | Absent | 2009 | 2012 | 670.9 | 2.6 | north |
| Exclosure | 12.8 | 48 | Absent | ||||||||
| Second Look | HS | Exclosure | 40.9 | 17.1 | 35 | Absent | 2012 | 2013 | 532.6 | 4.1 | north |
| Control | 20.3 | 57 | Present | ||||||||
| ShakenBake | HS | Control | 43.6 | 24.1 | 66 | Present | 2009 | 2012 | 564.1 | 25.9 | southeast |
| Exclosure | 19.9 | 69 | Present | ||||||||
| Sorry About That | HS | Exclosure | 45.7 | 15.6 | 37 | Absent | 2010 | 2012 | 608.3 | 26.5 | northwest |
| Control | 13.3 | 30 | Absent | ||||||||
| Treed Bear | HS | Control | 55.6 | 21.0 | 39 | Absent | 2009 | 2012 | 531.6 | 14.2 | north |
| Exclosure | 22.9 | 39 | Absent | ||||||||
| BloodyRun | SH | Exclosure | 29.5 | 30.2 | 64 | Absent | 2012 | 2009 | 569.2 | 4.1 | south |
| Control | 20.0 | 48 | Present | ||||||||
| Bradford 40 | SH | Control | 38.7 | 10.6 | 44 | Absent | 2012 | 2011 | 669.9 | 0.2 | north |
| Exclosure | 19.7 | 28 | Absent | ||||||||
| BuntsRun | SH | Exclosure | 29.9 | 16.3 | 34 | Absent | 2012 | 2008 | 544.6 | 2.8 | south |
| Control | 23.2 | 50 | Absent | ||||||||
| Compressor | SH | Control | 32.1 | 21.6 | 46 | Present | 2012 | 2012 | 538.6 | 29.8 | south |
| Exclosure | 17.0 | 62 | Absent | ||||||||
| First Hunt | SH | Exclosure | 30.0 | 28.1 | 53 | Absent | 2012 | 2011 | 513.7 | 2.9 | southeast |
| Control | 24.1 | 48 | Absent | ||||||||
| Irvine Run | SH | Exclosure | 40.1 | 14.9 | 47 | Absent | 2012 | 2010 | 513.9 | 1.6 | east |
| Control | 14.6 | 27 | Absent | ||||||||
| McKean 37 | SH | Control | 47.2 | 18.5 | 49 | Present | 2012 | 2010 | 676.7 | 7.8 | east |
| Exclosure | 16.9 | 46 | Absent | ||||||||
| Potter6 | SH | Control | 40.8 | 15.3 | 36 | Absent | 2012 | 2012 | 687.8 | 4.8 | northeast |
| Exclosure | 17.2 | 39 | Present | ||||||||
| Regen 134 | SH | Exclosure | 32.8 | 19.3 | 38 | Present | 2012 | 2005 | 555.3 | 3.2 | southeast |
| Control | 31.8 | 61 | Present | ||||||||
| Spring Creek | SH | Exclosure | 39.5 | 22.8 | 44 | Absent | 2012 | 2011 | 544.4 | 3.6 | southeast |
| Control | 28.5 | 57 | Absent |
Appendix B
| (A) All Desirable Seedling Density 2017 | ||
| Effect | F-Value | p-Value |
| Sequence | F1,20.0 = 0.25 | 0.624 |
| Exclosure | F1,20.0 = 2.34 | 0.142 |
| Exclosure × Sequence | F1,20.0 = 0.07 | 0.798 |
| (B) All Desirable Seedling Height 2017 | ||
| Effect | F-Value | p-Value |
| Sequence | F1,20.0 = 0.00 | 0.984 |
| Exclosure | F1,20.0 = 1.75 | 0.201 |
| Exclosure × Sequence | F1,20.0 = 0.00 | 0.993 |
References
- Albright, T.A.; McWilliams, W.H.; Widmann, R.H.; Butler, B.J.; Crocker, S.J.; Kurtz, C.M.; Lehman, S.; Lister, T.W.; Miles, P.D.; Morin, R.S.; et al. Pennsylvania Forests 2014. Resource Bulletin. NRS-111; USDA Forest Service, Northern Research Station: Newtown Square, PA, USA, 2017.
- Widmann, R.H.; Crawford, S.; Kurtz, C.M.; Nelson, M.D.; Miles, P.D.; Morin, R.S.; Rienmann, R. New York Forests, 2012. Resource Bulletin NRS-98; USDA Forest Service, Northern Research Station: Newtown Square, PA, USA, 2015.
- Kizlinski, M.L.; Orwig, D.A.; Cobb, R.C.; Foster, D.R. Direct and indirect ecosystem consequences of an invasive pest on forests dominated by eastern hemlock. J. Biogeogr. 2002, 29, 1489–1503. [Google Scholar] [CrossRef]
- Royo, A.A.; Peterson, C.J.; Stanovick, J.S.; Carson, W.P. Evaluating the ecological impacts of salvage logging: Can natural and anthropogenic disturbances promote coexistence? Ecology 2016, 97, 1566–1582. [Google Scholar] [CrossRef]
- Ward, J.S.; Stephens, G.R. Influence of crown class on survival and development of Betula lenta in Connecticut, U.S.A. Can. J. For. Res. 1996, 26, 277–288. [Google Scholar] [CrossRef]
- Fei, S.; Gould, P.J.; Steiner, K.C.; Finley, J.C.; McDill, M.E. Forest regeneration composition and development in upland, mixed-oak forests. Tree Physiol. 2005, 25, 1495–1500. [Google Scholar] [CrossRef]
- Plotkin, A.B.; Foster, D.; Carlson, J.; Magill, A. Survivors, not invaders, control forest development following simulated hurricane. Ecology 2013, 94, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Morin, R.S.; Cook, G.W.; Barnett, C.J.; Butler, B.J.; Crocker, S.J.; Hatfield, M.A.; Kurtz, C.M.; Lister, T.W.; Luppold, W.G.; McWilliams, W.H.; et al. West Virginia Forests 2013. Resource Bulletin NRS-105; USDA Forest Service, Northern Research Station: Newtown Square, PA, USA, 2016.
- Abrams, M.D. The red maple paradox: What explains the widespread expansion of red maple in eastern forests? Bioscience 1998, 48, 355–364. [Google Scholar] [CrossRef]
- Burns, R.M.; Honkala, B.H. (Eds.) Silvics of North America; United States Department of Agriculture, Forest Service: Washington, DC, USA, 1990.
- Baker, F.S. A revised tolerance table. J. For. 1949, 47, 179–181. [Google Scholar]
- de la Cretaz, A.L.; Kelty, M.J. Establishment and control of hay-scented fern: A native invasive species. Biol. Invasions 1999, 1, 223–236. [Google Scholar] [CrossRef]
- Horsley, S.B.; Marquis, D.A. Interference by weeds and deer with Allegheny (Pennsylvania, USA) hardwood reproduction. Can. J. For. Res. 1983, 13, 61–69. [Google Scholar] [CrossRef]
- Matlack, G.R. Secondary dispersal of seed across snow in Betula lenta, a gap-colonizing tree species. J. Ecol. 1989, 77, 853–869. [Google Scholar] [CrossRef]
- Godman, R.M.; Krefting, L.W. Factors important to yellow birch establishment in Upper Michigan. Ecology 1960, 41, 18–28. [Google Scholar] [CrossRef]
- Marquis, D.A. Regeneration of Birch and Associated Hardwoods after Patch Cutting. Research Paper NE-32; USDA Forest Service, Northeastern Forest Experiment Station: Upper Darby, PA, USA, 1964.
- Bressette, J.W.; Beck, H.; Beauchamp, V.B. Beyond the browse line: Complex cascade effects mediated by white-tailed deer. Oikos 2012, 121, 1749–1760. [Google Scholar] [CrossRef]
- Horsley, S.B.; Stout, S.L.; deCalesta, D.S. White-tailed deer impact on the vegetation dynamics of a northern hardwood forest. Ecol. Appl. 2003, 13, 98–118. [Google Scholar] [CrossRef]
- Nuttle, T.; Royo, A.A.; Adams, M.B.; Carson, W.P. Historic disturbance regimes promote tree diversity only under low browsing regimes in eastern deciduous forest. Ecol. Monogr. 2013, 83, 3–17. [Google Scholar] [CrossRef]
- Fownes, J.H.; Harrington, R.A. Seedling response to gaps: Separating effects of light and nitrogen. For. Ecol. Manag. 2004, 203, 297–310. [Google Scholar] [CrossRef]
- Crabtree, R.C.; Bazzaz, F.A. Seedling response of four birch species to simulated nitrogen deposition: Ammonium vs. nitrate. Ecol. Appl. 1993, 3, 315–321. [Google Scholar] [CrossRef]
- Falxa-Raymond, N.; Patterson, A.E.; Schuster, W.S.F.; Griffin, K.L. Oak loss increases foliar nitrogen, δ15N and growth rates of Betula lenta in a northern temperate deciduous forest. Tree Physiol. 2012, 32, 1092–1101. [Google Scholar] [CrossRef]
- Mou, P.; Fahey, T.J.; Hughes, J.W. Effects of soil disturbance on vegetation recovery and nutrient accumulation following whole-tree harvest of a northern hardwood ecosystem. J. Appl. Ecol. 1993, 30, 661–675. [Google Scholar] [CrossRef]
- Marquis, D.A.; Ernst, R.L.; Stout, S.L. (Eds.) Prescribing Silvicultural Treatments in Hardwood Stands of the Alleghenies (Revised), Gen. Tech. Rep. NE-96; USDA Forest Service Northeastern Research Station: Radnor, PA, USA, 1992; p. 101.
- Hannah, P.R. Regeneration of northern hardwoods in the northeast with the shelterwood method. North J. Appl. For. 1991, 8, 99–104. [Google Scholar]
- Horsley, S.B. Regeneration success and plant species diversity of Allegheny hardwood stands after Roundup application and shelterwood cutting. N. J. Appl. For. 1994, 11, 109–116. [Google Scholar]
- Nyland, R.D. (Ed.) Silviculture—Concepts and Applications, 2nd ed.; McGraw-Hill: New York, NY, USA, 2002; p. 682.
- Ristau, T.E.; Horsley, S.B. When is pin cherry (Prunus pensylvanica L.) a problem in Allegheny hardwoods? North J. Appl. For. 2006, 23, 204–210. [Google Scholar]
- Bailey, S.W.; Horsley, S.B.; Long, R.P. Thirty years of change in forest soils of the Allegheny Plateau, Pennsylvania. Soil Sci. Soc. Am. J. 2005, 69, 681–690. [Google Scholar] [CrossRef]
- Redding, J. History of deer population trends and forest cutting on the Allegheny National Forest. In Proceedings of the 10th Central Hardwood Forest Conference; Gottschalk, K.W., Fosbroke, S.L.C., Eds.; Gen. Tech. Rep. NE-197; USDA Forest Service: Morgantown, WV, USA, 1995; pp. 214–224. [Google Scholar]
- Horsley, S.B.; McCormick, L.H.; Groninger, J.W. Effects of timing of Oust application on survival of hardwood seedlings. North. Jour. Appl. For. 1992, 9, 22–27. [Google Scholar]
- Iverson, L.R.; Dale, M.E.; Scott, C.T.; Prasad, A. A GIS-derived integrated moisture index to predict forest composition and productivity of Ohio forests (USA). Landsc. Ecol. 1997, 12, 331–348. [Google Scholar] [CrossRef]
- Stout, S.L.; Nyland, R.D. Role of species composition in relative density measurement in Allegheny hardwoods. Can. J. For. Res. 1986, 16, 574–579. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS System for Windows, 9.4; SAS Institute Inc.: Cary, NC, USA, 2013. [Google Scholar]
- Littell, R.C.; Milliken, G.A.; Stroup, W.W.; Wolfinger, R.D.; Schabenberger, O. SAS for Mixed Models, 2nd ed.; SAS Institute, Inc.: Cary, NC, USA, 2006. [Google Scholar]
- Brown, M.B.; Forsythe, A.B. Robust tests for the equality of variances. J. Am. Stat. Assoc. 1974, 69, 364–367. [Google Scholar] [CrossRef]
- Anderson, J.J.; Dulka, J.J. Environmental fate of sulfometuron methyl in aerobic soils. J. Agric. Food Chem. 1985, 33, 596–602. [Google Scholar] [CrossRef]
- Harvey, J.J.; Dulka, J.J.; Anderson, J.J. Properties of sulfometuron methyl affecting its environmental fate: Aqueous hydrolysis and photolysis, mobility and adsorption on soils, and bioaccumulation potential. J. Agric. Food Chem. 1985, 33, 590–596. [Google Scholar] [CrossRef]
- Ostrofsky, W.D.; McCormack, J.M.L. Silvicultural management of beech and the beech bark disease. N. J. Appl. For. 1986, 3, 89–91. [Google Scholar] [CrossRef]
- Bose, A.K.; Wagner, R.G.; Roth, B.E.; Weiskittel, A.R. Influence of browsing damage and overstory cover on regeneration of American beech and sugar maple nine years following understory herbicide release in central Maine. New For. 2018, 49, 67–85. [Google Scholar] [CrossRef]
- Ristau, T.E.; Stoleson, S.H.; Horsley, S.B.; deCalesta, D.S. Ten-year response of the herbaceous layer to an operational herbicide-shelterwood treatment in a northern hardwood forest. For. Ecol. Manag. 2011, 262, 970–979. [Google Scholar] [CrossRef]
- Kelty, M.J.; Nyland, R.D. Regenerating Adirondack northern hardwoods by shelterwood cutting and control of deer density. J. For. 1981, 79, 22–26. [Google Scholar]
- Ray, D.G.; Nyland, R.D.; Yanai, R.D. Patterns of early cohort development following shelterwood cutting in three Adirondack northern hardwood stands. For. Ecol. Manag. 1999, 119, 1–11. [Google Scholar] [CrossRef]
- Nelson, A.S.; Wagner, R.G. Improving the composition of beech-dominated northern hardwood understories in northern Maine. N. J. Appl. For. 2011, 28, 186–193. [Google Scholar] [CrossRef]
- Bell, F.W.; Newmaster, S.G. The effects of silvicultural disturbances on the diversity of seed-producing plants in the boreal mixedwood forest. Can. J. For. Res. 2002, 32, 1180–1191. [Google Scholar] [CrossRef]
- Krueger, L.M.; Peterson, C.J.; Royo, A.; Carson, W.P. Evaluating relationships among tree growth rate, shade tolerance, and browse tolerance following disturbance in an eastern deciduous forest. Can. J. For. Res. 2009, 39, 2460–2469. [Google Scholar] [CrossRef]
- Kern, C.C.; Reich, P.B.; Montgomery, R.A.; Strong, T.F. Do deer and shrubs override canopy gap size effects on growth and survival of yellow birch, northern red oak, eastern white pine, and eastern hemlock seedlings? For. Ecol. Manag. 2012, 267, 134–143. [Google Scholar] [CrossRef]
- Kain, M.; Battaglia, L.; Royo, A.; Carson, W.P. Over-browsing in Pennsylvania creates a depauperate forest dominated by an understory tree: Results from a 60-year-old deer exclosure. J. Torrey Bot. Soc. 2011, 138, 322–326. [Google Scholar] [CrossRef]
- Healy, W.M. Forage preferences of tame deer in a Northwest Pennsylvania clear-cutting. J. Wildl. Manag. 1971, 35, 717–723. [Google Scholar] [CrossRef]
- Tubbs, C.H. The Influence of Light, Moisture, and Seedbed on Yellow Birch Regeneration; Research Paper NC-27; US Dept. of Agriculture, Forest Service, North Central Forest Experiment Station: St. Paul, MN, USA, 1969.
- Ribbens, E.; Silander, J.A.; Pacala, S.W. Seedling recruitment in forests: Calibrating models to predict patterns of tree seedling dispersion. Ecology 1994, 75, 1794–1806. [Google Scholar] [CrossRef]
- Caspersen, J.P.; Saprunoff, M. Seedling recruitment in a northern temperate forest: The relative importance of supply and establishment limitation. Can. J. For. Res. 2005, 35, 978–989. [Google Scholar] [CrossRef]
- Houle, G. Seed dispersal and seedling recruitment of Betula alleghaniensis: Spatial inconsistency in time. Ecology 1998, 79, 807–818. [Google Scholar] [CrossRef]
- Iverson, L.R.; Prasad, A.M. A.M. A GIS-derived integrated moisture index. In Characteristics of Mixed Oak Forest Ecosystems in Southern Ohio Prior to the Reintroduction of Fire; Gen. Tech. Rep. NE-299; U.S. Department of Agriculture, Forest Service: Newtown Square, PA, USA, 2003; pp. 29–41. [Google Scholar]
- Prasad, A.M.; Iverson, L.R.; Peters, M.P.; Matthews, S.N. Climate Change Tree Atlas. Available online: http://www.nrs.fs.fed.us/atlas (accessed on 9 September 2018).
- Elliott, K.J.; Swank, W.T. Impacts of drought on tree mortality and growth in a mixed hardwood forest. J. Veg. Sci. 1994, 5, 229–236. [Google Scholar] [CrossRef]
- Nowacki, G.J.; Abrams, M.D. Community and edaphic analysis of mixed oak forests in the ridge and valley province of central Pennsylvania. In Proceedings of the 8th Central Hardwood Forest Conference, Radnor, PA, USA, 4–6 March 1991; Gen. Tech. Rep NE-148. pp. 247–260. [Google Scholar]
- Beaudet, M.; Messier, C. Growth and morphological responses of yellow birch, sugar maple, and beech seedlings growing under a natural light gradient. Can. J. For. Res. 1998, 28, 1007–1015. [Google Scholar] [CrossRef]
- Holmes, W.E.; Zak, D.R. Soil microbial control of nitrogen loss following clear-cut harvest in northern hardwood ecosystems. Ecol. Appl. 1999, 9, 202–215. [Google Scholar] [CrossRef]
- Catovsky, S.; Bazzaz, F.A. The role of resource interactions and seedling regeneration in maintaining a positive feedback in hemlock stands. J. Ecol. 2000, 88, 100–112. [Google Scholar] [CrossRef]
- Ristau, T.E.; Horsley, S.B. Pin cherry effects on Allegheny hardwood stand development. Can. J. For. Res. 1999, 29, 73–84. [Google Scholar] [CrossRef]





| Effect | F-Value | p-Value |
|---|---|---|
| Sequence | F1,23.5 = 0.01 | 0.942 |
| Exclosure | F1,21.4 = 0.00 | 1.000 |
| Exclosure × Sequence | F1,21.4 = 6.87 | 0.016 |
| Year | F2,69.2 = 7.48 | 0.001 |
| Sequence × Year | F2,69.2 = 7.97 | 0.001 |
| Exclosure × Year | F2,69.2 = 0.30 | 0.744 |
| Exclosure × Sequence × Year | F2,69.2 = 0.43 | 0.651 |
| Sequence × Seed Source | F2,29.1 = 6.02 | 0.006 |
| Effect | F-Value | p-Value |
|---|---|---|
| Sequence | F1,19.3 = 1.46 | 0.241 |
| Exclosure | F1,49.2 = 6.62 | 0.013 |
| Exclosure × Sequence | F1,49.2 = 0.00 | 0.999 |
| Year | F2,55.0 = 46.53 | <0001 |
| Sequence × Year | F2,55.0 = 0.42 | 0.658 |
| Exclosure × Year | F2,56.1 = 0.55 | 0.579 |
| Exclosure × Sequence × Year | F2,56.1 = 0.44 | 0.646 |
| Mean IMI | F1,19.2 = 6.25 | 0.022 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Royo, A.A.; Pinchot, C.C.; Stanovick, J.S.; Stout, S.L. Timing is Not Everything: Assessing the Efficacy of Pre- Versus Post-Harvest Herbicide Applications in Mitigating the Burgeoning Birch Phenomenon in Regenerating Hardwood Stands. Forests 2019, 10, 324. https://doi.org/10.3390/f10040324
Royo AA, Pinchot CC, Stanovick JS, Stout SL. Timing is Not Everything: Assessing the Efficacy of Pre- Versus Post-Harvest Herbicide Applications in Mitigating the Burgeoning Birch Phenomenon in Regenerating Hardwood Stands. Forests. 2019; 10(4):324. https://doi.org/10.3390/f10040324
Chicago/Turabian StyleRoyo, Alejandro A., Cornelia C. Pinchot, John S. Stanovick, and Susan L. Stout. 2019. "Timing is Not Everything: Assessing the Efficacy of Pre- Versus Post-Harvest Herbicide Applications in Mitigating the Burgeoning Birch Phenomenon in Regenerating Hardwood Stands" Forests 10, no. 4: 324. https://doi.org/10.3390/f10040324
APA StyleRoyo, A. A., Pinchot, C. C., Stanovick, J. S., & Stout, S. L. (2019). Timing is Not Everything: Assessing the Efficacy of Pre- Versus Post-Harvest Herbicide Applications in Mitigating the Burgeoning Birch Phenomenon in Regenerating Hardwood Stands. Forests, 10(4), 324. https://doi.org/10.3390/f10040324