

Automatic Segmentation of Histological Images of Mouse Brains


 ,
, 
Abstract
:1. Introduction
Objective
- Management of high resolution histological images (up to 1.5 Gb each).
- Image treatment (up/down-scaling, re-sampling, curve approximation) and conversion (ROI to PNG formats and vice versa) for initial and final steps of the training pipeline.
- Training and test U-Net and Attention U-Net architectures with two different size of the images as input for the segmentation task.
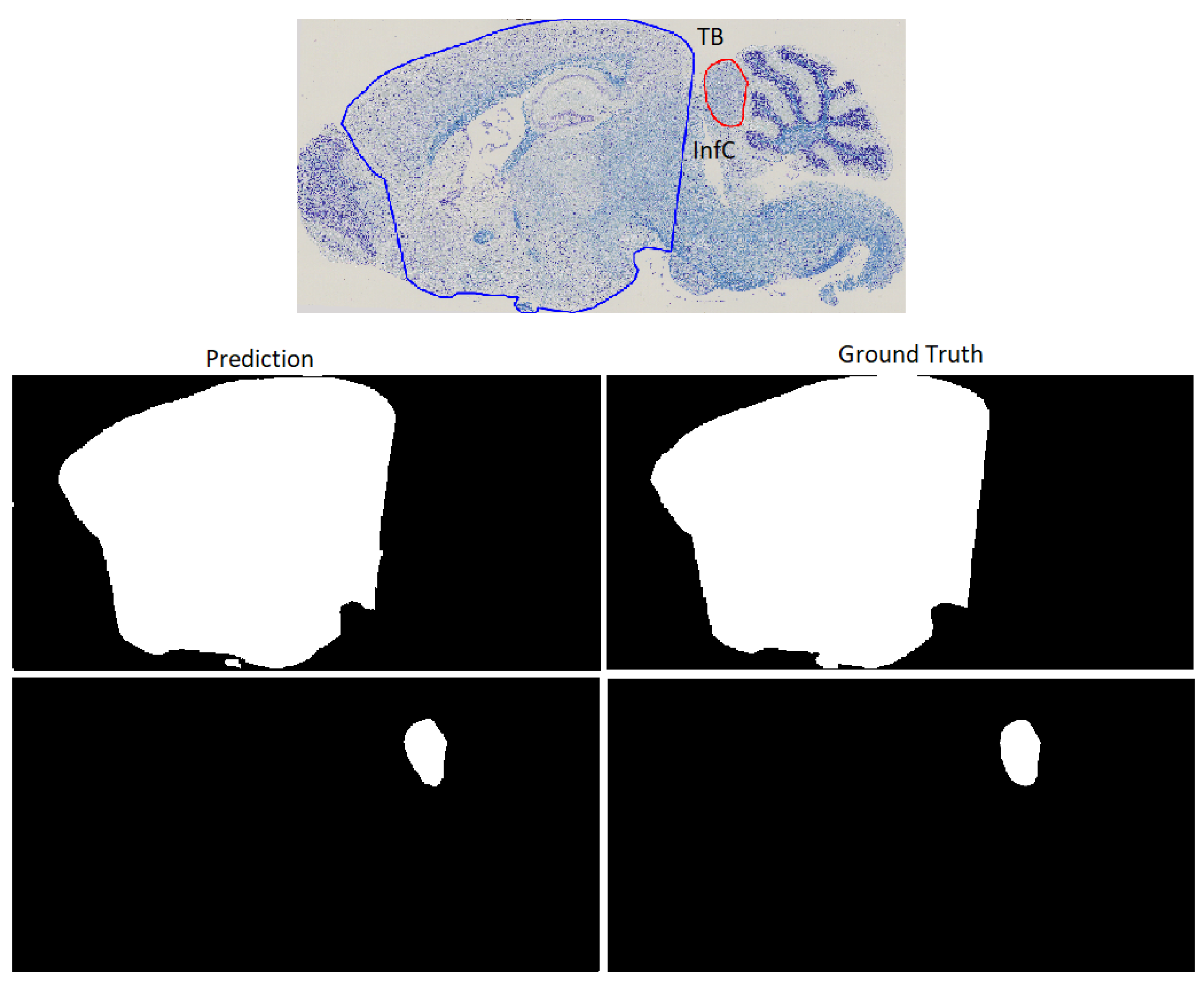
- Creation and deploy of a usable tool to automatically segment histological mouse brain images for 24 regions of interest, which would be significantly faster than human annotators.
2. Materials and Methods
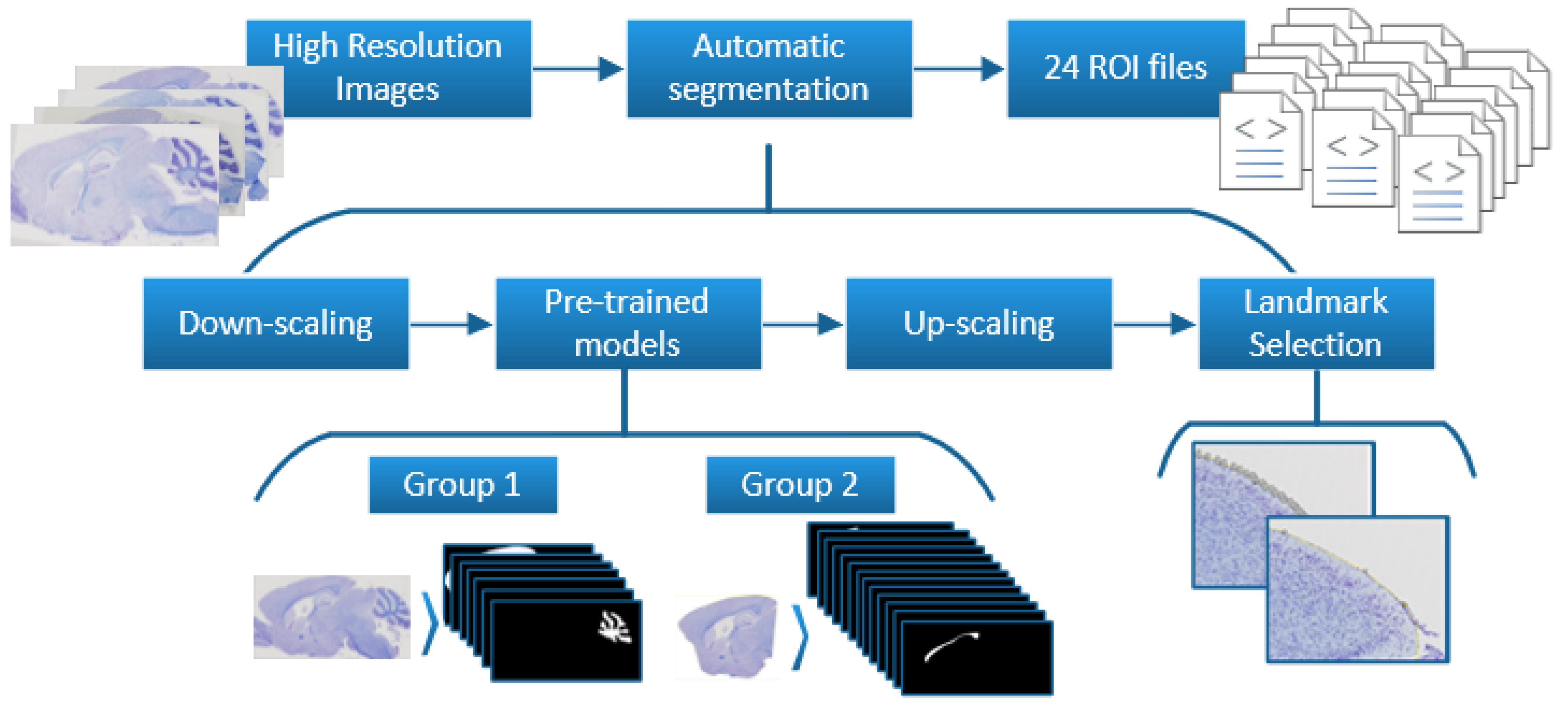
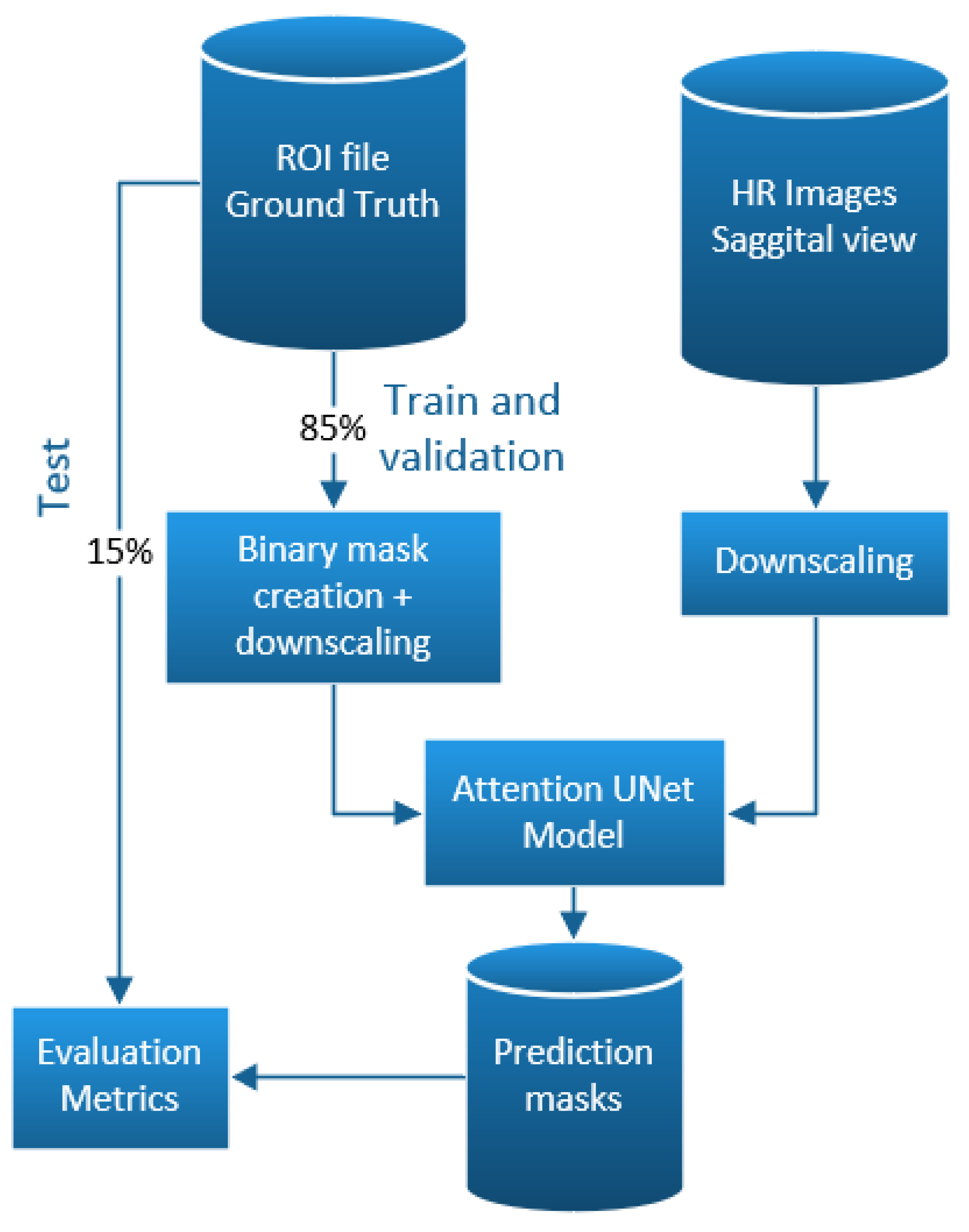
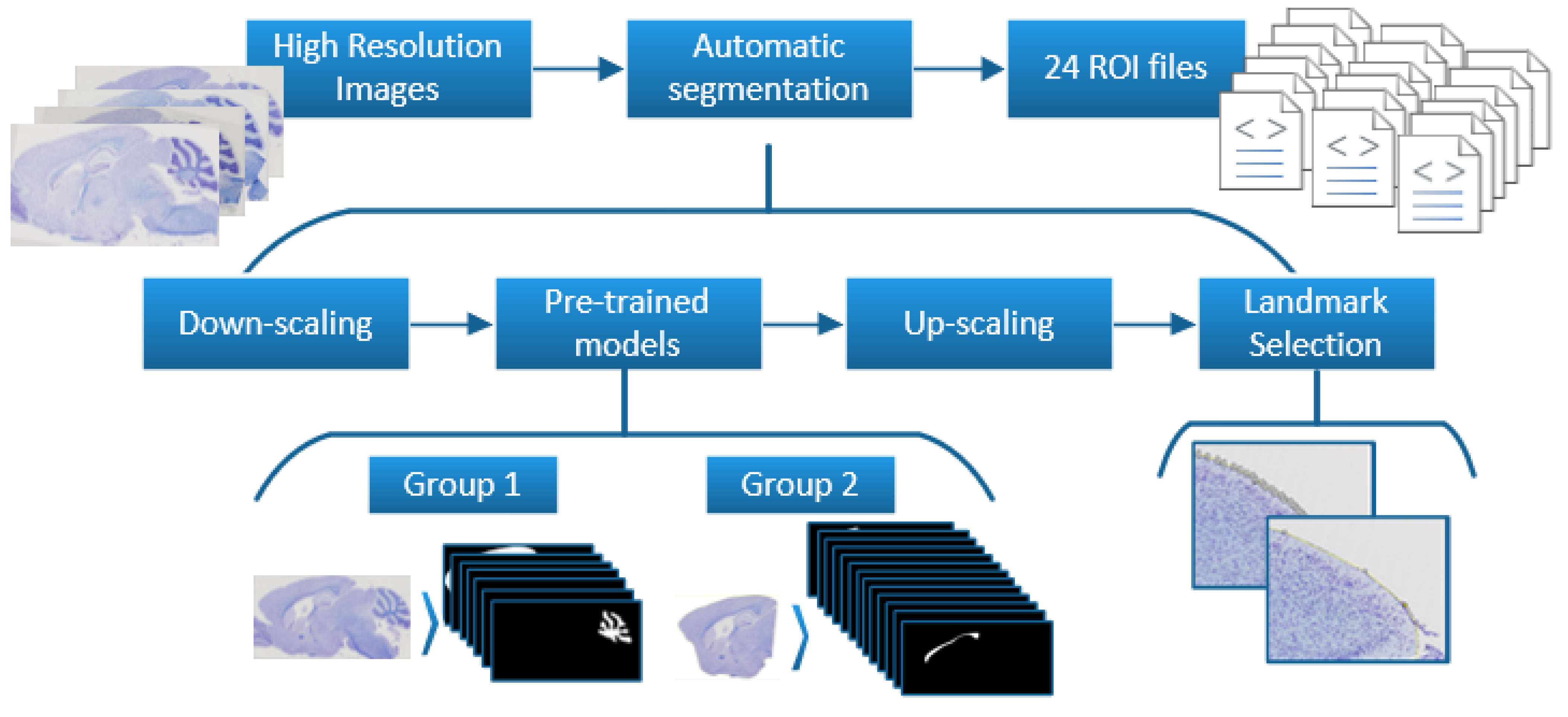
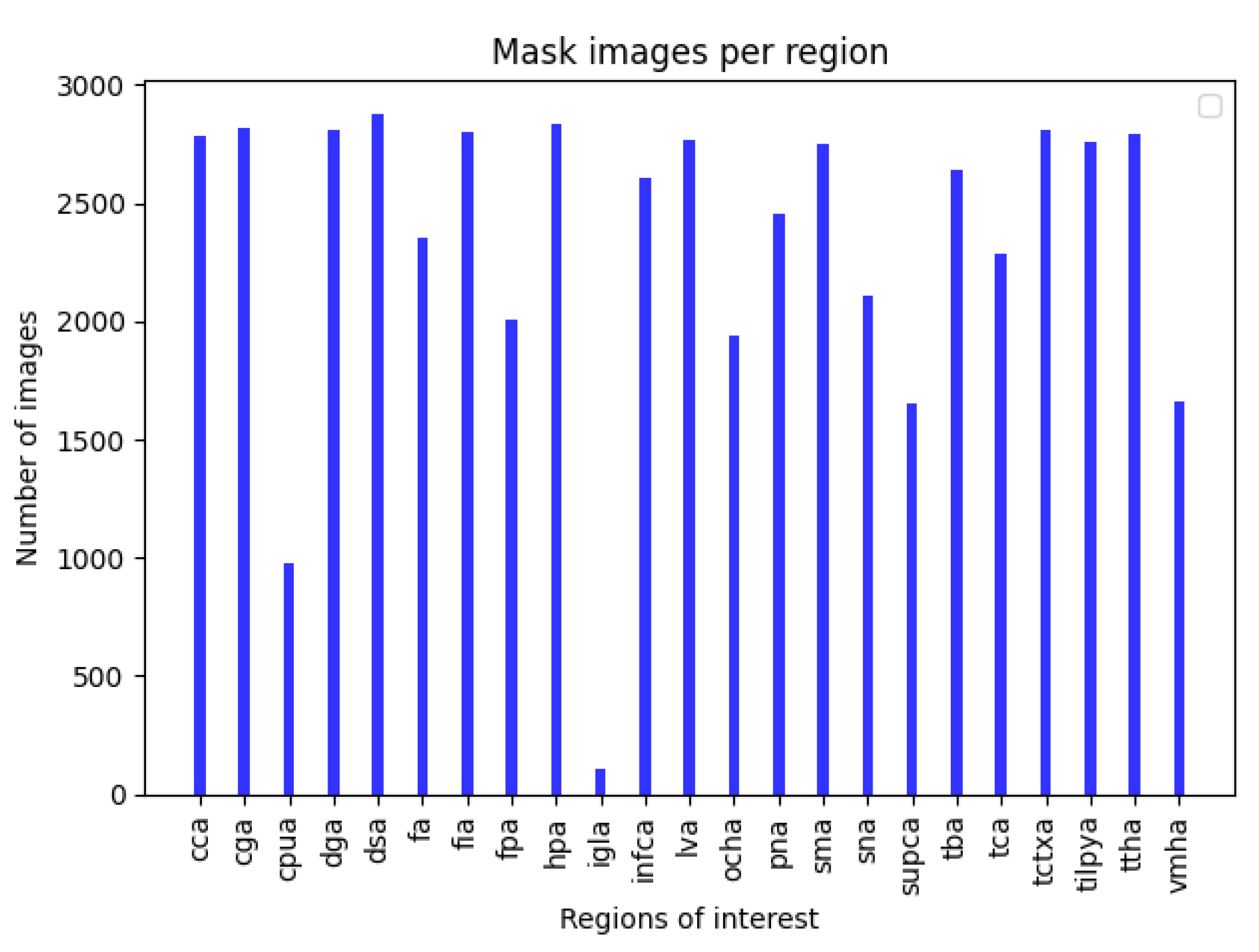
2.1. Dataset Preparation
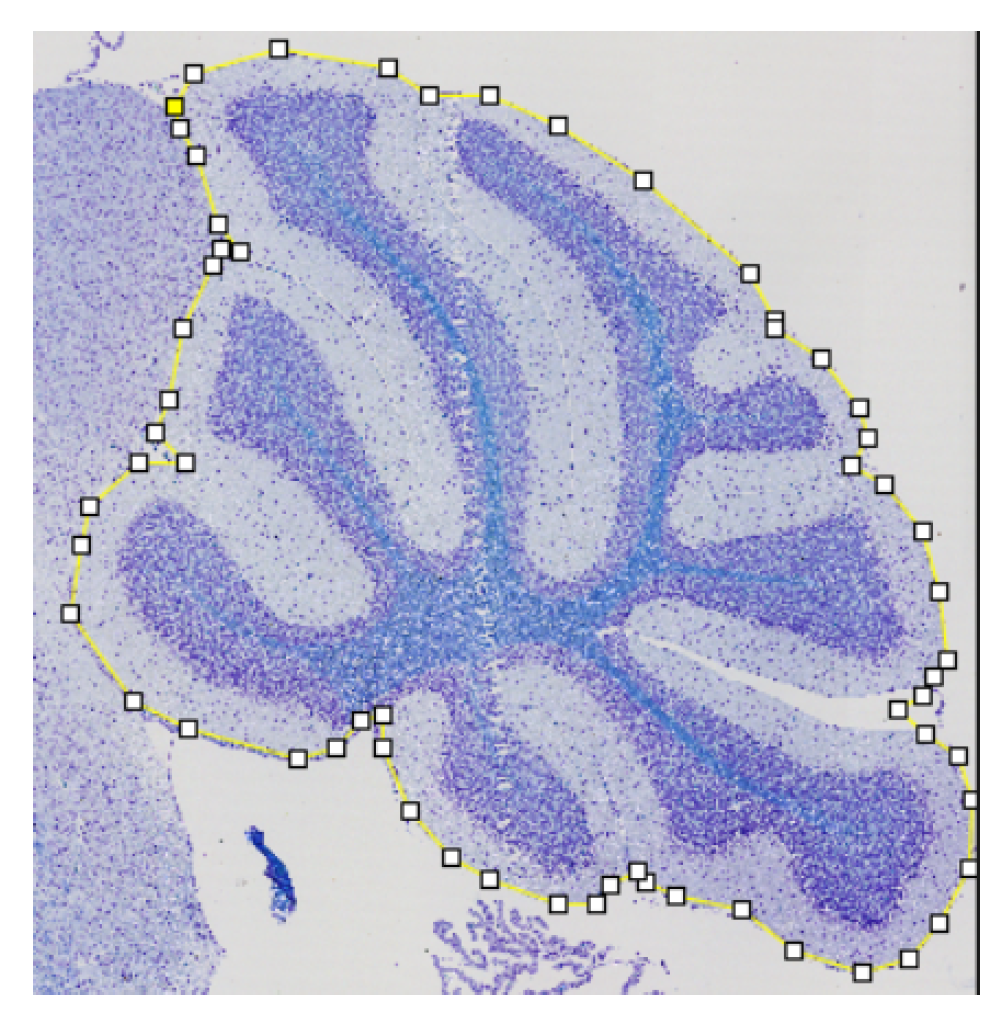
2.1.1. Landmarks to Binary Masks
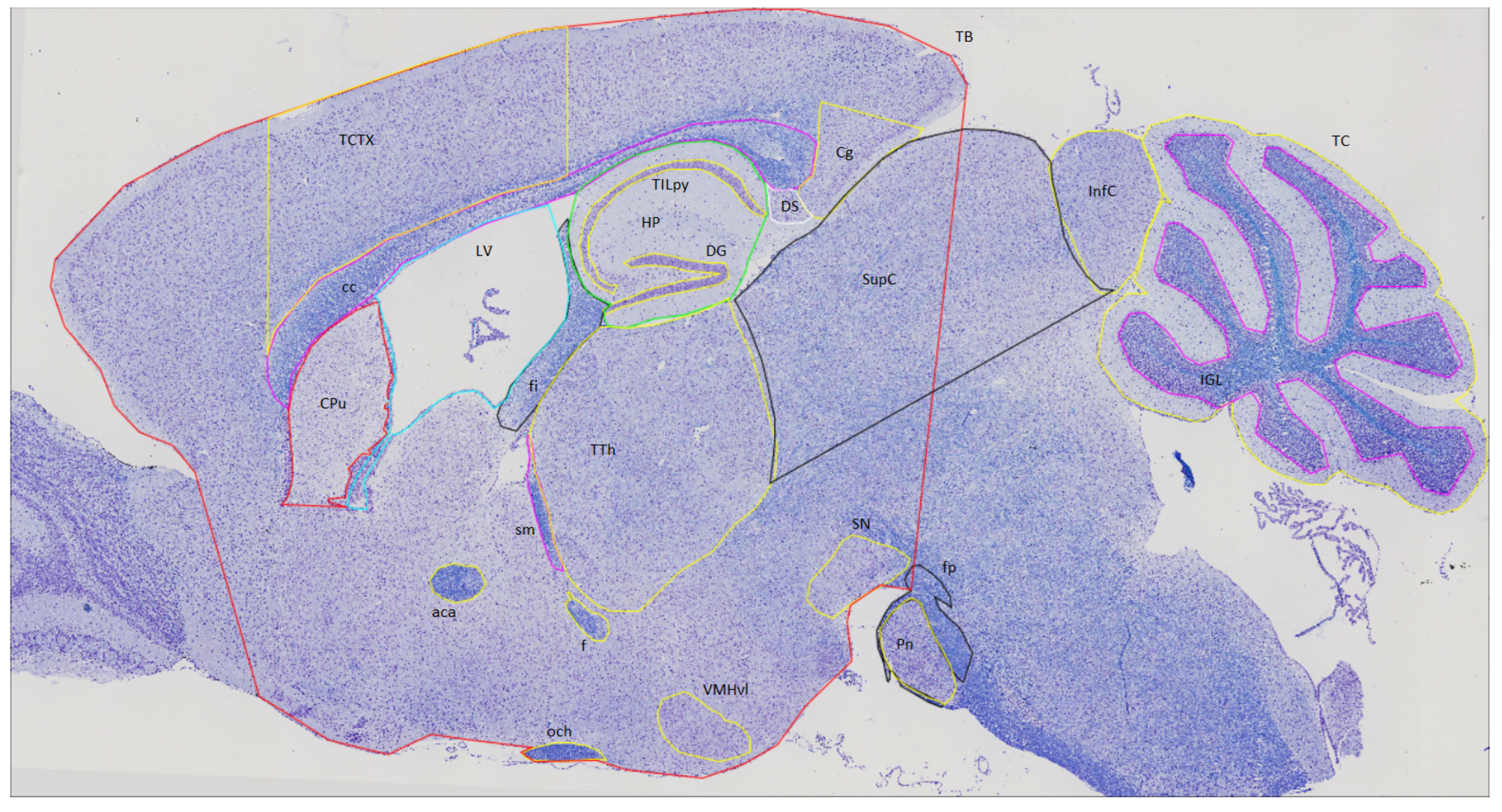
2.1.2. Brain Division
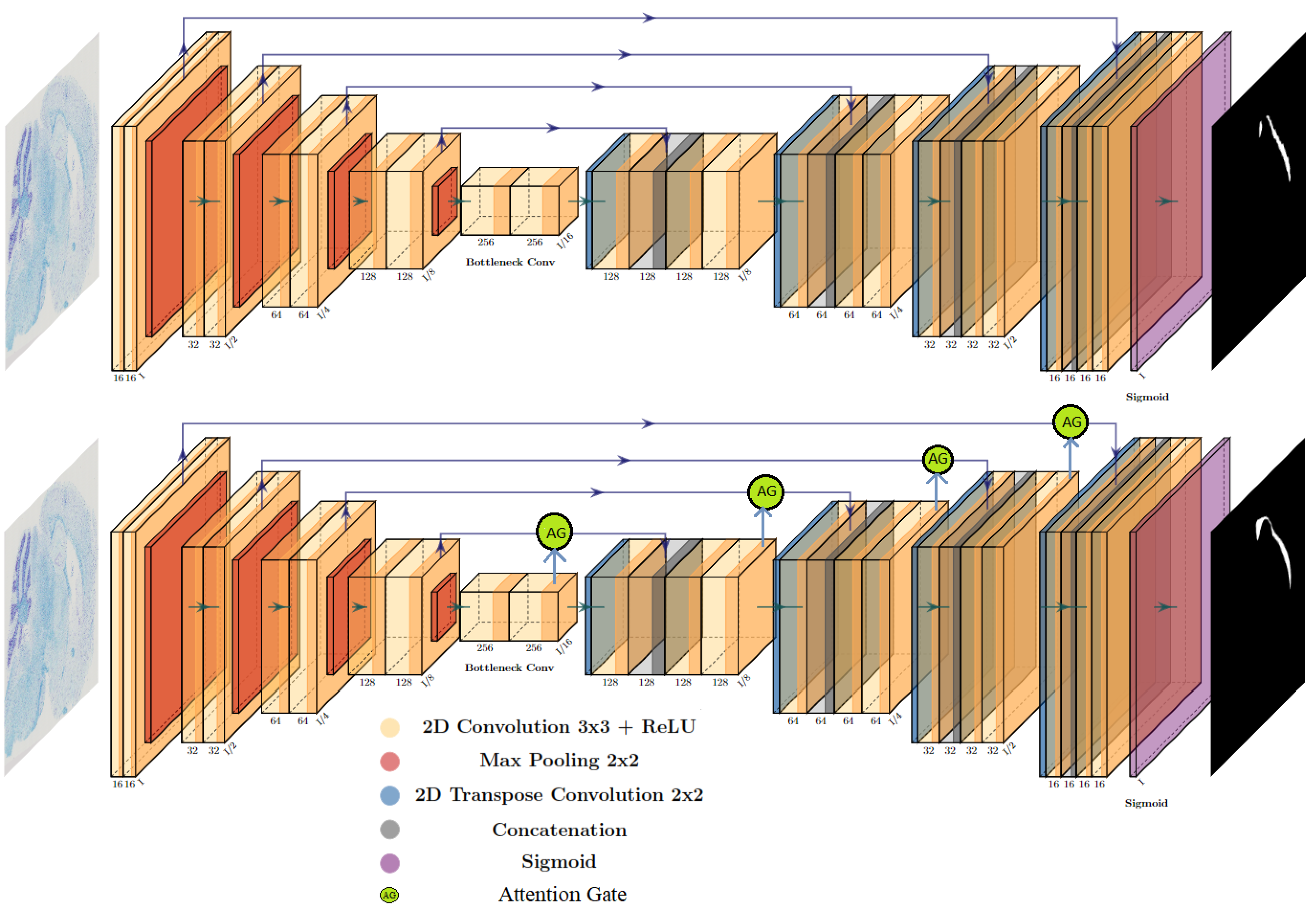
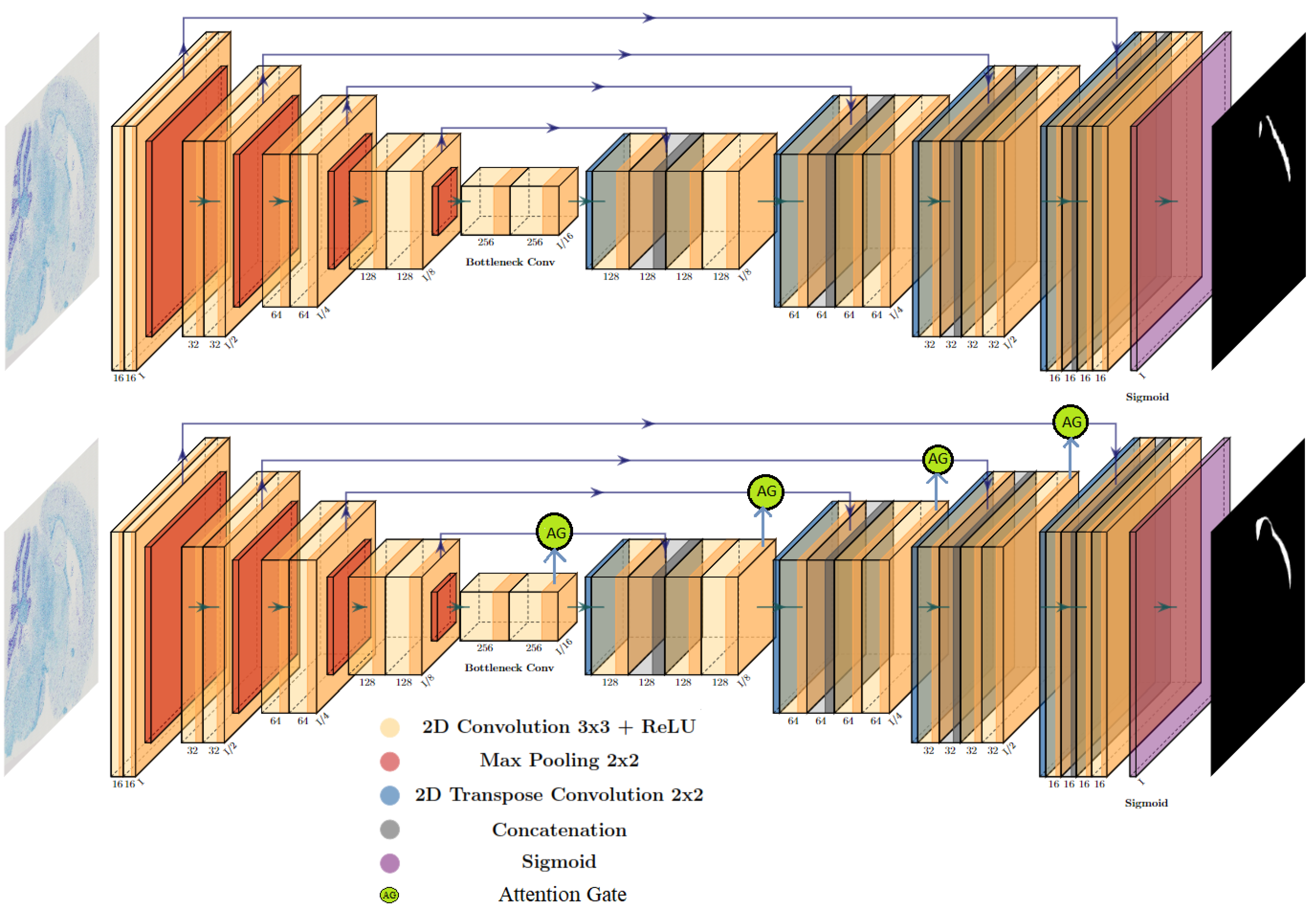
2.2. Deep Learning Models
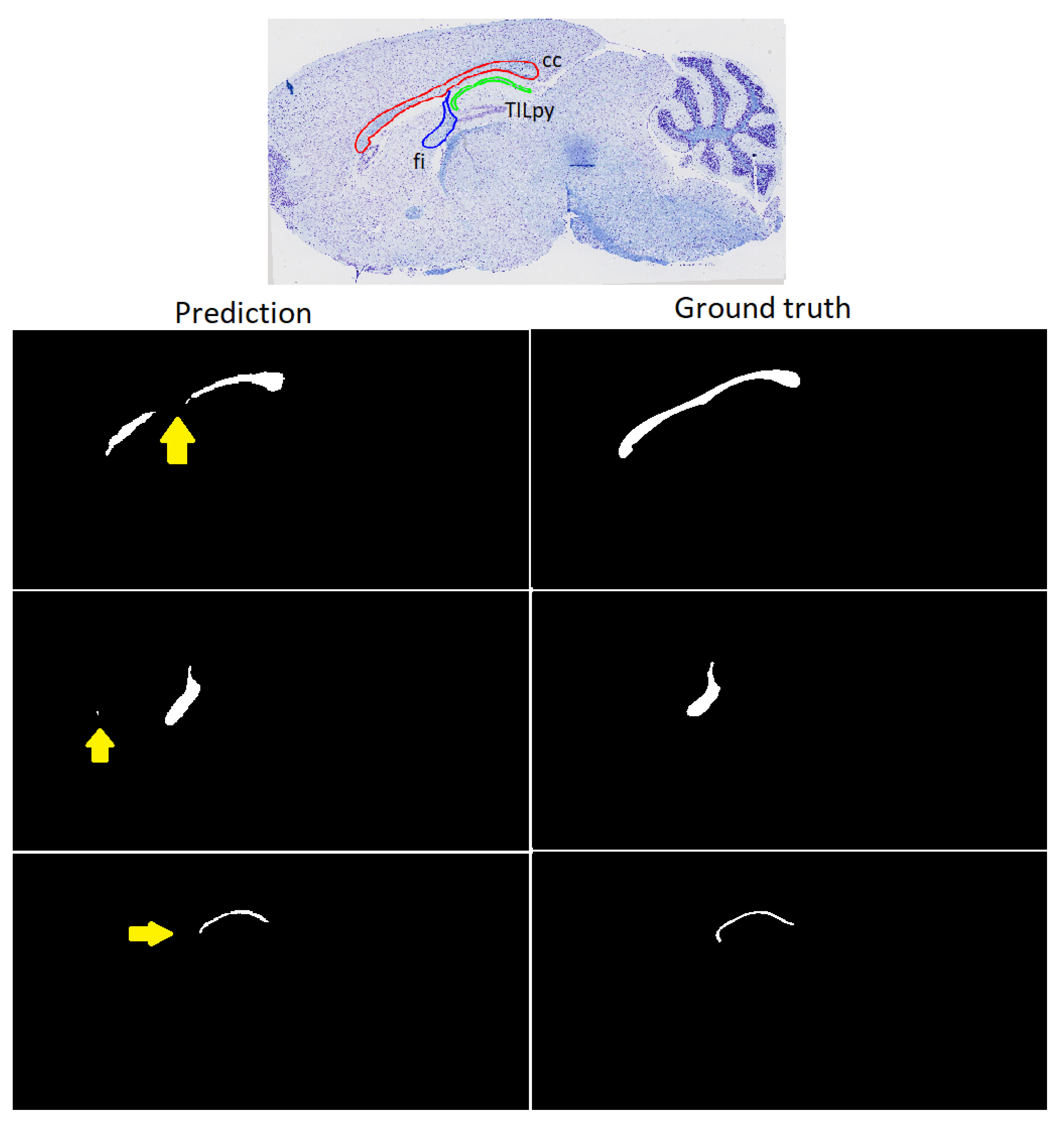
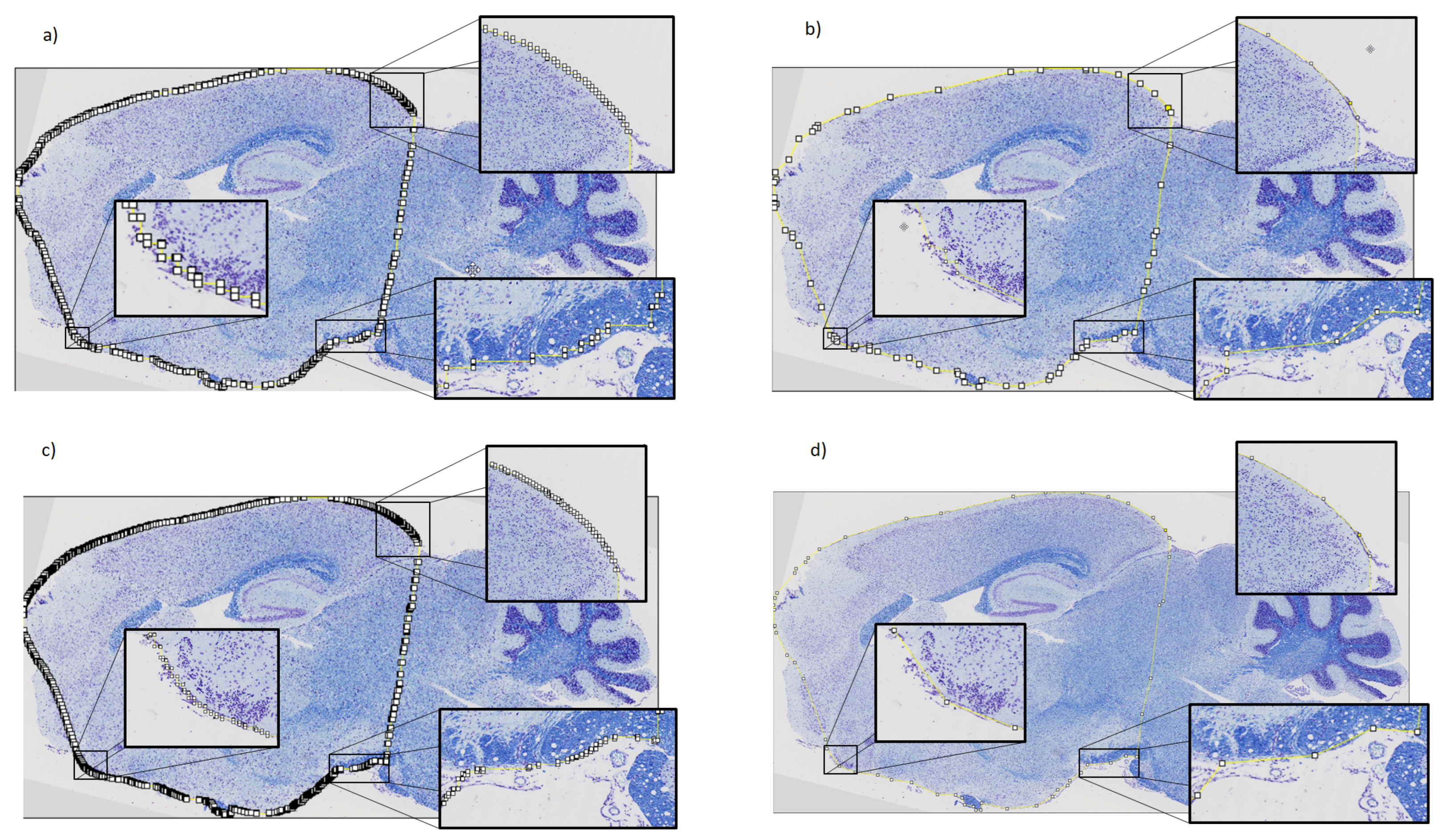
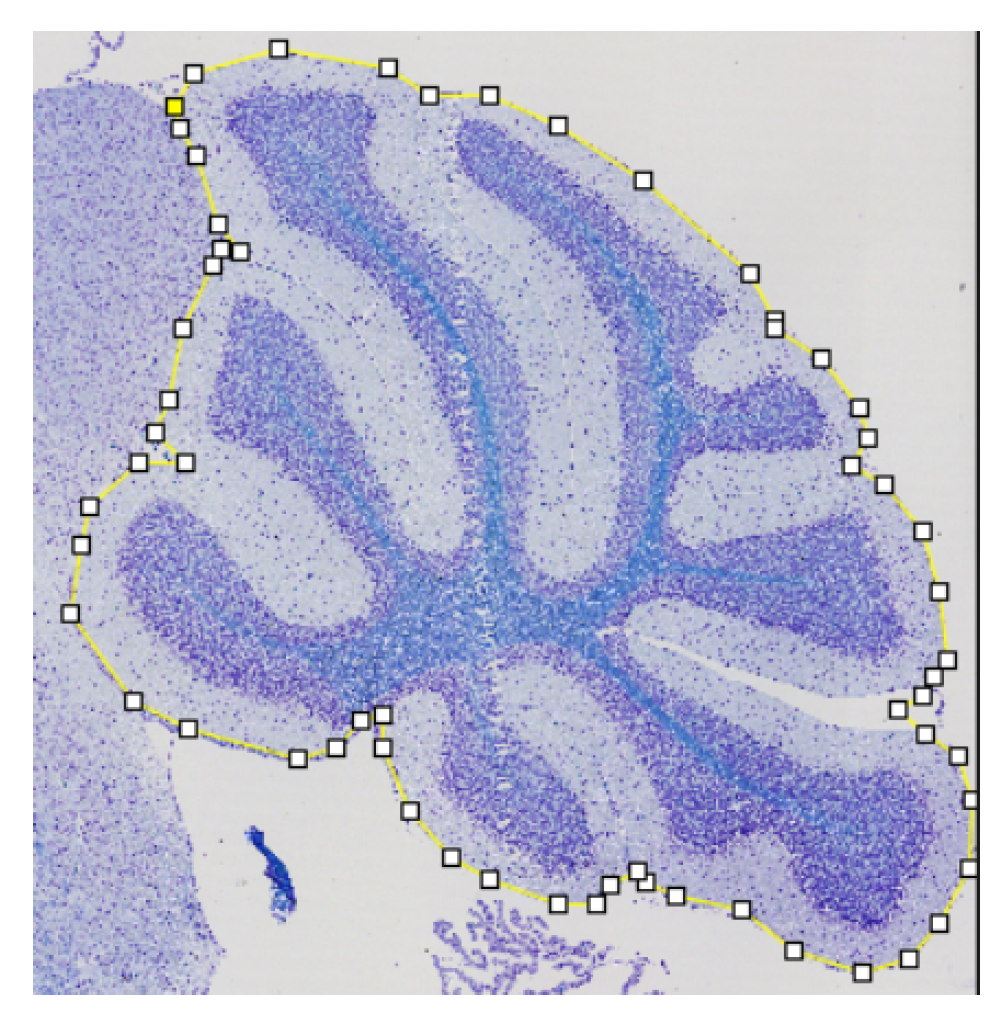
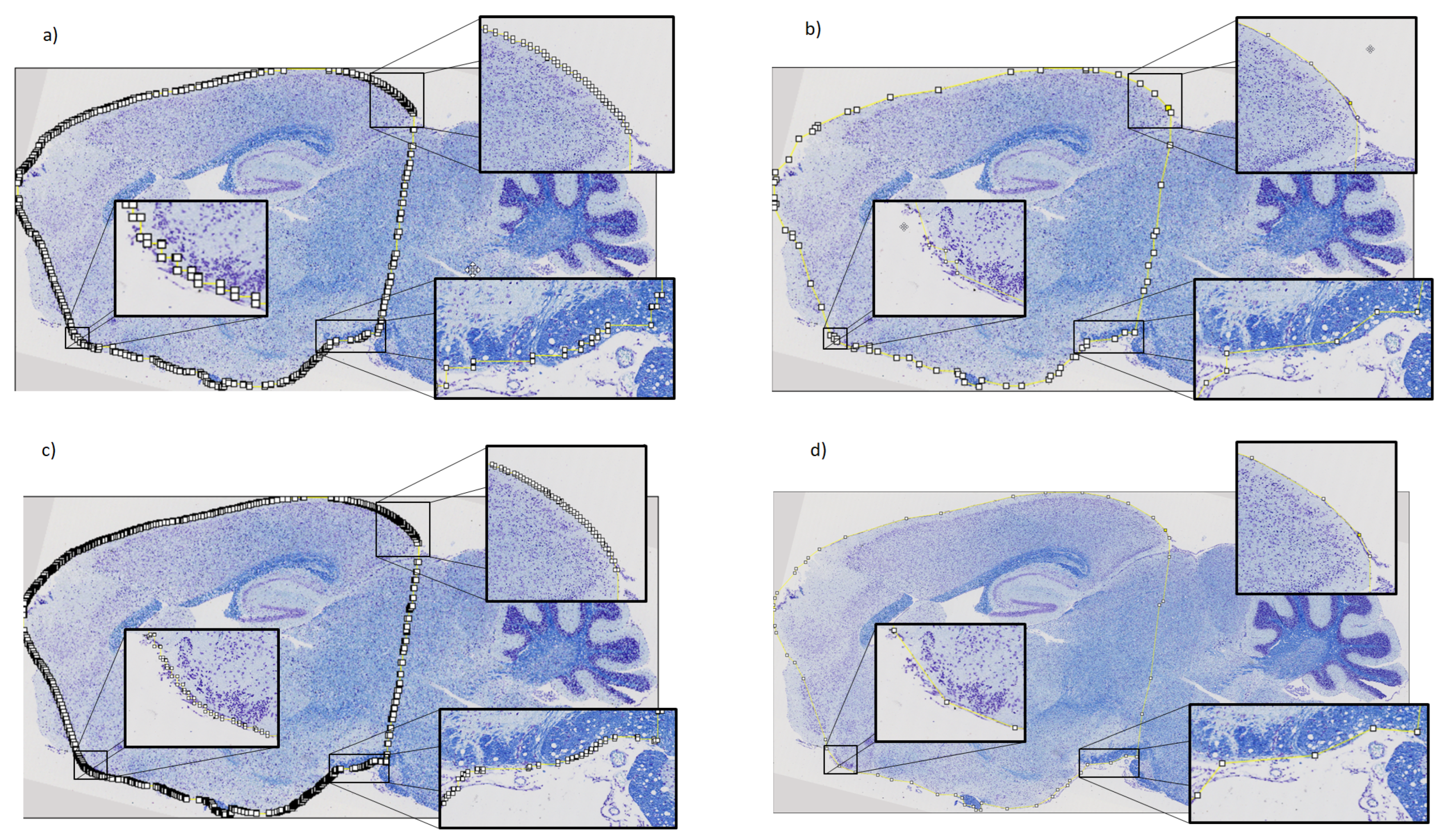
2.3. Refinement and Landmarks
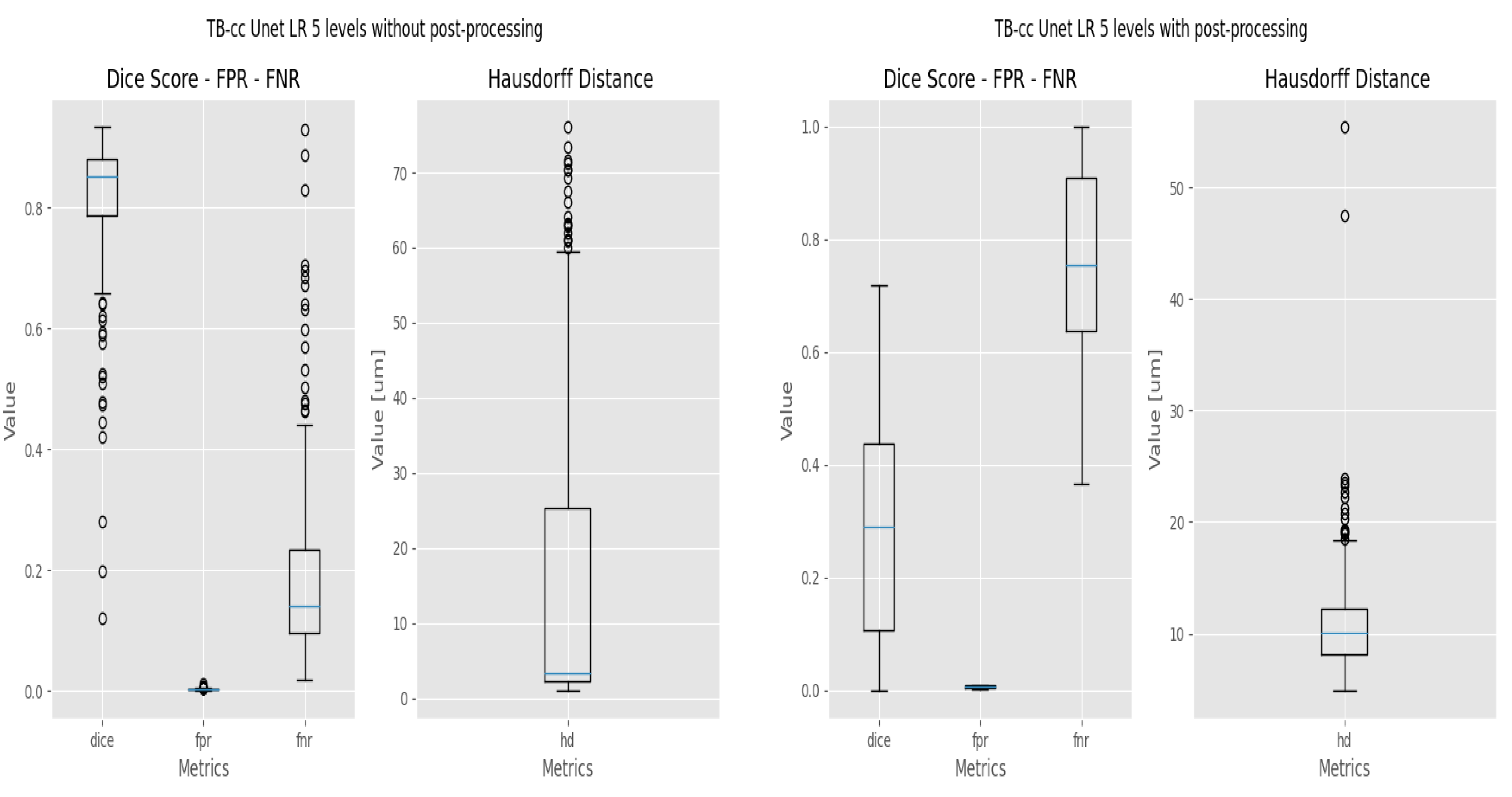
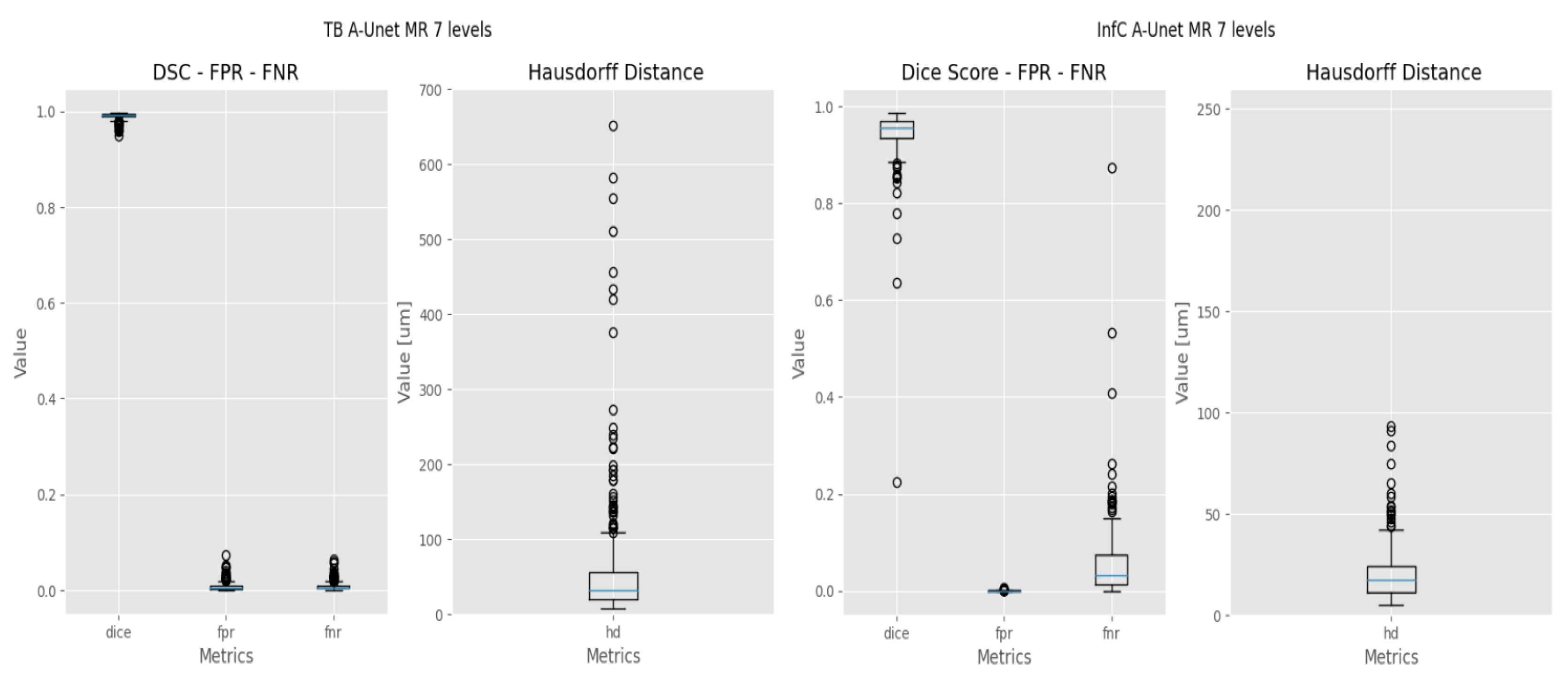
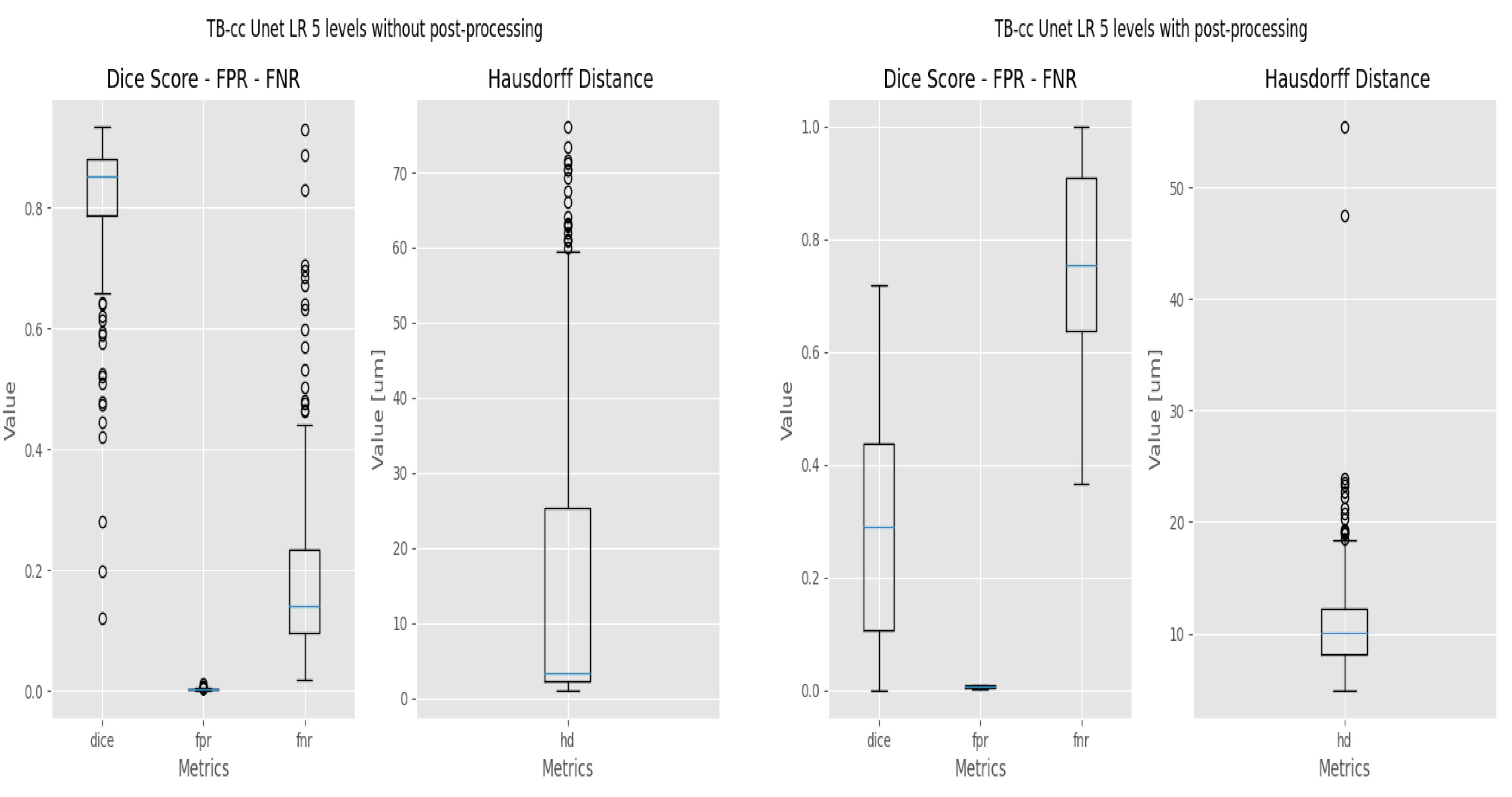
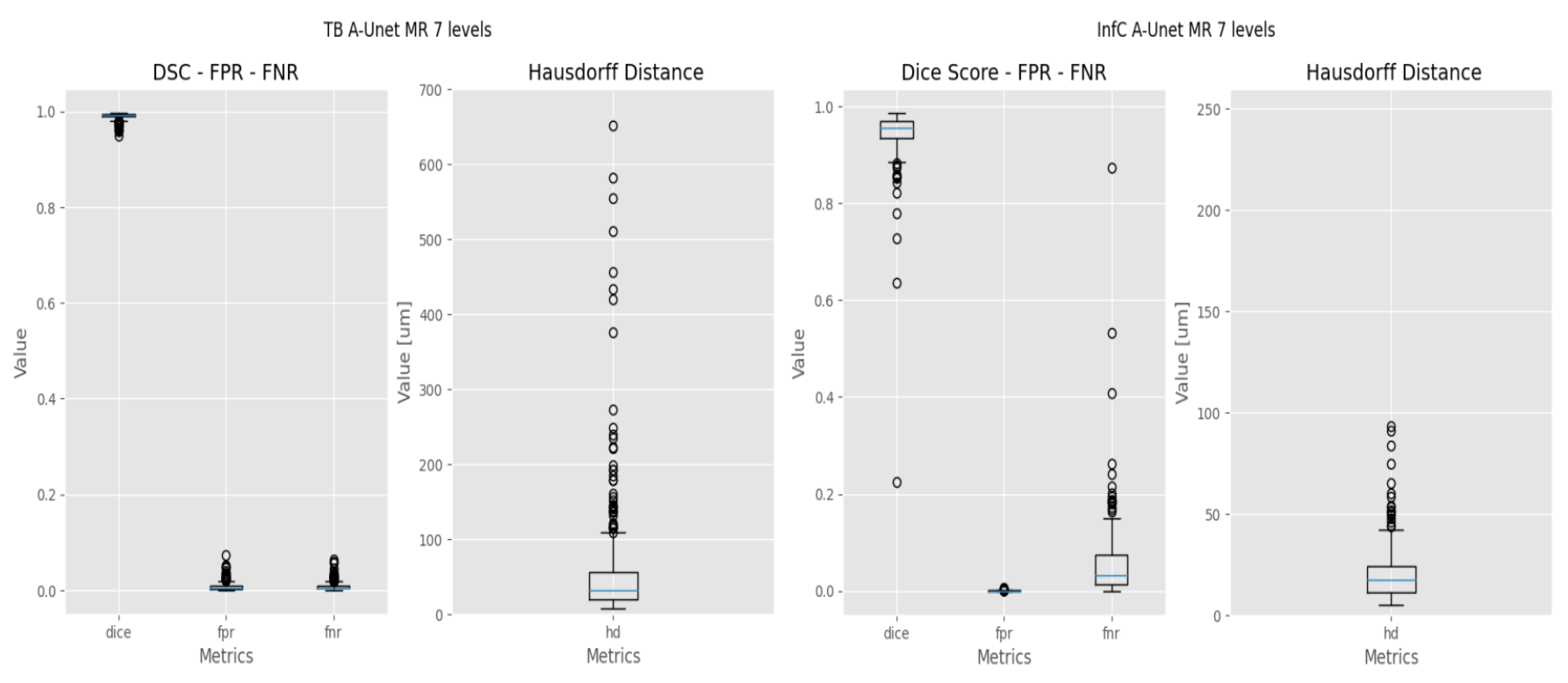
2.4. Evaluation Metrics
- The Dice coefficient, or Dice similarity coefficient [19], is a metric commonly used to evaluate the accuracy of segmentation results (Equation (1)). It measures the overlap between the predicted segmentation and the ground truth by calculating the ratio of twice the intersection of the two regions to the sum of their sizes.
- The False Positive Rate (FPR) [20] is a metric that measures the proportion of incorrect positive predictions made by the model (Equation (2)). A lower False Positive Rate indicates better performance, as it indicates a lower rate of false alarms or incorrect positive predictions.where FP = False Positive, TN = True Negative.
- The False Negative Rate (FNR). Ref. [20] measures the proportion of missed positive predictions by the model (Equation (3)). A lower False Negative Rate is desired as it signifies a lower rate of missed detections or incorrectly classified negatives, indicating better sensitivity and accuracy in capturing the target structure or region.where TP = True Positive, FN = False Negative.Both FPR and FNR will be used to evaluate the response of the models at pixel level.
- The Hausdorff Distance (HD) [21] measures the dissimilarity between two sets of points or contours (Equation (4)). It quantifies the maximum distance between any point in one set to the closest point in the other set.where:
- d(a, B) represents the minimum distance between a point a in set A and the closest point in set B.
- d(b, A) represents the minimum distance between a point b in set B and the closest point in set A.
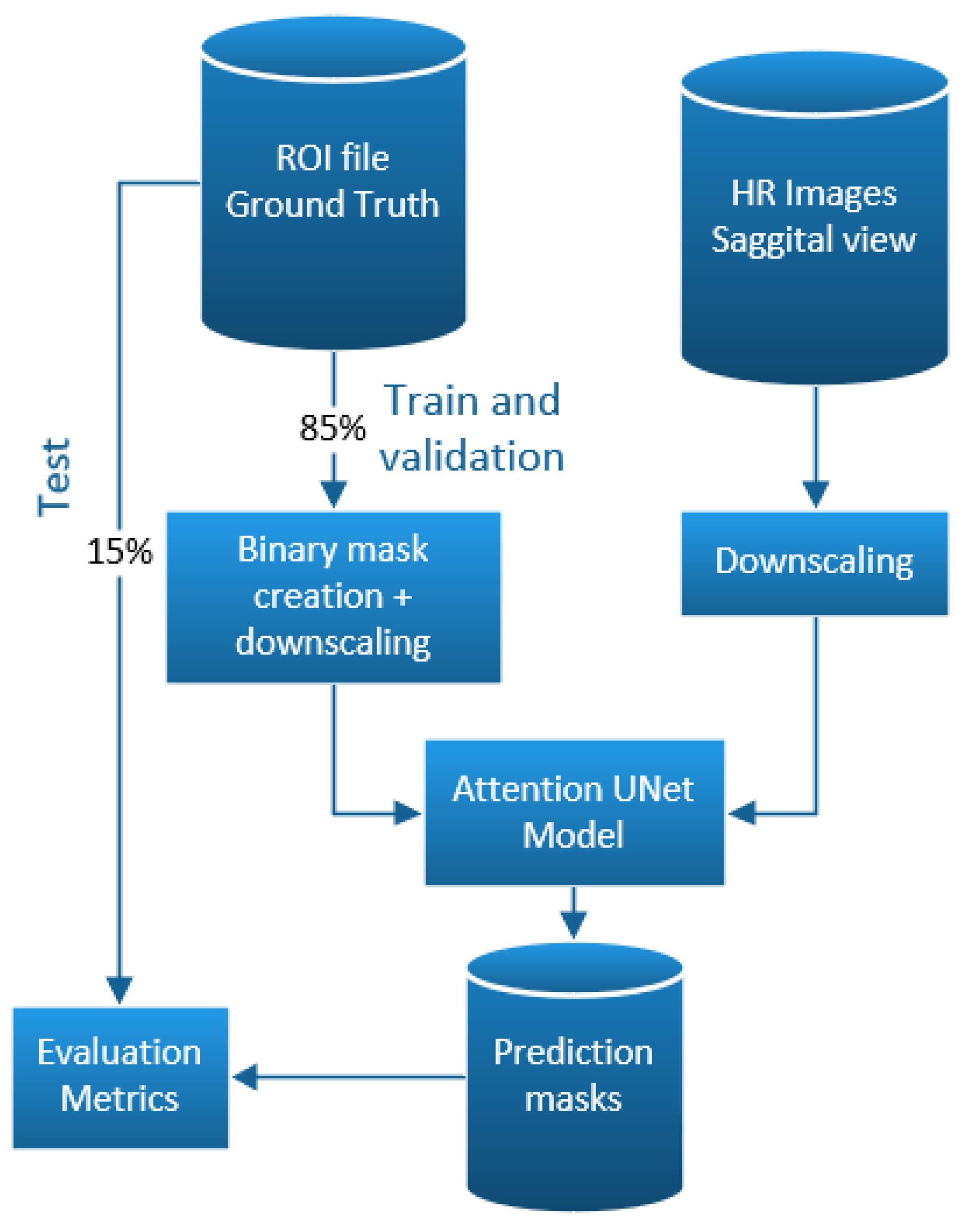
2.5. Implementation
3. Results
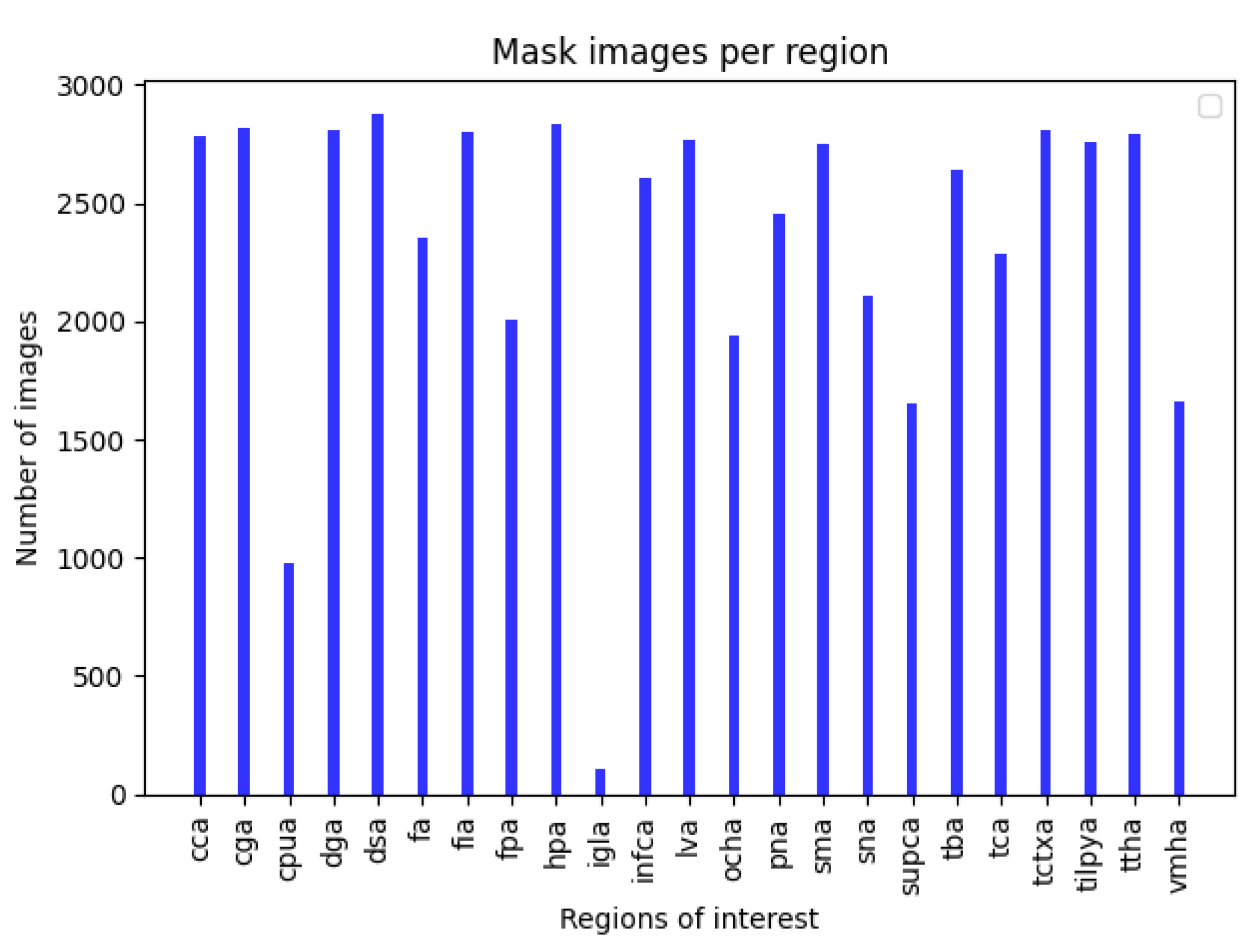
3.1. Data Preparation
3.2. Deep Learning
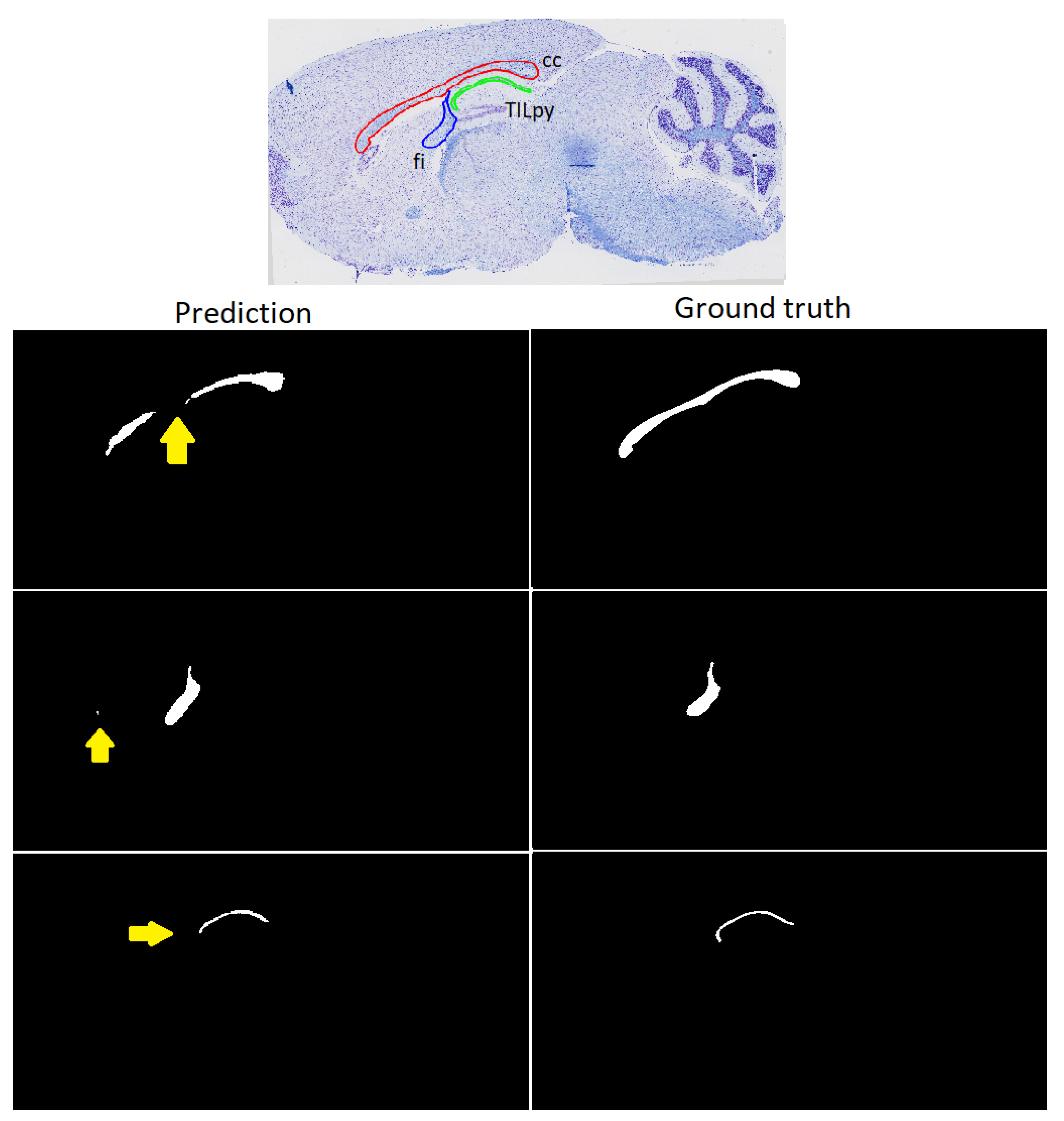
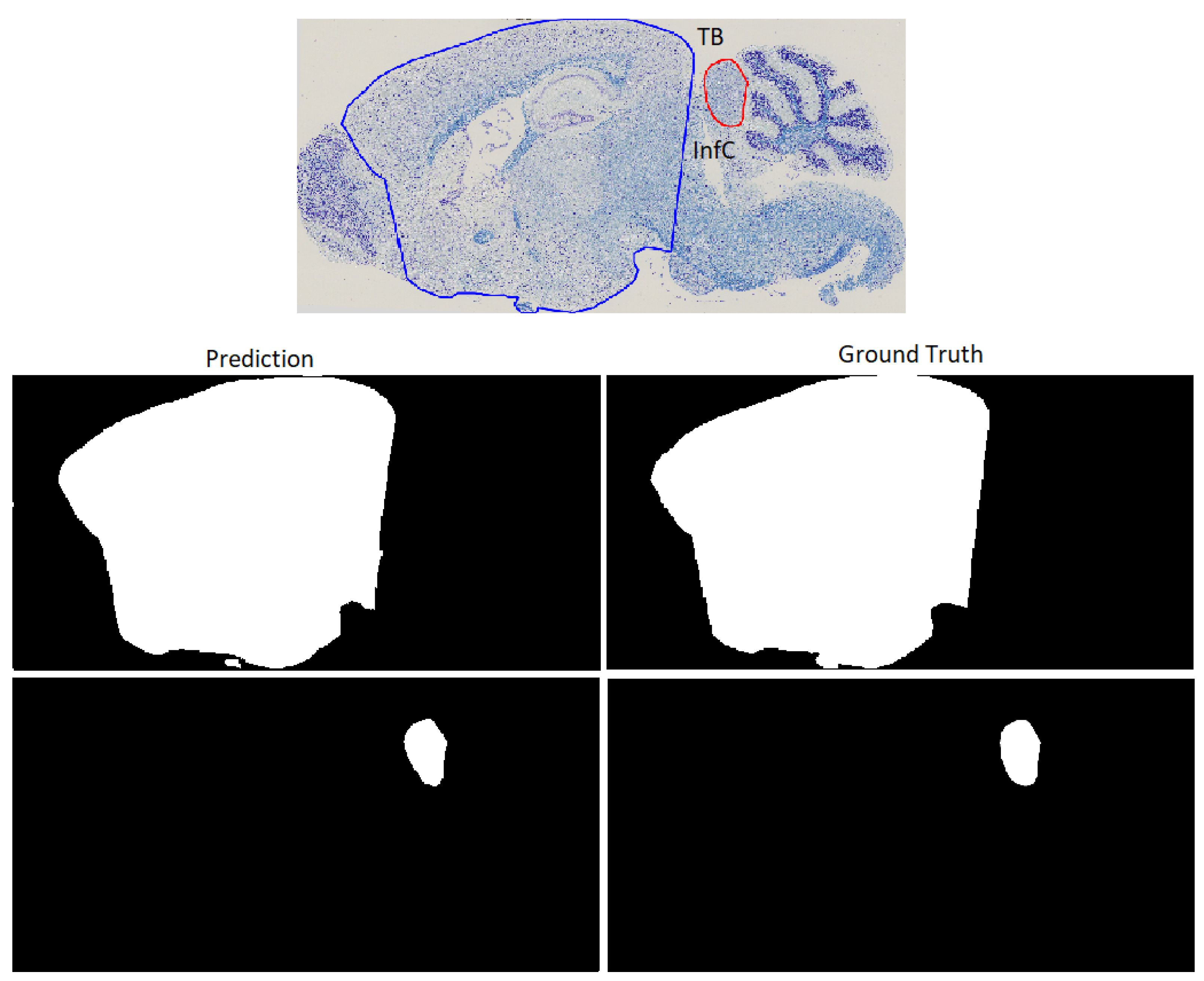
3.3. Refinement and Landmark
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bossert, L.; Hagendorff, T. Animals and AI. The role of animals in AI research and application—An overview and ethical evaluation. Technol. Soc. 2021, 67, 101678. [Google Scholar] [CrossRef]
- Collins, F.S.; Rossant, J.; Wurst, W. A mouse for all reasons. Cell 2007, 128, 9–13. [Google Scholar] [PubMed]
- Mohun, T.; Adams, D.J.; Baldock, R.; Bhattacharya, S.; Copp, A.J.; Hemberger, M.; Houart, C.; Hurles, M.E.; Robertson, E.; Smith, J.C.; et al. Deciphering the Mechanisms of Developmental Disorders (DMDD): A new programme for phenotyping embryonic lethal mice. Dis. Model. Mech. 2013, 6, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.; Mikhaleva, A.; Vrcelj, K.; Vancollie, V.; Wagner, C.; Demeure, N.; Whitley, H.; Kannan, M.; Balz, R.; Anthony, L.; et al. Large-scale neuroanatomical study uncovers 198 gene associations in mouse brain morphogenesis. Nat. Commun. 2019, 10, 3465. [Google Scholar] [CrossRef] [PubMed]
- Kretz, P.F.; Wagner, C.; Montillot, C.; Hugel, S.; Morella, I.; Kannan, M.; Mikhaleva, A.; Fischer, M.C.; Milhau, M.; Brambilla, R.; et al. Dissecting the autism-associated 16p11. 2 locus identifies multiple drivers in brain phenotypes and unveils a new role for the major vault protein. Genome Biol. 2023, in press. [Google Scholar]
- Groeneboom, N.E.; Yates, S.C.; Puchades, M.A.; Bjaalie, J.G. Nutil: A pre-and post-processing toolbox for histological rodent brain section images. Front. Neuroinform. 2020, 14, 37. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Liu, L.; Lei, X.; Mandal, M.; Lu, C. An unsupervised method for histological image segmentation based on tissue cluster level graph cut. Comput. Med. Imaging Graph. 2021, 93, 101974. [Google Scholar] [CrossRef] [PubMed]
- Yates, S.C.; Groeneboom, N.E.; Coello, C.; Lichtenthaler, S.F.; Kuhn, P.H.; Demuth, H.U.; Hartlage-Rübsamen, M.; Roßner, S.; Leergaard, T.; Kreshuk, A.; et al. QUINT: Workflow for quantification and spatial analysis of features in histological images from rodent brain. Front. Neuroinform. 2019, 13, 75. [Google Scholar] [CrossRef] [PubMed]
- Puchades, M.A.; Csucs, G.; Ledergerber, D.; Leergaard, T.B.; Bjaalie, J.G. Spatial registration of serial microscopic brain images to three-dimensional reference atlases with the QuickNII tool. PLoS ONE 2019, 14, e0216796. [Google Scholar] [CrossRef] [PubMed]
- Sommer, C.; Straehle, C.; Koethe, U.; Hamprecht, F.A. Ilastik: Interactive learning and segmentation toolkit. In Proceedings of the 2011 IEEE International Symposium on Biomedical Imaging: From Nano to Macro, Chicago, IL, USA, 30 March–2 April 2011; pp. 230–233. [Google Scholar]
- Xu, X.; Guan, Y.; Gong, H.; Feng, Z.; Shi, W.; Li, A.; Ren, M.; Yuan, J.; Luo, Q. Automated brain region segmentation for single cell resolution histological images based on Markov random field. Neuroinformatics 2020, 18, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Mesejo, P.; Ugolotti, R.; Cagnoni, S.; Di Cunto, F.; Giacobini, M. Automatic segmentation of hippocampus in histological images of mouse brains using deformable models and random forest. In Proceedings of the 2012 25th IEEE International Symposium on Computer-Based Medical Systems (CBMS), Rome, Italy, 20–22 June 2012; pp. 1–4. [Google Scholar]
- Barzekar, H.; Ngu, H.; Lin, H.H.; Hejrati, M.; Valdespino, S.R.; Chu, S.; Bingol, B.; Hashemifar, S.; Ghosh, S. Multiclass Semantic Segmentation to Identify Anatomical Sub-Regions of Brain and Measure Neuronal Health in Parkinson’s Disease. arXiv 2023, arXiv:2301.02925. [Google Scholar]
- Ronneberger, O.; Fischer, P.; Brox, T. U-Net: Convolutional Networks for Biomedical Image Segmentation. In Proceedings of the Medical Image Computing and Computer-Assisted Intervention–MICCAI 2015: 18th International Conference, Munich, Germany, 5–9 October 2015; Proceedings, Part III. pp. 234–241. [Google Scholar]
- Hamida, A.B.; Devanne, M.; Weber, J.; Truntzer, C.; Derangère, V.; Ghiringhelli, F.; Forestier, G.; Wemmert, C. Weakly Supervised Learning using Attention gates for colon cancer histopathological image segmentation. Artif. Intell. Med. 2022, 133, 102407. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.C.; Wagner, C.; Gagliardi, L.; Kretz, P.F.; Fischer, M.C.; Kessler, P.; Kannan, M.; Yalcin, B. A method for parasagittal sectioning for neuroanatomical quantification of brain structures in the adult mouse. Curr. Protoc. Mouse Biol. 2018, 8, e48. [Google Scholar] [CrossRef] [PubMed]
- Visvalingam, M.; Whyatt, J.D. The Douglas-Peucker algorithm for line simplification: Re-evaluation through visualization. In Computer Graphics Forum; Wiley Online Library: Hoboken, NJ, USA, 1990; Volume 9, pp. 213–225. [Google Scholar]
- Bimanjaya, A.; Handayani, H.H.; Rachmadi, R.F. Extraction of Road Network in Urban Area from Orthophoto Using Deep Learning and Douglas-Peucker Post-Processing Algorithm. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2023; Volume 1127, p. 012047. [Google Scholar]
- Zou, K.H.; Warfield, S.K.; Bharatha, A.; Tempany, C.M.; Kaus, M.R.; Haker, S.J.; Wells, W.M., III; Jolesz, F.A.; Kikinis, R. Statistical validation of image segmentation quality based on a spatial overlap index1: Scientific reports. Acad. Radiol. 2004, 11, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Moral, E.; Martins, R.; Wolf, D.; Rives, P. A new metric for evaluating semantic segmentation: Leveraging global and contour accuracy. In Proceedings of the 2018 IEEE Intelligent Vehicles Symposium (iv), Changshu, China, 26–30 June 2018; pp. 1051–1056. [Google Scholar]
- Sudha, N. Robust Hausdorff distance measure for face recognition. Pattern Recognit. 2007, 40, 431–442. [Google Scholar]
- Walsh, C.; Holroyd, N.A.; Finnerty, E.; Ryan, S.G.; Sweeney, P.W.; Shipley, R.J.; Walker-Samuel, S. Multifluorescence high-resolution episcopic microscopy for 3D imaging of adult murine organs. Adv. Photonics Res. 2021, 2, 2100110. [Google Scholar] [CrossRef]
- Scharrenberg, R.; Richter, M.; Johanns, O.; Meka, D.P.; Rücker, T.; Murtaza, N.; Lindenmaier, Z.; Ellegood, J.; Naumann, A.; Zhao, B.; et al. TAOK2 rescues autism-linked developmental deficits in a 16p11. 2 microdeletion mouse model. Mol. Psychiatry 2022, 27, 4707–4721. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
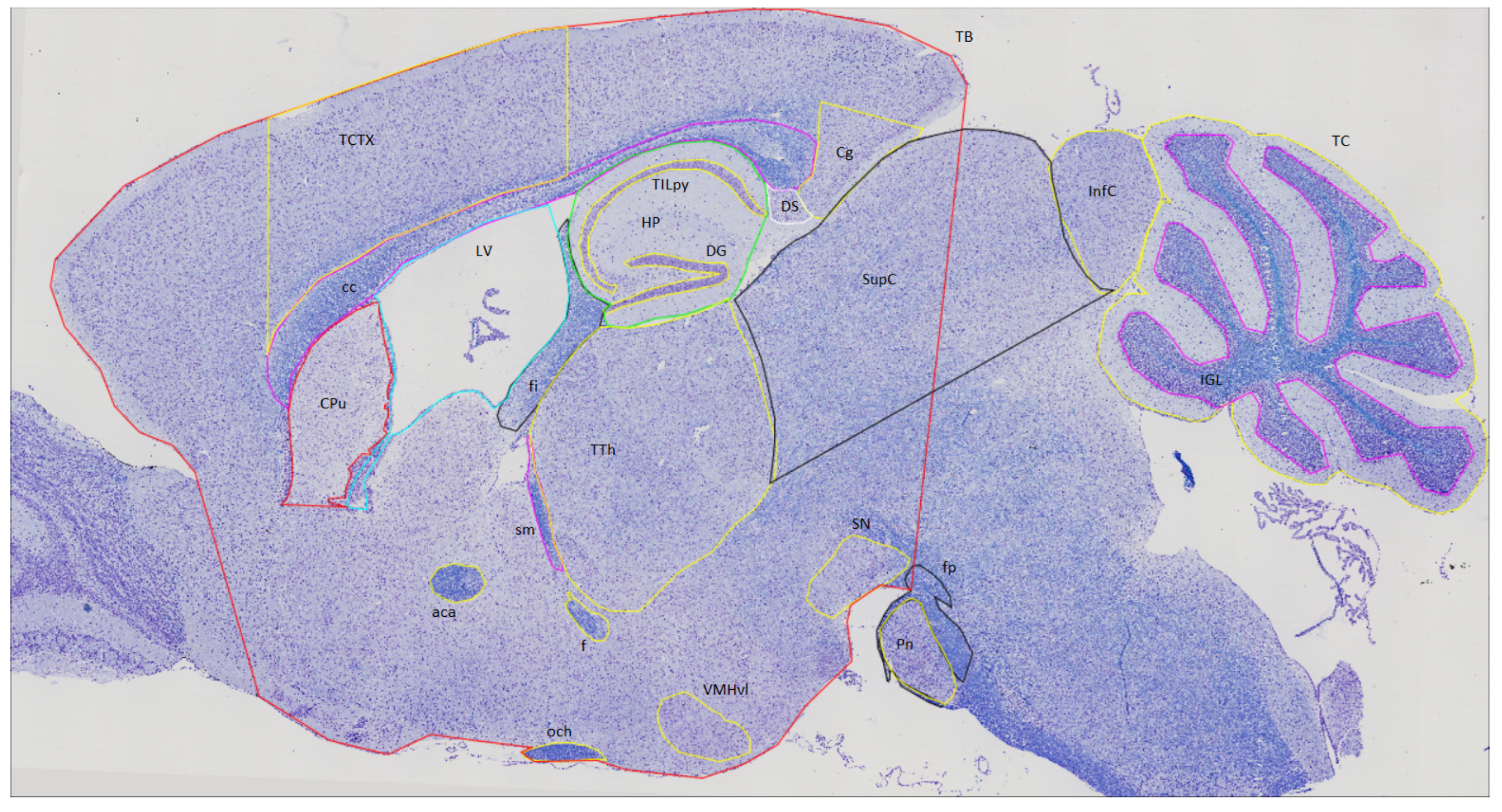
| GROUP 1 | GROUP 2 | ||||
|---|---|---|---|---|---|
| Name | Tag | Name | Tag | Name | Tag |
| Inferior Colliculus | InfC | anterior commisure | aca | Hippocampus | HP |
| Superior Colliculus | SupC | corpus callosum | cc | Lateral Ventricle | LV |
| Intra Granular Layer | IGL | Cingulate cortex | Cg | optic chiasm | och |
| Total Cerebellum | TC | Caudate Putamen | CPu | stria medularis | sm |
| Substancia Nigra | SN | Dentate Gyrus | DG | Total Cortical area | TCTX |
| Pontine nucleus | Pn | Dorsal Subiculum | DS | pyramidal cell layer | TILpy |
| fibers of the pons | fp | fornix | f | Thalamus | TTH |
| Total Brain | TB | fimbria | fi | Ventro Median Hypothalamus | VMHvl |
| TAG | DSC | FPR | FNR | HD m |
|---|---|---|---|---|
| aca | 0.9660 ± 0.0394 | 0.0001 ± 0.0001 | 0.0276 ± 0.0573 | 5.2889 ± 11.0271 |
| cc | 0.9365 ± 0.0347 | 0.0012 ± 0.0007 | 0.0421 ± 0.0463 | 15.1756 ± 14.8965 |
| f | 0.8828 ± 0.0957 | 0.0001 ± 0.0002 | 0.0822 ± 0.1161 | 12.686 ± 40.1613 |
| fi | 0.9179 ± 0.081 | 0.0004 ± 0.0004 | 0.0798 ± 0.1035 | 27.9981 ± 28.0597 |
| fp | 0.7047 ± 0.1758 | 0.0013 ± 0.0015 | 0.2383 ± 0.2184 | 66.8368 ± 52.7969 |
| och | 0.9214 ± 0.0801 | 0.0001 ± 0.0002 | 0.0613 ± 0.1021 | 12.8222 ± 22.2288 |
| sm | 0.8775 ± 0.0927 | 0.0003 ± 0.0004 | 0.0997 ± 0.1256 | 31.4965 ± 52.4748 |
| TB | 0.9914 ± 0.0062 | 0.0079 ± 0.0085 | 0.0078 ± 0.008 | 42.9213 ± 42.1242 |
| TCTX | 0.978 ± 0.028 | 0.0010 ± 0.0008 | 0.0205 ± 0.0387 | 20.3151 ± 28.2891 |
| TC | 0.9902 ± 0.0053 | 0.001 ± 0.0006 | 0.0087 ± 0.0083 | 24.1482 ± 23.9261 |
| IGL | 0.9267 ± 0.0865 | 0.0036 ± 0.0042 | 0.0546 ± 0.124 | 33.7096 ± 43.5162 |
| LV | 0.9452 ± 0.1069 | 0.0005 ± 0.001 | 0.0493 ± 0.1058 | 41.9781 ± 64.002 |
| TTh | 0.9515 ± 0.0268 | 0.0021 ± 0.0017 | 0.0492 ± 0.0435 | 34.1868 ± 16.472 |
| CPu | 0.7918 ± 0.2614 | 0.001 ± 0.0013 | 0.1846 ± 0.2654 | 43.735 ± 42.5232 |
| HP | 0.9848 ± 0.0068 | 0.0004 ± 0.0002 | 0.0122 ± 0.0109 | 12.6344 ± 7.6397 |
| TILpy | 0.8852 ± 0.0812 | 0.0003 ± 0.0001 | 0.0814 ± 0.105 | 18.8284 ± 24.7633 |
| DG | 0.9302 ± 0.077 | 0.0002 ± 0.0002 | 0.0465 ± 0.0862 | 8.0014 ± 11.5809 |
| Pn | 0.9500 ± 0.060 | 0.0002 ± 0.0002 | 0.0391 ± 0.0667 | 9.2620 ± 8.7277 |
| SN | 0.7647 ± 0.1958 | 0.0006 ± 0.0007 | 0.2014 ± 0.2184 | 28.2791 ± 20.1335 |
| Cg | 0.9087 ± 0.0612 | 0.0007 ± 0.0006 | 0.0817 ± 0.0873 | 23.341 ± 19.6141 |
| DS | 0.8842 ± 0.0784 | 0.0001 ± 0.0001 | 0.1012 ± 0.1103 | 10.8247 ± 6.6032 |
| InfC | 0.9442 ± 0.0613 | 0.0006 ± 0.0005 | 0.0542 ± 0.0784 | 19.608 ± 13.8624 |
| SupC | 0.9483 ± 0.0249 | 0.0035 ± 0.0032 | 0.0479 ± 0.0364 | 11.7670 ± 6.0383 |
| VMHvl | 0.8039 ± 0.1754 | 0.0005 ± 0.0004 | 0.1797 ± 0.2095 | 23.7869 ± 23.7924 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
{kind=link}
Cisneros, J.; Lalande, A.; Yalcin, B.; Meriaudeau, F.; Collins, S. Automatic Segmentation of Histological Images of Mouse Brains. Algorithms 2023, 16, 553. https://doi.org/10.3390/a16120553
Cisneros J, Lalande A, Yalcin B, Meriaudeau F, Collins S. Automatic Segmentation of Histological Images of Mouse Brains. Algorithms. 2023; 16(12):553. https://doi.org/10.3390/a16120553
Chicago/Turabian StyleCisneros, Juan, Alain Lalande, Binnaz Yalcin, Fabrice Meriaudeau, and Stephan Collins. 2023. "Automatic Segmentation of Histological Images of Mouse Brains" Algorithms 16, no. 12: 553. https://doi.org/10.3390/a16120553
APA StyleCisneros, J., Lalande, A., Yalcin, B., Meriaudeau, F., & Collins, S. (2023). Automatic Segmentation of Histological Images of Mouse Brains. Algorithms, 16(12), 553. https://doi.org/10.3390/a16120553
{kind=link}