Effect of Phosphinothricin on Transgenic Downy Birch (Betula pubescens Ehrh.) Containing bar or GS1 Genes



Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Visual Injury after Herbicide Treatment
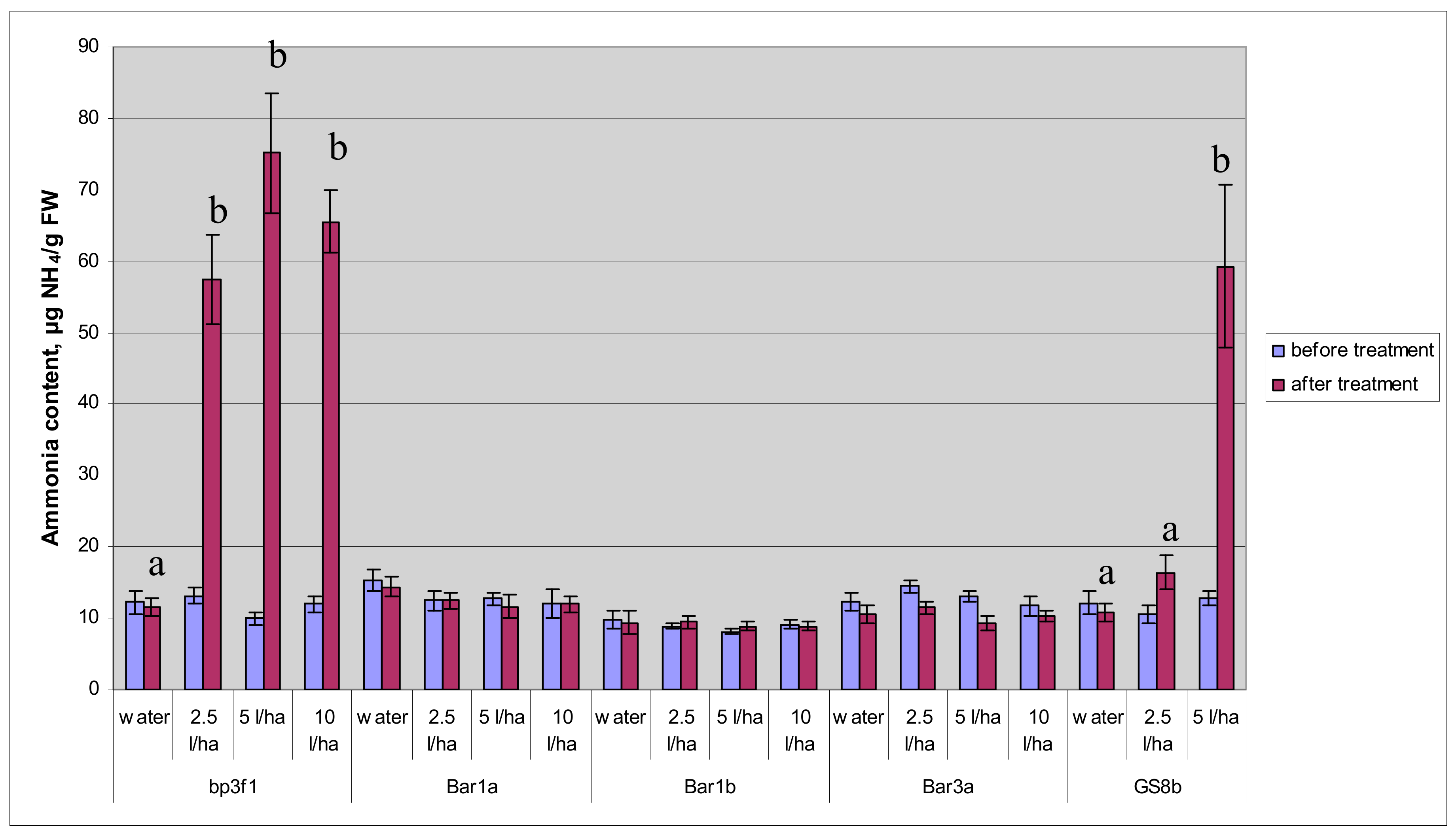
3.2. Ammonium Accumulation
3.3. Leaf Pigments (Chlorophyll and Carotenoids) Content
3.4. Relative Water Content (RWC)
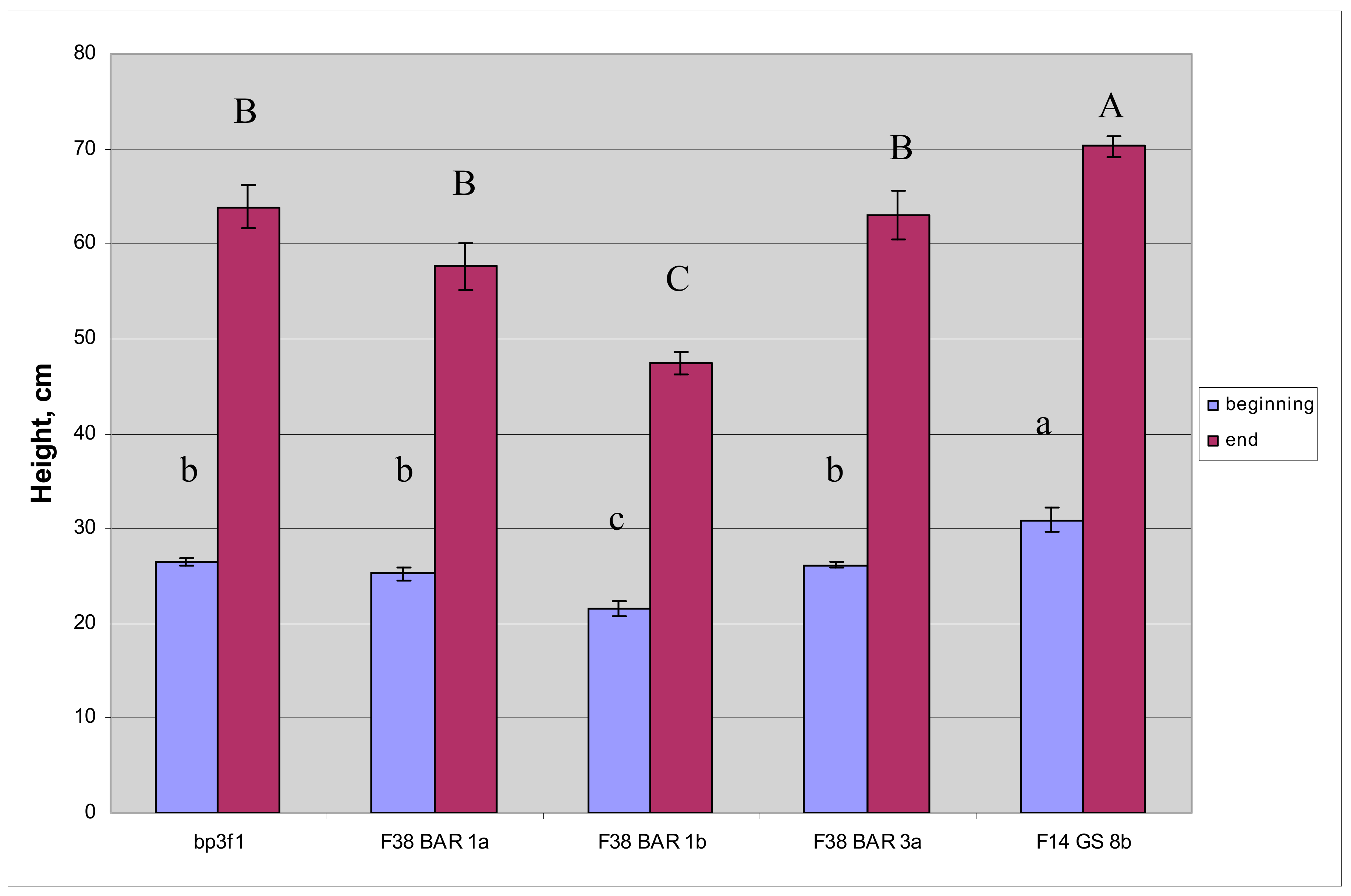
3.5. Height and Biomass
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Dayan, F.E.; Cantrell, C.L.; Duke, S.O. Natural products in crop protection. Bioorg. Medic. Chem. 2009, 17, 4022–4034. [Google Scholar] [CrossRef] [PubMed]
- Forde, B.G.; Lea, P.J. Glutamate in plants: Metabolism, regulation and signalling. J. Exp. Bot. 2007, 58, 2339–2358. [Google Scholar] [CrossRef] [PubMed]
- OECD. Consensus Document on General Information Concerning the Genes and their Enzymes that Confer Tolerance to Phosphinothricin Herbicide, SERIES on Harmonization of Regulatory Oversight in Biotechnology No. 11, 1999, OECD, Paris. Available online: www.oecd.org/env/biotrack/46815628.pdf (accessed on 30 August 2019).
- Kita, Y.; Hanafy, M.; Deguchi, M. Generation and characterization of herbic ide-resistant soybean plants expressing novel phosphinothricin N-acetyltransferase genes. Breed. Sci. 2009, 59, 245–251. [Google Scholar] [CrossRef][Green Version]
- Cui, Y.; Liu, Z.; Li, Y.; Zhou, F.; Chen, H.; Lin, Y. Corrigendum: Application of a novel phosphinothricin N-acetyltransferase (RePAT) gene in developing glufosinate-resistant rice. Sci. Rep. 2018, 8, 46937. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Lee, O.R.; Lee, S.; Kim, K.-T.; Yang, D.-C. Isolation and characterization of a theta glutathione S-transferase gene from Panax ginseng Meyer. J. Ginseng Res. 2012, 36, 449–460. [Google Scholar] [CrossRef]
- Pascual, M.B.; Jing, Z.P.; Kirby, E.G.; Canovas, F.M.; Gallardo, F. Response of transgenic poplar overexpressing cytosolic glutamine synthetase to phosphinothricin. Phytochemistry 2008, 69, 382–389. [Google Scholar] [CrossRef]
- Cai, H.; Zhou, Y.; Xiao, J.; Li, X.; Zhang, Q.; Lian, X. Overexpressed glutamine synthetase gene modifies nitrogen metabolism and abiotic stress responses in rice. Plant Cell Rep. 2009, 28, 527–537. [Google Scholar] [CrossRef]
- Tian, Y.-S.; Xu, J.; Zhao, W.; Xing, X.-J.; Fu, X.-Y.; Peng, R.-H.; Yao, Q.-H. Identification of a phosphinothricin-resistant mutant of rice glutamine synthetase using DNA shuffling. Sci. Rep. 2015, 5, 15495. [Google Scholar] [CrossRef]
- Abdeen, A.; Miki, B. The pleiotropic effects of the bar gene and glufosinate on the Arabidopsis transcriptome. Plant Biotech. J. 2009, 7, 266–282. [Google Scholar] [CrossRef]
- Tachibana, K.; Watanabe, T.; Sekizawa, Y.; Takematsu, T. Accumulation of ammonium in plants treated with bialaphos. J. Pestic. Sci. 1986, 11, 33–37. [Google Scholar] [CrossRef]
- Tachibana, K.; Watanabe, T.; Sekizawa, Y.; Takematsu, T. Inhibition of glutamine synthetase and quantitative changes of free amino acids in shoots of bialaphos-treated Japanese barnyard millet. J. Pestic. Sci. 1986, 11, 27–31. [Google Scholar] [CrossRef][Green Version]
- Wendler, C.; Barniske, M.; Wild, A. Effect of phosphinothricin (glufosinate) on photosynthesis and photorespiration of C3 and C4 plants. Photosynt. Res. 1990, 24, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Downs, C.G.; Christey, M.C.; Maddocks, J.F.S.; Stevenson, D.G. Hairy roots of Brassica napus: I. Applied glutamine overcomes the effect of phosphinothricin treatment. Plant Cell Rep. 1994, 14, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Takano, H.K.; Beffa, R.; Preston, C.; Westra, P.; Dayan, F.E. Reactive oxygen species trigger the fast action of glufosinate. Planta 2019, 249, 1837–1849. [Google Scholar] [CrossRef] [PubMed]
- Heap, I.; Duke, S.O. Overview of glyphosate-resistant weeds worldwide. Pest Manag. Sci. 2018, 74, 1040–1049. [Google Scholar] [CrossRef]
- Jalaludin, A.; Yu, Q.; Zoellner, P.; Beffac, R.; Powlesa, S.B. Characterisation of glufosinate resistance mechanisms in Eleusine indica. Pest Manag. Sci. 2017, 73, 1091–1100. [Google Scholar] [CrossRef] [PubMed]
- McNabb, K.; South, D.B.; Mitchell, R.J. Weed management systems for forest nurseries and woodlands. In Handbook of Weed Management Systems; Smith, A.E., Ed.; Marcel Dekker: New York, NY, USA, 1995; pp. 667–771. [Google Scholar]
- Padegimas, L.; Shulga, O.A.; Skryabin, K.G. Phosphinothricin-resistant transgenic nicotiana-tabacum and solanum-tuberosum plants. Mol. Biol. 1994, 28, 294–297. [Google Scholar]
- Shestibratov, K.A.; Bulatova, I.V.; Novikov, P.S. In vitro response of transgenic aspen containing glutamine synthetase gene GSI to the sublethal dose of phosphinothricin. Appl. Biochem. Microbiol. 2010, 46, 763–768. [Google Scholar] [CrossRef]
- Lebedev, V.G.; Schestibratov, K.A.; Shadrina, T.E.; Bulatova, I.V.; Abramochkin, D.G.; Miroshnikov, A.I. Cotransformation of aspen and birch with three T-DNA regions from two different replicons in one Agrobacterium tumefaciens strain. Russ. J. Genet. 2010, 46, 1282–1289. [Google Scholar] [CrossRef]
- Shestibratov, K.; Lebedev, V.; Podrezov, A.; Salmova, M. Transgenic aspen and birch trees for Russian plantation forests. BMC Proc. 2011, 5 (Suppl. 7), 124. [Google Scholar] [CrossRef]
- De Block, M.; De Brouwer, D.; Tenning, P. Transformation of Brassica napus and Brassica oleracea using Agrobacterium tumefaciens and the expression of the bar and neo genes in the transgenic plants. Plant Physiol. 1989, 91, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Weatherburn, M. Phenol-hypochloride reaction for ammonium. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Barr, H.D.; Weatherley, P.E. A re-examination of the relative turgidity technique for estimating water deficit in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef]
- De Block, M. Factors influencing the tissue culture and the Agrobacterium tumefaciens-mediated transformation of hybrid aspen and poplar clones. Plant Physiol. 1990, 93, 1110–1116. [Google Scholar] [CrossRef]
- Confalonieri, M.; Belenghi, B.; Balestrazzi, A.; Negri, S.; Facciotto, G.; Schenone, G.; Delledonne, M. Transformation of elite white poplar (Populus alba L.) cv. ‘Villafranca’ and evaluation of herbicide resistance. Plant Cell Rep. 2000, 19, 978–982. [Google Scholar] [CrossRef]
- Lebedev, V.G.; Faskhiev, V.N.; Kovalenko, N.P.; Schestibratov, K.A.; Miroshnikov, A.I. Testing transgenic aspen plants with bar gene for herbicide resistance under semi-natural conditions. Acta Nat. 2016, 8, 92–101. [Google Scholar] [CrossRef]
- Harcourt, R.L.; Kyozuka, J.; Floyd, R.B.; Bateman, K.S.; Tanaka, H.; Decroocq, V.; Llewellyn, D.J.; Zhu, X.; Peacock, W.J.; Dennis, E.S. Insect- and herbicide-resistant transgenic eucalypts. Mol. Breed. 2000, 6, 307–315. [Google Scholar] [CrossRef]
- Quinn, J.P. Evolving strategies for the genetic engineering of herbicide resistance in plants. Biotech. Adv. 1990, 8, 321–333. [Google Scholar] [CrossRef]
- Merino, I.; Contreras, A.; Jing, Z.-P.; Gallardo, F.; Cánovas, F.M.; Gómez, L. Plantation forestry under global warming: Hybrid poplars with improved thermotolerance provide new insights on the in vivo function of small heat shock protein chaperones. Plant Physiol. 2014, 164, 978–991. [Google Scholar] [CrossRef] [PubMed]
- Nolte, S.A.; Young, B.G.; Mungur, R.; Lightfoot, D.A. The glutamate dehydrogenase gene gdhA increased the resistance of tobacco to glufosinate. Weed Res. 2004, 44, 335–339. [Google Scholar] [CrossRef]
- Miflin, B.J.; Lea, P.J. Amino acid metabolism. Annu. Rev. Plant Physiol. 1997, 28, 299–329. [Google Scholar] [CrossRef]
- Asano, Y.; Ito, Y.; Fukami, M.; Sugiura, K.; Fujiie, A. Herbicide-resistant transgenic creeping bentgrass plants obtained by electroporation using an altered buffer. Plant Cell Rep. 1998, 17, 963–967. [Google Scholar] [CrossRef] [PubMed]
- Pornprom, T.; Chompoo, J.; Grace, B. Glufosinate tolerance in hybrid corn varieties based on decreasing ammonium accumulation. Weed Biol. Manag. 2003, 3, 41–45. [Google Scholar] [CrossRef]
- Krenchinski, F.H.; Carbonari, C.A.; Cesco, V.J.S.; Albrecht, A.J.P.; de Lara Campos Arcuri, M.; de Godoy Maia, I.; Velini, E.D. Glufosinate resistance level is proportional to phosphinothricin acetyltransferase gene expression in glufosinate-resistant maize. J. Agric. Food Chem. 2018, 66, 12641–12650. [Google Scholar] [CrossRef]
- James, D.; Borphukan, B.; Fartyal, D.; Ram, B.; Singh, J.; Manna, M.; Sheri, V.; Panditi, V.; Yadav, R.; Achary, V.M.M. Concurrent Overexpression of OsGS1;1 and OsGS2 genes in transgenic rice (Oryza sativa L.): Impact on tolerance to abiotic stresses. Front. Plant Sci. 2018, 9, 786. [Google Scholar] [CrossRef] [PubMed]
- Carbonari, C.A.; Latorre, D.O.; Gomes, G.L.; Velini, E.D.; Owens, D.K.; Pan, Z.; Dayan, F.E. Resistance to glufosinate is proportional to phosphinothricin acetyltransferase expression and activity in LibertyLink(®) and WideStrike(®) cotton. Planta 2016, 243, 925–933. [Google Scholar] [CrossRef]
- Tsai, C.J.; Wang, C.S.; Wang, C.Y. Physiological characteristics of glufosinate resistance in rice. Weed Sci. 2006, 54, 634–640. [Google Scholar] [CrossRef]
- Coetzer, E.; Al-Khatib, K. Photosynthetic inhibition and ammonium accumulation in Palmer amaranth after glufosinate application. Weed Sci. 2001, 49, 454–459. [Google Scholar] [CrossRef]
- De Block, M.; Sonville, A.D.; Debrouwer, D. The selection mechanism of phosphinothricin is influenced by the metabolic status of the tissue. Planta 1995, 197, 619–626. [Google Scholar] [CrossRef]
- Barsch, A.; Carvalho, H.G.; Cullimore, J.V.; Niehaus, K. GC–MS based metabolite profiling implies three interdependent ways of ammonium assimilation in Medicago truncatula root nodules. J. Biotech. 2006, 127, 79–83. [Google Scholar] [CrossRef]
- Salas-Perez, R.A.; Saski, C.A.; Noorai, R.E.; Srivastava, S.K.; Lawton-Rauh, A.L.; Nichols, R.L.; Roma-Burgos, N. RNA-Seq transcriptome analysis of Amaranthus palmeri with differential tolerance to glufosinate herbicide. PLoS ONE 2011, 13, e0195488. [Google Scholar] [CrossRef]
- Ahn, I.-P. Glufosinate ammonium-induced pathogen inhibition and defense responses culminate in disease protection in bar-transgenic rice. Plant Physiol. 2008, 146, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Selzer, L.J.; Busso, C.A. Pigments and photosynthesis of understory grasses: Light irradiance and soil moisture effects. Russ. J. Plant Physiol. 2016, 63, 224–234. [Google Scholar] [CrossRef]
- Keča, N.; Tkaczyk, M.; Żółciak, A.; Stocki, M.; Kalaji, H.M.; Nowakowska, J.A.; Oszako, T. Survival of European ash seedlings treated with phosphite after infection with the Hymenoscyphus fraxineus and Phytophthora species. Forests 2018, 9, 442. [Google Scholar] [CrossRef]
- Reddy, K.N.; Zablotowicz, R.M.; Bellaloui, N.; Ding, W. Glufosinate effects on nitrogen nutrition, growth, yield, and seed composition in glufosinate-resistant and glufosinate-sensitive soybean. Int. J. Agron. 2011, 2011, 109280. [Google Scholar] [CrossRef]
- Li, Z.; Keasling, J.D.; Niyogi, K.K. Overlapping photoprotective function of vitamin E and carotenoids in Chlamydomonas. Plant Physiol. 2012, 158, 313–323. [Google Scholar] [CrossRef]
- Kozaki, A.; Takeba, G. Photorespiration protects C3 plants from photooxidation. Nature 1996, 384, 557–560. [Google Scholar] [CrossRef]
- Yamasaki, S.; Dillenburg, L.R. Measurements of leaf relative water content in Araucaria angustifolia. Rev. Bras. Fisiol. Veg. 1999, 11, 69–75. [Google Scholar]
- Dayan, F.E. Current status and future prospects in herbicide discovery. Plants 2019, 8, 341. [Google Scholar] [CrossRef] [PubMed]
- Meilan, R.; Han, K.-H.; Ma, C.; DiFazio, S.P.; Eaton, J.A.; Hoien, E.A.; Stanton, B.J.; Crockett, R.P.; Taylor, M.L.; James, R.R.; et al. The CP4 transgene provides high levels of tolerance to Roundup herbicide in field-grown hybrid poplars. Can. J. For. Res. 2002, 32, 967–976. [Google Scholar] [CrossRef]
- Gallardo, F.; Fu, J.; Canton, F.R.; Garcia-Gutierrez, A.; Canovas, F.M.; Kirby, E.G. Expression of a conifer glutamine synthetase gene in transgenic poplar. Planta 1999, 210, 19–26. [Google Scholar] [CrossRef] [PubMed]
- García, M.J.; Palma-Bautista, C.; Rojano-Delgado, A.M.; Bracamonte, E.; Portugal, J.; Alcántara-de la Cruz, R.; De Prado, R. The triple amino acid substitution TAP-IVS in the EPSPS gene confers high glyphosate resistance to the superweed Amaranthus hybridus. Int. J. Mol. Sci. 2019, 20, 2396. [Google Scholar] [CrossRef]
- Brunharo, C.; Takano, H.; Mallory-Smith, C.; Dayan, F.; Hanson, B. Role of Glutamine synthetase isogenes and herbicide metabolism in the mechanism of resistance to glufosinate in Lolium perenne L. spp. multiflorum biotypes from Oregon. J. Agric. Food Chem. 2019, 67, 8431–8440. [Google Scholar] [CrossRef]
- Alcántara-de la Cruz, R.; Domínguez-Martínez, P.A.; da Silveira, H.M.; Cruz-Hipólito, H.E.; Palma-Bautista, C.; Vázquez-García, J.G.; Domínguez-Valenzuela, J.A.; De Prado, R. Management of glyphosate-resistantweeds in Mexican citrus groves: Chemical alternatives and economic viability. Plants 2019, 8, 325. [Google Scholar] [CrossRef]
- Christ, B.; Hochstrasser, R.; Guyer, L.; Francisco, R.; Aubry, S.; Hörtensteiner, S.; Weng, J.K. Non-specific activities of the major herbicide-resistance gene BAR. Nat. Plants 2017, 3, 937–945. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Birch Line | Concentration of the Basta Herbicide Solution% | Visual Scores of Necrosis Measured in 3, 7, and 14, or 28 Days after Treatment | ||
|---|---|---|---|---|
| 3 | 7 | 14 or 28 | ||
| bp3f1 (non-transgenic control) | 0.5 | 2–3 | 4 | 5 |
| 1 | 2–3 | 4 | 5 | |
| 2 | 3 | 5 | 5 | |
| Bar1a | 0.5 | 0 | 0 | 0 |
| 1 | 0 | 0 | 0–1 * | |
| 2 | 0 | 0–1 * | 0–1 ** | |
| Bar1b | 0.5 | 0 | 0 | 0 |
| 1 | 0 | 0 | 0 | |
| 2 | 0 | 0–1 * | 0–1 ** | |
| Bar3a | 0.5 | 0 | 0 | 0 |
| 1 | 0 | 0 | 0 | |
| 2 | 0 | 0–1 * | 0–1 ** | |
| GS8b | 0.5 | 2 | 4 | 5 |
| 1 | 2–3 | 4 | 5 | |
| Birch Line | Concentration of the Basta Herbicide Solution % | Visual Scores of Necrosis Measured in 1, 3, 7, 14, and 28 Days after Treatment | ||||
|---|---|---|---|---|---|---|
| 1 | 3 | 7 | 14 | 28 | ||
| bp3f1 (non-transgenic control) | 0.5% | 0–1 ** | 1–2 | 3–4 | 4 | 4–5 **** |
| 1% | 0–1 ** | 1–2 | 4 | 5 | 5 | |
| 2% | 0–1 *** | 1–2 | 4 | 5 | 5 | |
| Bar1a | 0.5% | 0 | 0 | 0 | 0 | 0 |
| 1% | 0 | 0 | 0–1 ** | 0–1 ** | 0–1 ** | |
| 2% | 0 | 0–1 * | 0–1 ** | 0–1 ** | 0–1 ** | |
| Bar1b | 0.5% | 0 | 0 | 0 | 0 | 0 |
| 1% | 0 | 0 | 0–1 ** | 0–1 ** | 0–1 ** | |
| 2% | 0 | 0–1 * | 0–1 ** | 0–1 *** | 0–1 *** | |
| Bar3a | 0.5% | 0 | 0 | 0 | 0 | 0 |
| 1% | 0 | 0 | 0–1 ** | 0–1 ** | 0–1 ** | |
| 2% | 0 | 0–1 * | 0–1 ** | 0–1 ** | 0–1 ** | |
| GS8b | 0.5% | 0–1 ** | 1–2 | 2–3 | 4 | 4–5 **** |
| 1% | 0–1 ** | 1–2 | 3–4 | 5 | 5 | |
| Birch Line | Herbicide Content in Water % | Days after Treatment 1 | Chlorophyll 2, µg/mg FW | Carotenoids 2, µg/mg FW | ||
|---|---|---|---|---|---|---|
| A | B | A + B | ||||
| bp3f1 (non-transgenic control) | 0 (control) | 0 | 1.860 | 0.409 | 2.269 | 0.632 |
| 3 | 1.847 | 0.429 | 2.276 | 0.585 | ||
| 7 | 1.838 | 0.414 | 2.252 | 0.537 | ||
| 0.5 | 0 | 1.716 a | 0.381 | 2.097 | 0.650 a | |
| 3 | 1.498 b | 0.391 | 1.889 | 0.468 b | ||
| 7 | - | - | - | - | ||
| 1 | 0 | 1.807 a | 0.392 | 2.199 a | 0.647 a | |
| 3 | 1.474 b | 0.356 | 1.830 b | 0.475 b | ||
| 7 | - | - | - | - | ||
| 2 | 0 | 1.967 a | 0.441 a | 2.409 a | 0.691 a | |
| 3 | 1.378 b | 0.361 b | 1.739 b | 0.416 b | ||
| 7 | - | - | - | - | ||
| Bar1a | 0 (control) | 0 | 1.715 | 0.385 | 2.100 | 0.581 |
| 3 | 1.692 | 0.391 | 2.083 | 0.590 | ||
| 7 | 1.744 | 0.417 | 2.161 | 0.616 | ||
| 0.5 | 0 | 1.754 | 0.427 | 2.181 | 0.551 | |
| 3 | 1.782 | 0.457 | 2.239 | 0.607 | ||
| 7 | 1.729 | 0.429 | 2.158 | 0.618 | ||
| 1 | 0 | 1.814 | 0.416 | 2.230 | 0.587 | |
| 3 | 1.730 | 0.439 | 2.169 | 0.548 | ||
| 7 | 1.748 | 0.424 | 2.172 | 0.517 | ||
| 2 | 0 | 1.791 | 0.391 | 2.182 | 0.617 | |
| 3 | 1.741 | 0.409 | 2.150 | 0.574 | ||
| 7 | 1.665 | 0.364 | 2.029 | 0.549 | ||
| Bar1b | 0 (control) | 0 | 1.781 | 0.449 | 2.230 | 0.625 |
| 3 | 1.760 | 0.439 | 2.199 | 0.575 | ||
| 7 | 1.870 | 0.446 | 2.316 | 0.575 | ||
| 0.5 | 0 | 2.101 | 0.606 | 2.708 | 0.702 | |
| 3 | 1.945 | 0.564 | 2.509 | 0.667 | ||
| 7 | 1.913 | 0.552 | 2.464 | 0.658 | ||
| 1 | 0 | 1.997 | 0.519 | 2.517 | 0.717 | |
| 3 | 2.031 | 0.537 | 2.569 | 0.656 | ||
| 7 | 1.891 | 0.513 | 2.404 | 0.567 | ||
| 2 | 0 | 2.040 | 0.518 | 2.558 | 0.712 a | |
| 3 | 1.980 | 0.523 | 2.503 | 0.569 ab | ||
| 7 | 1.693 | 0.454 | 2.148 | 0.529 b | ||
| Bar3a | 0 (control) | 0 | 1.748 | 0.404 | 2.152 | 0.588 |
| 3 | 1.793 | 0.411 | 2.204 | 0.583 | ||
| 7 | 1.675 | 0.399 | 2.074 | 0.513 | ||
| 0.5 | 0 | 1.646 | 0.361 | 2.007 | 0.591 | |
| 3 | 1.627 | 0.350 | 1.976 | 0.551 | ||
| 7 | 1.658 | 0.379 | 2.036 | 0.496 | ||
| 1 | 0 | 1.684 | 0.379 | 2.064 | 0.561 | |
| 3 | 1.452 | 0.306 | 1.759 | 0.497 | ||
| 7 | 1.586 | 0.370 | 1.956 | 0.483 | ||
| 2 | 0 | 1.617 a | 0.325 | 1.941 a | 0.592 a | |
| 3 | 1.399 b | 0.294 | 1.692 b | 0.472 b | ||
| 7 | 1.151 c | 0.261 | 1.411 c | 0.381 c | ||
| GS8b | 0 (control) | 0 | 1.560 | 0.343 | 1.903 | 0.595 |
| 3 | 1.619 | 0.365 | 1.984 | 0.535 | ||
| 7 | 1.774 | 0.389 | 2.163 | 0.607 | ||
| 0.5 | 0 | 1.579 | 0.354 | 1.933 | 0.580 | |
| 3 | 1.592 | 0.345 | 1.937 | 0.541 | ||
| 7 | - | - | - | - | ||
| 1 | 0 | 1.602 a | 0.397 a | 1.999 a | 0.601 a | |
| 3 | 1.257 b | 0.279 b | 1.536 b | 0.470 b | ||
| 7 | - | - | - | - | ||
| Birch Line | Herbicide Content in Water % | First Season 1 | Second Season 1 | |||
|---|---|---|---|---|---|---|
| Same Day | 3 Days | 7 Days | Same Day | 3 Days | ||
| bp3f1 (non-transgenic control) | 0 (control) | 82.2 | 82.4 a | 81.3 a | 83.6 | 82.9 a |
| 0.5 | 82.0 | 47.4 b | 28.6 b | 86.7 | 78.8 b | |
| 1 | 81.8 | 41.4 b | 29.3 b | 83.9 | 78.1 b | |
| 2 | 83.0 | 42.6 b | 22.1 b | 84.3 | 70.1 c | |
| Bar1a | 0 (control) | 82.6 | 83.2 | 81.5 | 87.3 | 87.6 |
| 0.5 | 82.2 | 83.5 | 81.2 | 89.5 | 87.2 | |
| 1 | 83.1 | 83.1 | 82.0 | 86.1 | 87.6 | |
| 2 | 82.9 | 78.6 | 79.3 | 86.5 | 86.7 | |
| Bar1b | 0 (control) | 82.2 | 81.4 a | 81.3 a | 84.6 | 84.5 |
| 0.5 | 83.1 | 82.1 a | 83.3 a | 85.9 | 86.3 | |
| 1 | 82.1 | 83.7 a | 82.9 a | 85.1 | 85.8 | |
| 2 | 81.5 | 72.7 b | 75.0 b | 86.5 | 86.4 | |
| Bar3a | 0 (control) | 83.3 | 83.0 | 82.2 | 83.7 | 85.3 |
| 0.5 | 81.5 | 82.7 | 82.9 | 86.9 | 87.0 | |
| 1 | 81.1 | 83.4 | 83.2 | 85.0 | 88.4 | |
| 2 | 82.9 | 77.7 | 77.9 | 84.6 | 85.5 | |
| GS8b | 0 (control) | 83.0 | 82.0 a | 81.9 a | 84.2 | 84.7 a |
| 0.5 | 82.8 | 64.5 b | 34.5 b | 86.4 | 82.8 ab | |
| 1 | 81.6 | 48.7 c | 30.5 b | 82.6 | 77.2 b | |
| Herbicide Content in Water % | Bar1a | Bar1b | Bar3a |
|---|---|---|---|
| 0 (control treatment) | 57.6 ± 2.5 | 47.4 ± 1.2 | 63.0 ± 2.6 |
| 0.5 | 57.8 ± 4.7 | 48.2 ± 3.4 | 63.9 ± 2.9 |
| 1 | 53.5 ± 2.4 | 51.1 ± 5.0 | 69.4 ± 3.0 |
| 2 | 54.1 ± 1.9 | 52.6 ± 1.1 | 64.7 ± 3.9 |
| Herbicide Content in Water % | Bar1a | Bar1b | Bar3a |
|---|---|---|---|
| 0 (control treatment) | 29.8 ± 3.4 | 24.9 ± 2.3 | 29.4 ± 2.1 |
| 0.5 | 24.7 ± 2.0 | 25.4 ± 1.6 | 23.5 ± 1.2 |
| 1 | 26.3 ± 2.7 | 21.6 ± 2.0 | 25.5 ± 1.6 |
| 2 | 24.5 ± 2.6 | 24.5 ± 1.9 | 24.4 ± 1.6 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lebedev, V.G.; Krutovsky, K.V.; Shestibratov, K.A. Effect of Phosphinothricin on Transgenic Downy Birch (Betula pubescens Ehrh.) Containing bar or GS1 Genes. Forests 2019, 10, 1067. https://doi.org/10.3390/f10121067
Lebedev VG, Krutovsky KV, Shestibratov KA. Effect of Phosphinothricin on Transgenic Downy Birch (Betula pubescens Ehrh.) Containing bar or GS1 Genes. Forests. 2019; 10(12):1067. https://doi.org/10.3390/f10121067
Chicago/Turabian StyleLebedev, Vadim G., Konstantin V. Krutovsky, and Konstantin A. Shestibratov. 2019. "Effect of Phosphinothricin on Transgenic Downy Birch (Betula pubescens Ehrh.) Containing bar or GS1 Genes" Forests 10, no. 12: 1067. https://doi.org/10.3390/f10121067
APA StyleLebedev, V. G., Krutovsky, K. V., & Shestibratov, K. A. (2019). Effect of Phosphinothricin on Transgenic Downy Birch (Betula pubescens Ehrh.) Containing bar or GS1 Genes. Forests, 10(12), 1067. https://doi.org/10.3390/f10121067