Nitrogen and Phosphorus Resorption in Planted Forests Worldwide

 ,
, 

Abstract
1. Introduction
2. Materials and Methods
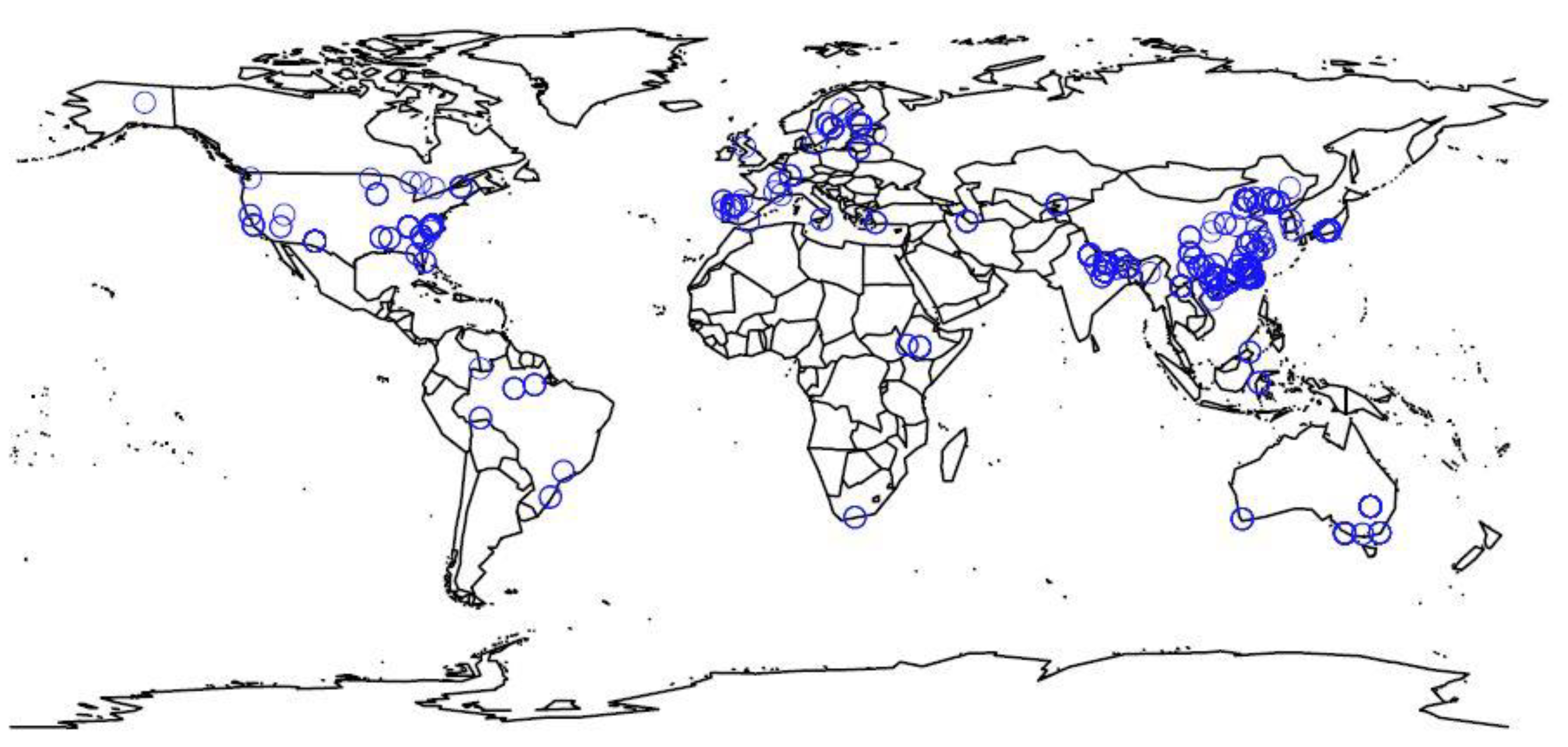
2.1. Data Description
2.2. Statistical Analysis
3. Results
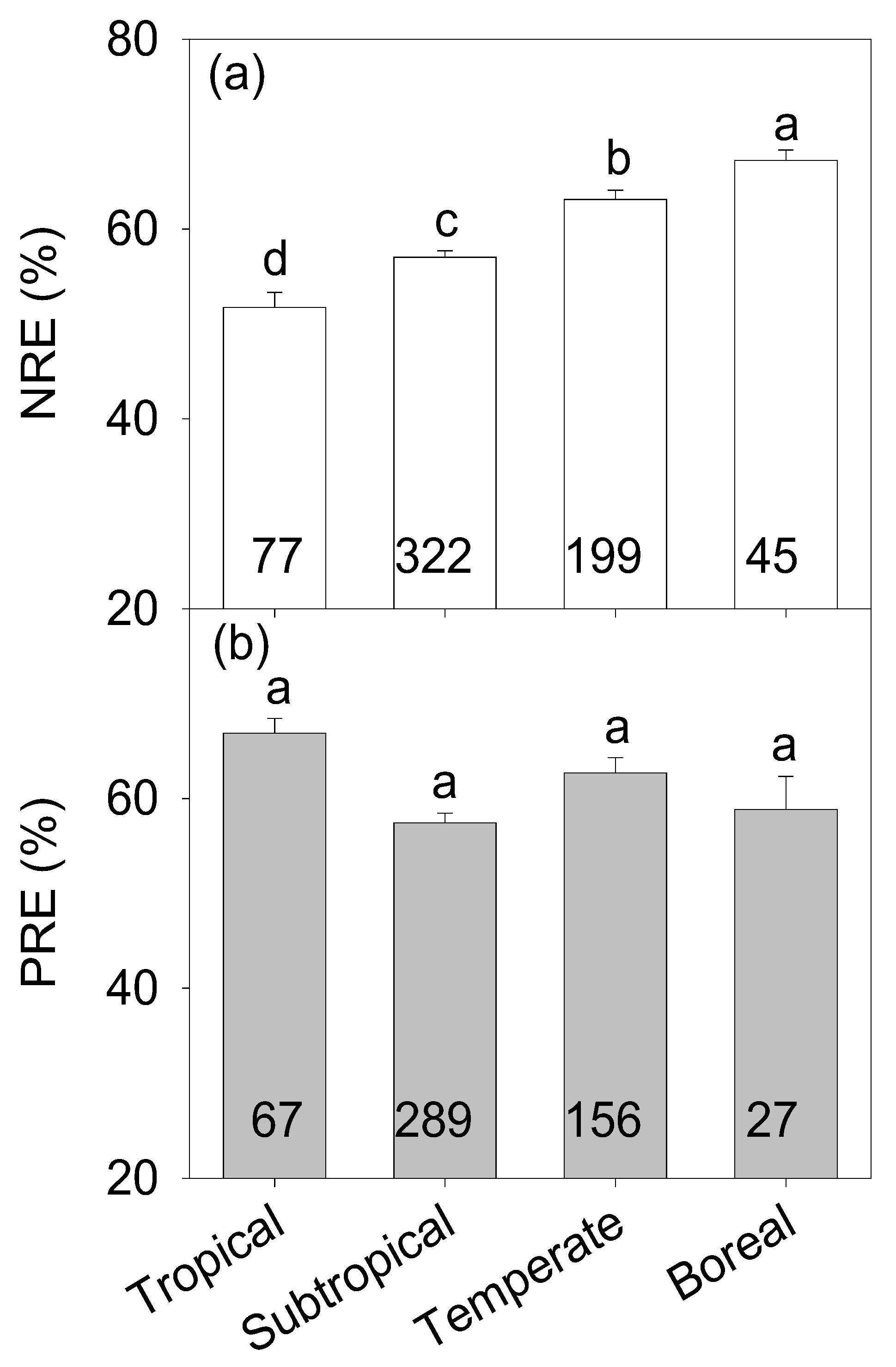
3.1. Nutrient Resorption Patterns in Planted Forests
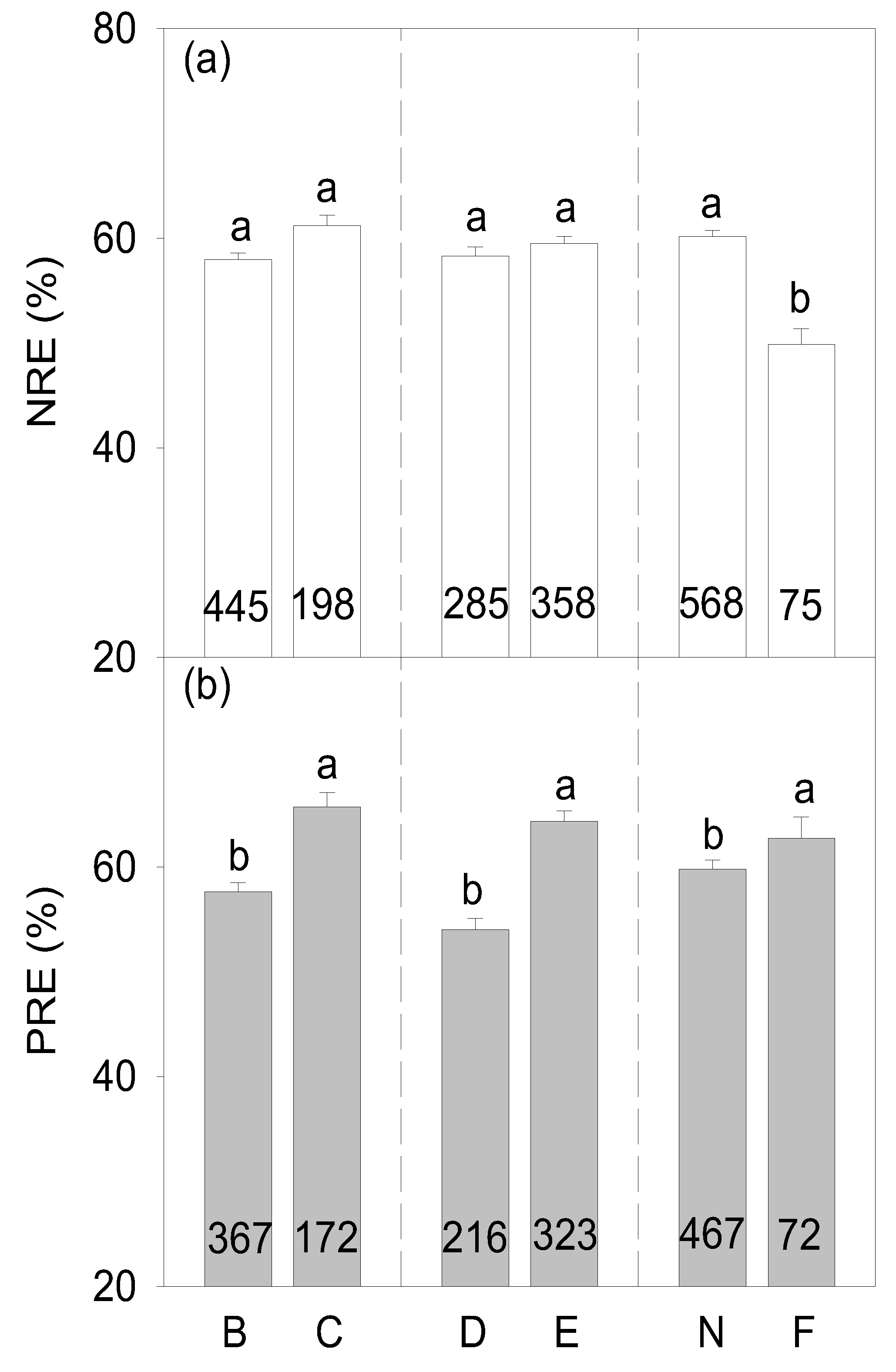
3.2. Controls of Nutrient Resorption in Planted Forests
4. Discussion
4.1. Nutrient Resorption Patterns in Planted Forests
4.2. Controls of Nutrient Resorption in Planted Forests
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vergutz, L.; Manzoni, S.; Porporato, A.; Novais, R.F.; Jackson, R.B. Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecol. Monogr. 2012, 82, 205–220. [Google Scholar] [CrossRef]
- Sokolov, A.P.; Kicklighter, D.W.; Melillo, J.M.; Felzer, B.S.; Schlosser, C.A.; Cronin, T.W. Consequences of considering carbon-nitrogen interactions on the feedbacks between climate and the terrestrial carbon cycle. J. Clim. 2008, 21, 3776–3796. [Google Scholar] [CrossRef]
- Aerts, R. Nutrient Resorption from Senescing Leaves of Perennials: Are there General Patterns? J. Ecol. 1996, 84, 597–608. [Google Scholar] [CrossRef]
- Yan, T.; Lu, X.T.; Zhu, J.J.; Yang, K.; Yu, L.Z.; Gao, T. Changes in nitrogen and phosphorus cycling suggest a transition to phosphorus limitation with the stand development of larch plantations. Plant Soil 2018, 422, 385–396. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H. Global-scale patterns of nutrient resorption associated with latitude, temperature and precipitation. Glob. Ecol. Biogeogr. 2009, 18, 11–18. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Fan, Z.X.; Zhao, Q.; Wang, M.C.; Ran, J.Z.; Huang, H.; Niklas, K.J. Global data analysis shows that soil nutrient levels dominate foliar nutrient resorption efficiency in herbaceous species. Front. Plant Sci. 2018, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Killingbeck, K.T. Nutrients in senesced leaves: Keys to the search for potential resorption and resorption proficiency. Ecology 1996, 77, 1716–1727. [Google Scholar] [CrossRef]
- Schmidt, M.; Veldkamp, E.; Corre, M.D. Tree-microbial biomass competition for nutrients in a temperate deciduous forest, central Germany. Plant Soil 2016, 408, 227–242. [Google Scholar] [CrossRef]
- Achat, D.L.; Pousse, N.; Nicolas, M.; Augusto, L. Nutrient remobilization in tree foliage as affected by soil nutrients and leaf life span. Ecol. Monogr. 2018, 88, 408–428. [Google Scholar] [CrossRef]
- Aerts, R.; Chapin, F.S. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. Adv. Ecol. Res. 2000, 30, 1–67. [Google Scholar] [CrossRef]
- Deng, M.F.; Liu, L.L.; Jiang, L.; Liu, W.X.; Wang, X.; Li, S.P.; Yang, S.; Wang, B. Ecosystem scale trade-off in nitrogen acquisition pathways. Nat. Ecol. Evol. 2018, 2, 1724–1734. [Google Scholar] [CrossRef] [PubMed]
- Brant, A.N.; Chen, H.Y.H. Patterns and mechanisms of nutrient resorption in plants. Crit. Rev. Plant Sci. 2015, 34, 471–486. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H. Negative effects of fertilization on plant nutrient resorption. Ecology 2015, 96, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.Z.; Liu, L.L.; Peng, S.S.; Penuelas, J.; Zeng, H.; Piao, S.L. Age-related modulation of the nitrogen resorption efficiency response to growth requirements and soil nitrogen availability in a temperate Pine plantation. Ecosystems 2016, 19, 698–709. [Google Scholar] [CrossRef]
- Sedjo, R.A. The potential of High-Yield Plantation Forestry for Meeting Timber Needs. In Planted Forests: Contributions to the Quest for Sustainable Societies; Boyle, J.R., Winjum, J.K., Kavanagh, K., Jensen, E.C., Eds.; Springer: Dordrecht, The Netherlands, 1999; pp. 339–359. [Google Scholar]
- Yan, T.; Lu, X.T.; Yang, K.; Zhu, J.J. Leaf nutrient dynamics and nutrient resorption: A comparison between larch plantations and adjacent secondary forests in Northeast China. J. Plant Ecol. 2016, 9, 165–173. [Google Scholar] [CrossRef]
- Sun, X.; Kang, H.; Chen, H.Y.H.; Bjorn, B.; Samuel, B.F.; Liu, C. Biogeographic patterns of nutrient resorption from Quercus variabilis Blume leaves across China. Plant Biol. 2016, 18, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.Y.; Han, W.X.; Chen, Y.H.; Fang, J.Y. Resorption proficiency and efficiency of leaf nutrients in woody plants in eastern China. J. Plant Ecol. 2013, 6, 408–417. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Lu, X.T.; Hartmann, H.; Keller, A.; Han, X.G.; Trumbore, S.; Phillips, R.P. Foliar nutrient resorption differs between arbuscular mycorrhizal and ectomycorrhizal trees at local and global scales. Glob. Ecol. Biogeogr. 2018, 27, 875–885. [Google Scholar] [CrossRef]
- Yan, T.; Zhu, J.J.; Yang, K. Leaf nitrogen and phosphorus resorption of woody species in response to climatic conditions and soil nutrients: A meta-analysis. J. For. Res. 2018, 29, 905–913. [Google Scholar] [CrossRef]
- Han, W.X.; Tang, L.Y.; Chen, Y.H.; Fang, J.Y. Relationship between the relative limitation and resorption efficiency of nitrogen vs phosphorus in woody plants. PLoS ONE 2013, 8, e83366. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, J.N.; Wang, J.Y.; Guo, Z.W.; Wang, G.G.; Zeng, D.H.; Wu, T.G. Tree stoichiometry and nutrient resorption along a chronosequence of Metasequoia glyptostroboides forests in coastal China. For. Ecol. Manag. 2018, 430, 445–450. [Google Scholar] [CrossRef]
- Maeda, Y.; Tashiro, N.; Enoki, T.; Urakawa, R.; Hishi, T. Effects of species replacement on the relationship between net primary production and soil nitrogen availability along a topographical gradient: Comparison of belowground allocation and nitrogen use efficiency between natural forests and plantations. For. Ecol. Manag. 2018, 422, 214–222. [Google Scholar] [CrossRef]
- Xu, S.; Zhou, G.; Tang, X.; Wang, W.; Wang, G.; Ma, K.; Han, S.; Du, S.; Li, S.; Yan, J.; et al. Different spatial patterns of nitrogen and phosphorus resorption efficiencies in China’s forests. Sci. Rep. 2017, 7, 10584. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.Y.; Li, L.H. Soil water status influences plant nitrogen use: A case study. Plant Soil 2007, 301, 303–313. [Google Scholar] [CrossRef]
- Austin, A.T.; Yahdjian, L.; Stark, J.M.; Belnap, J.; Porporato, A.; Norton, U.; Ravetta, D.A.; Schaeffer, S.M. Water pulses and biogeochemical cycles in arid and semiarid ecosystems. Oecologia 2004, 141, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Guo, W.H.; Yu, M.K.; Wang, G.G.; Wu, T.G. Latitudinal patterns of leaf N, P stoichiometry and nutrient resorption of Metasequoia glyptostroboides along the eastern coastline of China. Sci. Total Environ. 2018, 618, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.L.; Addo-Danso, S.D.; Wu, P.F.; Li, S.B.; Zou, X.H.; Zhang, Y.; Ma, X.Q. Leaf resorption efficiency in relation to foliar and soil nutrient concentrations and stoichiometry of Cunninghamia lanceolata with stand development in southern China. J. Soils Sediments 2016, 16, 1448–1459. [Google Scholar] [CrossRef]
- Machado, M.R.; Sampaio, P.D.; Ferraz, J.; Camara, R.; Pereira, M.G. Nutrient retranslocation in forest species in the Brazilian Amazon. Acta Sci.-Agron. 2016, 38, 93–101. [Google Scholar] [CrossRef]
- Ye, G.F.; Zhang, S.J.; Zhang, L.H.; Lin, Y.M.; Wei, S.D.; Liao, M.M.; Lin, G.H. Age-related changes in nutrient resorption patterns and tannin concentration of Casuarina equisetifolia plantations. J. Trop. For. Sci. 2012, 24, 546–556. [Google Scholar] [CrossRef]
- Lal, C.B.; Annapurna, C.; Raghubanshi, A.S.; Singh, J.S. Foliar demand and resource economy of nutrients in dry tropical forest species. J. Veg. Sci. 2001, 12, 5–14. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Li, L.H.; Han, X.G.; Huang, J.H.; Jiang, G.M.; Wan, S.Q.; Zhang, W.H.; Chen, Q.S. Nitrogen resorption from senescing leaves in 28 plant species in a semi-arid region of northern China. J. Arid Environ. 2005, 63, 191–202. [Google Scholar] [CrossRef]
- Pang, D.B.; Wang, G.Z.; Li, G.J.; Sun, Y.L.; Liu, Y.G.; Zhou, J.X. Ecological stoichiometric characteristics of two typical plantations in the Karst ecosystem of southwestern China. Forests 2018, 9, 56. [Google Scholar] [CrossRef]
- MacDicken, K.; Jonsson, Ö.; Piña, L.; Marklund, L.; Maulo, S.; Contessa, V.; Adikari, Y.; Garzuglia, M.; Lindquist, E.; Reams, G.; et al. Global Forest Resources Assessment 2015: How Are the World’S Forests Changing? Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2015; pp. 1–54. [Google Scholar]
- Van Dijk, A.I.J.M.; Keenan, R.J. Planted forests and water in perspective. For. Ecol. Manag. 2007, 251, 1–9. [Google Scholar] [CrossRef]
- Fang, J.Y.; Chen, A.P.; Peng, C.H.; Zhao, S.Q.; Ci, L.J. Changes in forest biomass carbon storage in China between 1949 and 1998. Science 2001, 292, 2320–2322. [Google Scholar] [CrossRef] [PubMed]
- Goodale, C.L.; Apps, M.J.; Birdsey, R.A.; Field, C.B.; Heath, L.S.; Houghton, R.A.; Jenkins, J.C.; Kohlmaier, G.H.; Kurz, W.; Liu, S.; et al. Forest carbon sinks in the northern Hemisphere. Ecol. Appl. 2002, 12, 891–899. [Google Scholar] [CrossRef]
- Netzer, F.; Schmid, C.; Herschbach, C.; Rennenberg, H. Phosphorus-nutrition of European beech (Fagus sylvatica L.) during annual growth depends on tree age and P-availability in the soil. Environ. Exp. Bot. 2017, 137, 194–207. [Google Scholar] [CrossRef]
- Weatherall, A.; Proe, M.F.; Craig, J.; Cameron, A.D.; Midwood, A.J. Internal cycling of nitrogen, potassium and magnesium in young Sitka spruce. Tree Physiol. 2006, 26, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.S.; Niklas, K.J.; Liu, Y.; Fang, X.M.; Wan, S.Z.; Wang, H. Nitrogen and phosphorus additions alter nutrient dynamics but not resorption efficiencies of Chinese fir leaves and twigs differing in age. Tree Physiol. 2015, 35, 1106–1117. [Google Scholar] [CrossRef] [PubMed]
- Reed, S.C.; Townsend, A.R.; Davidson, E.A.; Cleveland, C.C. Stoichiometric patterns in foliar nutrient resorption across multiple scales. New Phytol. 2012, 196, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Van Heerwaarden, L.M.; Toet, S.; Aerts, R. Current measures of nutrient resorption efficiency lead to a substantial underestimation of real resorption efficiency: Facts and solutions. Oikos 2003, 101, 664–669. [Google Scholar] [CrossRef]
- Oleksyn, J.; Reich, P.B.; Zytkowiak, R.; Karolewski, P.; Tjoelker, M.G. Nutrient conservation increases with latitude of origin in European Pinus sylvestris populations. Oecologia 2003, 136, 220–235. [Google Scholar] [CrossRef] [PubMed]
- Bengtson, P.; Falkengren-Grerup, U.; Bengtsson, G. Relieving substrate limitation-soil moisture and temperature determine gross N transformation rates. Oikos 2005, 111, 81–90. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef] [PubMed]
- Cramer, M.D.; Hawkins, H.J.; Verboom, G.A. The importance of nutritional regulation of plant water flux. Oecologia 2009, 161, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Wright, I.J.; Westoby, M. Nutrient concentration, resorption and lifespan leaf traits of Australian sclerophyll species. Funct. Ecol. 2003, 17, 10–19. [Google Scholar] [CrossRef]
- Augusto, L.; Achat, D.L.; Jonard, M.; Vidal, D.; Ringeval, B. Soil parent material-A major driver of plant nutrient limitations in terrestrial ecosystems. Glob. Chang. Biol. 2017, 23, 3808–3824. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.P.; Chen, X.; Huang, B.; Chi, G.Y. Distribution changes of phosphorus in soil-plant systems of larch plantations across the chronosequence. Forests 2018, 9, 563. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Li, L.H.; Han, X.G.; Huang, J.H.; Wan, S.Q. Foliar nitrogen dynamics and nitrogen resorption of a sandy shrub Salix gordejevii in northern China. Plant Soil 2005, 278, 183–193. [Google Scholar] [CrossRef]
- Liu, J.T.; Gu, Z.J.; Shao, H.B.; Zhou, F.; Peng, S.Y. N–P stoichiometry in soil and leaves of Pinus massoniana forest at different stand ages in the subtropical soil erosion area of China. Environ. Earth Sci. 2016, 75, 1091. [Google Scholar] [CrossRef]
- Escudero, A.; Del Arco, J.M.; Sanz, I.C.; Ayala, J. Effects of leaf longevity and retranslocation efficiency on the retention time of nutrients in the leaf biomass of different woody species. Oecologia 1992, 90, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Norris, M.D.; Reich, P.B. Modest enhancement of nitrogen conservation via retranslocation in response to gradients in N supply and leaf N status. Plant Soil 2009, 316, 193–204. [Google Scholar] [CrossRef]
- Zhao, G.S.; Shi, P.L.; Wu, J.S.; Xiong, D.P.; Zong, N.; Zhang, X.Z. Foliar nutrient resorption patterns of four functional plants along a precipitation gradient on the Tibetan Changtang Plateau. Ecol. Evol. 2017, 7, 7201–7212. [Google Scholar] [CrossRef] [PubMed]
- Hayes, P.; Turner, B.L.; Lambers, H.; Laliberte, E. Foliar nutrient concentrations and resorption efficiency in plants of contrasting nutrient-acquisition strategies along a 2-million-year dune chronosequence. J. Ecol. 2014, 102, 396–410. [Google Scholar] [CrossRef]
- Deng, M.F.; Liu, L.L.; Sun, Z.Z.; Piao, S.L.; Ma, Y.C.; Chen, Y.W.; Wang, J.; Qiao, C.L.; Wang, X.; Li, P. Increased phosphate uptake but not resorption alleviates phosphorus deficiency induced by nitrogen deposition in temperate Larix principis-rupprechtii plantations. New Phytol. 2016, 212, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Chen, Q.; Elser, J.J.; He, N.; Wu, H.; Zhang, G.; Wu, J.; Bai, Y.; Han, X. Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability. Ecol. Lett. 2010, 13, 1390–1399. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.N.; Lu, J.Y.; Yang, H.M.; Zhang, X.; Luo, C.L.; Zhao, Y.X. Resorption of nitrogen, phosphorus and potassium from leaves of lucerne stands of different ages. Plant Soil 2014, 383, 301–312. [Google Scholar] [CrossRef]
- Kobe, R.K.; Lepczyk, C.A.; Iyer, M. Resorption efficiency decreases with increasing green leaf nutrients in a global data set. Ecology 2005, 86, 2780–2792. [Google Scholar] [CrossRef]
- Tsujii, Y.; Onoda, Y.; Kitayama, K. Phosphorus and nitrogen resorption from different chemical fractions in senescing leaves of tropical tree species on Mount Kinabalu, Borneo. Oecologia 2017, 185, 171–180. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Resorption | Fixed Term | df | F, p |
|---|---|---|---|
| NRE | Climate zones | 3 | 3.56 * |
| Broadleaf vs. Conifer | 1 | 0.56 | |
| Deciduous vs. Evergreen | 1 | 2.48 | |
| Non-N-fixing tree vs. N-fixing tree | 1 | 15.85 *** | |
| PRE | Climate zones | 3 | 0.79 |
| Broadleaf vs. Conifer | 1 | 4.74 * | |
| Deciduous vs. Evergreen | 1 | 12.93 *** | |
| Non-N-fixing tree vs. N-fixing tree | 1 | 5.06 * |
| Variables | Regression | n | r2 | Excluded Variables | |
|---|---|---|---|---|---|
| NRE | MAP | y = 67.48 − 0.01 × MAP | 476 | 0.09 *** | Age, Ngreen, Pgreen, LAT |
| MAP, Pgreen | y = 62.65 − 0.01 × MAP + 2.70 × Pgreen | 476 | 0.12 *** | Age, Ngreen, LAT | |
| MAP, Pgreen, LAT | y = 54.68 − 0.01 × MAP + 2.36 × Pgreen + 0.21 × LAT | 476 | 0.14 *** | Age, Ngreen | |
| PRE | Pgreen | y = 66.11 − 4.12 × Pgreen | 476 | 0.05 *** | Age, Ngreen, LAT, MAP |
| Pgreen, Ngreen | y = 60.31 − 5.33 × Pgreen + 0.41 × Ngreen | 476 | 0.07 *** | Age, LAT, MAP |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, D.; Geng, Q.; Li, Q.; Luo, Y.; Vogel, J.; Shi, Z.; Ruan, H.; Xu, X. Nitrogen and Phosphorus Resorption in Planted Forests Worldwide. Forests 2019, 10, 201. https://doi.org/10.3390/f10030201
Jiang D, Geng Q, Li Q, Luo Y, Vogel J, Shi Z, Ruan H, Xu X. Nitrogen and Phosphorus Resorption in Planted Forests Worldwide. Forests. 2019; 10(3):201. https://doi.org/10.3390/f10030201
Chicago/Turabian StyleJiang, Dalong, Qinghong Geng, Qian Li, Yiqi Luo, Jason Vogel, Zheng Shi, Honghua Ruan, and Xia Xu. 2019. "Nitrogen and Phosphorus Resorption in Planted Forests Worldwide" Forests 10, no. 3: 201. https://doi.org/10.3390/f10030201
APA StyleJiang, D., Geng, Q., Li, Q., Luo, Y., Vogel, J., Shi, Z., Ruan, H., & Xu, X. (2019). Nitrogen and Phosphorus Resorption in Planted Forests Worldwide. Forests, 10(3), 201. https://doi.org/10.3390/f10030201