Factors Affecting the Distribution of Pine Pitch Canker in Northern Spain

 , ,
, ,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description and Sampling Procedure
2.2. Statistical Analysis
2.2.1. Spatial Autocorrelation Analyses
2.2.2. Univariate Analyses
2.2.3. Multivariate
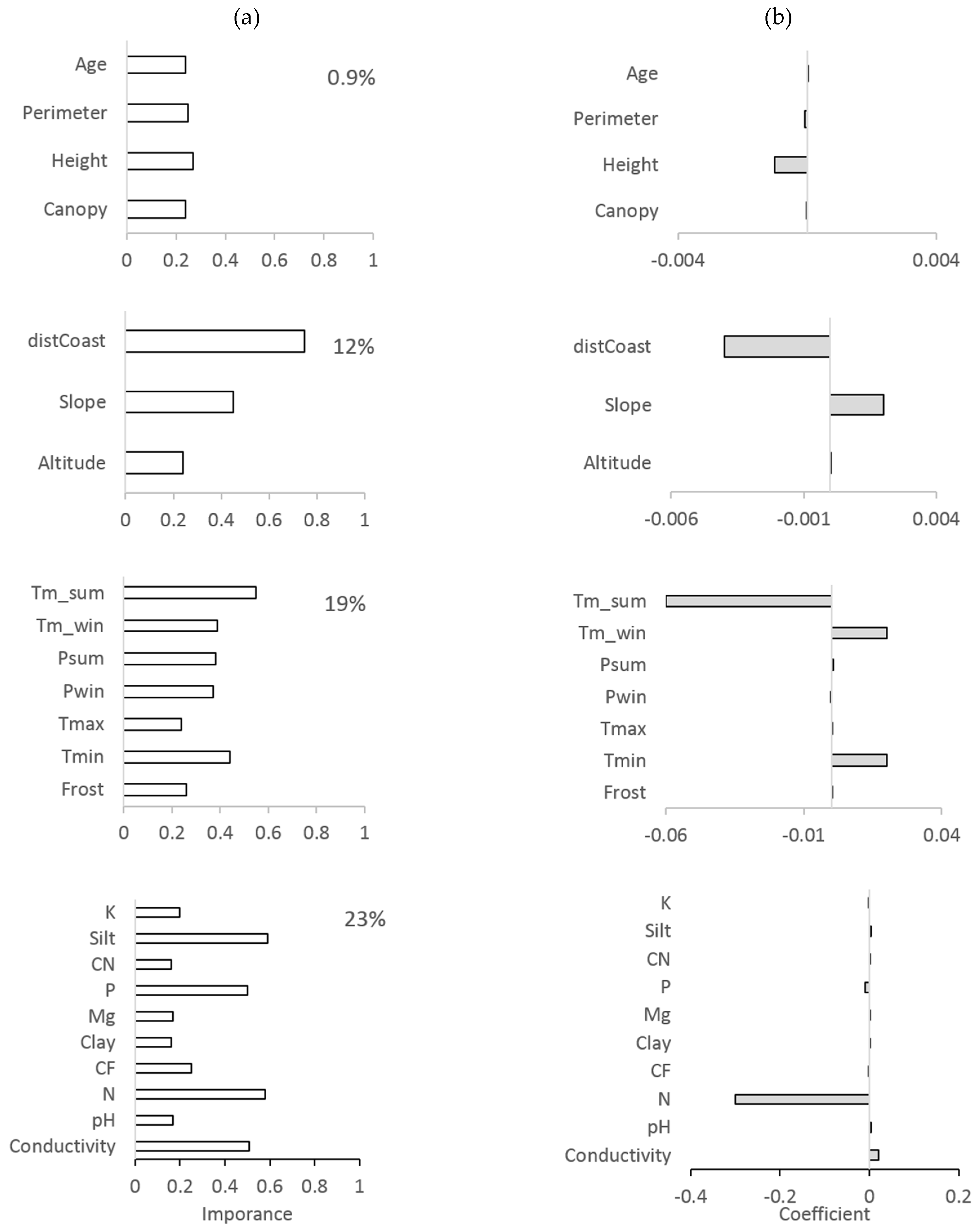
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PPC | Pine Pitch Canker |
| Asl | above sea level |
References
- Nirenberg, H.I.; O’Donnell, K. New Fusarium species and combinations within the Gibberella fujikuroi species complex. Mycologia 1998, 434–458. [Google Scholar] [CrossRef]
- Vettraino, A.; Potting, R.; Raposo, R. EU legislation on forest plant health: An overview with a focus on Fusarium circinatum. Forests 2018, 9, 568. [Google Scholar] [CrossRef]
- Wingfield, M.J.; Hammerbacher, A.; Ganley, R.J.; Steenkamp, E.T.; Gordon, T.R.; Wingfield, B.D.; Coutinho, T.A. Pitch canker caused by Fusarium circinatum—A growing threat to pine plantations and forests worldwide. Australas. Plant Pathol. 2008, 37, 319–334. [Google Scholar] [CrossRef]
- Hepting, G.H.; Roth, E.R. Pitch canker, a new disease of some southern pines. J. For. 1946, 44, 742–744. [Google Scholar]
- McCain, A.H.; Koehler, C.S.; Tjosvold, S.A. Pitch canker threatens California pines. Calif. Agric. 1987, 41, 22–23. [Google Scholar]
- Hepting, G.H.; Roth, E.R. Host relations and spread of the pine pitch canker disease. Phytopathology 1953, 43, 475. [Google Scholar]
- Wingfield, M.J.; Jacobs, A.; Coutinho, T.A.; Ahumada, R.; Wingfield, B.D. First report of the pitch canker fungus, Fusarium circinatum, on pines in Chile. Plant Pathol. 2002, 51, 397. [Google Scholar] [CrossRef]
- Viljoen, A.; Wingfield, M.J.; Kemp, G.H.J.; Marasas, W.O. Susceptibility of pines in South Africa to the pitch canker fungus Fusarium subglutinans f. sp. Pini. Plant Pathol. 1995, 44, 877–882. [Google Scholar] [CrossRef]
- Muramoto, M.; Dwinell, L.D. Pitch canker of Pinus luchuensis in Japan. Plant Dis. 1990, 74, 530. [Google Scholar] [CrossRef]
- Guerra-Santos, J.J. Pitch canker on Monterey pine in Mexico. In Forestry and Forest Products. Current and Potential Impacts of Pitch Canker in Radiata Pine, Proceedings of the IMPACT Monterey Workshop, Monterey, CA, USA, 30 November–3 December 1998; Technical Report; Devey, M., Matheson, C., Gordon, T., Eds.; CSIRO: Canberra, Australia, 1999; Volume 112, pp. 58–61. [Google Scholar]
- Cho, W.D.; Shin, H.D. List of Plant Diseases in Korea, 4th ed.; Korean Society of Plant Pathology: Seoul, Korea, 2004. [Google Scholar]
- Alonso, R.; Bettucci, L. First report of the pitch canker fungus Fusarium circinatum affecting Pinus taeda seedlings in Uruguay. Australas. Plant Dis. Notes 2009, 4, 91–92. [Google Scholar]
- Steenkamp, E.T.; Rodas, C.A.; Kvas, M.; Wingfield, M.J. Fusarium circinatum and pitch canker of Pinus in Colombia. Australas. Plant Pathol. 2012, 41, 483–491. [Google Scholar] [CrossRef]
- Pfenning, L.H.; Costa, S.D.S.; de Melo, M.P.; Costa, H.; Ventura, J.A.; Auer, C.G.; Santos, Á.F.D. First report and characterization of Fusarium circinatum, the causal agent of pitch canker in Brazil. Trop. Plant Pathol. 2014, 39, 210–216. [Google Scholar] [CrossRef]
- Landeras, E.; García, P.; Fernández, Y.; Braña, M. Outbreak of pitch canker caused by Fusarium circinatum on Pinus spp. in Northern Spain. Plant Disease. 2005, 89, 1015. [Google Scholar] [CrossRef]
- EPPO (European and Mediterranean Plant Protection Organization). First report of Gibberella circinata in France. EPPO Reporting Service 2006-05 no. 104. Paris, France. 2006. Available online: http://www.eppo. int/PUBLICATIONS/reporting/reporting_service.htm (accessed on 5 February 2019).
- Carlucci, A.; Colatruglio, L.; Frisullo, S. First report of pitch canker caused by Fusarium circinatum on Pinus halepensis and Pinus pinea in Apulia (Southern Italy). Plant Dis. 2007, 91, 1683. [Google Scholar] [CrossRef]
- Bragança, H.; Diogo, E.; Moniz, F.; Amaro, P. First report of pitch canker on pines caused by Fusarium circinatum in Portugal. Plant Dis. 2009, 93, 1079. [Google Scholar] [CrossRef] [PubMed]
- Dwinell, L.D.; Adams, D.; Guerra-Santos, J.J.; Aquirre, J.R.M. Pitch canker disease of Pinus radiata. In Proceedings of the 7th International Congress of Plant Pathology, Edinburgh, UK, 9–16 August 1998; pp. 9–16. [Google Scholar]
- Martínez-Álvarez, P.; Pando, V.; Diez, J.J. Alternative species to replace Monterey pine plantations affected by pitch canker caused by Fusarium circinatum in northern Spain. Plant Pathol. 2014, 63, 1086–1094. [Google Scholar] [CrossRef]
- CAB International. Gibberella Circinata (Pitch Canker); Crop Protection Compendium: Wallingford, UK, 2007. [Google Scholar]
- Gordon, T.R.; Storer, A.J.; Wood, D.L. The pitch canker epidemic in California. Plant Dis. 2001, 85, 1128–1139. [Google Scholar] [CrossRef] [PubMed]
- Hermoso, E.; Carballo, J.; Fernandez-Golfin, J. Structural characterization of Pinus radiata D. Don Timber from Pais Vasco (Spain) according to Standard Modifications. Maderas-Cienc. Tecnol. 2007, 9, 223–232. [Google Scholar]
- Martín-Rodrigues, N.; Espinel, S.; Sanchez-Zabala, J.; Ortíz, A.; González-Murua, C.; Duñabeitia, M.K. Spatial and temporal dynamics of the colonization of Pinus radiata by Fusarium circinatum, of conidiophora development in the pith and of traumatic resin duct formation. New Phytol. 2013, 198, 1215–1227. [Google Scholar] [CrossRef]
- Bezos, D.; Martínez-Álvarez, P.; Diez, J.J.; Fernández, M.M. The pine shoot beetle Tomicus piniperda as a plausible vector of Fusarium circinatum in northern Spain. Ann. For. Sci. 2015, 72, 1079–1088. [Google Scholar] [CrossRef]
- Romón, P.; Iturrondobeitia, J.C.; Gibson, K.; Lindgren, B.S.; Goldarazena, A. Quantitative association of bark beetles with pitch canker fungus and effects of verbenone on their semiochemical communication in Monterey pine forests in northern Spain. Environ. Entomol. 2007, 36, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Gordon, T.R. Pitch Canker. In Infectious Forest Diseases; CABI Publishing: Oxon, UK, 2013; pp. 376–391. [Google Scholar]
- Dwinell, L.D.; Barrows-Broaddus, J.B.; Kuhlman, E.G. Pitch canker: A disease complex. Plant Dis. 1985, 69, 270–276. [Google Scholar] [CrossRef]
- Gordon, T.R.; Reynolds, G.J. Plasticity in plant-microbe interactions: A perspective based on the pitch canker pathosystem. Phytoparasitica 2017, 45, 1–8. [Google Scholar] [CrossRef]
- Kelley, W.D.; Williams, J.C. Incidence of pitch canker among clones of loblolly pine in seed orchards. Plant Dis. 1982, 66, 1171–1173. [Google Scholar] [CrossRef]
- Bezos, D.; Lomba, J.M.; Martinez-Alvarez, P.; Fernández, M.; Diez, J.J. Effects of Pruning in Monterrey Pine Plantations Affected by Fusarium circinatum. For. Syst. 2012, 21, 481–488. [Google Scholar]
- Eschen, R.; Rigaux, L.; Sukovata, L.; Vettraino, A.M.; Marzano, M.; Grégoire, J.-C. Phytosanitary inspection of woody plants for planting at European Union entry points: A practical enquiry. Biol. Invasions 2015, 17, 2403–2413. [Google Scholar] [CrossRef]
- Martín-García, J. Environmentally-friendly methods for controlling pine pitch canker. Plant Pathol. 2019. [Google Scholar] [CrossRef]
- Ganley, R.J.; Watt, M.S.; Manning, L.; Iturritxa, E. A global climatic risk assessment of pitch canker disease. Can. J. For. Res. 2009, 39, 2246–2256. [Google Scholar] [CrossRef]
- Möykkynen, T.; Capretti, P.; Pukkala, T. Modelling the potential spread of Fusarium circinatum, the causal agent of pitch canker in Europe. Ann. For. Sci. 2015, 72, 169–181. [Google Scholar] [CrossRef]
- Vivas, M.; Zas, R.; Sampedro, L.; Solla, A. Environmental maternal effects mediate the resistance of Maritime pine to biotic stress. PLoS ONE 2013, 8, e70148. [Google Scholar] [CrossRef]
- Fraedrich, B.R.; Witcher, W. Influence of fertilizations on pitch canker development on three southern pine species. Plant Dis. 1982, 66, 938–940. [Google Scholar] [CrossRef]
- Wikler, K.; Storer, A.J.; Newman, W.; Gordon, T.R.; Wood, D.L. The dynamics of an introduced pathogen in a native Monterey pine (Pinus radiata) forest. For. Ecol. Manag. 2003, 179, 209–221. [Google Scholar] [CrossRef]
- Gordon, T.R.; Swett, C.L.; Wingfield, M.J. Management of Fusarium diseases affecting conifers. Crop Prot. 2015, 73, 28–39. [Google Scholar] [CrossRef]
- ICP-Forests. Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; Thünen Institute of Forest Ecosystems: Eberswalde, Germany, 2017. [Google Scholar]
- Bascomb, C.L. Rapid method for the determination of cation-exchange capacity of calcareous and non-calcareous soils. J. Sci. Food Agric. 1964, 15, 821–823. [Google Scholar] [CrossRef]
- Ninyerola, M.; Pons, X.; Roure, J.M. Altas climático digital de la Península Ibérica. In Metodología y Aplicaciones en Bioclimatología y Geobotánica; Universidad Autónoma de Barcelona: Bellaterra, Spain, 2005. [Google Scholar]
- Peres-Neto, P.R.; Legendre, P. Estimating and controlling for spatial structure in the study of ecological communities. Glob. Ecol. Biogeogr. 2010, 19, 174–184. [Google Scholar] [CrossRef]
- Goodchild, M.F. Spatial Autocorrelation; Geo Books: Norwich, CT, USA, 1986; Volume 47. [Google Scholar]
- Neter, J.; Wasserman, W.; Kutner, M.H. Applied Linear Regression Models; Richard D. Irwin: Boston, MA, USA, 1989. [Google Scholar]
- Naimi, B.; Hamm, N.A.; Groen, T.A.; Skidmore, A.K.; Toxopeus, A.G. Where is positional uncertainty a problem for species distribution modelling? Ecography 2014, 37, 191–203. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Blank, L.; Linker, R.; Carmel, Y. A multiscale analysis of herbaceous species richness in a Mediterranean ecosystem. J. Plant Ecol. 2013, 6, 113–121. [Google Scholar] [CrossRef]
- Krasnov, H.; Cohen, Y.; Goldshtein, E.; Mendelsohn, O.; Silberstein, M.; Gazit, Y.; Blank, L. The effect of local and landscape variables on Mediterranean fruit fly dynamics in citrus orchards utilizing the ecoinformatics approach. J. Pest Sci. 2018. [Google Scholar] [CrossRef]
- Calcagno, V.; de Mazancourt, C. glmulti: An R package for easy automated model selection with (generalized) linear models. J. Stat. Softw. 2010, 34, 1–29. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N. A protocol for conducting and presenting results of regression-type analyses. Methods Ecol. Evol. 2016, 7, 636–645. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S.; Adler, D.; Bates, D.; Baud-Bovy, G.; Ellison, S.; Firth, D.; Friendly, M.; Gorjanc, G.; Graves, S. Package ‘Car’; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Bjørnstad, O.N. Package ‘ncf’: Spatial Nonparametric Covariance Functions; R Core Development Team: Vienna, Austria, 2006. [Google Scholar]
- Blank, L.; Blaustein, L. A multi-scale analysis of breeding site characteristics of the endangered fire salamander (Salamandra infraimmaculata) at its extreme southern range limit. Hydrobiologia 2014, 726, 229–244. [Google Scholar] [CrossRef]
- Blank, L.; Cohen, Y.; Borenstein, M.; Shulhani, R.; Lofthouse, M.; Sofer, M.; Shtienberg, D. Variables associated with severity of bacterial canker and wilt caused by Clavibacter michiganensis subsp. michiganensis in tomato greenhouses. Phytopathology 2016, 106, 254–261. [Google Scholar] [CrossRef] [PubMed]
- MacNally, R.; Walsh, C.J. Hierarchical partitioning public-domain software. Biodivers. Conserv. 2004, 13, 659–660. [Google Scholar]
- Nagelkerke, N.J. A note on a general definition of the coefficient of determination. Biometrika 1991, 78, 691–692. [Google Scholar] [CrossRef]
- Nakazawa, M. fmsb: Functions for Medical Statistics Book with Some Demographic Data; R Package Version 0.4; R Core Development Team: Vienna, Austria, 2014; p. 5. [Google Scholar]
- R Core Development Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://cran. r-project.org (accessed on 5 February 2019).
- Bulman, L.S.; Bradshaw, R.E.; Fraser, S.; Martín-García, J.; Barnes, I.; Musolin, D.L.; La Porta, N.; Woods, A.J.; Diez, J.J.; Koltay, A. A worldwide perspective on the management and control of Dothistroma needle blight. For. Pathol. 2016, 46, 472–488. [Google Scholar] [CrossRef]
- Pietrzykowski, M. Macronutrient accumulation and relationships in a Scots pine (Pinus sylvestris L.) ecosystem on reclaimed opencast lignite mine spoil heaps in central Poland. In Proceedings of the 25th Annual Meeting of ASMR and 10th IALR, “New Opportunities to Apply Our Science”, Richmond, VA, USA, 14–19 June 2008; pp. 856–877. [Google Scholar]
- Inman, A.R.; Kirkpatrick, S.C.; Gordon, T.R.; Shaw, D.V. Limiting effects of low temperature on growth and spore germination in Gibberella circinata, the cause of pitch canker in pine species. Plant Dis. 2008, 92, 542–545. [Google Scholar] [CrossRef]
- Sakamoto, J.M.; Gordon, T.R. Factors influencing infection of mechanical wounds by Fusarium circinatum on Monterey pines (Pinus radiata). Plant Pathol. 2006, 55, 130–136. [Google Scholar] [CrossRef]
- Blakeslee, G.M.; Jokela, E.J.; Hollis, C.H.; Wilson, D.S.; Lante, W.D.; Allen, J.E. Pitch canker in young loblolly pines: Influence of precommercial thinning and fertilization on disease incidence and severity. South. J. Appl. For. 1999, 23, 139–143. [Google Scholar]
- Laucirica, J.M.; Muguruza, J.R. Presencia de Fusarium subglutinans F. sp. pini en viveros de pino radiata en Bizkaia. In Proceedings of the XIV Reunión anual del Grupo de Trabajo Fitosanitario de Forestales, Parques y Jardines, Marbella, Spain, 19–21 November 1997; pp. 18–20. [Google Scholar]
- Bezos, D.; Martínez-Álvarez, P.; Sanz-Ros, A.; Martín-García, J.; Fernandez, M.; Diez, J. Fungal Communities Associated with Bark Beetles in Pinus radiata Plantations in Northern Spain Affected by Pine Pitch Canker, with Special Focus on Fusarium Species. Forests 2018, 9, 698. [Google Scholar] [CrossRef]
- Datnoff, L.E.; Elmer, W.H.; Huber, D.M. Mineral Nutrition and Plant Disease; American Phytopathological Society (APS Press): St Paul, MN, USA, 2007. [Google Scholar]
- Fisher, R.F.; Garbett, W.S.; Underhill, E.M. Effects of fertilization on healthy and pitch canker-infected pines. South. J. Appl. For. 1981, 5, 77–79. [Google Scholar]
- Kozlowski, T.T. Growth and development of trees. In Seed Germination, Ontogeny and Shoot Growth; Academic Press: New York, NY, USA, 1971; Volume I. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Abbreviation | Units | Average | Min | Max | |
|---|---|---|---|---|---|---|
| Pathology | Cankers | Canker | proportion | 0.20 | 0.00 | 0.64 |
| Defoliation | Defoliation | proportion | 0.14 | 0.00 | 0.66 | |
| Dieback | Dieback | proportion | 0.16 | 0.00 | 0.48 | |
| Climate | Average annual precipitation | Precipitation | mm | 1232 | 702 | 1735 |
| Average annual maximal temperature | Tmax | °C | 18.3 | 17.0 | 21.0 | |
| Average annual minimal temperature | Tmin | °C | 7.7 | 6.0 | 9.0 | |
| Average summer temperature | Tm_sum | °C | 18.6 | 17.4 | 19.8 | |
| Average winter temperature | Tm_win | °C | 8.66 | 7.2 | 10.2 | |
| Average summer precipitation | Psum | mm | 202.1 | 82.4 | 251.6 | |
| Average winter precipitation | Pwin | mm | 355 | 165.8 | 453.3 | |
| Frost period | Frost | Number of months | 5.3 | 3.0 | 7.0 | |
| Topography | Slope | Slope | Degree | 15.8 | 5.0 | 35.0 |
| Elevation | Elevation | Meters asl | 360.5 | 92.0 | 898.0 | |
| Distance from the eastern boarder of Cantabria | distEast | Km | 19.3 | 2.9 | 39.5 | |
| Distance from the coast | distCoast | Km | 19 | 2.0 | 39.0 | |
| Soil | pH | pH | No units | 4.6 | 3.8 | 6.6 |
| Cationic exchange capacity | Conductivity | MS/cm | 0.08 | 0.03 | 0.6 | |
| Coarse fragments | CF | G/100 gr | 5.3 | 0.0 | 58.0 | |
| Percentage of sand | Sand | % | 56.7 | 12.6 | 80.0 | |
| Percentage of silt | Silt | % | 17.9 | 3.3 | 44.6 | |
| Percentage of clay | Clay | % | 24.2 | 1.4 | 49.7 | |
| Organic matter | OM | G/100 gr | 3.5 | 0.7 | 9.4 | |
| Potassium | K | G/100 gr | 61.94 | 13.00 | 365.00 | |
| Phosphorus | P | Mg/kg | 1.2 | 0.0 | 5.5 | |
| Calcium | Ca | Meq/100 gr | 1.5 | 0.03 | 29.1 | |
| Magnesium | Mg | Meq/100 gr | 0.25 | 0.03 | 0.84 | |
| C/N ratio | CN | No units | 11.9 | 6.2 | 16.7 | |
| Nitrogen | N | G/100 gr | 0.2 | 0.05 | 0.44 | |
| Stand characteristics | Canopy cover | Canopy | % | 44.3 | 17.7 | 81.4 |
| Mean diameter | DBH | cm | 25.1 | 13.8 | 51.5 | |
| Stand age | Age | years | 22.6 | 5.0 | 56.0 | |
| Average height | Height | m | 16.6 | 10.5 | 27.3 | |
| Average perimeter | Perimeter | cm | 80.6 | 47.7 | 169.3 |
| Independante Variable | Canker | Defoliation | Dieback |
|---|---|---|---|
| Tm_sum | −0.19 | 0.15 | 0.03 |
| Tm_win | 0.13 | 0.09 | 0.03 |
| Psum | 0.08 | −0.28 | −0.32 |
| Pwin | −0.14 | −0.3 | −0.35 |
| Precipitation * | −0.05 | −0.27 | −0.36 |
| Tmax | −0.08 | 0.26 | 0.25 |
| Tmin | 0.19 | −0.03 | 0.13 |
| Frost | −0.11 | 0.09 | 0.02 |
| Slope | 0.11 | −0.1 | 0.08 |
| Elevation | −0.1 | −0.12 | −0.01 |
| DistEast * | −0.35 | 0.09 | −0.12 |
| DistCoast | −0.29 | 0.07 | −0.12 |
| pH | 0.09 | −0.02 | 0.08 |
| Conductivity | −0.07 | 0.02 | 0.17 |
| CF | 0 | −0.14 | 0.18 |
| Sand * | −0.24 | −0.06 | −0.1 |
| Silt | 0.11 | −0.06 | 0.05 |
| Clay | 0.12 | 0.14 | 0.09 |
| OM * | −0.16 | −0.01 | 0.13 |
| P | −0.2 | 0.1 | 0.1 |
| K | 0.06 | −0.09 | 0.02 |
| Ca * | −0.17 | −0.17 | 0.04 |
| Mg | −0.07 | −0.24 | 0.05 |
| N | −0.19 | −0.07 | 0.13 |
| CN | 0.03 | 0.16 | 0.09 |
| Canopy | 0.03 | −0.06 | −0.28 |
| DBH * | 0.08 | −0.07 | 0.06 |
| Age | 0 | 0.09 | 0.25 |
| Perimeter | 0.05 | −0.15 | −0.05 |
| Height | −0.01 | −0.16 | −0.13 |
| Canker | |||
| Defoliation | 0.04 | ||
| Dieback | 0.13 | 0.5 |
| Group of Variables | Canker | Defoliation | Dieback |
|---|---|---|---|
| Climatic | |||
| Pwin | - | 39 | 45 |
| Psum | - | - | 31 |
| Topography and spatial | |||
| distCoast | 73 | - | - |
| Soil | |||
| - | - | - | - |
| Stand characteristics | |||
| Canopy | - | - | 49 |
| Age | - | - | 35 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blank, L.; Martín-García, J.; Bezos, D.; Vettraino, A.M.; Krasnov, H.; Lomba, J.M.; Fernández, M.; Diez, J.J. Factors Affecting the Distribution of Pine Pitch Canker in Northern Spain. Forests 2019, 10, 305. https://doi.org/10.3390/f10040305
Blank L, Martín-García J, Bezos D, Vettraino AM, Krasnov H, Lomba JM, Fernández M, Diez JJ. Factors Affecting the Distribution of Pine Pitch Canker in Northern Spain. Forests. 2019; 10(4):305. https://doi.org/10.3390/f10040305
Chicago/Turabian StyleBlank, Lior, Jorge Martín-García, Diana Bezos, Anna Maria Vettraino, Helena Krasnov, José M. Lomba, Mercedes Fernández, and Julio J. Diez. 2019. "Factors Affecting the Distribution of Pine Pitch Canker in Northern Spain" Forests 10, no. 4: 305. https://doi.org/10.3390/f10040305
APA StyleBlank, L., Martín-García, J., Bezos, D., Vettraino, A. M., Krasnov, H., Lomba, J. M., Fernández, M., & Diez, J. J. (2019). Factors Affecting the Distribution of Pine Pitch Canker in Northern Spain. Forests, 10(4), 305. https://doi.org/10.3390/f10040305