Temperate Agroforestry Systems and Insect Pollinators: A Review


Abstract
1. Introduction
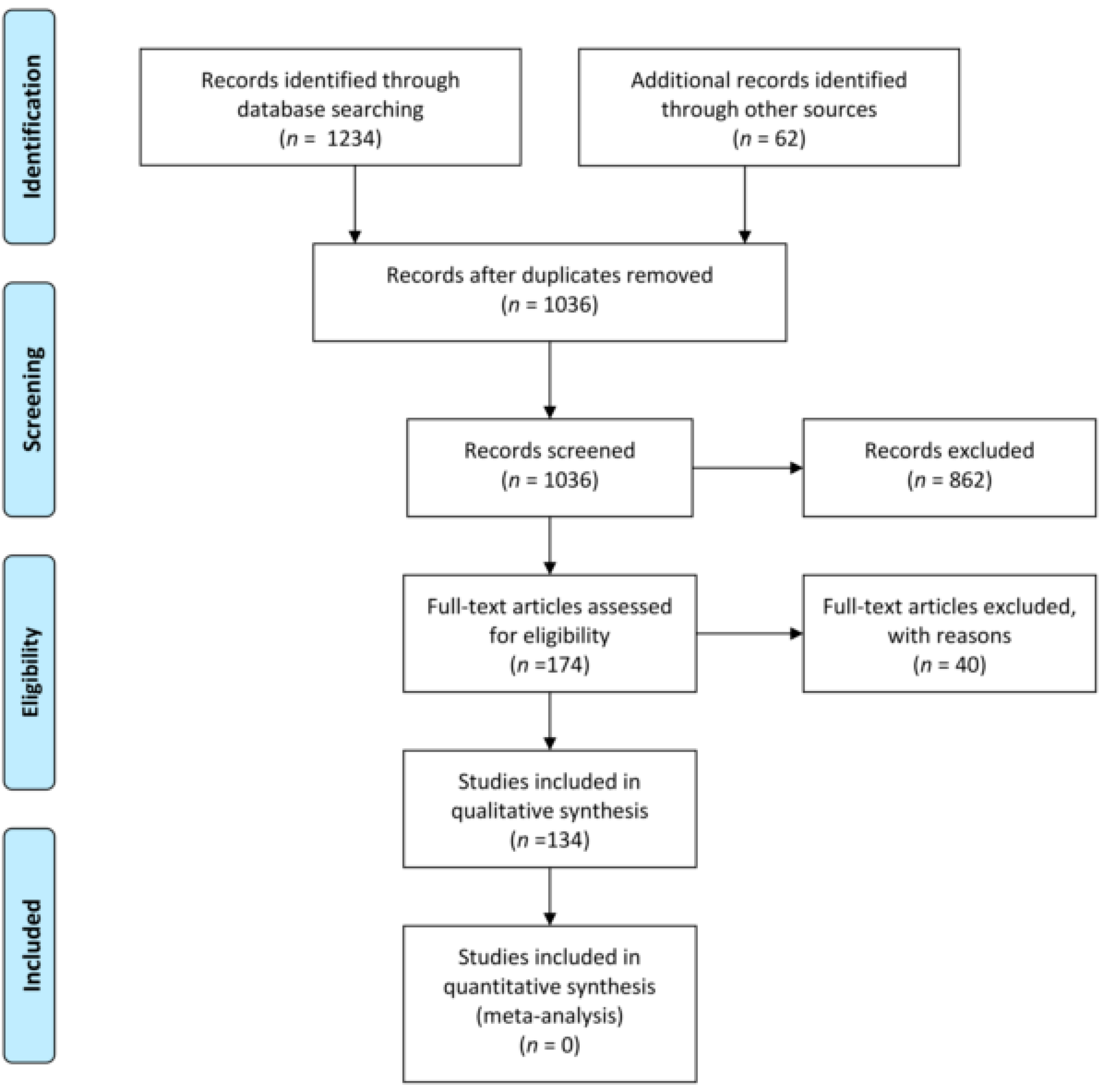
2. Literature Review
3. Foraging Resources
3.1. Pollen and Nectar
3.2. Resins and Oils
3.3. Microclimate Modification
3.4. Nesting and Egg-Laying Sites
4. Habitat Connectivity
4.1. Site Connectivity
4.2. Landscape Connectivity
4.3. Barrier
5. Pesticide Exposure Mitigation
5.1. Spray Drift Mitigation
5.2. Runoff Mitigation
5.3. Pesticide Accumulation
5.4. Refuge from Pesticides
6. Crop Pollination Services
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ollerton, J.; Winfree, R.; Tarrant, S. How many flowering plants are pollinated by animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Klein, A.M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Eilers, E.J.; Kremen, C.; Greenleaf, S.S.; Garber, A.K.; Klein, A.-M. Contribution of pollinator-mediated crops to nutrients in the human food supply. PLoS ONE 2011, 6, e21363. [Google Scholar] [CrossRef] [PubMed]
- Kearns, C.A.; Inouye, D.W. Pollinators, flowering plants, and conservation biology. BioScience 1997, 47, 297–307. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Imperatriz-Fonseca, V.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; et al. Safeguarding pollinators and their values to human well-being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef]
- McGregor, S.E. Insect Pollination of Cultivated Crop Plants, 1st ed.; United States Department of Agriculture: Washington, DC, USA, 1976.
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef]
- IPBES. Assessment Report on Pollinators, Pollination and Food Production; Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services: Bonn, Germany, 2016. [Google Scholar]
- Kremen, C.; Williams, N.M.; Thorp, R.W. Crop pollination from native bees at risk from agricultural intensification. Proc. Natl. Acad. Sci. USA 2002, 99, 16812–16816. [Google Scholar] [CrossRef]
- Williams, N.M.; Kremen, C. Resource distributions among habitats determine solitary bee offspring production in a mosaic landscape. Ecol. Appl. 2007, 17, 910–921. [Google Scholar] [CrossRef]
- Tallamy, D.W.; Shropshire, K.J. Ranking lepidopteran use of native versus introduced plants. Conserv. Biol. 2009, 23, 941–947. [Google Scholar] [CrossRef]
- Fiedler, A.K.; Landis, D.A.; Arduser, M. Rapid shift in pollinator communities following invasive species removal. Restor. Ecol. 2012, 20, 593–602. [Google Scholar] [CrossRef]
- Kevan, P.G. Pollinators as bioindicators of the state of the environment: Species, activity and diversity. Agric. Ecosyst. Environ. 1999, 74, 373–393. [Google Scholar] [CrossRef]
- Dover, J.; Sotherton, N.; Gobbett, K. Reduced pesticide inputs on cereal field margins: The effects on butterfly abundance. Ecol. Entomol. 1990, 15, 17–24. [Google Scholar] [CrossRef]
- Whitehorn, P.R.; O’Connor, S.; Wackers, F.L.; Goulson, D. Neonicotinoid pesticide reduces bumble bee colony growth and queen production. Science 2012, 336, 351–352. [Google Scholar] [CrossRef]
- Colla, S.R.; Otterstatter, M.C.; Gegear, R.J.; Thomson, J.D. Plight of the bumble bee: Pathogen spillover from commercial to wild populations. Biol. Conserv. 2006, 129, 461–467. [Google Scholar] [CrossRef]
- Cameron, S.A.; Lozier, J.D.; Strange, J.P.; Koch, J.B.; Cordes, N.; Solter, L.F.; Griswold, T.L. Patterns of widespread decline in North American bumble bees. Proc. Natl. Acad. Sci. USA 2011, 108, 662–667. [Google Scholar] [CrossRef]
- Kearns, C.A.; Inouye, D.W.; Waser, N.M. Endangered mutualisms: The conservation of plant-pollinator interactions. Annu. Rev. Ecol. Syst. 1998, 29, 83–112. [Google Scholar] [CrossRef]
- Steffan-Dewenter, I.; Westphal, C. The interplay of pollinator diversity, pollination services and landscape change. J. Appl. Ecol. 2008, 45, 737–741. [Google Scholar] [CrossRef]
- Klein, A.-M.; Brittain, C.; Hendrix, S.D.; Thorp, R.; Williams, N.; Kremen, C. Wild pollination services to California almond rely on semi-natural habitat. J. Appl. Ecol. 2012, 49, 723–732. [Google Scholar] [CrossRef]
- Morandin, L.A.; Kremen, C. Hedgerow restoration promotes pollinator populations and exports native bees to adjacent fields. Ecol. Appl. 2013, 23, 829–839. [Google Scholar] [CrossRef]
- Schoeneberger, M.M.; Bentrup, G.; Patel-Weynand, T. Agroforestry: Enhancing Resiliency in U.S. Agricultural landscapes under Changing Conditions; U.S. Department of Agriculture, U.S. Forest Service: Washington, DC, USA, 2017.
- Jose, S. Agroforestry for ecosystem services and environmental benefits: An overview. Agrofor. Syst. 2009, 76, 1–10. [Google Scholar] [CrossRef]
- Udawatta, R.P.; Rankoth, L.M.; Jose, S. Agroforestry and biodiversity. Sustainability 2019, 11, 2879. [Google Scholar] [CrossRef]
- Moher, D. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. Ann. Intern. Med. 2009, 151, 264. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, A.M.; Mc Guckin, C. The Systematic Literature Review Method: Trials and Tribulations of Electronic Database Searching at Doctoral Level; SAGE Publications: London, UK, 2016. [Google Scholar]
- Morandin, L.A.; Kremen, C. Bee preference for native versus exotic plants in restored agricultural hedgerows. Restor. Ecol. 2013, 21, 26–32. [Google Scholar] [CrossRef]
- Garratt, M.P.D.; Senapathi, D.; Coston, D.J.; Mortimer, S.R.; Potts, S.G. The benefits of hedgerows for pollinators and natural enemies depends on hedge quality and landscape context. Agric. Ecosyst. Environ. 2017, 247, 363–370. [Google Scholar] [CrossRef]
- Sutter, L.; Albrecht, M.; Jeanneret, P. Landscape greening and local creation of wildflower strips and hedgerows promote multiple ecosystem services. J. Appl. Ecol. 2018, 55, 612–620. [Google Scholar] [CrossRef]
- Minarro, M.; Prida, E. Hedgerows surrounding organic apple orchards in north-west Spain: Potential to conserve beneficial insects. Agric. For. Entomol. 2013, 15, 382–390. [Google Scholar] [CrossRef]
- Ponisio, L.C.; Gaiarsa, M.P.; Kremen, C. Opportunistic attachment assembles plant-pollinator networks. Ecol. Lett. 2017, 20, 1261–1272. [Google Scholar] [CrossRef]
- Rollin, O.; Bretagnolle, V.; Decourtye, A.; Aptel, J.; Michel, N.; Vaissiere, B.E.; Henry, M. Differences of floral resource use between honey bees and wild bees in an intensive farming system. Agric. Ecosyst. Environ. 2013, 179, 78–86. [Google Scholar] [CrossRef]
- Sardiñas, H.S.; Kremen, C. Pollination services from field-scale agricultural diversification may be context-dependent. Agric. Ecosyst. Environ. 2015, 207, 17–25. [Google Scholar] [CrossRef]
- Timberlake, T.P.; Vaughan, I.P.; Memmott, J. Phenology of farmland floral resources reveals seasonal gaps in nectar availability for bumblebees. J. Appl. Ecol. 2019, 56, 1585–1596. [Google Scholar] [CrossRef]
- Varah, A.; Jones, H.; Smith, J.; Potts, S.G. Enhanced biodiversity and pollination in UK agroforestry systems. J. Sci. Food Agric. 2013, 93, 2073–2075. [Google Scholar] [CrossRef] [PubMed]
- Cole, L.J.; Brocklehurst, S.; Robertson, D.; Harrison, W.; McCracken, D.I. Exploring the interactions between resource availability and the utilisation of semi-natural habitats by insect pollinators in an intensive agricultural landscape. Agric. Ecosyst. Environ. 2017, 246, 157–167. [Google Scholar] [CrossRef]
- Hannon, L.E.; Sisk, T.D. Hedgerows in an agri-natural landscape: Potential habitat value for native bees. Biol. Conserv. 2009, 142, 2140–2154. [Google Scholar] [CrossRef]
- Hanley, M.E.; Franco, M.; Dean, C.E.; Franklin, E.L.; Harris, H.R.; Haynes, A.G.; Rapson, S.R.; Rowse, G.; Thomas, K.C.; Waterhouse, B.R.; et al. Increased bumblebee abundance along the margins of a mass flowering crop: Evidence for pollinator spill-over. Oikos 2011, 120, 1618–1624. [Google Scholar] [CrossRef]
- Kremen, C.; M′Gonigle, L.K.; Ponisio, L.C. Pollinator community assembly tracks changes in floral resources as restored hedgerows mature in agricultural landscapes. Front. Ecol. Evol. 2018, 6, 170. [Google Scholar] [CrossRef]
- M′Gonigle, L.K.; Ponisio, L.C.; Cutler, K.; Kremen, C. Habitat restoration promotes pollinator persistence and colonization in intensively managed agriculture. Ecol. Appl. 2015, 25, 1557–1565. [Google Scholar] [CrossRef]
- Whitaker, D.; Carroll, A.; Montevecchi, W. Elevated numbers of flying insects and insectivorous birds in riparian buffer strips. Can. J. Zool. Rev. Can. Zool. 2000, 78, 740–747. [Google Scholar] [CrossRef]
- Sobczyk, D. Butterflies (Lepidoptera) of young midfield shelterbelts. Pol. J. Ecol. 2004, 52, 449–453. [Google Scholar]
- Dover, J.; Sparks, T. A review of the ecology of butterflies in British hedgerows. J. Environ. Manag. 2000, 60, 51–63. [Google Scholar] [CrossRef]
- Luppi, M.; Dondina, O.; Orioli, V.; Bani, L. Local and landscape drivers of butterfly richness and abundance in a human-dominated area. Agric. Ecosyst. Environ. 2018, 254, 138–148. [Google Scholar] [CrossRef]
- Dainese, M.; Montecchiari, S.; Sitzia, T.; Sigura, M.; Marini, L. High cover of hedgerows in the landscape supports multiple ecosystem services in Mediterranean cereal fields. J. Appl. Ecol. 2017, 54, 380–388. [Google Scholar] [CrossRef]
- Dover, J.W.; Sparks, T.H.; Greatorex-Davies, J.N. The importance of shelter for butterflies in open landscapes. J. Insect Conserv. 1997, 1, 89–97. [Google Scholar] [CrossRef]
- Meier, K.; Kuusemets, V.; Luig, J.; Mander, U. Riparian buffer zones as elements of ecological networks: Case study on Pamassius mnemosyne distribution in Estonia. Ecol. Eng. 2005, 24, 531–537. [Google Scholar] [CrossRef]
- Rosas-Ramos, N.; Banos-Picon, L.; Tormos, J.; Asis, J.D. The complementarity between ecological infrastructure types benefits natural enemies and pollinators in a Mediterranean vineyard agroecosystem. Ann. Appl. Biol. 2019, 175, 193–201. [Google Scholar] [CrossRef]
- Froidevaux, J.S.P.; Broyles, M.; Jones, G. Moth responses to sympathetic hedgerow management in temperate farmland. Agric. Ecosyst. Environ. 2019, 270, 55–64. [Google Scholar] [CrossRef]
- Merckx, T.; Feber, R.E.; Mclaughlan, C.; Bourn, N.A.D.; Parsons, M.S.; Townsend, M.C.; Riordan, P.; Macdonald, D.W. Shelter benefits less mobile moth species: The field-scale effect of hedgerow trees. Agric. Ecosyst. Environ. 2010, 138, 147–151. [Google Scholar] [CrossRef]
- Merckx, T.; Marini, L.; Feber, R.E.; Macdonald, D.W. Hedgerow trees and extended-width field margins enhance macro-moth diversity: Implications for management. J. Appl. Ecol. 2012, 49, 1396–1404. [Google Scholar] [CrossRef]
- Haenke, S.; Kovács-Hostyánszki, A.; Fründ, J.; Batáry, P.; Jauker, B.; Tscharntke, T.; Holzschuh, A. Landscape configuration of crops and hedgerows drives local syrphid fly abundance. J. Appl. Ecol. 2014, 51, 505–513. [Google Scholar] [CrossRef]
- Schirmel, J.; Albrecht, M.; Bauer, P.-M.; Sutter, L.; Pfister, S.C.; Entling, M.H. Landscape complexity promotes hoverflies across different types of semi-natural habitats in farmland. J. Appl. Ecol. 2018, 55, 1747–1758. [Google Scholar] [CrossRef]
- Macdonald, K.J.; Kelly, D.; Tylianakis, J.M. Do local landscape features affect wild pollinator abundance, diversity and community composition on Canterbury farms? N. Z. J. Ecol. 2018, 42, 262–268. [Google Scholar] [CrossRef]
- Stubbs, C.S.; Jacobson, H.A.; Osgood, E.A.; Drummond, F.A. TB148: Alternative Forage Plants for Native (Wild) Bees Associated with Lowbush Blueberry, Vaccinium spp., in Maine; Maine Agricultural & Forest Experiment Station: Orono, ME, USA, 1992. [Google Scholar]
- Loose, J.L.; Drummond, F.A.; Stubbs, C.; Woods, S. Conservation and Management of Native Bees in Cranberry; Maine Agricultural & Forest Experiment Station: Orono, ME, USA, 2005. [Google Scholar]
- Baude, M.; Kunin, W.E.; Boatman, N.D.; Conyers, S.; Davies, N.; Gillespie, M.A.K.; Morton, R.D.; Smart, S.M.; Memmott, J. Historical nectar assessment reveals the fall and rise of floral resources in Britain. Nature 2016, 530, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Somme, L.; Moquet, L.; Quinet, M.; Vanderplanck, M.; Michez, D.; Lognay, G.; Jacquemart, A.-L. Food in a row: Urban trees offer valuable floral resources to pollinating insects. Urban Ecosyst. 2016, 19, 1149–1161. [Google Scholar] [CrossRef]
- Batra, S.W.T. Red maple (Acer rubrum l.), an important early spring food resource for honey bees and other insects. J. Kans. Entomol. Soc. 1985, 58, 169–172. [Google Scholar]
- Ostaff, D.P.; Mosseler, A.; Johns, R.C.; Javorek, S.; Klymko, J.; Ascher, J.S. Willows (Salix spp.) as pollen and nectar sources for sustaining fruit and berry pollinating insects. Can. J. Plant Sci. 2015, 95, 505–516. [Google Scholar] [CrossRef]
- Crane, E.; Walker, P. Some nectar characteristics of certain important honey sources. Pszczel. Zesz. Naukowe 1985, 29, 29–45. [Google Scholar]
- Tasei, J.-N.; Aupinel, P. Nutritive value of 15 single pollens and pollen mixes tested on larvae produced by bumblebee workers (Bombus terrestris, Hymenoptera: Apidae). Apidologie 2008, 39, 397–409. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Salignon, M.; Le Conte, Y.; Belzunces, L.P.; Decourtye, A.; Kretzschmar, A.; Suchail, S.; Brunet, J.-L.; Alaux, C. Influence of pollen nutrition on honey bee health: Do pollen quality and diversity matter? PLoS ONE 2013, 8, e72016. [Google Scholar] [CrossRef]
- Russo, L.; Danforth, B. Pollen preferences among the bee species visiting apple (Malus pumila) in New York. Apidologie 2017, 48, 806–820. [Google Scholar] [CrossRef]
- Filipiak, M. Key pollen host plants provide balanced diets for wild bee larvae: A lesson for planting flower strips and hedgerows. J. Appl. Ecol. 2019, 56, 1410–1418. [Google Scholar] [CrossRef]
- Fowler, J. Specialist bees of the Northeast: Host plants and habitat conservation. Northeast. Nat. 2016, 23, 305–320. [Google Scholar] [CrossRef]
- Dötterl, S.; Vereecken, N.J. The chemical ecology and evolution of bee–flower interactions: A review and perspectives. Can. J. Zool. 2010, 88, 668–697. [Google Scholar] [CrossRef]
- Carvell, C.; Bourke, A.F.G.; Dreier, S.; Freeman, S.N.; Hulmes, S.; Jordan, W.C.; Redhead, J.W.; Sumner, S.; Wang, J.; Heard, M.S. Bumblebee family lineage survival is enhanced in high-quality landscapes. Nature 2017, 543, 547–549. [Google Scholar] [CrossRef] [PubMed]
- Wood, T.J.; Kaplan, I.; Szendrei, Z. Wild bee pollen diets reveal patterns of seasonal foraging resources for honey bees. Front. Ecol. Evol. 2018, 6, 210. [Google Scholar] [CrossRef]
- Ponisio, L.C.; M’Gonigle, L.K.; Kremen, C. On-farm habitat restoration counters biotic homogenization in intensively managed agriculture. Glob. Chang. Biol. 2016, 22, 704–715. [Google Scholar] [CrossRef]
- Donkersley, P. Trees for bees. Agric. Ecosyst. Environ. 2019, 270–271, 79–83. [Google Scholar] [CrossRef]
- Staley, J.T.; Botham, M.S.; Amy, S.R.; Hulmes, S.; Pywell, R.F. Experimental evidence for optimal hedgerow cutting regimes for Brown hairstreak butterflies. Insect Conserv. Divers. 2018, 11, 213–218. [Google Scholar] [CrossRef]
- Staley, J.T.; Botham, M.S.; Chapman, R.E.; Amy, S.R.; Heard, M.S.; Hulmes, L.; Savage, J.; Pywell, R.F. Little and late: How reduced hedgerow cutting can benefit Lepidoptera. Agric. Ecosyst. Environ. 2016, 224, 22–28. [Google Scholar] [CrossRef]
- Policarová, J.; Cardinal, S.; Martins, A.C.; Straka, J. The role of floral oils in the evolution of apid bees (Hymenoptera: Apidae). Biol. J. Linn. Soc. 2019, 128, 486–497. [Google Scholar] [CrossRef]
- Portman, Z.M.; Orr, M.C.; Griswold, T. A review and updated classification of pollen gathering behavior in bees (Hymenoptera, Apoidea). J. Hymenopt. Res. 2019, 71, 171–208. [Google Scholar] [CrossRef]
- Cane, J.H.; Griswold, T.; Parker, F.D. Substrates and materials used for nesting by North American Osmia bees (Hymenoptera: Apiformes: Megachilidae). Ann. Entomol. Soc. Am. 2007, 100, 350–358. [Google Scholar] [CrossRef]
- Wcislo, W.T.; Cane, J.H. Floral resource utilization by solitary bees (Hymenoptera: Apoidea) and exploitation of their stored foods by natural enemies. Annu. Rev. Entomol. 1996, 41, 257–286. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, M. Notes on Dianthidium Simile (Cresson) (Hymenoptera: Megachilidae) in Michigan. Gt. Lakes Entomol. 2018, 40, 1. [Google Scholar]
- Simone-Finstrom, M.; Spivak, M. Propolis and bee health: The natural history and significance of resin use by honey bees. Apidologie 2010, 41, 295–311. [Google Scholar] [CrossRef]
- Simone-Finstrom, M.; Borba, R.S.; Wilson, M.; Spivak, M. Propolis counteracts some threats to honey bee health. Insects 2017, 8, 46. [Google Scholar] [CrossRef] [PubMed]
- Greenaway, W.; Scaysbrook, T.; Whatley, F.R. The composition and plant origins of propolis: A report of work at Oxford. Bee World 1990, 71, 107–118. [Google Scholar] [CrossRef]
- Bankova, V.S.; de Castro, S.L.; Marcucci, M.C. Propolis: Recent advances in chemistry and plant origin. Apidologie 2000, 31, 3–15. [Google Scholar] [CrossRef]
- König, B. Plant sources of propolis. Bee World 1985, 66, 136–139. [Google Scholar] [CrossRef]
- Drescher, N.; Klein, A.-M.; Schmitt, T.; Leonhardt, S.D. A clue on bee glue: New insight into the sources and factors driving resin intake in honeybees (Apis mellifera). PLoS ONE 2019, 14, e0210594. [Google Scholar] [CrossRef]
- Ghisalberti, E.L. Propolis: A review. Bee World 1979, 60, 59–84. [Google Scholar] [CrossRef]
- Corbet, S.A. Pollination and the weather. Israel J. Plant Sci. 1990, 39, 13–30. [Google Scholar]
- Vicens, N.; Bosch, J. Weather-dependent pollinator activity in an apple orchard, with special reference to Osmia cornuta and Apis mellifera (Hymenoptera: Megachilidae and Apidae). Environ. Entomol. 2000, 29, 413–420. [Google Scholar] [CrossRef]
- McNaughton, K.G. Effects of windbreaks on turbulent transport and microclimate. Agric. Ecosyst. Environ. 1988, 22, 17–39. [Google Scholar] [CrossRef]
- Baldwin, C.S. The influence of field windbreaks on vegetable and specialty crops. Agric. Ecosyst. Environ. 1988, 22, 191–203. [Google Scholar] [CrossRef]
- Norton, R.L. Windbreaks: Benefits to orchard and vineyard crops. Agric. Ecosyst. Environ. 1988, 22, 205–213. [Google Scholar] [CrossRef]
- Papanikolaou, A.D.; Kühn, I.; Frenzel, M.; Schweiger, O. Semi-natural habitats mitigate the effects of temperature rise on wild bees. J. Appl. Ecol. 2017, 54, 527–536. [Google Scholar] [CrossRef]
- Hill, D.B.; Webster, T.C. Apiculture and forestry (bees and trees). Agrofor. Syst. 1995, 29, 313–320. [Google Scholar] [CrossRef]
- Haydak, M.H. Wintering of bees in Minnesota. J. Econ. Entomol. 1958, 51, 332–334. [Google Scholar] [CrossRef]
- Merrill, J.H. Value of winter protection for bees. J. Econ. Entomol. 1923, 16, 125–130. [Google Scholar] [CrossRef]
- Brittain, C.; Kremen, C.; Klein, A.-M. Biodiversity buffers pollination from changes in environmental conditions. Glob. Chang. Biol. 2013, 19, 540–547. [Google Scholar] [CrossRef]
- Lewis, T. Patterns of distribution of insects near a windbreak of tall trees. Ann. Appl. Biol. 1970, 65, 213–220. [Google Scholar] [CrossRef]
- Pinzauti, M. The influence of the wind on nectar secretion from the melon and on the flight of bees: The use of an artificial wind-break. Apidologie 1986, 17, 63–72. [Google Scholar] [CrossRef]
- Pasek, J.E. Influence of wind and windbreaks on local dispersal of insects. Agric. Ecosyst. Environ. 1988, 22–23, 539–554. [Google Scholar] [CrossRef]
- Peri, P.L.; Bloomberg, M. Windbreaks in southern Patagonia, Argentina: A review of research on growth models, windspeed reduction, and effects on crops. Agrofor. Syst. 2002, 56, 129–144. [Google Scholar] [CrossRef]
- Potts, S.G.; Vulliamy, B.; Roberts, S.; O′Toole, C.; Dafni, A.; Ne′eman, G.; Willmer, P. Role of nesting resources in organising diverse bee communities in a Mediterranean landscape. Ecol. Entomol. 2005, 30, 78–85. [Google Scholar] [CrossRef]
- Sardiñas, H.S.; Ponisio, L.C.; Kremen, C. Hedgerow presence does not enhance indicators of nest-site habitat quality or nesting rates of ground-nesting bees. Restor. Ecol. 2016, 24, 499–505. [Google Scholar] [CrossRef]
- Sardiñas, H.S.; Tom, K.; Ponisio, L.C.; Rominger, A.; Kremen, C. Sunflower (Helianthus annuus) pollination in California′s Central Valley is limited by native bee nest site location. Ecol. Appl. 2016, 26, 438–447. [Google Scholar] [CrossRef]
- Kremen, C.; M′Gonigle, L.K. Small-scale restoration in intensive agricultural landscapes supports more specialized and less mobile pollinator species. J. Appl. Ecol. 2015, 52, 602–610. [Google Scholar] [CrossRef]
- Kay, S.; Kühn, E.; Albrecht, M.; Sutter, L.; Szerencsits, E.; Herzog, F. Agroforestry can enhance foraging and nesting resources for pollinators with focus on solitary bees at the landscape scale. Agrofor. Syst. 2019, 1–9. [Google Scholar] [CrossRef]
- Shuler, R.E.; Roulston, T.H.; Farris, G.E. Farming practices influence wild pollinator populations on squash and pumpkin. J. Econ. Entomol. 2005, 98, 790–795. [Google Scholar] [CrossRef]
- Kim, J.; Williams, N.; Kremen, C. Effects of Cultivation and Proximity to Natural Habitat on Ground-nesting Native Bees in California Sunflower Fields. J. Kans. Entomol. Soc. 2006, 79, 309–320. [Google Scholar] [CrossRef]
- Svensson, B.; Lagerlöf, J.; Svensson, B.G. Habitat preferences of nest-seeking bumble bees (Hymenoptera: Apidae) in an agricultural landscape. Agric. Ecosyst. Environ. 2000, 77, 247–255. [Google Scholar] [CrossRef]
- Kells, A.R.; Goulson, D. Preferred nesting sites of bumblebee queens (Hymenoptera: Apidae) in agroecosystems in the UK. Biol. Conserv. 2003, 109, 165–174. [Google Scholar] [CrossRef]
- Osborne, J.L.; Martin, A.P.; Shortall, C.R.; Todd, A.D.; Goulson, D.; Knight, M.E.; Hale, R.J.; Sanderson, R.A. Quantifying and comparing bumblebee nest densities in gardens and countryside habitats. J. Appl. Ecol. 2008, 45, 784–792. [Google Scholar] [CrossRef]
- Lye, G.; Park, K.; Osborne, J.; Holland, J.; Goulson, D. Assessing the value of Rural Stewardship schemes for providing foraging resources and nesting habitat for bumblebee queens (Hymenoptera: Apidae). Biol. Conserv. 2009, 142, 2023–2032. [Google Scholar] [CrossRef]
- Maudsley, M. A review of the ecology and conservation of hedgerow invertebrates in Britain. J. Environ. Manag. 2000, 60, 65–76. [Google Scholar] [CrossRef]
- Pywell, R.F.; James, K.L.; Herbert, I.; Meek, W.R.; Carvell, C.; Bell, D.; Sparks, T.H. Determinants of overwintering habitat quality for beetles and spiders on arable farmland. Biol. Conserv. 2005, 123, 79–90. [Google Scholar] [CrossRef]
- Coulthard, E.; McCollin, D.; Littlemore, J. The use of hedgerows as flight paths by moths in intensive farmland landscapes. J. Insect Conserv. 2016, 20, 345–350. [Google Scholar] [CrossRef]
- Cranmer, L.; McCollin, D.; Ollerton, J. Landscape structure influences pollinator movements and directly affects plant reproductive success. Oikos 2012, 121, 562–568. [Google Scholar] [CrossRef]
- Klaus, F.; Bass, J.; Marholt, L.; Müller, B.; Klatt, B.; Kormann, U. Hedgerows have a barrier effect and channel pollinator movement in the agricultural landscape. J. Landsc. Ecol. 2015, 8, 22–31. [Google Scholar] [CrossRef]
- Graham, J.B.; Nassauer, J.I. Wild bee abundance in temperate agroforestry landscapes: Assessing effects of alley crop composition, landscape configuration, and agroforestry area. Agrofor. Syst. 2019, 93, 837–850. [Google Scholar] [CrossRef]
- Foeldesi, R.; Kovacs-Hostyanszki, A. Hoverfly (Diptera: Syrphidae) community of a cultivated arable field and the adjacent hedgerow near Debrecen, Hungary. Biologia 2014, 69, 381–388. [Google Scholar] [CrossRef]
- Rands, S.A.; Whitney, H.M. Field margins, foraging distances and their impacts on nesting pollinator success. PLoS ONE 2011, 6, e25971. [Google Scholar] [CrossRef] [PubMed]
- Walther-Hellwig, K.; Frankl, R. Foraging habitats and foraging distances of bumblebees, Bombus spp. (Hym., Apidae), in an agricultural landscape. J. Appl. Entomol. 2000, 124, 299–306. [Google Scholar] [CrossRef]
- Dover, J.; Fry, G.L.A. Experimental simulation of some visual and physical components of a hedge and the effects on butterfly behaviour in an agricultural landscape. Entomol. Exp. Appl. 2001, 100, 221–233. [Google Scholar] [CrossRef]
- Wratten, S.; Bowie, M.; Hickman, J.; Evans, A.; Sedcole, J.; Tylianakis, J. Field boundaries as barriers to movement of hover flies (Diptera: Syrphidae) in cultivated land. Oecologia 2003, 134, 605–611. [Google Scholar] [CrossRef]
- Ouin, A.; Burel, F. Influence of herbaceous elements on butterfly diversity in hedgerow agricultural landscapes. Agric. Ecosyst. Environ. 2002, 93, 45–53. [Google Scholar] [CrossRef]
- Campagne, P.; Affre, L.; Baumel, A.; Roche, P.; Tatoni, T. Fine-scale response to landscape structure in Primula vulgaris Huds.: Does hedgerow network connectedness ensure connectivity through gene flow? Popul. Ecol. 2009, 51, 209–219. [Google Scholar] [CrossRef]
- Ucar, T.; Hall, F.R.; Tew, J.E.; Hacker, J.K. Wind tunnel studies on spray deposition on leaves of tree species used for windbreaks and exposure of honey bees. Pest Manag. Sci. 2003, 59, 358–364. [Google Scholar] [CrossRef]
- Ucar, T.; Hall, F.R. Windbreaks as a pesticide drift mitigation strategy: A review. Pest Manag. Sci. 2001, 57, 663–675. [Google Scholar] [CrossRef]
- Otto, S.; Loddo, D.; Baldoin, C.; Zanin, G. Spray drift reduction techniques for vineyards in fragmented landscapes. J. Environ. Manag. 2015, 162, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Otto, S.; Lazzaro, L.; Finizio, A.; Zanin, G. Estimating ecotoxicological effects of pesticide drift on nontarget arthropods in field hedgerows. Environ. Toxicol. Chem. 2009, 28, 853–863. [Google Scholar] [CrossRef] [PubMed]
- Lazzaro, L.; Otto, S.; Zanin, G. Role of hedgerows in intercepting spray drift: Evaluation and modelling of the effects. Agric. Ecosyst. Environ. 2008, 123, 317–327. [Google Scholar] [CrossRef]
- Longley, M.; Čilgi, T.; Jepson, P.C.; Sotherton, N.W. Measurements of pesticide spray drift deposition into field boundaries and hedgerows: 1. Summer applications. Environ. Toxicol. Chem. 1997, 16, 165–172. [Google Scholar] [CrossRef]
- Wenneker, M.; Heijne, B.; van de Zande, J.C. Effect of natural windbreaks on drift reduction in orchard spraying. Commun. Agric. Appl. Biol. Sci. 2005, 70, 961–969. [Google Scholar]
- Chen, L.; Liu, C.; Zhang, L.; Zou, R.; Zhang, Z. Variation in tree species ability to capture and retain airborne fine particulate matter (pm 2.5). Sci. Rep. 2017, 7, 1–11. [Google Scholar]
- Mercer, G.N. Modelling to determine the optimal porosity of shelterbelts for the capture of agricultural spray drift. Environ. Model. Softw. 2009, 24, 1349–1352. [Google Scholar] [CrossRef]
- Pavlidis, G. Pollution control by agroforestry systems: A short review. Eur. Water 2017, 59, 297–301. [Google Scholar]
- Pavlidis, G.; Tsihrintzis, V.A. Environmental benefits and control of pollution to surface water and groundwater by agroforestry systems: A review. Water Resour. Manag. 2018, 32, 1–29. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, X.; Zhang, M.; Dahlgren, R.A.; Eitzel, M. A review of vegetated buffers and a meta-analysis of their mitigation efficacy in reducing nonpoint source pollution. J. Environ. Qual. 2010, 39, 76–84. [Google Scholar] [CrossRef]
- Reichenberger, S.; Bach, M.; Skitschak, A.; Frede, H.-G. Mitigation strategies to reduce pesticide inputs into ground- and surface water and their effectiveness; A review. Sci. Total Environ. 2007, 384, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, Q.; Blom-Zandstra, M.; Gupta, S.; Joner, E.J. Utilising the synergy between plants and rhizosphere microorganisms to enhance breakdown of organic pollutants in the environment. Environ. Sci. Pollut. Res. Int. 2005, 12, 34–48. [Google Scholar] [CrossRef] [PubMed]
- Longley, M.; Sotherton, N.W. Measurements of pesticide spray drift deposition into field boundaries and hedgerows: 2. Autumn applications. Environ. Toxicol. Chem. 1997, 16, 173–178. [Google Scholar] [CrossRef]
- Kjær, C.; Bruus, M.; Bossi, R.; Løfstrøm, P.; Andersen, H.V.; Nuyttens, D.; Larsen, S.E. Pesticide drift deposition in hedgerows from multiple spray swaths. J. Pestic. Sci. 2014, 39, 14–21. [Google Scholar]
- Davis, B.N.K.; Williams, C.T. Buffer zone widths for honeybees from ground and aerial spraying of insecticides. Environ. Pollut. 1990, 63, 247–259. [Google Scholar] [CrossRef]
- Felsot, A.S.; Unsworth, J.B.; Linders, J.B.H.J.; Roberts, G.; Rautman, D.; Harris, C.; Carazo, E. Agrochemical spray drift; assessment and mitigation-a review. J. Environ. Sci. Health Part. B Pestic. Food Contam. Agric. Wastes 2011, 46, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Botías, C.; David, A.; Hill, E.M.; Goulson, D. Contamination of wild plants near neonicotinoid seed-treated crops, and implications for non-target insects. Sci. Total Environ. 2016, 566, 269–278. [Google Scholar] [CrossRef]
- Long, E.Y.; Krupke, C.H. Non-cultivated plants present a season-long route of pesticide exposure for honey bees. Nat. Commun. 2016, 7, 1–12. [Google Scholar] [CrossRef]
- Zaady, E.; Katra, I.; Shuker, S.; Knoll, Y.; Shlomo, S. Tree belts for decreasing aeolian dust-carried pesticides from cultivated areas. Geosciences 2018, 8, 286. [Google Scholar] [CrossRef]
- Krupke, C.H.; Hunt, G.J.; Eitzer, B.D.; Andino, G.; Given, K. Multiple routes of pesticide exposure for honey bees living near agricultural fields. PLoS ONE 2012, 7, e29268. [Google Scholar] [CrossRef]
- Gilchrist, A.; Barker, A.; Handley, J.F. Pathways through the landscape in a changing climate: The role of landscape structure in facilitating species range expansion through an urbanised region. Landsc. Res. 2016, 41, 26–44. [Google Scholar] [CrossRef]
- Krosby, M.; Tewksbury, J.; Haddad, N.M.; Hoekstra, J. Ecological connectivity for a changing climate. Conserv. Biol. 2010, 24, 1686–1689. [Google Scholar] [CrossRef] [PubMed]
- Morandin, L.A.; Long, R.F.; Kremen, C. Pest control and pollination cost-benefit analysis of hedgerow restoration in a simplified agricultural landscape. J. Econ. Entomol. 2016, 109, 1020–1027. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.D.; Lewis, T. The effects of windbreaks on the blossom-visiting fauna of apple orchards and on yield. Ann. Appl. Biol. 1972, 72, 229–238. [Google Scholar] [CrossRef]
- Castle, D.; Grass, I.; Westphal, C. Fruit quantity and quality of strawberries benefit from enhanced pollinator abundance at hedgerows in agricultural landscapes. Agric. Ecosyst. Environ. 2019, 275, 14–22. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Kremen, C.; Morales, J.M.; Bommarco, R.; Cunningham, S.A.; Carvalheiro, L.G.; Chacoff, N.P.; Dudenhöffer, J.H.; Greenleaf, S.S.; et al. Stability of pollination services decreases with isolation from natural areas despite honey bee visits. Ecol. Lett. 2011, 14, 1062–1072. [Google Scholar] [CrossRef]
- Kort, J. Benefits of windbreaks to field and forage crops. Agric. Ecosyst. Environ. 1988, 22, 165–190. [Google Scholar] [CrossRef]
- Ticknor, K.A. Design and use of field windbreaks in wind erosion control systems. Agric. Ecosyst. Environ. 1988, 22, 123–132. [Google Scholar] [CrossRef]
- Gathmann, A.; Tscharntke, T. Foraging ranges of solitary bees. J. Anim. Ecol. 2002, 71, 757–764. [Google Scholar] [CrossRef]
- Moisan-DeSerres, J.; Chagnon, M.; Fournier, V. Influence of windbreaks and forest borders on abundance and species richness of native pollinators in lowbush blueberry fields in Québec, Canada. Can. Entomol. 2015, 147, 432–442. [Google Scholar] [CrossRef]
- Ponisio, L.C.; de Valpine, P.; M′Gonigle, L.K.; Kremen, C. Proximity of restored hedgerows interacts with local floral diversity and species′ traits to shape long-term pollinator metacommunity dynamics. Ecol. Lett. 2019, 22, 1048–1060. [Google Scholar] [CrossRef] [PubMed]
- Diekoetter, T.; Billeter, R.; Crist, T.O. Effects of landscape connectivity on the spatial distribution of insect diversity in agricultural mosaic landscapes. Basic Appl. Ecol. 2008, 9, 298–307. [Google Scholar] [CrossRef]
- Krewenka, K.M.; Holzschuh, A.; Tscharntke, T.; Dormann, C.F. Landscape elements as potential barriers and corridors for bees, wasps and parasitoids. Biol. Conserv. 2011, 144, 1816–1825. [Google Scholar] [CrossRef]
- Stanley, J.; Preetha, G. Pesticide Toxicity to Non-Target Organisms: Exposure, Toxicity and Risk Assessment Methodologies; Springer: Dordrecht, The Netherlands, 2016. [Google Scholar]
- Bischof, M.; Coffey, T.; Drennan, M. The effectiveness of riparian hedgerows at intercepting drift from aerial pesticide application. J. Environ. Qual. 2019, 48, 1481–1488. [Google Scholar]
- Davis, B.N.K.; Brown, M.J.; Frost, A.J.; Yates, T.J.; Plant, R.A. The effects of hedges on spray deposition and on the biological impact of pesticide spray drift. Ecotoxicol. Environ. Saf. 1994, 27, 281–293. [Google Scholar] [CrossRef]
- Park, M.G.; Blitzer, E.J.; Gibbs, J.; Losey, J.E.; Danforth, B.N. Negative effects of pesticides on wild bee communities can be buffered by landscape context. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150299. [Google Scholar] [CrossRef]
- Venturini, E.M.; Drummond, F.A.; Hoshide, A.K.; Dibble, A.C.; Stack, L.B. Pollination reservoirs for wild bee habitat enhancement in cropping systems: A review. Agroecol. Sustain. Food Syst. 2017, 41, 101–142. [Google Scholar] [CrossRef]
- Kremen, C.; Williams, N.M.; Bugg, R.L.; Fay, J.P.; Thorp, R.W. The area requirements of an ecosystem service: Crop pollination by native bee communities in California. Ecol. Lett. 2004, 7, 1109–1119. [Google Scholar] [CrossRef]
- Nair, P.R. The coming of age of agroforestry. J. Sci. Food Agric. 2007, 87, 1613–1619. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
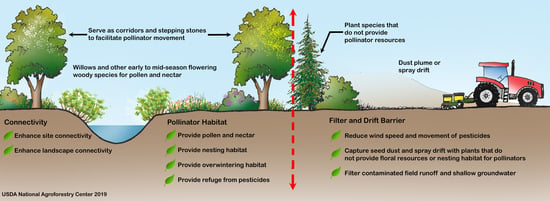
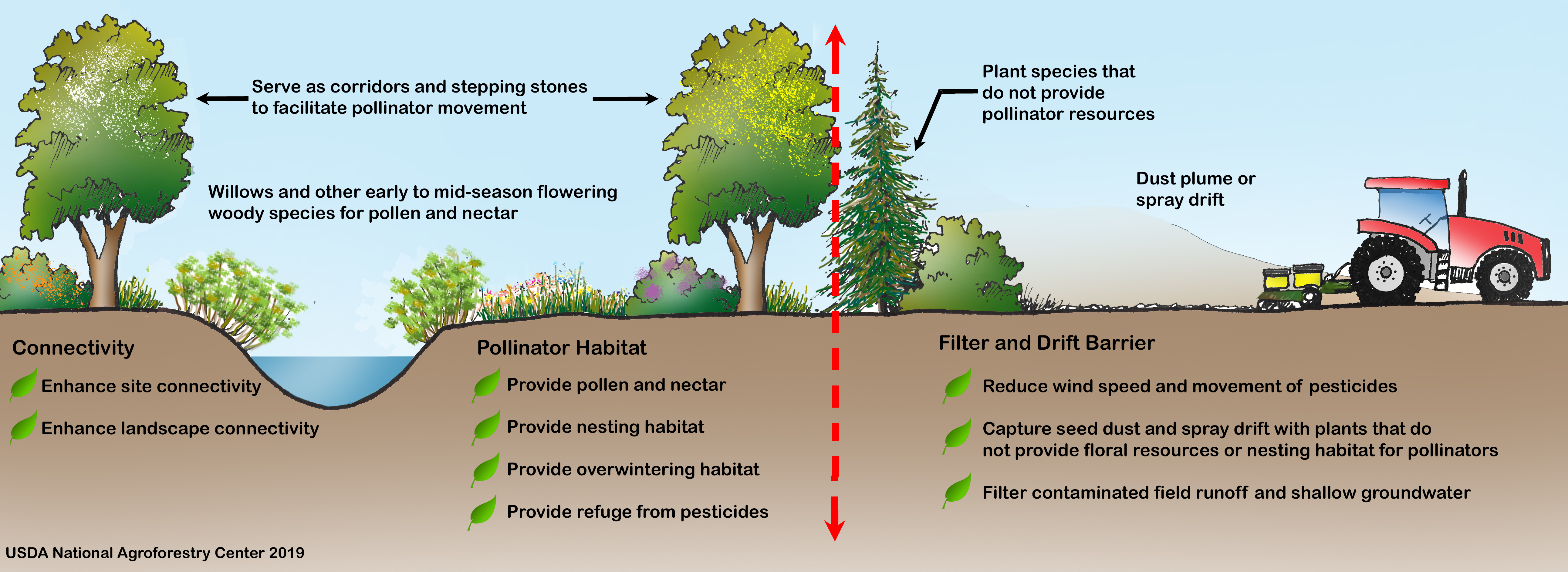
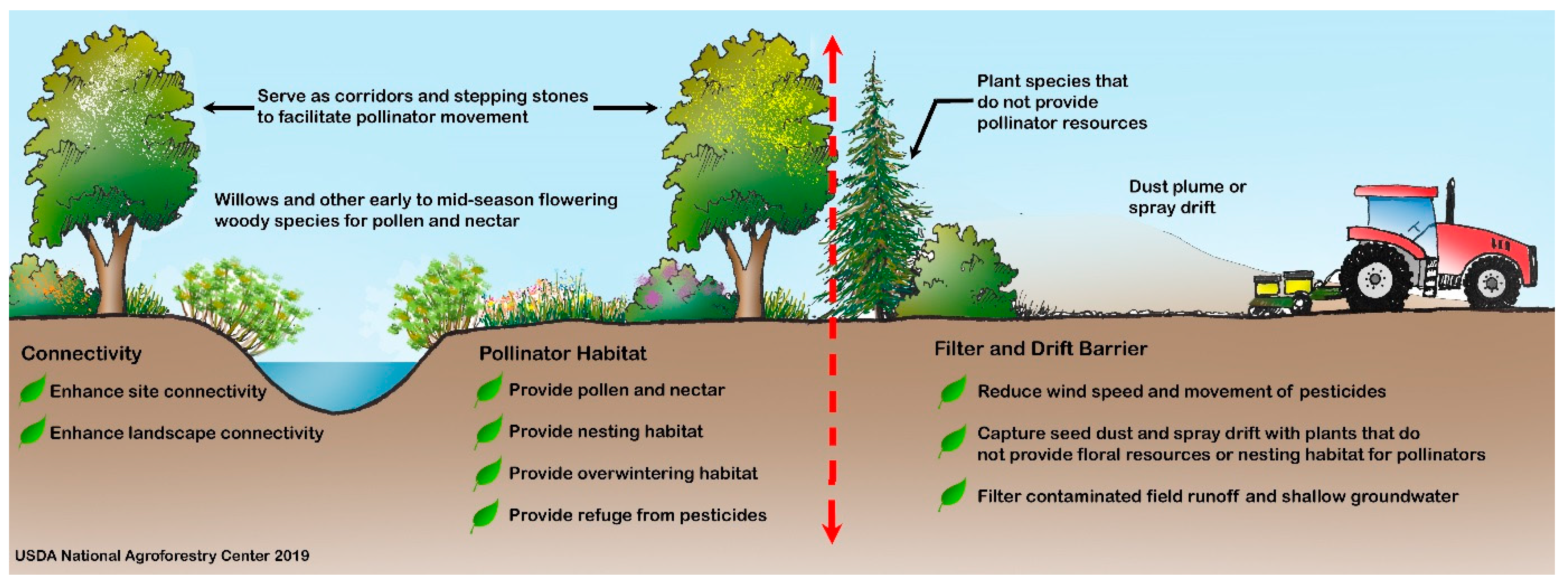
| Habitat Component or Ecosystem Service | Summary | Key References |
|---|---|---|
| Resins and oils | Honey bees harvest resins from tree buds, particularly poplar (Populus spp.), to make propolis, which provides antimicrobial and structural benefits for the colony. Other tree species, including pine (Pinus spp.), birch (Betula spp.), elm (Ulmus spp.), alder (Alnus spp.), beech (Fagus spp.), and horsechestnut (Aesculus spp.) are sources of resin. Some solitary bees also collect plant resins to include in brood cell linings and others use oils in brood cell provisions. | [75,76,78,82,83,84,86] |
| Early-season pollen and nectar | Woody species in temperate regions can provide important early-season sources of pollen and nectar. | [57,59,60,61,65,69,70] |
| Pollen protein quality | Willows (Salix spp.), cherry, and plum (Prunus spp.), and other woody species, offer pollen with high nutritive value. | [60,61,63,64,65] |
| Nectar sugar density | Tree and shrub flowers can provide nectar with relatively high sugar content and high flower densities. Hedgerows can provide greater nectar per unit area compared to woodlands and pastures. | [35,56,57,58,59,60,61,62] |
| Butterfly and moth larval hosts | Woody plants are important host plants for the larvae of many lepidopteran species (moths and butterflies). Some of the most highly used plant genera by lepidopteran species include oaks (Quercus spp.), cherry and plum (Prunus spp.), willows (Salix spp.), birch (Betula spp.), and poplar (Populus spp.). | [12,44,52,112] |
| Ground-nesting | Agroforestry practices can offer stable sites for ground-nesting bees and wasps in frequently disturbed agricultural landscapes. | [22,32,71,104,108,109,110] |
| Cavity-nesting | Shrub species with pithy centers such as elderberry (Sambucus spp.), sumac (Rhus spp), and brambles (Rubus spp.) can provide hollow tunnels for above-ground cavity-nesting bees. Dead trees and branches left in an agroforestry practice can also provide nesting sites. | [22,104,105] |
| Overwintering | Lepidopteran (butterflies and moths), Coleopteran (beetles), and other pollinators overwinter under bark and leaf litter found in hedgerows and other woody plantings. | [44,112,113] |
| Microclimate modification: wind | Windbreaks and other agroforestry practices can reduce winds and desiccation of pollen and floral parts, thereby enhancing pollinator foraging. Windbreaks can protect insect flight up to a distance equal to about 9 times its height. Agroforestry practices can help reduce winter mortality in honey bee hives by providing protection from winter winds. | [94,95,98,99] |
| Microclimate modification: temperature | Trees and shrubs can shade honey bee hives and reduce summer temperatures. Daytime air temperatures are several degrees warmer within a certain distance downwind of windbreaks and these elevated temperatures can increase pollinator activity and pollination if air temperatures at pollination time are below optimum. | [89,91,93] |
| Connectivity | Hedgerows and other linear agroforestry practices can facilitate pollinator movement across agricultural and urban landscapes at multiple spatial scales. These practices can provide spatially-distributed habitat that is within the foraging range of many pollinators, including short-distance foragers. | [22,53,104,114,115,116,117,118,119,120] |
| Barrier | Hedgerows may act as a barrier to pollinator dispersal and pollen transfer, depending on the landscape context and pollinator species. The orientation of plant rows may influence whether a hedgerow functions as a barrier or corridor. | [116,121,122,123,124] |
| Pesticide spray drift mitigation | Agroforestry practices can reduce pesticide exposure to pollinators by reducing spray drift from coming onto or leaving a farm by capturing particles and reducing wind speed. Windbreaks can reduce drift by up to 80% to 90%. Agroforestry buffers that are 2.5–3 m tall, with 40-50% porosity and fine, evergreen foliage are generally the most effective for drift prevention. | [125,126,127,128,129,130,131,132,133] |
| Pesticide runoff mitigation | Agroforestry practices can reduce pesticide exposure to pollinators by helping to capture pesticide runoff, prevent or slow pesticide movement through soil, and help break down some pesticides. | [134,135,136,137,138] |
| Refuge from pesticides | Agroforestry practices may serve as a safe haven for pollinators from pesticides, if adequately protected from spray drift. No-spray buffer zones may be necessary to protect the agroforestry planting. | [126,130,139,140,141,142] |
| Pesticide accumulation | Plants used in agroforestry practices can become contaminated with pesticides through aerial deposition and uptake through root systems. Plants contaminated by neonicotinoids through non-target drift of treated seed-coating dust during crop planting can negatively impact pollinators. | [143,144,145,146] |
| Adaptation to climate change | Agroforestry practices may offer ecological niches that allow pollinators to find suitable sites for thermal regulation under increasing temperatures and may serve as corridors and stepping stones to facilitate pollinator range shifts due to climate change. Landscapes that have a higher proportion of semi-natural habitats, including hedgerows and other woody plantings, may decrease the detrimental effects of warmer temperatures on pollinators. | [92,147,148] |
| Crop pollination | Agroforestry practices can provide increased pollination services and crop yields, including higher crop quality, although few studies have been conducted to document this direct agronomic benefit. | [30,46,149,150,151] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bentrup, G.; Hopwood, J.; Adamson, N.L.; Vaughan, M. Temperate Agroforestry Systems and Insect Pollinators: A Review. Forests 2019, 10, 981. https://doi.org/10.3390/f10110981
Bentrup G, Hopwood J, Adamson NL, Vaughan M. Temperate Agroforestry Systems and Insect Pollinators: A Review. Forests. 2019; 10(11):981. https://doi.org/10.3390/f10110981
Chicago/Turabian StyleBentrup, Gary, Jennifer Hopwood, Nancy Lee Adamson, and Mace Vaughan. 2019. "Temperate Agroforestry Systems and Insect Pollinators: A Review" Forests 10, no. 11: 981. https://doi.org/10.3390/f10110981
APA StyleBentrup, G., Hopwood, J., Adamson, N. L., & Vaughan, M. (2019). Temperate Agroforestry Systems and Insect Pollinators: A Review. Forests, 10(11), 981. https://doi.org/10.3390/f10110981