Spatial Patterns in Different Stages of Regeneration after Clear-Cutting of a Black Locust Forest in Central China



Abstract
1. Introduction
- (1)
- What are the regeneration spatial patterns of black locust trees in different growth stages following logging?
- (2)
- What is the spatial relation between stumps and regenerations?
- (3)
- What are the ecological processes behind these spatial patterns?
- (1)
- The extent of cluster of spatial pattern will show a downward trend with age due to impact of self-thinning and tending.
- (2)
- Spatial repulsion between stumps and ramets will happen at small scales, because sprouts of stumps suppress sprouts of roots and seedlings.
- (3)
- Silvicultural scheme, expansion and extension characteristics of black locust, and grazing will be the ecological processes behind these spatial patterns.
2. Materials and Methods
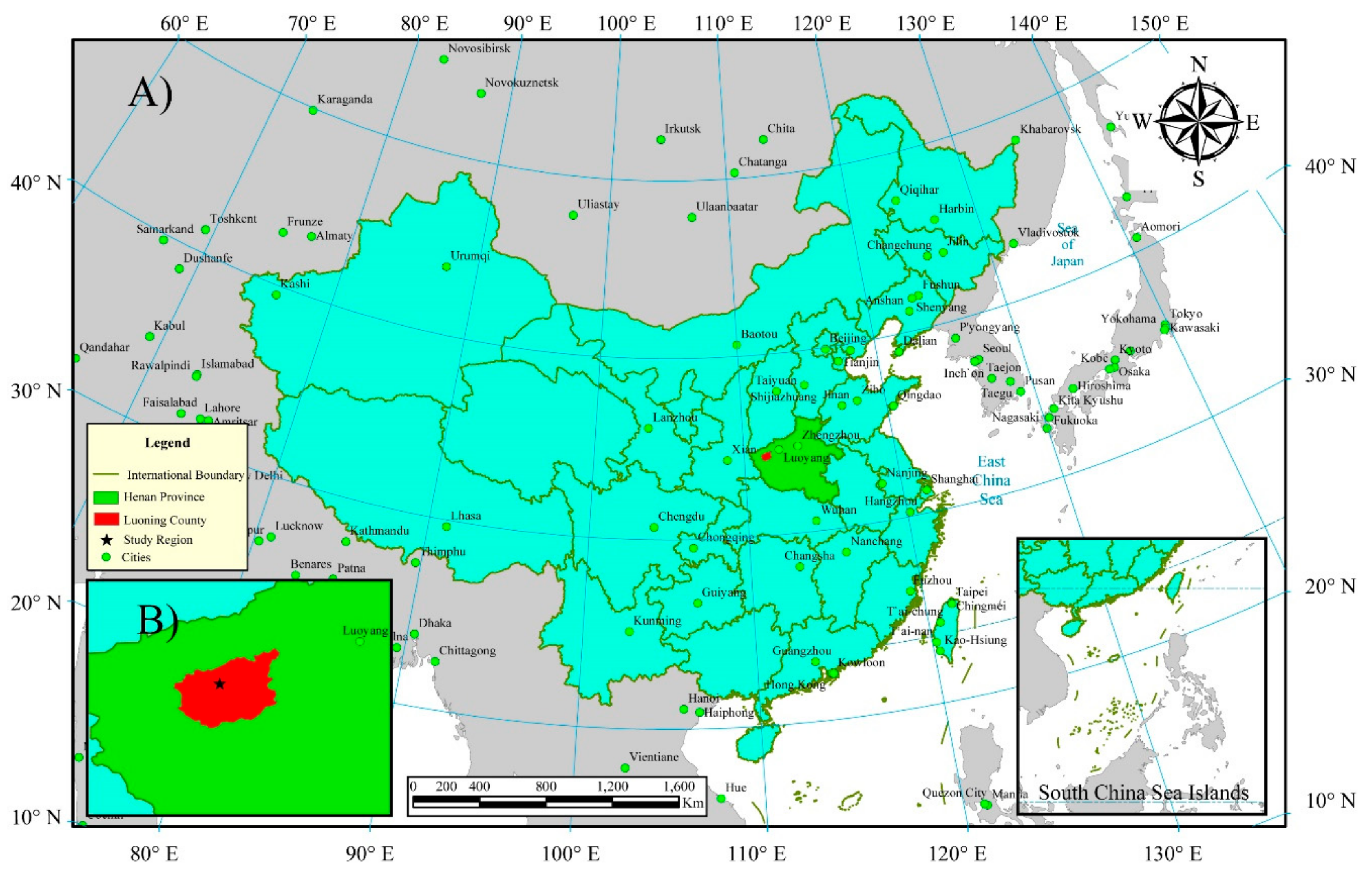
2.1. Study Area
2.2. Experiment Design and Data Collection
2.3. Data Analysis
2.3.1. Analysis of Basic Stand Characteristics
2.3.2. Correlation and Spacing Analysis
2.3.3. Fitting the Cluster Point Process to the Regeneration Data
3. Results
3.1. Basic Stand Characteristics
3.2. Correlation and Spacing Analysis
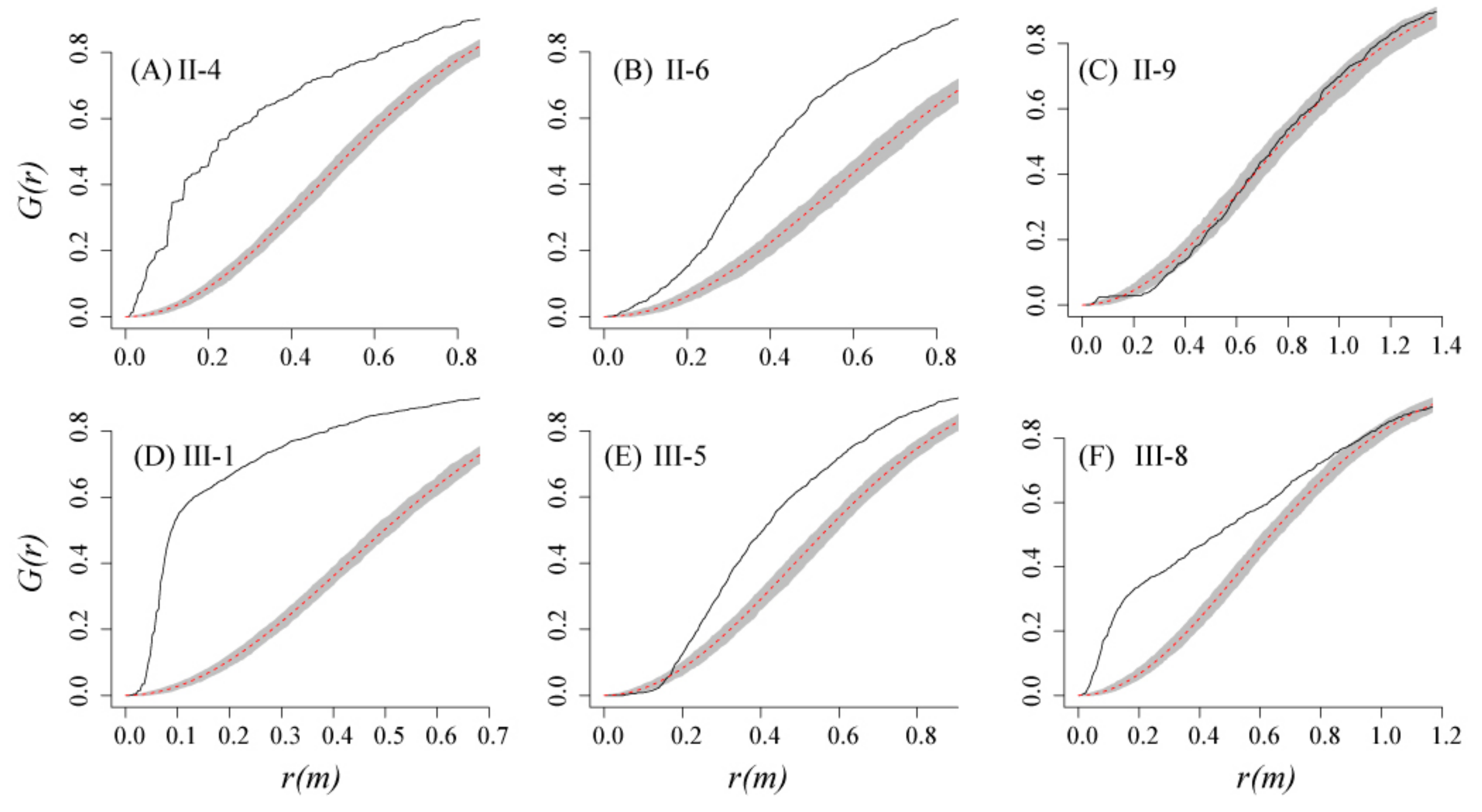
3.3. Fitting the Cluster Point Process to the Observed Data
4. Discussion
4.1. The Growth Differences in Intergenerational Stages
4.2. The Spatial Patterns in Different Growth Stages
4.3. The Ecological Processes behind These Spatial Patterns
4.4. Management Implications
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Yamagawa, H.; Ito, S.; Nakao, T. Restoration of semi-natural forest after clearcutting of conifer plantations in Japan. Landsc. Ecol. Eng. 2010, 6, 109. [Google Scholar] [CrossRef]
- Kruys, N.; Fridman, J.; Götmark, F.; Simonsson, P.; Gustafsson, L. Retaining trees for conservation at clearcutting has increased structural diversity in young Swedish production forests. For. Ecol. Manag. 2013, 304, 312–321. [Google Scholar] [CrossRef]
- Olson, M.G.; Meyer, S.R.; Wagner, R.G.; Seymour, R.S. Commercial thinning stimulates natural regeneration in spruce-fir stands. Can. J. For. Res. 2013, 44, 173–181. [Google Scholar] [CrossRef]
- Weis, W.; Rotter, V.; Göttlein, A. Water and element fluxes during the regeneration of Norway spruce with European beech: Effects of shelterwood-cut and clear-cut. For. Ecol. Manag. 2006, 224, 304–317. [Google Scholar] [CrossRef]
- Montoro Girona, M.; Lussier, J.M.; Morin, H.; Thiffault, N. Conifer regeneration after experimental shelterwood and seed-tree treatments in boreal forests: Finding silvicultural alternatives. Front. Plant. Sci. 2018, 9, 1145. [Google Scholar] [CrossRef]
- Boring, L.R.; Swank, W.T. The role of black locust (Robinia pseudoacacia) in forest succession. J. Ecol. 1984, 72, 749–766. [Google Scholar] [CrossRef]
- Yang, F. The Study on Biomass and Caloric Value of Locust Energy Forests in Luoning Hilly Region, Henan. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2013. [Google Scholar]
- Kurokochi, H.; Toyama, K.; Hogetsu, T. Regeneration of robinia pseudoacacia riparian forests after clear-cutting along the chikumagawa river in Japan. Plant Ecol. 2010, 210, 31–41. [Google Scholar] [CrossRef]
- Wang, B.; Liu, G.; Xue, S. Effect of black locust (Robinia pseudoacacia) on soil chemical and microbiological properties in the eroded hilly area of China’s loess plateau. Environ. Earth Sci. 2012, 65, 597–607. [Google Scholar] [CrossRef]
- Sitzia, T.; Campagnaro, T.; Dainese, M.; Cierjacks, A. Plant species diversity in alien black locust stands: A paired comparison with native stands across a north-Mediterranean range expansion. For. Ecol. Manag. 2012, 285, 85–91. [Google Scholar] [CrossRef]
- Call, L.J.; Nilsen, E.T. Analysis of interactions between the invasive tree-of-heaven (Ailanthus altissima) and the native black locust (Robinia pseudoacacia). Plant Ecol. 2005, 176, 275–285. [Google Scholar] [CrossRef]
- Benesperi, R.; Giuliani, C.; Zanetti, S.; Gennai, M.; Mariotti Lippi, M.; Guidi, T.; Nascimbene, J.; Foggi, B. Forest plant diversity is threatened by Robinia pseudoacacia(black-locust) invasion. Biodivers. Conserv. 2012, 21, 3555–3568. [Google Scholar] [CrossRef]
- Vítková, M.; Müllerová, J.; Sádlo, J.; Pergl, J.; Pyšek, P. Black locust (Robinia pseudoacacia) beloved and despised: A story of an invasive tree in Central Europe. For. Ecol. Manag. 2017, 384, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Kokutse, N.; Genet, M.; Fourcaud, T.; Zhang, Z. Effect of spatial variation of tree root characteristics on slope stability. A case study on Black Locust (Robinia pseudoacacia) and Arborvitae (Platycladus orientalis) stands on the Loess Plateau, China. Catena 2012, 92, 139–154. [Google Scholar] [CrossRef]
- Sands, R. Forestry in a global context. CABI 2006, 16, 633–635. [Google Scholar]
- Økland, T.; Rydgren, K.; Økland, R.H.; Storaunet, K.O.; Rolstad, J. Variation in environmental conditions, understorey species number, abundance and composition among natural and managed Picea abies forest stands. For. Ecol. Manag. 2003, 177, 17–37. [Google Scholar] [CrossRef]
- Lavoie, J.; Girona, M.M.; Morin, H. Vulnerability of Conifer Regeneration to Spruce Budworm Outbreaks in the Eastern Canadian Boreal Forest. Forests 2019, 10, 850. [Google Scholar] [CrossRef]
- Rytter, L.; Rytter, R.M. Productivity and sustainability of hybrid aspen (Populus tremula L. × P. Tremuloides Michx.) root sucker stands with varying management strategies. For. Ecol. Manag. 2017, 401, 223–232. [Google Scholar] [CrossRef]
- Ghalandarayeshi, S.; Nord-Larsen, T.; Johannsen, V.K.; Larsen, J.B. Spatial patterns of tree species in Suserup Skov – a semi-natural forest in Denmark. For. Ecol. Manag. 2017, 406, 391–401. [Google Scholar] [CrossRef]
- Nord-Larsen, T.; Bechsgaard, A.; Holm, M.; Holten-Andersen, P. Economic analysis of near-natural beech stand management in Northern Germany. For. Ecol. Manag. 2003, 184, 149–165. [Google Scholar] [CrossRef]
- Watt, A.S. Pattern and Process in the Plant Community. J. Ecol. 1947, 35, 1–22. [Google Scholar] [CrossRef]
- Chave, J. The problem of pattern and scale in ecology: What have we learned in 20 years? Ecol. Lett. 2013, 16, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Condit, R. Spatial patterns in the distribution of tropical tree species. Science 2000, 288, 1414–1418. [Google Scholar] [CrossRef] [PubMed]
- Levin, S.A. The Problem of Pattern and Scale in Ecology: The Robert H. MacArthur Award Lecture. Ecology 1992, 73, 1943–1967. [Google Scholar] [CrossRef]
- Leps, J. Can underlying mechanisms be deduced from observed patterns. Spat. Process. Plant Communities 1990, 1–11. [Google Scholar]
- Wild, J.; Kopecký, M.; Svoboda, M.; Zenáhlíková, J.; Edwards-Jonášová, M.; Herben, T. Spatial patterns with memory: Tree regeneration after stand-replacing disturbance in Picea abies mountain forests. J. Veg. Sci. 2014, 25, 1327–1340. [Google Scholar] [CrossRef]
- Getzin, S.; Wiegand, T.; Wiegand, K.; He, F. Heterogeneity influences spatial patterns and demographics in forest stands. J. Ecol. 2008, 96, 807–820. [Google Scholar] [CrossRef]
- Greig-Smith, P. Ecological Observations on Degraded and Secondary Forest in Trinidad, British West Indies: I. General Features of the Vegetation. J. Ecol. 1952, 40, 283–315. [Google Scholar] [CrossRef]
- Yang, X.; Yan, H.; Li, B.; Han, Y.; Song, B. Spatial distribution patterns of Symplocos congeners in a subtropical evergreen broad-leaf forest of southern China. J. For. Res. 2018, 29, 773–784. [Google Scholar] [CrossRef]
- Akhavan, R.; Sagheb-Talebi, K.; Zenner, E.K.; Safavimanesh, F. Spatial patterns in different forest development stages of an intact old-growth Oriental beech forest in the Caspian region of Iran. Eur. J. For. Res. 2012, 131, 1355–1366. [Google Scholar] [CrossRef]
- Dimov, L.D.; Chambers, J.L.; Lockhart, B.R. Spatial continuity of tree attributes in bottomland hardwood forests in the Southeastern United States. For. Sci. 2005, 51, 532–540. [Google Scholar]
- Salas, C.; LeMay, V.; Núñez, P.; Pacheco, P.; Espinosa, A. Spatial patterns in an old-growth Nothofagus obliqua forest in south-central Chile. For. Ecol. Manag. 2006, 231, 38–46. [Google Scholar] [CrossRef]
- Girona, M.M.; Morin, H.; Lussier, J.M.; Walsh, D. Radial growth response of black spruce stands ten years after experimental shelterwoods and seed-tree cuttings in boreal forest. Forests 2016, 7, 240. [Google Scholar] [CrossRef]
- Girona, M.M.; Rossi, S.; Lussier, J.M.; Walsh, D.; Morin, H. Understanding tree growth responses after partial cuttings: A new approach. PLoS ONE 2017, 12, e0172653. [Google Scholar]
- Gadow, K.V. The neighbourhood pattern-a new parameter for describing forest structures. Cent. Ges Forstwes. 1998, 115, 1–10. [Google Scholar]
- von Gadow, K.; Füldner, K. Zur bestandesbeschreibung in der forsteinrichtung. Forst und Holz 1993, 48, 602–606. [Google Scholar]
- Gadow, K.V.; Zhang, C.Y.; Wehenkel, C.; Pommerening, A.; Corral-Rivas, J.; Korol, M.; Myklush, S.; Hui, G.Y.; Kiviste, A.; Zhao, X.H. “Forest Structure and Diversity.” In Continuous Cover Forestry; Springer: Dordrecht, The Netherlands, 2012; pp. 29–83. [Google Scholar]
- Newton, J.; Ripley, B.D. Spatial Statistics; John Wiley & Sons: Hoboken, NJ, USA, 1984; Volume 40, ISBN 9780471725213. [Google Scholar]
- Ripley, B.D. Modelling Spatial Patterns. J. R. Stat. Soc. Ser. B 1977, 39, 172–192. [Google Scholar] [CrossRef]
- Ripley, B.D. Tests of “Randomness” for Spatial Point Patterns. J. R. Stat. Soc. Ser. B 1979, 41, 368–374. [Google Scholar] [CrossRef]
- Besag, J.E. Contribution to the discussion of Dr. Ripley’s paper. J. R. Stat. Soc. 1977, 39, 193–195. [Google Scholar]
- Baddeley, A.; Rubak, E.; Turner, R. Spatial Point Patterns: Methodology and Applications with R; Chapman and Hall/CRC: London, UK, 2015. [Google Scholar]
- Law, R.; Illian, J.; Burslem, D.F.R.P.; Gratzer, G.; Gunatilleke, C.V.S.; Gunatilleke, I.A.U.N. Ecoogical information from satial patterns of plants: Insights from point process theory. J. Ecol. 2009, 97, 616–628. [Google Scholar] [CrossRef]
- Turkington, R.; Harper, J.L. The growth, distribution and neighbour relationships of Trifolium repens in a permanent pasture: I. Ordination, pattern and contact. J. Ecol. 1979, 67, 201–218. [Google Scholar] [CrossRef]
- Wiegand, T.; Moloney, K.A. Rings, circles, and null-models for point pattern analysis in ecology. Oikos 2004, 104, 209–229. [Google Scholar] [CrossRef]
- Illian, J.; Penttinen, A.; Stoyan, H.; Stoyan, D. Statistical Analysis and Modelling of Spatial Point Patterns; John Wiley & Sons: Hoboken, NJ, USA, 2008; ISBN 9780470725160. [Google Scholar]
- Kowarik, I. Funktionen klonalen Wachstums von Bäumen bei der Brachflächen-Sukzession unter besonderer Beachtung von Robinia pseudoacacia. Verh. Ges. Ökol. 1996, 26, 173–181. [Google Scholar]
- Gyokusen, K. Spatial distribution and morphological features of root sprouts in niseakashia (Robinia pseudoacasia L.) growing under a coastal black pine forest. Bull. Kyushu Univ. For. 1991, 64, 13–28. [Google Scholar]
- Iliev, N.; Iliev, I.; Park, Y. Black Locust (Robina Pseudoacacia L.) in Bulgaria. J. Korean For. Soc. 2005, 94, 291. [Google Scholar]
- Shure, D.J.; Phillips, D.L.; Edward Bostick, P. Gap size and succession in cutover southern Appalachian forests: An 18 year study of vegetation dynamics. Plant Ecol. 2006, 185, 299–318. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, H.; Dong, B.; Zhou, M.; Ma, L.; Jia, Z.; Duan, J. Effects of canopy gap size on growth and spatial patterns of Chinese pine (Pinus tabulaeformis) regeneration. For. Ecol. Manag. 2017, 385, 46–56. [Google Scholar] [CrossRef]
- Wickham, H. Package ‘ggplot2’: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Stoyan, D.; Penttinen, A. Recent applications of point process methods in forestry statistics. Stat. Sci. 2000, 15, 61–78. [Google Scholar]
- Penttinen, A.; Stoyan, D.; Henttonen, H.M. Marked Point Processes in Forest Statistics. For. Sci. 1992, 38, 806–824. [Google Scholar]
- Haase, P. Spatial pattern analysis in ecology based on Ripley’s K-function: Introduction and methods of edge correction. J. Veg. Sci. 1995, 6, 575–582. [Google Scholar] [CrossRef]
- Loosmore, N.B.; Ford, E.D. Statistical inference using the G or K point pattern spatial statistics. Ecology 2006, 87, 1925–1931. [Google Scholar] [CrossRef]
- Zhang, H.; Xue, J. Spatial pattern and competitive relationships of moso bamboo in a native subtropical rainforest community. Forests 2018, 9, 774. [Google Scholar] [CrossRef]
- Matérn, B. Spatial Variation: Stochastic Models and their Applications to Some Problems in Forest Surveys and Other Sampling Investigations. Medd. fran Statens Skogsforskningsinstitut 1960, 49, 1–144. [Google Scholar]
- Jalilian, A.; Guan, Y.; Waagepetersen, R. Decomposition of Variance for Spatial Cox Processes. Scand. J. Stat. 2013, 40, 119–137. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Chang, L.W.; Yang, K.C.; Wang, H.H.; Sun, I.F. Point patterns of tree distribution determined by habitat heterogeneity and dispersal limitation. Oecologia 2011, 165, 175–184. [Google Scholar] [CrossRef]
- Wang, J.; Yan, Q.; Zhang, T.; Lu, D.; Xie, J.; Sun, Y.; Zhang, J.; Zhu, J. Converting larch plantations to larch-walnut mixed stands: Effects of spatial distribution pattern of larch plantations on the rodent-mediated seed dispersal of Juglans mandshurica. Forests 2018, 9, 716. [Google Scholar] [CrossRef]
- Yan, Q.L.; Zhu, J.J.; Yu, L.Z. Seed regeneration potential of canopy gaps at early formation stage in temperate secondary forests, Northeast China. PLoS ONE 2012, 7, e39502. [Google Scholar] [CrossRef]
- Zhang, X.Q.; Liu, J.; Welham, C.V.J.; Liu, C.C.; Li, D.N.; Chen, L.; Wang, R.Q. The effects of clonal integration on morphological plasticity and placement of daughter ramets in black locust (Robinia pseudoacacia). Flora Morphol. Distrib. Funct. Ecol. Plants 2006, 201, 547–554. [Google Scholar] [CrossRef]
- Alpert, P.; Mooney, H.A. Resource heterogeneity generated by shrubs and topography on coastal sand dunes. Vegetatio 1996, 122, 83–93. [Google Scholar] [CrossRef]
- Prévost, M.; Gauthier, M.M. Precommercial thinning increases growth of overstory aspen and understory balsam fir in a boreal mixedwood stand. For. Ecol. Manag. 2012, 278, 17–26. [Google Scholar] [CrossRef]
- Barrett, R.P.; Mebrahtu, T.; Hanover, J.W. Black locust: A multi-purpose tree species for temperate climates. In Advances in New Crops; Timber Press: Portland, OR, USA, 1990; pp. 278–283. [Google Scholar]
- Rebertus, A.J.; Williamson, G.B.; Moser, E.B. Fire-induced changes in Quercus laevis spatial pattern in Florida sandhills. J. Ecol. 1989, 77, 638–650. [Google Scholar] [CrossRef]
- Sterrett, J.P.; Chappell, W.E. The Effect of Auxin on Suckering in Black Locust. Weeds 1967, 15, 323–326. [Google Scholar] [CrossRef]
- Devaney, J.L.; Jansen, M.A.K.; Whelan, P.M. Spatial patterns of natural regeneration in stands of English yew (Taxus baccata L.); Negative neighbourhood effects. For. Ecol. Manag. 2014, 321, 52–60. [Google Scholar] [CrossRef]
- Dovčiak, M.; Frelich, L.E.; Reich, P.B. Discordance in spatial patterns of white pine (Pinus strobus) size-classes in a patchy near-boreal forest. J. Ecol. 2001, 89, 280–291. [Google Scholar] [CrossRef]
- He, F.; Duncan, R.P. Density-dependent effects on tree survival in an old-growth Douglas fir forest. J. Ecol. 2000, 88, 676–688. [Google Scholar] [CrossRef]
- Connell, J. On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees. Dyn. Popul. 1971, 298, 312. [Google Scholar]
- Janzen, D.H. Herbivores and the Number of Tree Species in Tropical Forests. Am. Nat. 1970, 104, 501–528. [Google Scholar] [CrossRef]
- Harrington, C.A. Factors influencing initial sprouting of red alder. Can. J. For. Res. 1984, 14, 357–361. [Google Scholar] [CrossRef]
- McIntire, E.J.B.; Fajardo, A. Beyond description: The active and effective way to infer processes from spatial patterns. Ecology 2009, 90, 46–56. [Google Scholar] [CrossRef]
- Shen, G.; Yu, M.; Hu, X.S.; Mi, X.; Ren, H.; Sun, I.F.; Ma, K. Species-area relationships explained by the joint effects of dispersal limitation and habitat heterogeneity. Ecology 2009, 90, 3033–3041. [Google Scholar] [CrossRef]
- Dessaint, F.; Chadoeuf, R.; Barralis, G. Spatial Pattern Analysis of Weed Seeds in the Cultivated Soil Seed Bank. J. Appl. Ecol. 1991, 28, 721–730. [Google Scholar] [CrossRef]
- Drössler, L.; Feldmann, E.; Glatthorn, J.; Annighöfer, P.; Kucbel, S.; Tabaku, V. What Happens after the Gap?—Size Distributions of Patches with Homogeneously Sized Trees in Natural and Managed Beech Forests in Europe. Open J. For. 2016, 6, 177. [Google Scholar] [CrossRef]
- Hubbell, S.P. Tree dispersion, abundance, and diversity in a tropical dry forest. Science 1979, 203, 1299–1309. [Google Scholar] [CrossRef] [PubMed]
- Petritan, I.C.; Marzano, R.; Petritan, A.M.; Lingua, E. Overstory succession in a mixed Quercus petraea—Fagus sylvatica old growth forest revealed through the spatial pattern of competition and mortality. For. Ecol. Manag. 2014, 326, 9–17. [Google Scholar] [CrossRef]
- Plotkin, J.B.; Chave, J.; Ashton, P.S. Cluster analysis of spatial patterns in malaysian tree species. Am. Nat. 2002, 160, 629–644. [Google Scholar] [CrossRef] [PubMed]
- Hubbell, S.P.; Foster, R.B.; O’Brien, S.T.; Harms, K.E.; Condit, R.; Wechsler, B.; Wright, S.J.; Loo De Lao, S. Light-gap disturbances, recruitment limitation, and tree diversity in a neotropical forest. Science 1999, 283, 554–557. [Google Scholar] [CrossRef]
- Veblen, T.T.; Ashton, D.H.; Schlegel, F.M. Tree Regeneration Strategies in a Lowland Nothofagus-Dominated Forest in South-Central Chile. J. Biogeogr. 1979, 6, 329–340. [Google Scholar] [CrossRef]
- Wiegand, T.; Gunatilleke, S.; Gunatilleke, N.; Okuda, T. Analyzing the spatial structure of a Sri Lankan tree species with multiple scales of clustering. Ecology 2007, 88, 3088–3102. [Google Scholar] [CrossRef]
- Sachs, T. Developmental processes and the evolution of plant clonality. In Ecology and Evolutionary Biology of Clonal Plants; Springer: Dordrecht, The Netherlands, 2002; pp. 263–278. [Google Scholar]
- Waagepetersen, R.P. An estimating function approach to inference for inhomogeneous Neyman-Scott processes. Biometrics 2007, 63, 252–258. [Google Scholar] [CrossRef]
- Barrette, M.; Bélanger, L.; De Grandpré, L.; Ruel, J.C. Cumulative effects of chronic deer browsing and clear-cutting on regeneration processes in second-growth white spruce stands. For. Ecol. Manag. 2014, 329, 69–78. [Google Scholar] [CrossRef]
- Speed, J.D.M.; Austrheim, G.; Hester, A.J.; Solberg, E.J.; Tremblay, J.P. Regional-scale alteration of clear-cut forest regeneration caused by moose browsing. For. Ecol. Manag. 2013, 289, 289–299. [Google Scholar] [CrossRef]
- Homyack, J.A.; Harrison, D.J.; Krohn, W.B. Structural differences between precommercially thinned and unthinned conifer stands. For. Ecol. Manag. 2004, 194, 131–143. [Google Scholar] [CrossRef]
- Varmola, M.; Salminen, H. Timing and intensity of precommercial thinning in Pinus sylvestris stands. Scand. J. For. Res. 2004, 19, 142–151. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot No. | Times of Clear-Cutting | Age of Stand | Recruits | Stumps | Average Diameter (cm) | Average Height (m) | Tending Felling | Grazing |
|---|---|---|---|---|---|---|---|---|
| II-4 | 1 | 4 | 3090 | 282 | 2.5 ± 1.25 | 2.88 ± 1.57 | No | No |
| II-6 | 1 | 6 | 1300 | 269 | 4.25 ± 1.75 | 4.49 ± 1.46 | Yes | Yes |
| II-9 | 1 | 9 | 625 | 228 | 5.32 ± 1.68 | 6.29 ± 1.94 | Yes | Yes |
| III-1 | 2 | 1 | 5171 | 467 | 1.34 ± 0.61 | 1.89 ± 0.92 | No | No |
| III-5 | 2 | 5 | 1135 | 478 | 3.49 ± 1.38 | 5.42 ± 1.86 | No | No |
| III-8 | 2 | 8 | 970 | 449 | 4.82 ± 1.76 | 7.35 ± 2.68 | No | No |
| Model | II-4 | II-6 | II-9 | III-1 | III-5 | III-8 | |
|---|---|---|---|---|---|---|---|
| CM | κ | 0.387 | 0.207 | 0.234 | 0.176 | 0.949 | 1.191 |
| r | 0.247 | 0.788 | 5.310 | 0.293 | 0.662 | 0.123 | |
| μ | - | - | - | - | - | - | |
| CV | κ | 0.217 | 0.188 | 0.136 | 0.157 | 0.791 | 1.106 |
| r | 0.301 | 0.475 | 4.692 | 0.196 | 0.457 | 0.075 | |
| μ | - | - | - | - | - | - | |
| M | κ | 0.266 | 0.166 | 0.117 | 0.026 | 0.584 | 1.102 |
| r | 0.441 | 0.930 | 6.158 | 3.341 | 0.921 | 0.128 | |
| μ | 2.803 | 3.045 | 3.085 | 33.828 | 1.170 | 0.496 | |
| V | κ | 0.191 | 0.142 | 0.057 | 0.023 | 0.435 | 1.010 |
| r | 0.379 | 0.592 | 6.106 | 1.972 | 0.694 | 0.086 | |
| μ | 3.910 | 3.548 | 6.352 | 38.155 | 1.573 | 0.542 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, K.; Shen, Z.; Yang, X.; Ma, L.; Duan, J.; Li, Y. Spatial Patterns in Different Stages of Regeneration after Clear-Cutting of a Black Locust Forest in Central China. Forests 2019, 10, 1066. https://doi.org/10.3390/f10121066
Zhang K, Shen Z, Yang X, Ma L, Duan J, Li Y. Spatial Patterns in Different Stages of Regeneration after Clear-Cutting of a Black Locust Forest in Central China. Forests. 2019; 10(12):1066. https://doi.org/10.3390/f10121066
Chicago/Turabian StyleZhang, Kaiquan, Zhan Shen, Xinchao Yang, Luyi Ma, Jie Duan, and Yun Li. 2019. "Spatial Patterns in Different Stages of Regeneration after Clear-Cutting of a Black Locust Forest in Central China" Forests 10, no. 12: 1066. https://doi.org/10.3390/f10121066
APA StyleZhang, K., Shen, Z., Yang, X., Ma, L., Duan, J., & Li, Y. (2019). Spatial Patterns in Different Stages of Regeneration after Clear-Cutting of a Black Locust Forest in Central China. Forests, 10(12), 1066. https://doi.org/10.3390/f10121066