Important Insect and Disease Threats to United States Tree Species and Geographic Patterns of Their Potential Impacts


Abstract
1. Introduction
2. Materials and Methods
2.1. Insect and Disease Agents and Severity Ratings
2.2. Geographic Patterns of Potential Forest Importance Value Loss
3. Results
3.1. Insect and Disease Agent Summary
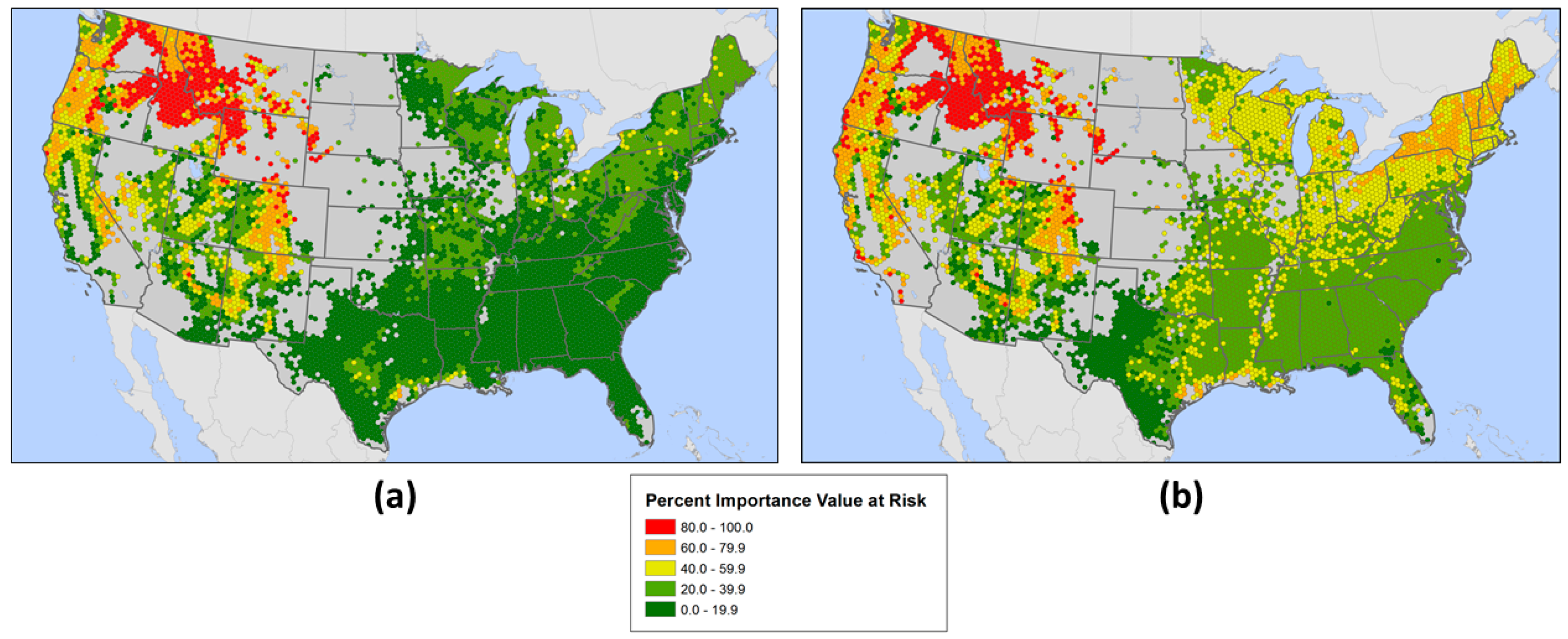
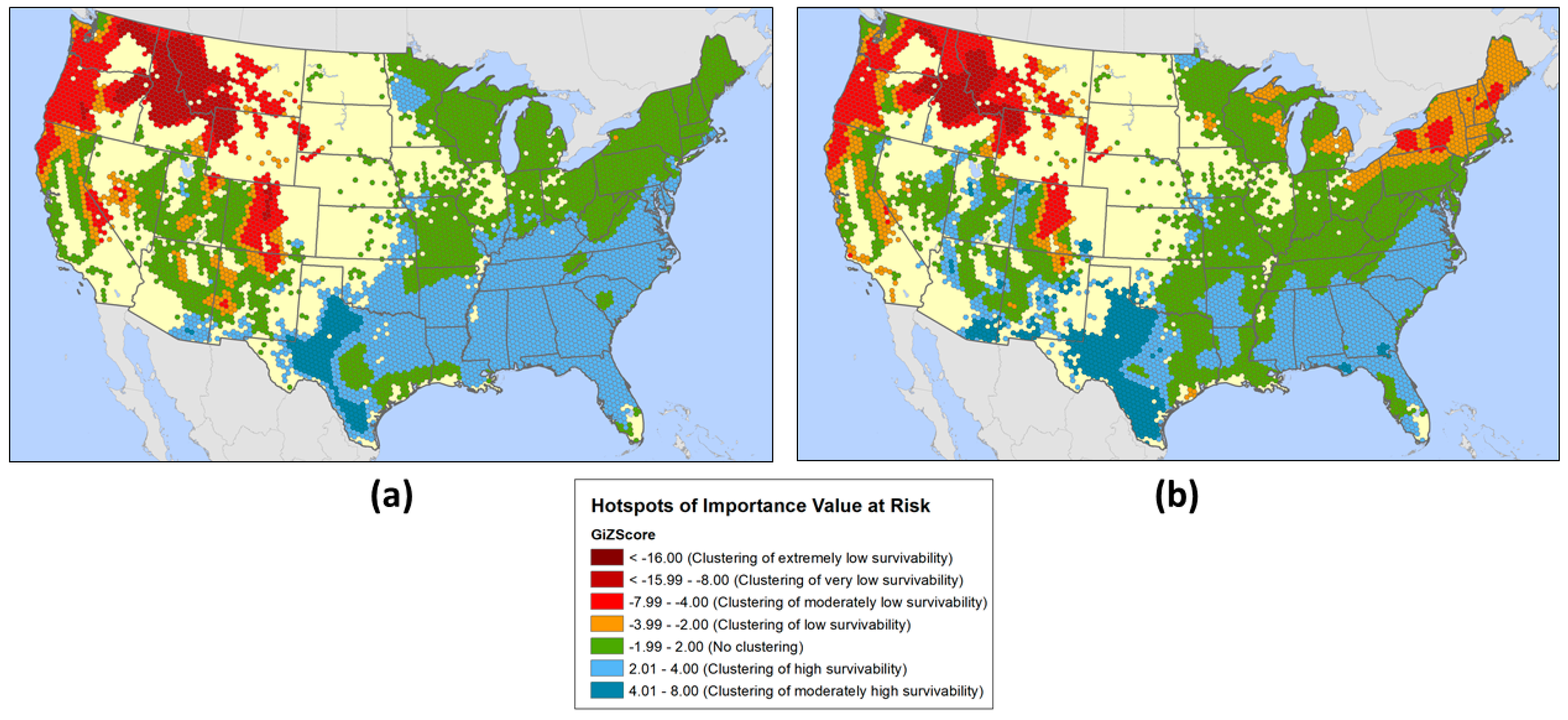
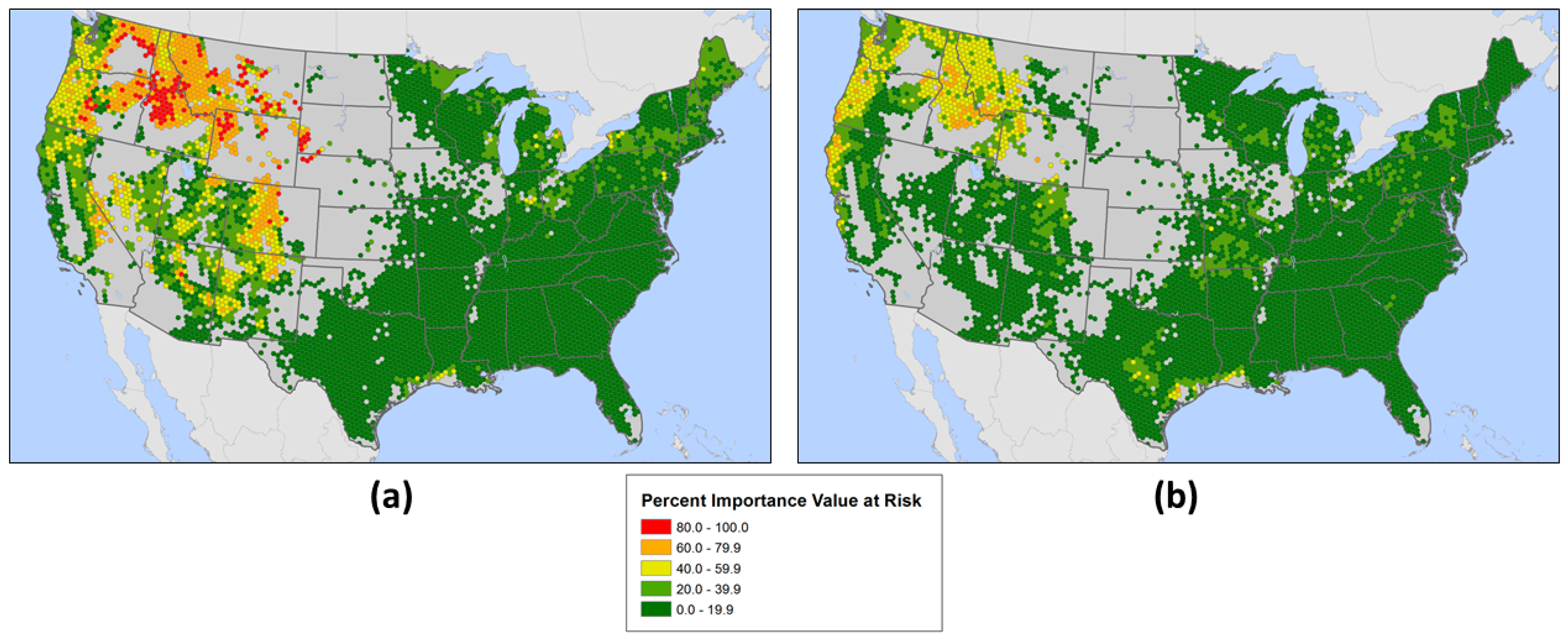
3.2. Geographic Patterns of Potential Insect and Disease Impacts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lovett, G.M.; Weiss, M.; Liebhold, A.M.; Holmes, T.P.; Leung, B.; Lambert, K.F.; Orwig, D.A.; Campbell, F.T.; Rosenthal, J.; McCullough, D.G.; et al. Nonnative forest insects and pathogens in the United States: Impacts and policy options. Ecol. Appl. 2016, 26, 1437–1455. [Google Scholar] [CrossRef] [PubMed]
- Tobin, P.C. Ecological Consequences of Pathogen and Insect Invasions. Curr. For. Rep. 2015, 1, 25–32. [Google Scholar] [CrossRef]
- Logan, J.A.; Regniere, J.; Powell, J.A. Assessing the impacts of global warming on forest pest dynamics. Front. Ecol. Environ. 2003, 1, 130–137. [Google Scholar] [CrossRef]
- Krist, F.J.; Ellenwood, J.R.; Woods, M.H.; McMahan, A.J.; Cowardin, J.P.; Ryerson, D.E.; Sapio, F.J.; Zweifler, M.O.; Romero, S.A. 2013–2027 National Insect and Disease Forest Risk Assessment; United States Department of Agriculture, Forest Service, Forest Health Technology Enterprise Team: Washington, DC, USA, 2014; p. 199.
- Teale, S.A.; Castello, J.D. The past as key to the future: A new perspective on forest health. In Forest Health: An Integrated Perspective; Castello, J.D., Teale, S.A., Eds.; Cambridge University Press: New York, NY, USA, 2011; pp. 3–16. [Google Scholar]
- Zhang, L.; Rubin, B.D.; Manion, P.D. Mortality: The essence of a healthy forest. In Forest Health: An Integrated Perspective; Castello, J.D., Teale, S.A., Eds.; Cambridge University Press: New York, NY, USA, 2011; pp. 17–49. [Google Scholar]
- Vorster, A.G.; Evangelista, P.H.; Stohlgren, T.J.; Kumar, S.; Rhoades, C.C.; Hubbard, R.M.; Cheng, A.S.; Elder, K. Severity of a mountain pine beetle outbreak across a range of stand conditions in Fraser Experimental Forest, Colorado, United States. For. Ecol. Manag. 2017, 389, 116–126. [Google Scholar] [CrossRef]
- Johnson, T.N.; Buskirk, S.W.; Hayward, G.D.; Raphael, M.G. Tree mortality after synchronized forest insect outbreaks: Effects of tree species, bole diameter, and cutting history. For. Ecol. Manag. 2014, 319, 10–17. [Google Scholar] [CrossRef]
- Windmuller-Campione, M.A.; Page, D.H.; Long, J.N. Does the practice of silviculture build resilience to the spruce beetle? A case study of treated and untreated spruce-fir stands in Northern Utah. J. For. 2017, 115, 559–567. [Google Scholar] [CrossRef]
- Dukes, J.S.; Pontius, J.; Orwig, D.; Garnas, J.R.; Rodgers, V.L.; Brazee, N.; Cooke, B.; Theoharides, K.A.; Stange, E.E.; Harrington, R.; et al. Responses of insect pests, pathogens, and invasive plant species to climate change in the forests of northeastern North America: What can we predict? Can. J. For. Res. 2009, 39, 231–248. [Google Scholar] [CrossRef]
- Sturrock, R.N.; Frankel, S.J.; Brown, A.V.; Hennon, P.E.; Kliejunas, J.T.; Lewis, K.J.; Worrall, J.J.; Woods, A.J. Climate change and forest diseases. Plant Pathol. 2011, 60, 133–149. [Google Scholar] [CrossRef]
- Parry, D.; Teale, S.A. Alien invasions: The effects of introduced species on forest structure and function. In Forest Health: An Integrated Perspective; Castello, J.D., Teale, S.A., Eds.; Cambridge University Press: New York, NY, USA, 2011; pp. 115–162. [Google Scholar]
- Gandhi, K.J.K.; Herms, D.A. Direct and indirect effects of alien insect herbivores on ecological processes and interactions in forests of eastern North America. Biol. Invasions 2010, 12, 389–405. [Google Scholar] [CrossRef]
- Loo, J. Ecological impacts of non-indigenous invasive fungi as forest pathogens. Biol. Invasions 2009, 11, 81–96. [Google Scholar] [CrossRef]
- Havill, N.P.; Montgomery, M.E.; Keena, M.A. Hemlock woolly adelgid and its hemlock hosts: A global perspective. In Implementation and Status of Biological Control of the Hemlock Woolly Adelgid; Onken, B., Reardon, R., Eds.; U.S. Forest Service, Forest Health Technology Enterprise Team: Morgantown, WV, USA, 2011; Volume FHTET-2011-04, pp. 3–14. [Google Scholar]
- Klooster, W.S.; Gandhi, K.J.K.; Long, L.C.; Perry, K.I.; Rice, K.B.; Herms, D.A. Ecological impacts of emerald ash borer in forests at the epicenter of the invasion in North America. Forests 2018, 9, 250. [Google Scholar] [CrossRef]
- Hughes, M.A.; Riggins, J.J.; Koch, F.H.; Cognato, A.I.; Anderson, C.; Formby, J.P.; Dreaden, T.J.; Ploetz, R.C.; Smith, J.A. No rest for the laurels: symbiotic invaders cause unprecedented damage to southern USA forests. Biol. Invasions 2017, 19, 2143–2157. [Google Scholar] [CrossRef]
- Castello, J.D.; Leopold, D.J.; Smallidge, P.J. Pathogens, patterns, and processes in forest ecosystems. Bioscience 1995, 45, 16–24. [Google Scholar] [CrossRef]
- Potter, K.M.; Paschke, J.L. Large-scale patterns of insect and disease activity in the conterminous United States, Alaska, and Hawaii from the national insect and disease survey, 2015. In Forest Health Monitoring: National Status, Trends and Analysis, 2016; Potter, K.M., Conkling, B.L., Eds.; General Technical Report SRS-222; United States Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2017; pp. 21–42. [Google Scholar]
- Liebhold, A.M.; McCullough, D.G.; Blackburn, L.M.; Frankel, S.J.; Von Holle, B.; Aukema, J.E. A highly aggregated geographical distribution of forest pest invasions in the USA. Divers. Distrib. 2013, 19, 1208–1216. [Google Scholar] [CrossRef]
- Holdenrieder, O.; Pautasso, M.; Weisberg, P.J.; Lonsdale, D. Tree diseases and landscape processes: The challenge of landscape pathology. Trends Ecol. Evol. 2004, 19, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Early, R.; Bradley, B.A.; Dukes, J.S.; Lawler, J.J.; Olden, J.D.; Blumenthal, D.M.; Gonzalez, P.; Grosholz, E.D.; Ibanez, I.; Miller, L.P.; et al. Global threats from invasive alien species in the twenty-first century and national response capacities. Nat. Commun. 2016, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Yemshanov, D.; Koch, F.H.; McKenney, D.W.; Downing, M.C.; Sapio, F. Mapping Invasive Species Risks with Stochastic Models: A Cross-Border United States-Canada Application for Sirex noctilio Fabricius. Risk Anal. 2009, 29, 868–884. [Google Scholar] [CrossRef] [PubMed]
- Shatz, A.J.; Rogan, J.; Sangermano, F.; Miller, J.; Elmes, A. Modeling the risk of spread and establishment for Asian longhorned beetle (Anoplophora glabripennis) in Massachusetts from 2008–2009. Geocarto Int. 2016, 31, 813–831. [Google Scholar] [CrossRef]
- Vaclavik, T.; Kanaskie, A.; Hansen, E.M.; Ohmann, J.L.; Meentemeyer, R.K. Predicting potential and actual distribution of sudden oak death in Oregon: Prioritizing landscape contexts for early detection and eradication of disease outbreaks. For. Ecol. Manag. 2010, 260, 1026–1035. [Google Scholar] [CrossRef]
- Trotter, R.T.; Morin, R.S.; Oswalt, S.N.; Liebhold, A. Changes in the regional abundance of hemlock associated with the invasion of hemlock woolly adelgid (Adelges tsugae Annand). Biol. Invasions 2013, 15, 2667–2679. [Google Scholar] [CrossRef]
- Morin, R.S.; Liebhold, A.M.; Pugh, S.A.; Crocker, S.J. Regional assessment of emerald ash borer, Agrilus planipennis, impacts in forests of the Eastern United States. Biol. Invasions 2017, 19, 703–711. [Google Scholar] [CrossRef]
- Potter, K.M.; Crane, B.S.; Hargrove, W.W. A United States national prioritization framework for tree species vulnerability to climate change. New For. 2017, 48, 275–300. [Google Scholar] [CrossRef]
- Bechtold, W.A.; Patterson, P.L. The Enhanced Forest Inventory and Analysis Program: National Sampling Design and Estimation Procedures; Gen. Tech. Rep. SRS-80; USDA Forest Service, Southern Research Station: Asheville, NC, USA, 2005; 85p.
- Potter, K.M. Do United States protected areas effectively conserve forest tree rarity and evolutionary distinctiveness? Biol. Conserv. 2018, 224, 34–46. [Google Scholar] [CrossRef]
- Leung, B.; Springborn, M.R.; Turner, J.A.; Brockerhoff, E.G. Pathway-level risk analysis: The net present value of an invasive species policy in the US. Front. Ecol. Environ. 2014, 12, 273–279. [Google Scholar] [CrossRef]
- Gilbert, G.S.; Webb, C.O. Phylogenetic signal in plant pathogen-host range. Proc. Natl. Acad. Sci. USA 2007, 104, 4979–4983. [Google Scholar] [CrossRef]
- Horst, R.K. Westcott’s Plant Disease Handbook, 8th ed.; Springer: Dordrecht, The Netherlands, 2013; 826p. [Google Scholar]
- United States Department of Agriculture Forest Health Protection. Damage Causing Agent (DCA) List. Available online: https://www.fs.fed.us/foresthealth/applied-sciences/mapping-reporting/gis-spatial-analysis/digital-mobile-sketch-mapping.shtml (accessed on 13 November 2018).
- Woudenberg, S.W.; Conkling, B.L.; O’Connell, B.M.; LaPoint, E.B.; Turner, J.A.; Waddell, K.L. The Forest Inventory and Analysis Database: Database Description and Users Manual Version 4.0 for Phase 2; USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2010; 336p.
- Little, E.L. Atlas of United States Trees. Volume 1. Conifers and Important Hardwoods; United States Department of Agriculture Forest Service: Washington, DC, USA, 1971; 9p, 200 maps.
- Viereck, L.A.; Little, E.L. Atlas of United States Trees. Volume 2. Alaska Trees and Common Shrubs; United States Department of Agriculture Forest Service: Washington, DC, USA, 1975; 19p, 105 maps.
- Little, E.L. Atlas of United States Trees. Volume 3. Minor Western Hardwoods; United States Department of Agriculture Forest Service: Washington, DC, USA, 1976; 13p, 290 maps.
- Little, E.L. Atlas of United States Trees. Volume 4. Minor Eastern Hardwoods; United States Department of Agriculture Forest Service: Washington, DC, USA, 1976; 17p, 230 maps.
- U.S. Department of Agriculture Natural Resource Conservation Service. The PLANTS Database. Available online: http://plants.usda.gov (accessed on 1 March 2019).
- Conover, W.J. Practical Nonparametric Stastics; John Wiley & Sons, Inc.: New York, NY, USA, 1971; 462p. [Google Scholar]
- SAS Institute Inc. The SAS System for Windows, Version 9.4; SAS Institute Inc.: Cary, NC, USA, 2013. [Google Scholar]
- Smith, W.B. Forest inventory and analysis: a national inventory and monitoring program. Environ. Pollut. 2002, 116, S233–S242. [Google Scholar] [CrossRef]
- Tinkham, W.T.; Mahoney, P.R.; Hudak, A.T.; Domke, G.M.; Falkowski, M.J.; Woodall, C.W.; Smith, A.M.S. Applications of the United States Forest Inventory and Analysis dataset: A review and future directions. Can. J. For. Res. 2018, 48, 1251–1268. [Google Scholar] [CrossRef]
- Smith, R.L.; Smith, T.M. Ecology and Field Biology, 6th ed.; Addison Wesley Longman: San Francisco, CA, USA, 2001; 771p. [Google Scholar]
- United States Department of Agriculture Forest Service. Alien Forest Pest Explorer Database. Available online: https://www.nrs.fs.fed.us/tools/afpe (accessed on 25 June 2018).
- White, D.; Kimerling, A.J.; Overton, W.S. Cartographic and geometric components of a global sampling design for environmental monitoring. Cartog. Geogr. Inf. 1992, 19, 5–22. [Google Scholar] [CrossRef]
- Potter, K.M.; Koch, F.H.; Oswalt, C.M.; Iannone, B.V. Data, data everywhere: Detecting spatial patterns in fine-scale ecological information collected across a continent. Landsc. Ecol. 2016, 31, 67–84. [Google Scholar] [CrossRef]
- Getis, A.; Ord, J.K. The analysis of spatial association by use of distance statistics. Geogr. Anal. 1992, 24, 189–206. [Google Scholar] [CrossRef]
- ESRI. ArcMap 10.3.1; Environmental Systems Research Institute Inc.: Redlands, CA, USA, 2015. [Google Scholar]
- Brockerhoff, E.G.; Liebhold, A.M. Ecology of forest insect invasions. Biol. Invasions 2017, 19, 3141–3159. [Google Scholar] [CrossRef]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Ajchler, M.; Lobocka, M.; Oszako, T. Pathogenic oomycetes of Phytophthora genus: A new threat to forests in Europe. Sylwan 2017, 161, 870–880. [Google Scholar]
- Foden, W.B.; Butchart, S.H.M.; Stuart, S.N.; Vie, J.C.; Akcakaya, H.R.; Angulo, A.; DeVantier, L.M.; Gutsche, A.; Turak, E.; Cao, L.; et al. Identifying the world’s most climate change vulnerable species: A systematic trait-based assessment of all birds, amphibians and corals. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Dumroese, R.K.; Williams, M.I.; Stanturf, J.A.; St. Clair, J.B. Considerations for restoring temperate forests of tomorrow: forest restoration, assisted migration, and bioengineering. New For. 2015, 46, 947–964. [Google Scholar] [CrossRef]
- United States Department of Agriculture Forest Health Protection. Insect and Disease Detection Survey Database (IDS). Available online: https://www.fs.fed.us/foresthealth/applied-sciences/mapping-reporting/gis-spatial-analysis/detection-surveys.shtml (accessed on 24 October 2018).
- United States Department of Agriculture Forest Service. Future of America’s Forest and Rangelands: Forest Service 2010 Resources Planning Act Assessment; U.S. Department of Agriculture Forest Service: Washington, DC, USA, 2012; 198p.
- United States Department of Agriculture Forest Service. Future of America’s Forests and Rangelands: Update to the 2010 Resources Planning Act Assessment; U.S. Department of Agriculture Forest Service: Washington, DC, USA, 2016; 250p.
- United States Department of Agriculture Forest Service. National Report on Sustainable Forests—2010; U.S. Department of Agriculture Forest Service: Washington, DC, USA, 2011; 134p.
- United States Department of Agriculture Forest Service. National Report on Sustainable Forests—2003; U.S. Department of Agriculture Forest Service: Washington, DC, USA, 2004; 139p.
- Smith, W.B.; Miles, P.D.; Perry, C.H.; Pugh, S.A. Forest resources of the United States, 2007; GTR-WO-78; U.S. Department of Agriculture Forest Service, Washington Office: Washington, DC, USA, 2009; 336p.
- Hudgins, E.J.; Liebhold, A.M.; Leung, B. Predicting the spread of all invasive forest pests in the United States. Ecol. Lett. 2017, 20, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Meddens, A.J.H.; Hicke, J.A.; Ferguson, C.A. Spatiotemporal patterns of observed bark beetle-caused tree mortality in British Columbia and the western United States. Ecol. Appl. 2012, 22, 1876–1891. [Google Scholar] [CrossRef] [PubMed]
- Royama, T. Analytical Population Dynamics; Chapman and Hall: London, UK, 1992. [Google Scholar]
- Chapman, T.B.; Veblen, T.T.; Schoennagel, T. Spatiotemporal patterns of mountain pine beetle activity in the southern Rocky Mountains. Ecology 2012, 93, 2175–2185. [Google Scholar] [CrossRef]
- Rocky Mountain Region Forest Health Protection. A Field Guide to Insects and Diseases of the Rocky Mountain Region; U.S. Department of Agriculture Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2010; 336p.
- Teale, S.A.; Castello, J.D. Regulators and terminators: The importance of biotic factors to a healthy forest. In Forest Health: An Integrated Perspective; Castello, J.D., Teale, S.A., Eds.; Cambridge University Press: New York, NY, USA, 2011; pp. 81–114. [Google Scholar]
- Edmonds, R.L.; Agee, J.K.; Gara, R.I. Forest Health and Protection, 2nd ed.; Waveland Press, Inc.: Long Grove, IL, USA, 2011; 667p. [Google Scholar]
- Weed, A.S.; Ayres, M.P.; Hicke, J.A. Consequences of climate change for biotic disturbances in North American forests. Ecol. Monogr. 2013, 83, 441–470. [Google Scholar] [CrossRef]
- Kolb, T.E.; Fettig, C.J.; Ayres, M.P.; Bentz, B.J.; Hicke, J.A.; Mathiasen, R.; Stewart, J.E.; Weed, A.S. Observed and anticipated impacts of drought on forest insects and diseases in the United States. For. Ecol. Manag. 2016, 380, 321–334. [Google Scholar] [CrossRef]
- Sniezko, R.A.; Koch, J. Breeding trees resistant to insects and diseases: Putting theory into application. Biol. Invasions 2017, 19, 3377–3400. [Google Scholar] [CrossRef]
- Aukema, J.E.; McCullough, D.G.; Von Holle, B.; Liebhold, A.M.; Britton, K.; Frankel, S.J. Historical accumulation of nonindigenous forest pests in the continental United States. Bioscience 2010, 60, 886–897. [Google Scholar] [CrossRef]
- Koch, F.H.; Yemshanov, D.; Colunga-Garcia, M.; Magarey, R.D.; Smith, W.D. Potential establishment of alien-invasive forest insect species in the United States: where and how many? Biol. Invasions 2011, 13, 969–985. [Google Scholar] [CrossRef]
- Aukema, J.E.; Leung, B.; Kovacs, K.; Chivers, C.; Britton, K.O.; Englin, J.; Frankel, S.J.; Haight, R.G.; Holmes, T.P.; Liebhold, A.M.; et al. Economic impacts of non-native forest insects in the continental United States. PLoS ONE 2011, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Tonini, F.; Jones, C.; Miranda, B.R.; Cobb, R.C.; Sturtevant, B.R.; Meentemeyer, R.K. Modeling epidemiological disturbances in LANDIS-II. Ecography 2018, 41, 2038–2044. [Google Scholar] [CrossRef]
- Scheller, R.M.; Kretchun, A.M.; Loudermilk, E.L.; Hurteau, M.D.; Weisberg, P.J.; Skinner, C. Interactions Among Fuel Management, Species Composition, Bark Beetles, and Climate Change and the Potential Effects on Forests of the Lake Tahoe Basin. Ecosystems 2018, 21, 643–656. [Google Scholar] [CrossRef]
- Rogers, B.M.; Jantz, P.; Goetz, S.J. Vulnerability of eastern US tree species to climate change. Glob. Chang. Biol. 2017, 23, 3302–3320. [Google Scholar] [CrossRef]
- Tuomola, J.; Yemshanov, D.; Huitu, H.; Hannunen, S. Mapping risks of pest invasions based on the spatio-temporal distribution of hosts. Manag. Biol. Invasion 2018, 9, 115–126. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Spp. | All Insect and Disease Agents | Exotic Insect and Disease Agents | ||||
|---|---|---|---|---|---|---|---|
| Hosts | Host/Agent Combs. | Mean Severity, Combs. | Hosts | Host/Agent Combs. | Mean Severity, Combs. | ||
| Acanthaceae | 1 | 1 | 1 | 3.00 | 0 | 0 | 0.00 |
| Agavaceae | 2 | 1 | 1 | 1.00 | 0 | 0 | 0.00 |
| Anacardiace | 4 | 2 | 2 | 2.00 | 0 | 0 | 0.00 |
| Annonaceae | 2 | 1 | 2 | 1.00 | 1 | 1 | 1.00 |
| Aquifoliace | 4 | 2 | 3 | 2.33 | 0 | 0 | 0.00 |
| Arecaceae | 8 | 8 | 18 | 2.22 | 0 | 0 | 0.00 |
| Betulaceae | 16 | 13 | 51 | 2.37 | 12 | 22 | 2.91 |
| Bignoniaceae | 3 | 2 | 8 | 1.50 | 0 | 0 | 0.00 |
| Boraginaceae | 3 | 1 | 2 | 1.00 | 0 | 0 | 0.00 |
| Burseraceae | 1 | 1 | 2 | 1.00 | 1 | 1 | 1.00 |
| Cactaceae | 1 | 1 | 2 | 1.00 | 0 | 0 | 0.00 |
| Cannabaceae | 4 | 4 | 13 | 1.00 | 1 | 1 | 1.00 |
| Combretaceae | 2 | 1 | 3 | 1.00 | 1 | 2 | 1.00 |
| Cornaceae | 7 | 7 | 22 | 1.77 | 3 | 3 | 6.00 |
| Cupressaceae | 33 | 33 | 103 | 1.69 | 17 | 21 | 1.81 |
| Ebenaceae | 2 | 2 | 3 | 3.67 | 0 | 0 | 0.00 |
| Ericaceae | 4 | 4 | 15 | 1.93 | 2 | 2 | 2.00 |
| Euphorbiaceae | 1 | 0 | 0 | 0.00 | 0 | 0 | 0.00 |
| Fabaceae | 22 | 17 | 49 | 1.08 | 2 | 3 | 1.00 |
| Fagaceae | 65 | 65 | 281 | 3.11 | 65 | 124 | 5.13 |
| Hamamelidaceae | 1 | 1 | 5 | 1.00 | 1 | 2 | 1.00 |
| Juglandaceae | 19 | 19 | 56 | 1.55 | 2 | 2 | 5.50 |
| Lauraceae | 3 | 3 | 12 | 2.92 | 3 | 7 | 4.29 |
| Magnoliaceae | 9 | 4 | 11 | 1.00 | 0 | 0 | 0.00 |
| Meliaceae | 1 | 1 | 2 | 1.00 | 0 | 0 | 0.00 |
| Moraceae | 5 | 5 | 15 | 2.20 | 4 | 6 | 3.33 |
| Myrtaceae | 1 | 0 | 0 | 0.00 | 0 | 0 | 0.00 |
| Nyctaginaceae | 1 | 0 | 0 | 0.00 | 0 | 0 | 0.00 |
| Oleaceae | 16 | 15 | 62 | 4.23 | 15 | 29 | 5.97 |
| Pinaceae | 68 | 68 | 307 | 3.12 | 30 | 40 | 4.40 |
| Platanaceae | 3 | 3 | 10 | 2.10 | 1 | 1 | 8.00 |
| Polygonaceae | 1 | 1 | 2 | 1.00 | 0 | 0 | 0.00 |
| Rhamnaceae | 3 | 1 | 2 | 1.00 | 1 | 1 | 1.00 |
| Rhizophoraceae | 1 | 1 | 3 | 2.33 | 1 | 2 | 3.00 |
| Rosaceae | 31 | 30 | 111 | 1.47 | 13 | 23 | 2.30 |
| Rutaceae | 4 | 0 | 0 | 0.00 | 0 | 0 | 0.00 |
| Salicaceae | 22 | 22 | 84 | 1.79 | 18 | 41 | 2.27 |
| Sapindaceae | 20 | 19 | 73 | 2.74 | 19 | 35 | 4.51 |
| Sapotaceae | 4 | 2 | 3 | 1.00 | 2 | 2 | 1.00 |
| Simaroubaceae | 1 | 0 | 0 | 0.00 | 0 | 0 | 0.00 |
| Styracaceae | 3 | 0 | 0 | 0.00 | 0 | 0 | 0.00 |
| Taxaceae | 4 | 3 | 6 | 2.50 | 2 | 3 | 1.00 |
| Theaceae | 2 | 0 | 0 | 0.00 | 0 | 0 | 0.00 |
| Tiliaceae | 3 | 3 | 9 | 1.67 | 3 | 6 | 2.00 |
| Ulmaceae | 7 | 7 | 22 | 4.77 | 7 | 15 | 4.93 |
| Verbenaceae | 1 | 1 | 2 | 1.00 | 1 | 1 | 1.00 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Potter, K.M.; Escanferla, M.E.; Jetton, R.M.; Man, G. Important Insect and Disease Threats to United States Tree Species and Geographic Patterns of Their Potential Impacts. Forests 2019, 10, 304. https://doi.org/10.3390/f10040304
Potter KM, Escanferla ME, Jetton RM, Man G. Important Insect and Disease Threats to United States Tree Species and Geographic Patterns of Their Potential Impacts. Forests. 2019; 10(4):304. https://doi.org/10.3390/f10040304
Chicago/Turabian StylePotter, Kevin M., Maria E. Escanferla, Robert M. Jetton, and Gary Man. 2019. "Important Insect and Disease Threats to United States Tree Species and Geographic Patterns of Their Potential Impacts" Forests 10, no. 4: 304. https://doi.org/10.3390/f10040304
APA StylePotter, K. M., Escanferla, M. E., Jetton, R. M., & Man, G. (2019). Important Insect and Disease Threats to United States Tree Species and Geographic Patterns of Their Potential Impacts. Forests, 10(4), 304. https://doi.org/10.3390/f10040304