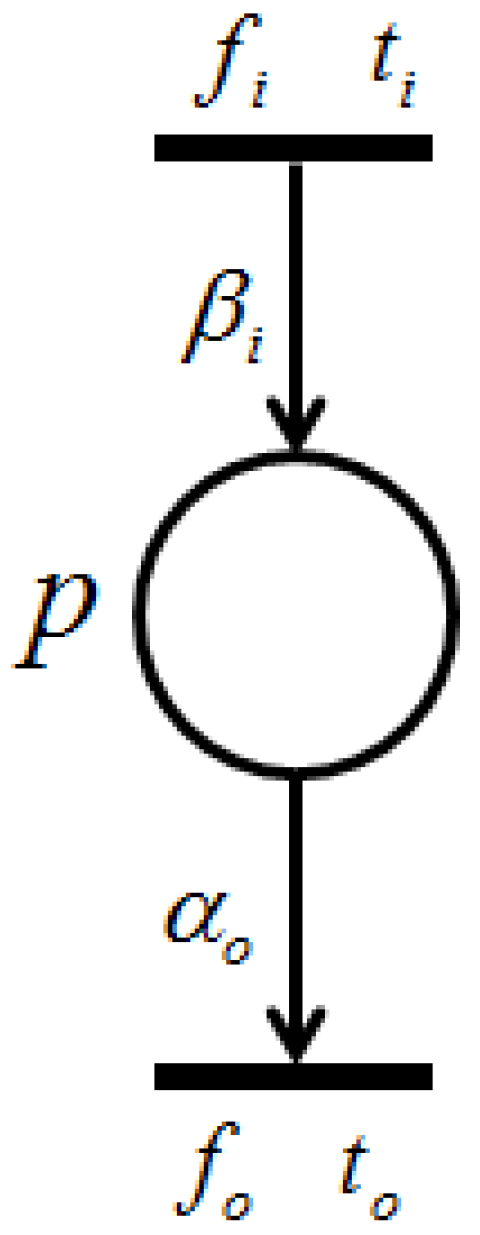1. Introduction
A Petri net [
1,
2] is a solid mathematical representation used in concurrent systems modeling, enabling a formal and clear representation of biological pathways at different time and scale levels [
3]. Structurally, a Petri net is a directed bipartite graph, including two types of nodes, places and transitions and tokens contained in places. A state of a Petri net is expressed by a token distribution over the places and can be changed by the firing of transitions. The first Petri net model of a biological pathway [
4] is a metabolic pathway, describing the conversion of metabolites through enzyme-catalyzed chemical reactions, which has been extensively studied regarding the structural [
5,
6,
7,
8] and dynamic [
9,
10,
11] properties.
A signaling pathway is another major biological pathway, consisting of cascades of activated/deactivated proteins or protein complexes, through which signals are propagated from the cell surface to the nucleus. A Petri net has been used as a structural modeling framework for a signal pathway in a knowledge representation [
12] and for data implementation in a database [
13].
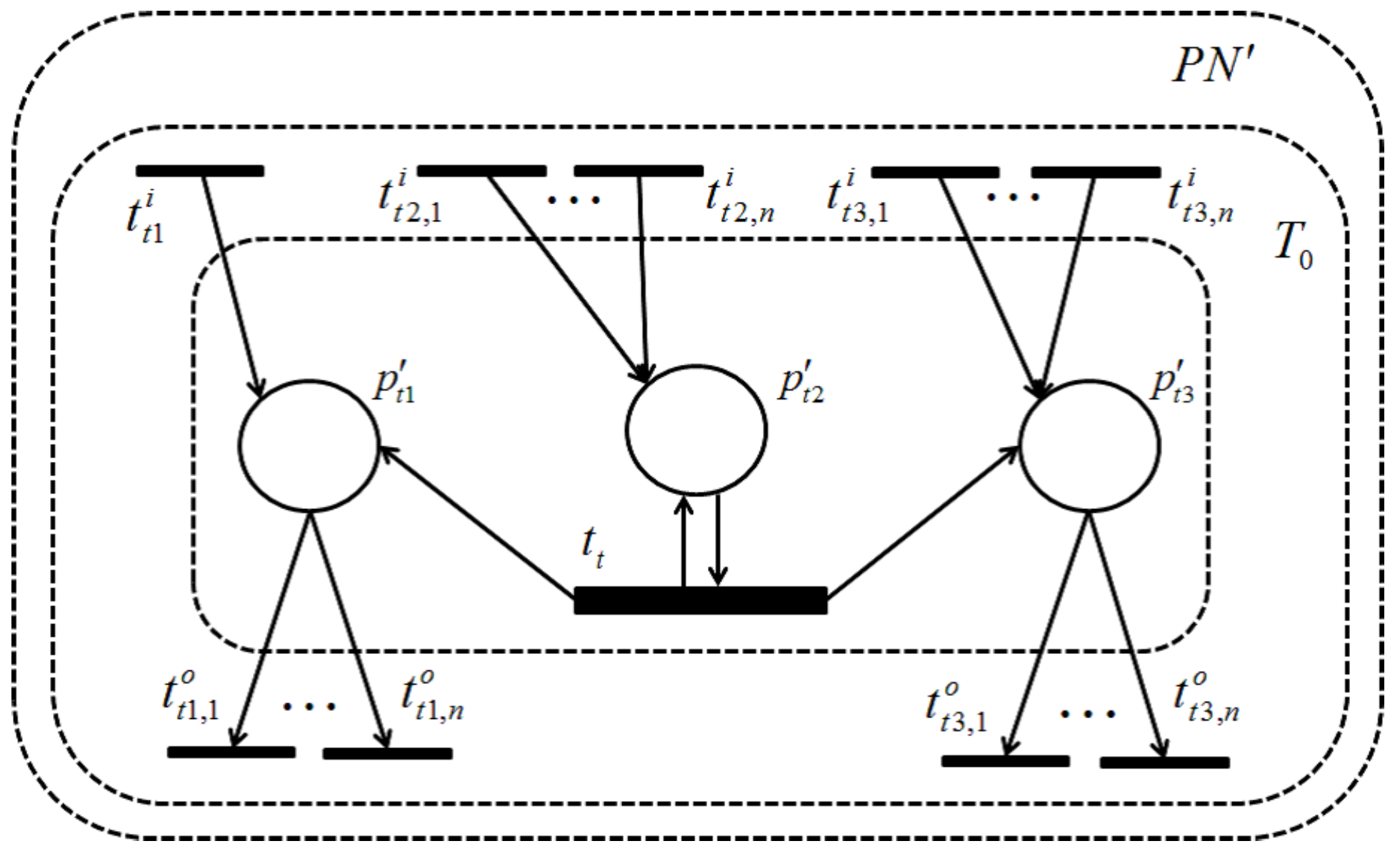
Modularization of a Petri net model into biologically-functional parts in a signaling pathway has been demonstrated using the fundamental behavioral property, T-invariant [
14], which was formalized in metabolic pathways before [
5,
6,
7,
8], and its extended property, the feasible T-invariant [
15]. Here, a T-invariant is a firing counting vector of transitions in a periodic firing sequence that leads back to the original token distribution where it starts from. A technique to automatically classify T-invariants into functional modules in a biological pathway was proposed in [
16] based on cluster analysis. Another classification of T-invariants in terms of biological network structure was demonstrated in [
17], where hierarchically-structured Petri net representations are derived automatically. T-invariant was further applied for the representation of the enzymatic activation process in a signaling pathway [
18]. The dynamic behaviors of a signaling pathway were examined in Petri net models for a token amount, reflecting the concentration levels of molecules obtained based on the order of the transition firings specified by the topological motifs in a signaling pathway [
19,
20], and were calculated after reducing the number of parameters through a simplification process of a Petri net model [
21].
The experimentally uncovered biological facts are usually summarized in a picture of a network structured by figures of various shapes and several type of arrows reflecting biological images. A Petri net allows us to model a biological pathway with maintaining its structural information, owing to the characteristics of the graphical representation of Petri net. Accordingly, Petri net-based modeling does not produce a negative effect of the abstraction in a modeling process compared to other modeling methods, such as differential equation-based methods.
The timed Petri net is a type of Petri net, in which time delay is associated with state transition, and is a promising candidate for modeling the dynamic property of a signaling pathway because the delay time of a transition is able to reflect the rate of a corresponding reaction in a signaling pathway. However, in the works above [
18,
19,
20,
21], a timed Petri net has not been employed.
The first attempt to use a timed Petri net for the modeling of a signaling pathway was in [
22] (the “time” Petri net, which has a different extension in terms of time to the “timed” Petri net, was applied to model metabolic pathways in [
11]), where the delay times were calculated based on a simple rule in which the sum of the consumption is equal to the production, so as to maintain the concentration equilibrium for each substance in a signaling pathway. Such Petri nets satisfying this simple rule are called retention-free Petri nets later and introduced as a new concept enabling a more flexible determination of the delay times using a concrete algorithm [
23]. Cycles and inhibitory arcs, which often appear in signaling pathways, are treated in [
24], but have not been incorporated owing to the difficulties in their structural handling.
These works [
22,
23,
24] have studied a Petri net-based methodology to calculate estimated values for non-valid reaction speeds from a valid reaction speed, namely a biologically-measured speed, in the signaling pathway. A simple question therefore arises. How many and which reactions should be biologically measured in order to obtain all of the delay times in a Petri net model through the proposed methodology? This paper answers this question by introducing the concept of a “dependency relation” over a transition set of a Petri net, by which the transition set is divided into equivalence classes in the sense of a dependency relation, namely an equivalence relation. In fact, the number of equivalence classes is the number of reactions that should be biologically measured.
One possible way to find such an equivalence class is to establish a series of linear equations for all places based on the retention-freeness, solve the simultaneous equations to obtain the dependent solutions and, finally, determine the transitions with a dependency relation. However, this will take plenty of computation time for a given large-scale signaling pathway model. To cope with this problem, this paper gives an efficient algorithm to reduce the size of a Petri net through a structural analysis and, finally, to discover these equivalence classes.
The remainder of this paper is organized as follows.
Section 2 provides the necessary definitions of Petri nets, and
Section 3 introduces a timed Petri net and retention-free Petri net (
). In
Section 4, the concept of a dependency relation over the transitions of
is introduced, and related properties are derived through a structural analysis. In
Section 5, based on the theoretical results of
Section 4, an algorithm is proposed and applied to the IL-3 Petri net model to show its usefulness. Finally,
Section 6 provides some concluding remarks and describes some future works.
3. Timed Petri Net and Retention-Free Petri Net
Petri nets are extended by assigning a delay time of the firing to each transition to facilitate a system-level understanding through means of a simulation. Such an extended Petri net is called a timed Petri net.
Definition 5. Timed Petri net is defined by , where D is a set of positive numbers expressing the firing delay times (or delay time for short) of transitions in T. The firing rule of a is as follows:initially, all of the tokens are in a non-reserved state; once a transition is decided to have fired, the tokens required for firing are changed from the non-reserved state to the reserved state;
when the delay time of has passed, fires to remove the reserved tokens from the input places of and put non-reserved tokens into the output places of .
In a timed Petri net, the firing times of a transition per unit of time are called firing frequency . represents the maximum firing frequency of . The delay time of is given by the reciprocal of .
Note that firing frequencies indicate reaction speed data in a biological pathway.
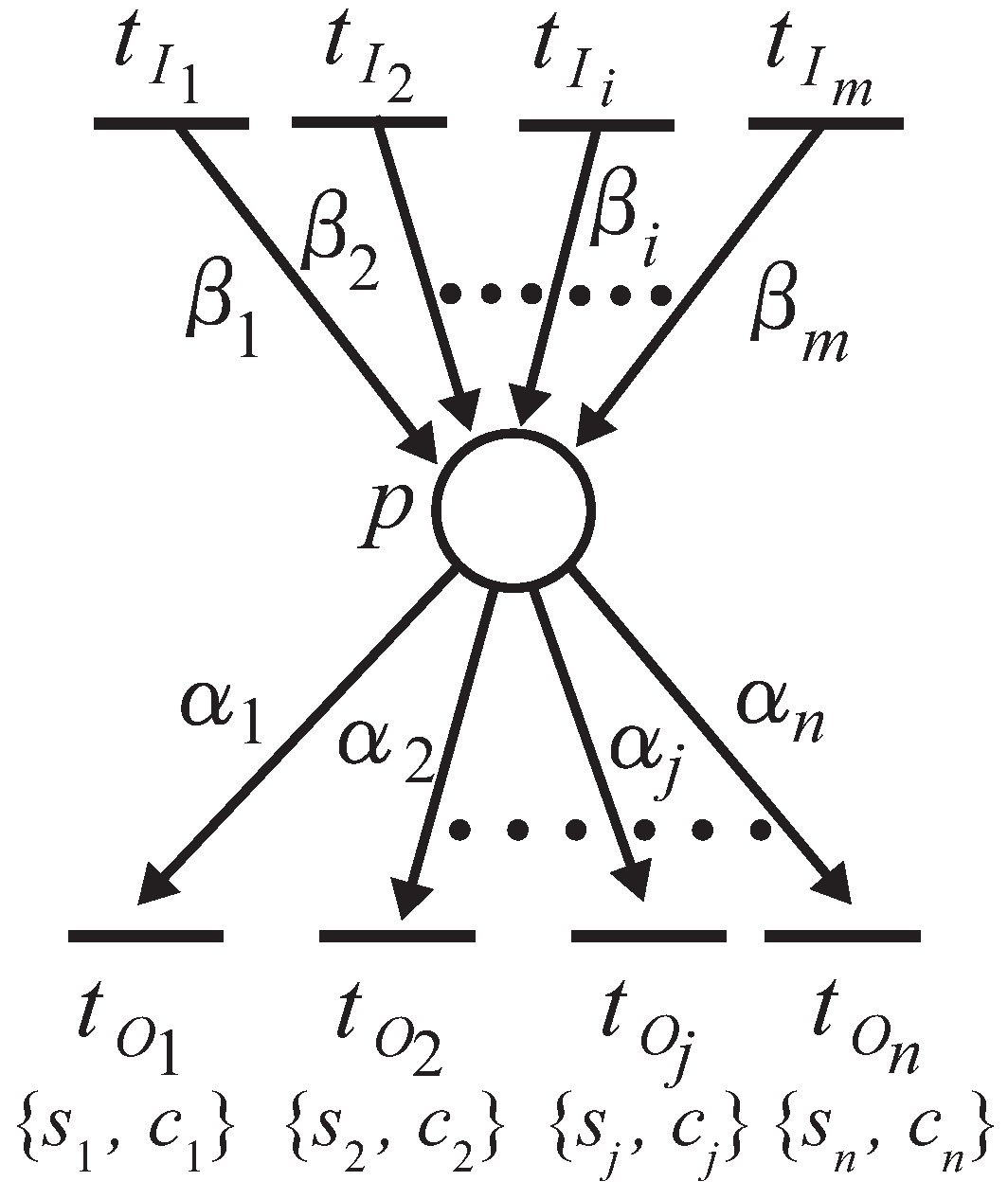
We show here the method of conflict resolution using stochastic decision rules. To resolve the conflict firing problem, there are three methods that can be adopted: priority, probabilistic choice and alternate firing [
25]. The proposed firing rule given in this paper is to combine a probabilistic choice and alternate firing. We introduce a stochastic approach to determine the firings for a series of transitions in conflict (
Figure 5), as defined in the following definition.
Definition 6. Suppose a place p possesses output transitions, , as shown in Figure 5. Then, the firing rule is as follows [
23]
:Each unreserved token deposited to input place p is assigned to be reserved by the transition that satisfies the following expression:where is decided by the following condition: When the number of reserved tokens of is not less than the required token number for the firing, the firing of is decided;
After the delay time of has passed, fires to remove the reserved tokens from the input place of and deposits unreserved tokens into the output places of .
In the above expression (
2),
is the arc weight of
;
is the firing probability of transition
, which represents the proportion of the firing frequency of each transition within the total firing frequency of the transitions in conflict. Probability
is assigned to the corresponding transition
, which is given as a constant in advance according to the event. Variable
is an accumulated number of tokens that
has reserved so far, and thus,
represents the number of firing times of transition
from the beginning. Expression (
2) is designed to reserve the token to such a transition
that has the largest difference between the calculated firing probability
and the given firing probability
among all transitions in conflict.
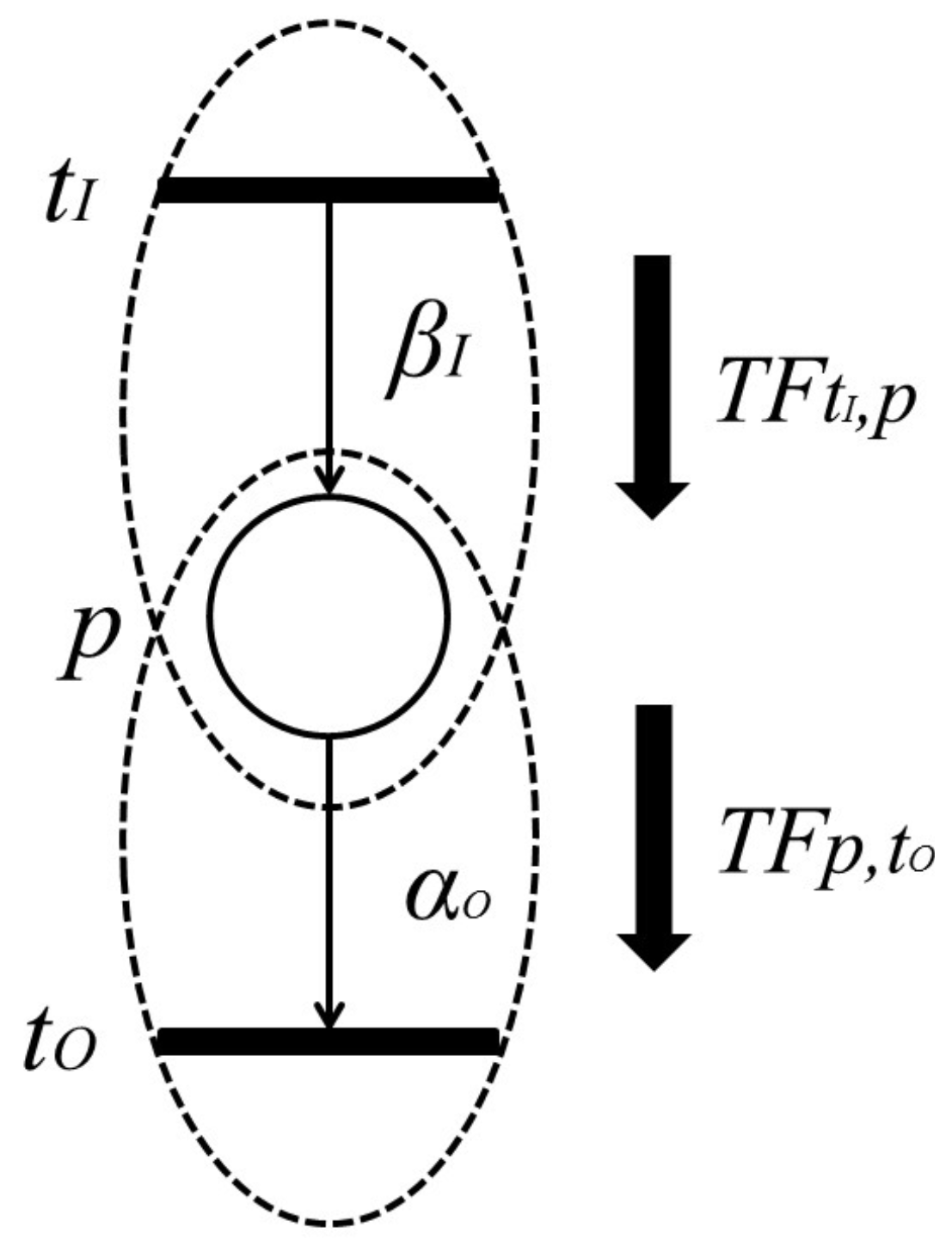
Definition 7. [
23]
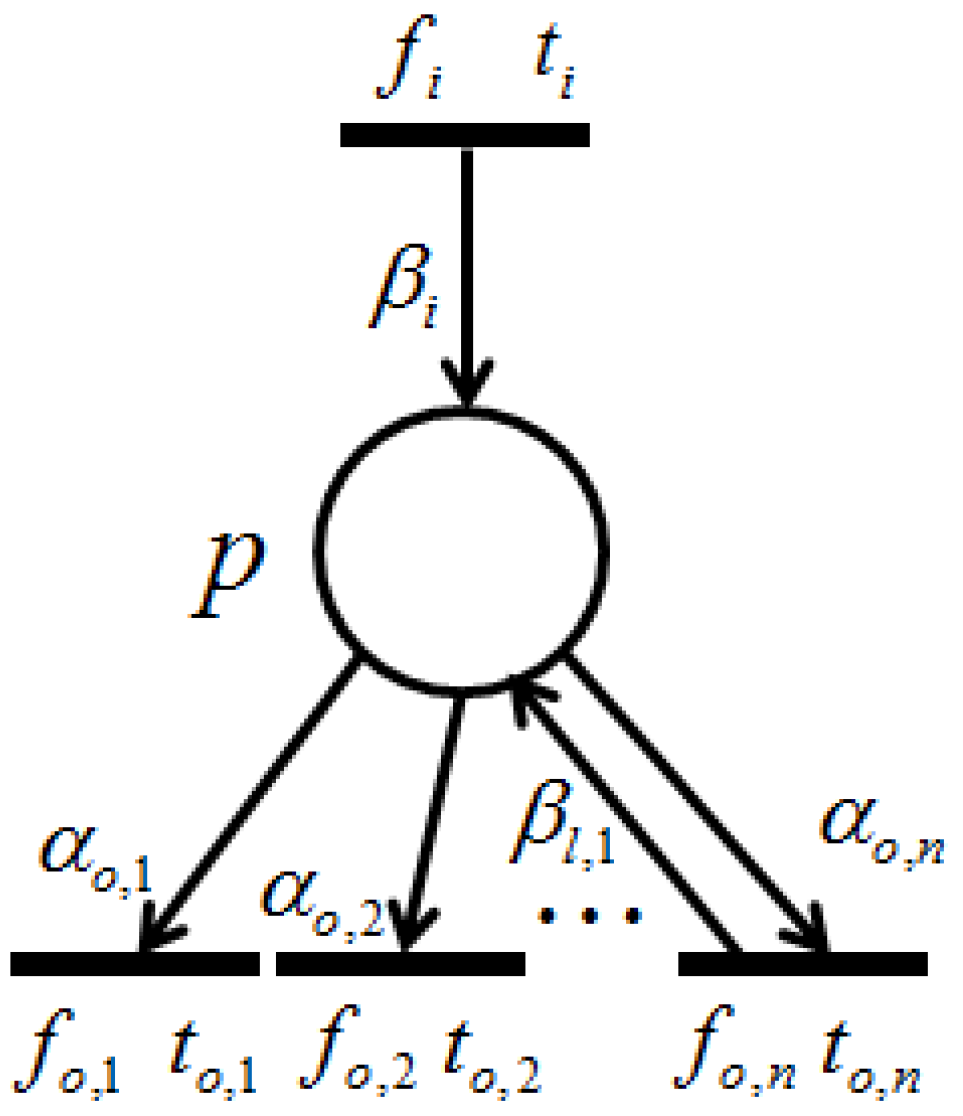
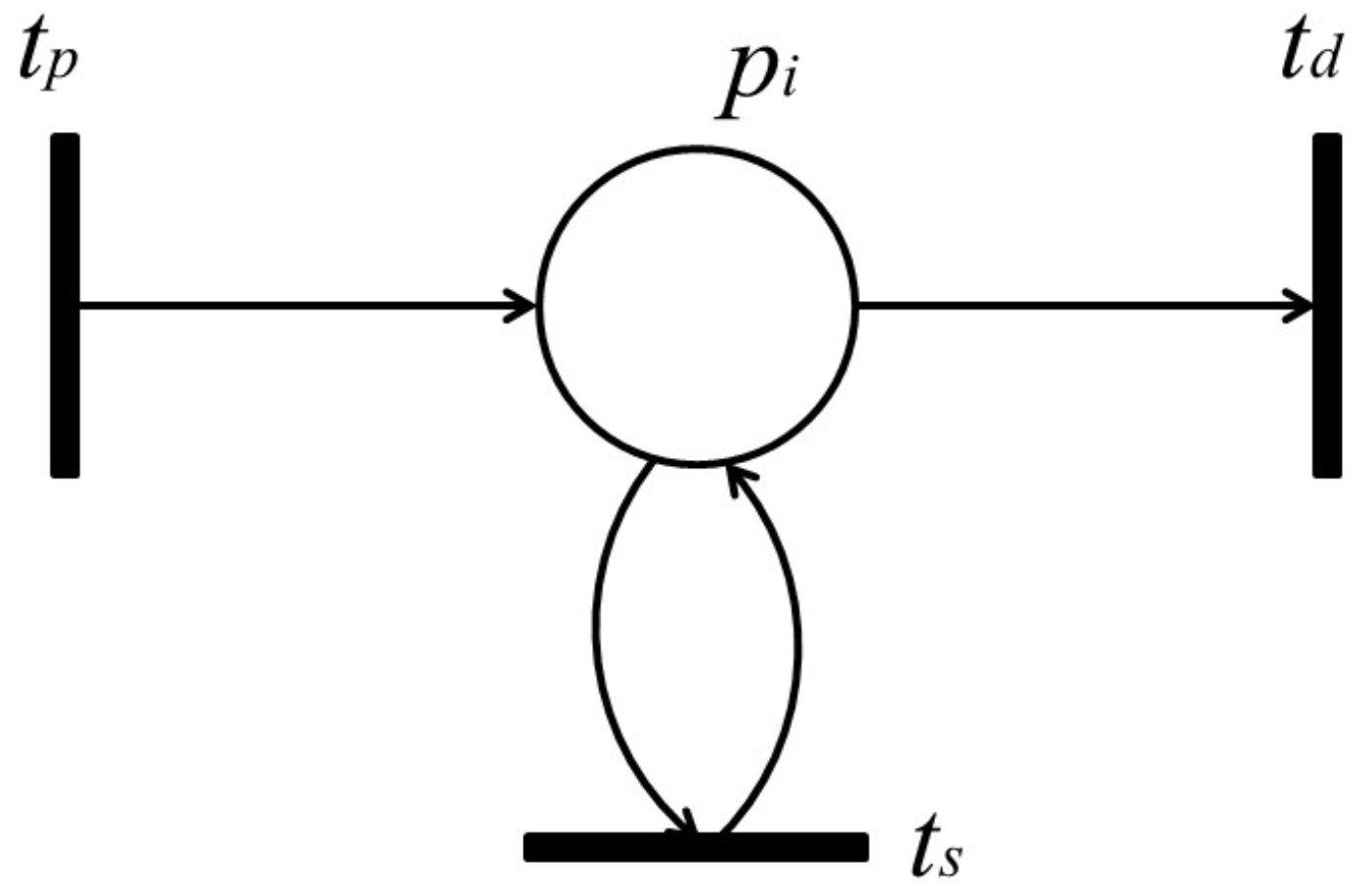
With the firing of transition , token amounts flowing into place p per unit of time are called “input token-flow” and are denoted by . On the other hand, with the firing of transition , token amounts flowing out of place p per unit of time are called “output token-flow” and are denoted by . and (shown in Figure 6) are defined by the following equations, respectively:where and are the firing frequencies of and , respectively, and and are the weights of and , respectively. Based on this definition, the following proposition holds.
Proposition 1. [
23]
Let p be a place with input transitions and output transitions . Then, and are the total input token-flow and the total output token-flow for place p, respectively. Furthermore, when firing frequency f is at the maximum firing rate, , input token-flow and output token flow become the maximum, and , respectively. These maximum token-flows satisfy the following equations. The following requirement is trivial.
Proposition 2. [
23]
In a timed Petri net, the total output token-flow is not more than the total input token-flow for each place p: In a signaling pathway, a reaction not only modifies the formation of molecules, as mentioned above, but also controls the amounts of produced substances. Although the amount of a substance will change along the signal propagation, it cannot be increased indefinitely; otherwise, an abnormal accumulation of a substance may cause a crucial problem in the signaling pathway. Based on this observation, we introduced the notion of a “retention-free” pathway for a timed Petri net. In a retention-free Petri net, the total token amounts flowed-in and flowed-out for each place per unit of time are equivalent.
Definition 8. [
23]
A timed Petri net is called a retention-free Petri net () (satisfying Proposition 1) if the total input token-flow and total output token-flow are equivalent at any place of , that is, In fact, there exists at least one T-invariant in an
, which indicates a firing counting vector of transitions in a periodic firing sequence [
26] that leads back to the marking where it starts. This is because actually, the firing frequencies of all of the transitions construct a T-invariant.
4. Dependency Relation and Dependent Shrink
The firing frequency of a certain transition can be calculated if the firing frequencies of the transitions attached to that transition have been determined thus far, i.e., when these transitions are dependent on each other with respect to the firing frequencies. To provide a clear and precise definition of this dependency of the transitions, we introduce a new concept called a “dependency relation” for transitions, which is formalized based on the retention-free concept presented in the previous section. After that, we propose a method for shrinking a Petri net subnet, induced by the transitions satisfying a dependency relation, into a single transition. In other words, if a subnet can be shrunk, then the firing frequency of any transition in the subnet can be deduced based on the firing frequency of a certain transition inside the subnet.
4.1. Dependency Relation
Definition 9. Let be a timed Petri net and and be two transitions. Here, and are said to be related to each other if and only if the following equation holds:where and are the firing frequencies of and , and is a positive rational. This is called a dependency relation and is denoted by . The above equation means if two transitions are related to each other, then firing frequencies are dependent. Based on this definition, we have the following propositions.
Theorem 1. A dependency relation is an equivalence relation.
Proof. We only need to prove that a dependency relation is reflexive, symmetric and transitive. Let t, , and be transitions of a Petri net. A dependency relation is clearly reflexive and symmetric because (1) holds and (2) if , then , and vice versa. Suppose and hold, then , which means the dependency relation is transitive. ☐
Based on the Definition 9, we have the following proposition for a retention-free Petri net.
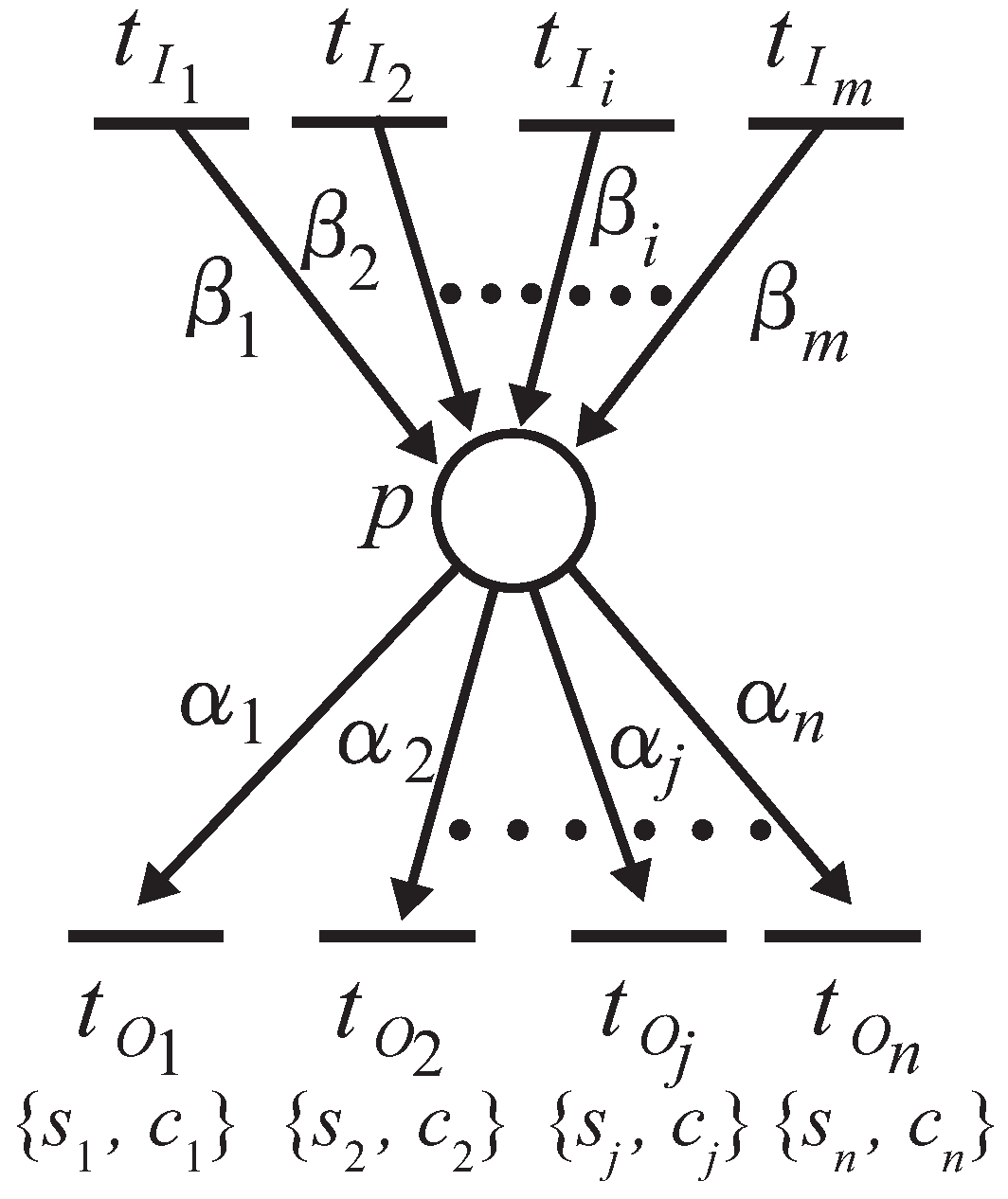
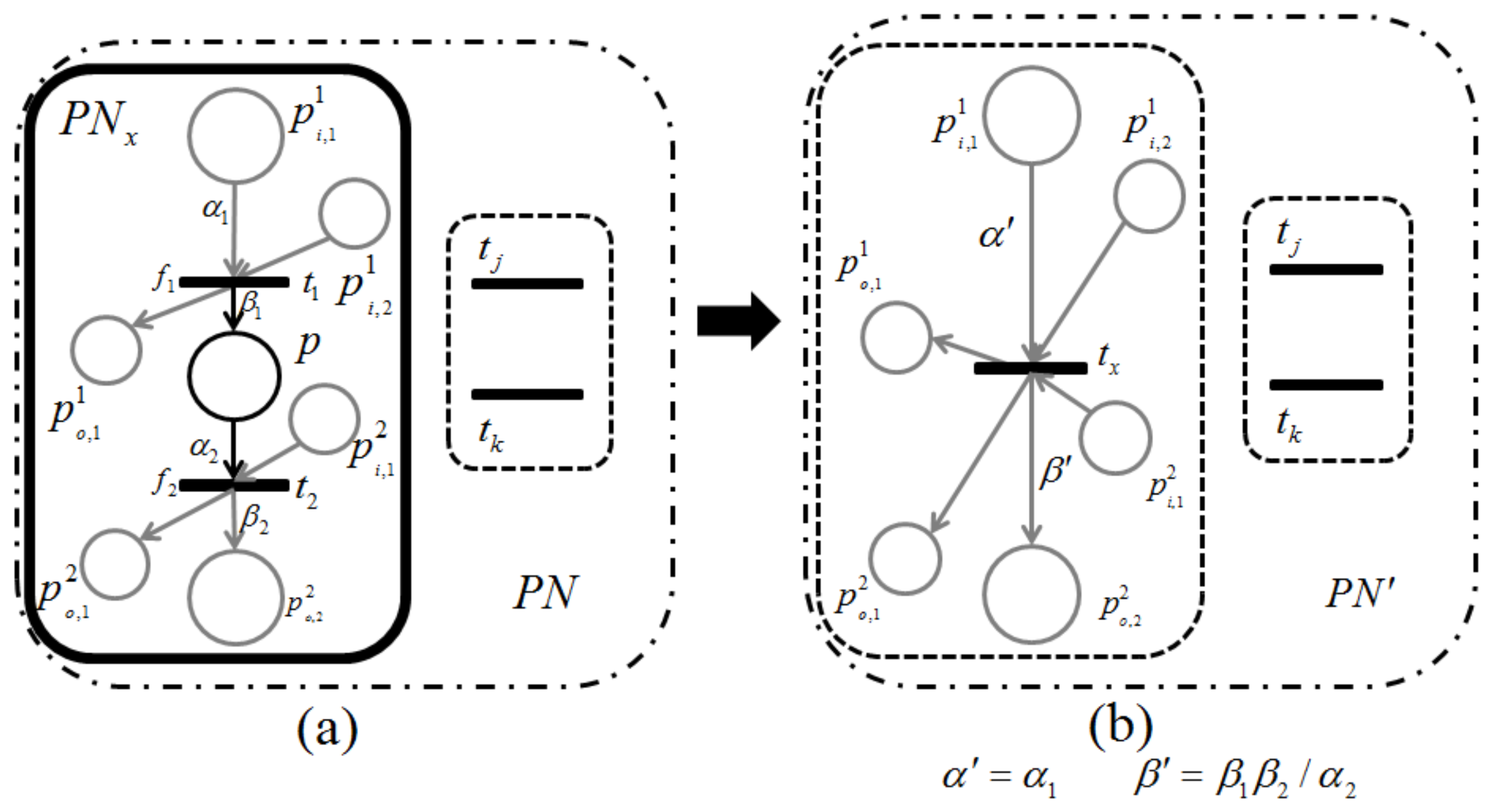
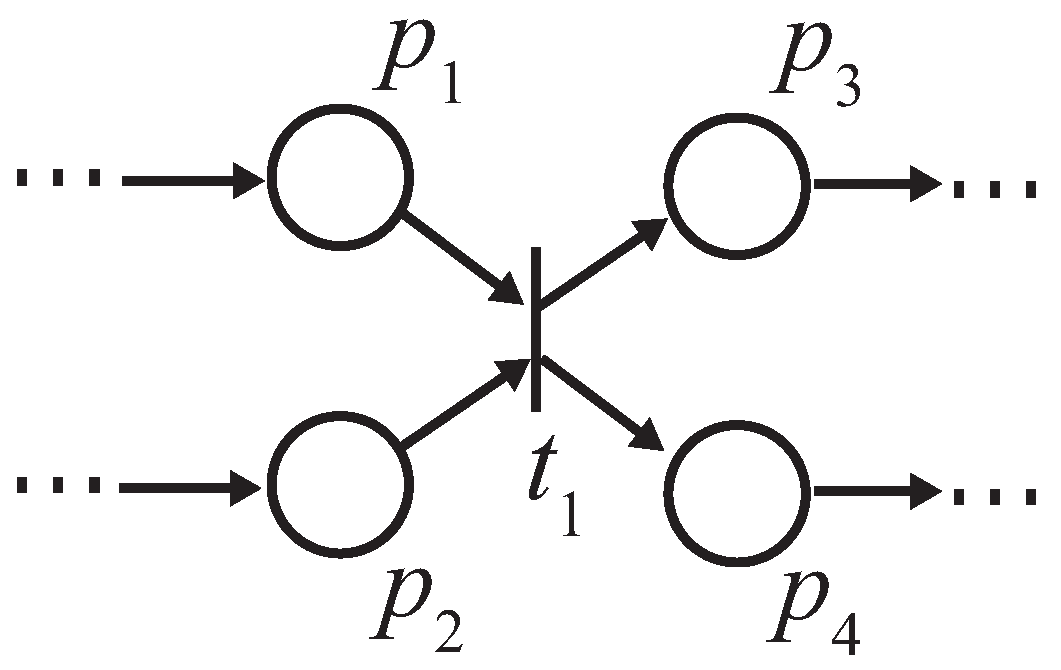
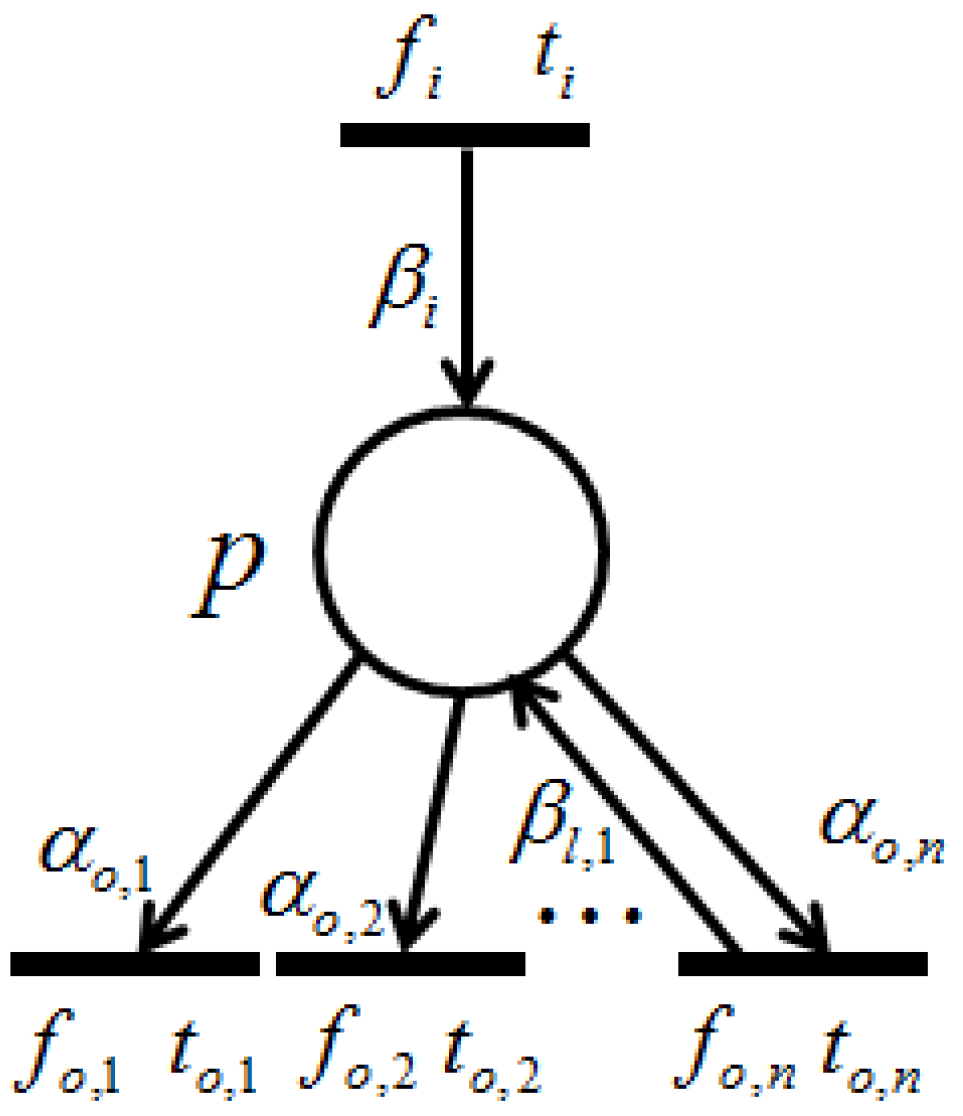
Proposition 3. Let and be two retention-free nets consisting of a place with single input and single output transitions (called a single-input and single-output structure), and a conflict place with multiple transitions (called a conflict structure), respectively. (i) In , the input transition and output transition satisfy a dependency relation; (ii) in , all output transitions satisfy their dependency relation with one another.
Proof. (i) Suppose
is a net, as shown in
Figure 7. Because
Figure 7 is retention free, the amount of input and output token flows through place
p must be equal, i.e.,
. Thus, the firing frequency
can be expressed as
. (ii) Without a loss of generality, herein we suppose place
p possesses a single input transition (because we only need to consider the dependency relation of the output transitions), as shown in
Figure 8.
Since the given firing probabilities of all output transitions,
, satisfy:
the firing frequencies,
,
, ⋯,
, can be expressed through the following equations:
which means that all output transitions satisfy the dependency relations with one another. ☐
The two nets shown above are elementary structures of , which will be applied in the following discussion, as well as in our algorithm.
As shown in Theorem 1 above, a dependency relation is an equivalence relation. Hence, the transition set of a timed Petri net can be divided into independent subsets, i.e., equivalence classes. In the following subsection, to indicate a block of a retention-free Petri net, in which all transitions satisfy a dependency relation, we provide a definition of “dependent shrink”.
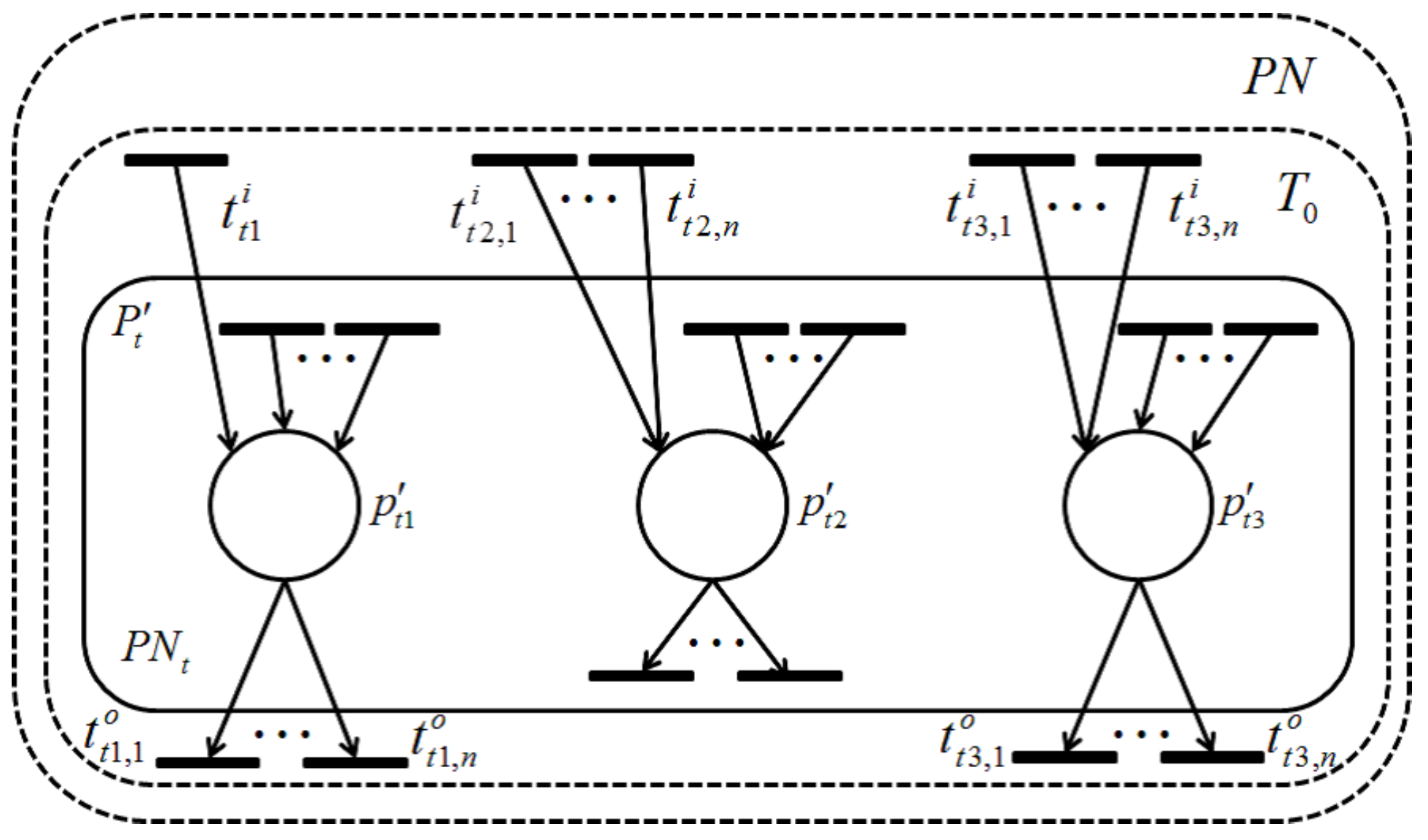
4.2. Dependent Shrink
Definition 10. Let be a retention-free Petri net.- (i)
Let be a subset of T, where x () is a positive integer, and all of the transitions in satisfy a dependency relation. A Petri net induced by is defined as , where , and .
- (ii)
The transformation of by constructing all transitions of and any place satisfying into a single transition, denoted by , is called a dependent shrink. In addition, the resultant net transformed from is called a dependent shrunk net, where, , , , α and β of ’s input and output arcs are decided, such that the input and output token flows of the places, included in , are kept equivalent before and after shrinking.
The following lemma shows us that the dependency relation of the transitions does not change before and after a dependent shrink, i.e., if two transitions are out of the dependency relation, then they will not satisfy the dependency relation after a dependent shrink; and vice versa.
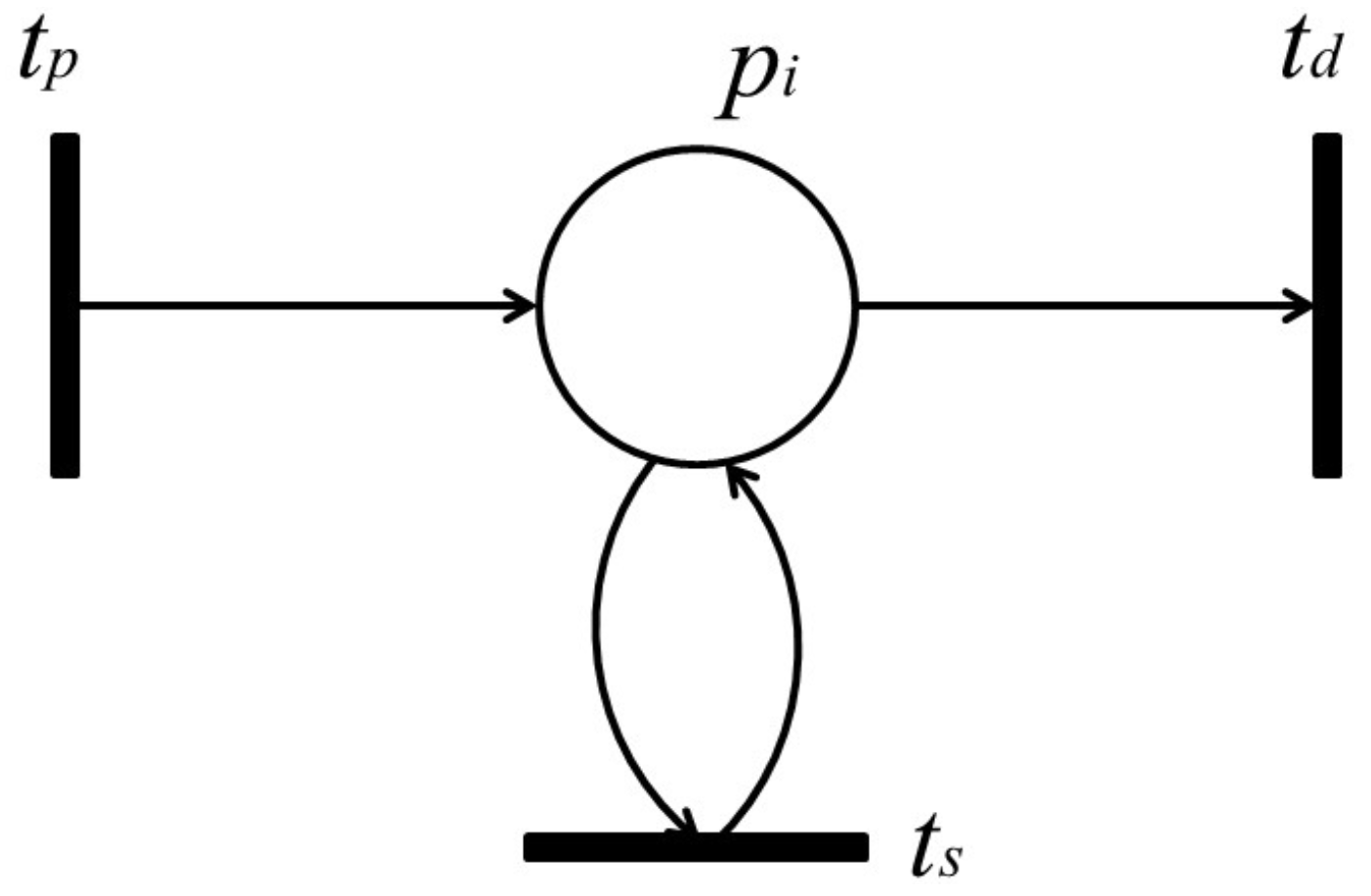
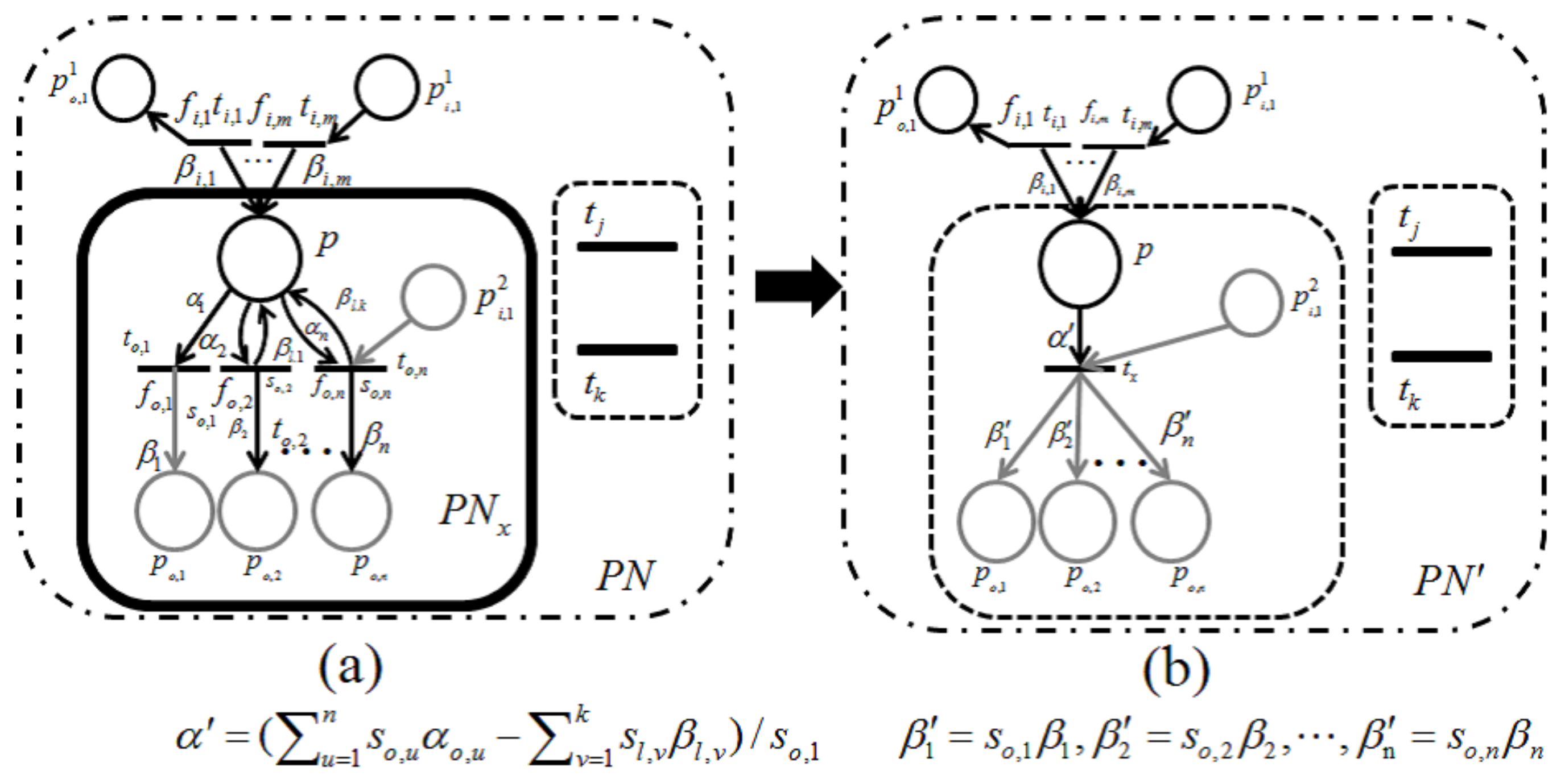
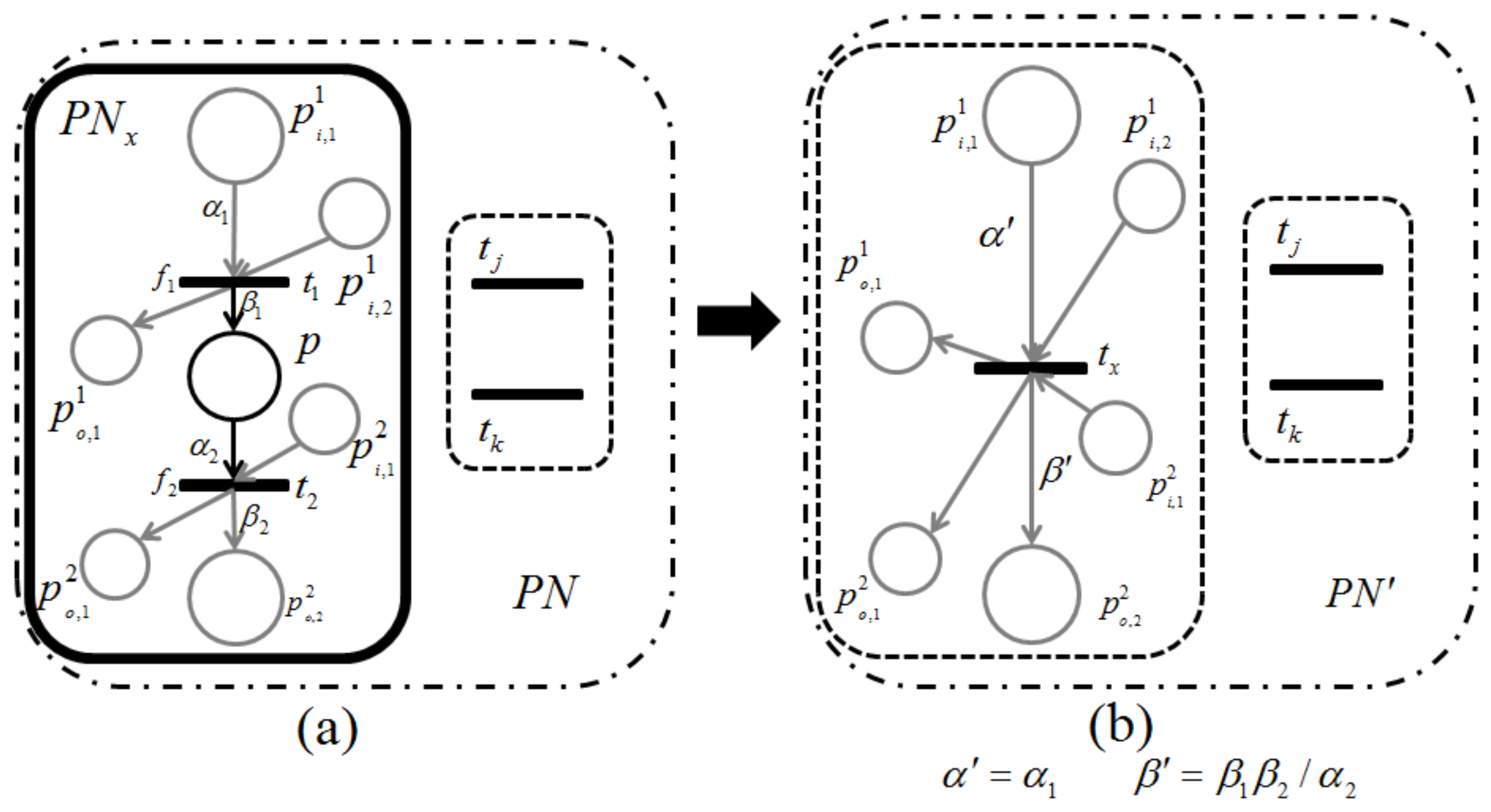
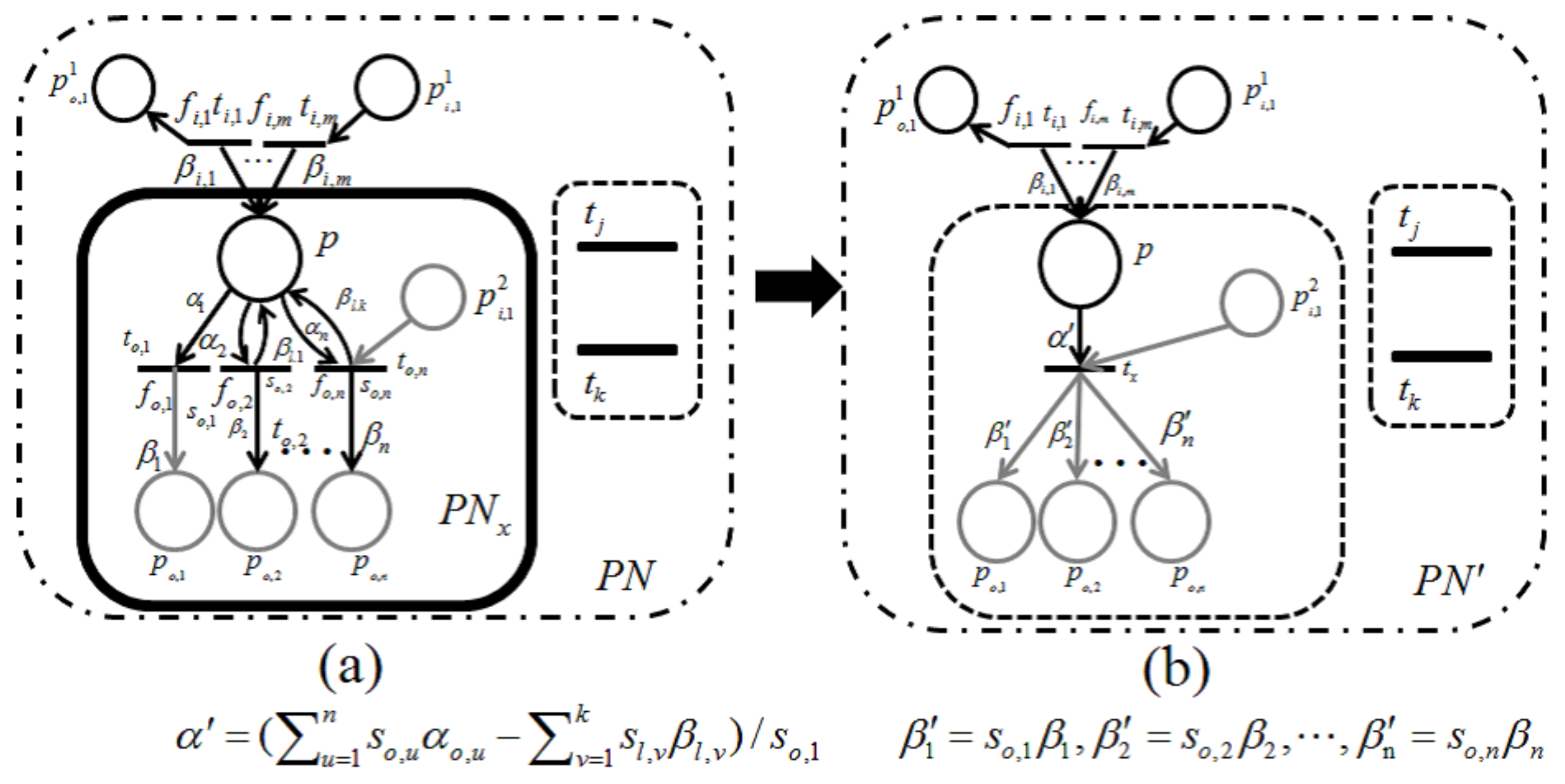
Lemma 1. Let be a retention-free net including a subnet of Figure 9a or Figure 10a and be its dependent shrunk net by applying a dependent shrink for , where . Suppose and ; then, the following claims hold.- Claim 1:
If in , then in ; otherwise, in .
- Claim 2:
If in , then in ; otherwise, in .
Proof. Recall that a single-input and single-output structure (
Figure 7) and a conflict structure (
Figure 8) are the elementary structures of
considered in this paper.
s surrounded by the bold line in
Figure 9a and
Figure 10a are basically the same as in
Figure 7 and
Figure 8, respectively, but these are extended by attaching additional places and arcs (pale parts) in order to make them more general. In addition, two transitions,
and
, are appended for the proof of Lemma 1 below.
For the case of
Figure 9,
p and its input and output transitions (
and
) are shrunk into a single transition,
, and the weights of the new input and output arcs are given as
and
, respectively. On the other hand, for the case of
Figure 10, all output transitions of place
p are shrunk into a single transition,
, and the weights of the new input and output arcs are set to:
Here,
should be satisfied; otherwise,
is not a retention-free Petri net. For both of these two cases, the moderated tokens on
and
arising from the firings of
and
of
Figure 9a and the firings of
of
Figure 10a have no changes before or after shrinking
; that is, the dependency relation between
and
is not changed, and Claim 1 therefore holds.
Let us see Claim 2.
should be the input or output transition of
p in
Figure 9 and one of the output transitions of
p in
Figure 10. Similar to the proof of Claim 1, the moderated tokens on
and
arising from the firings of
and
of
Figure 9a and the firings of
of
Figure 10a have no changes before or after shrinking
. Hence, if
(or
) in
, then
(or
) in
; oppositely, if
(or
) in
, then
(or
) is related to any transition of
Figure 9 or
Figure 10. Therefore, this lemma holds. ☐
The changes of weights given in the proof of Lemma 1 are also applied in our proposed algorithm.
In the following subsection, the uniqueness of a dependent shrink is proven. This means that if a retention-free Petri net Q can be transformed into a dependent shrunk net, the order of transitions in Q for the transformation does not affect the final resultant net.
4.3. Uniqueness on Dependent Shrink
In this section, we introduce the concept of a dependent transition set that is a maximum set of transitions with a dependency relation and discuss the uniqueness of the resultant net through dependent shrink for retention-free Petri nets.
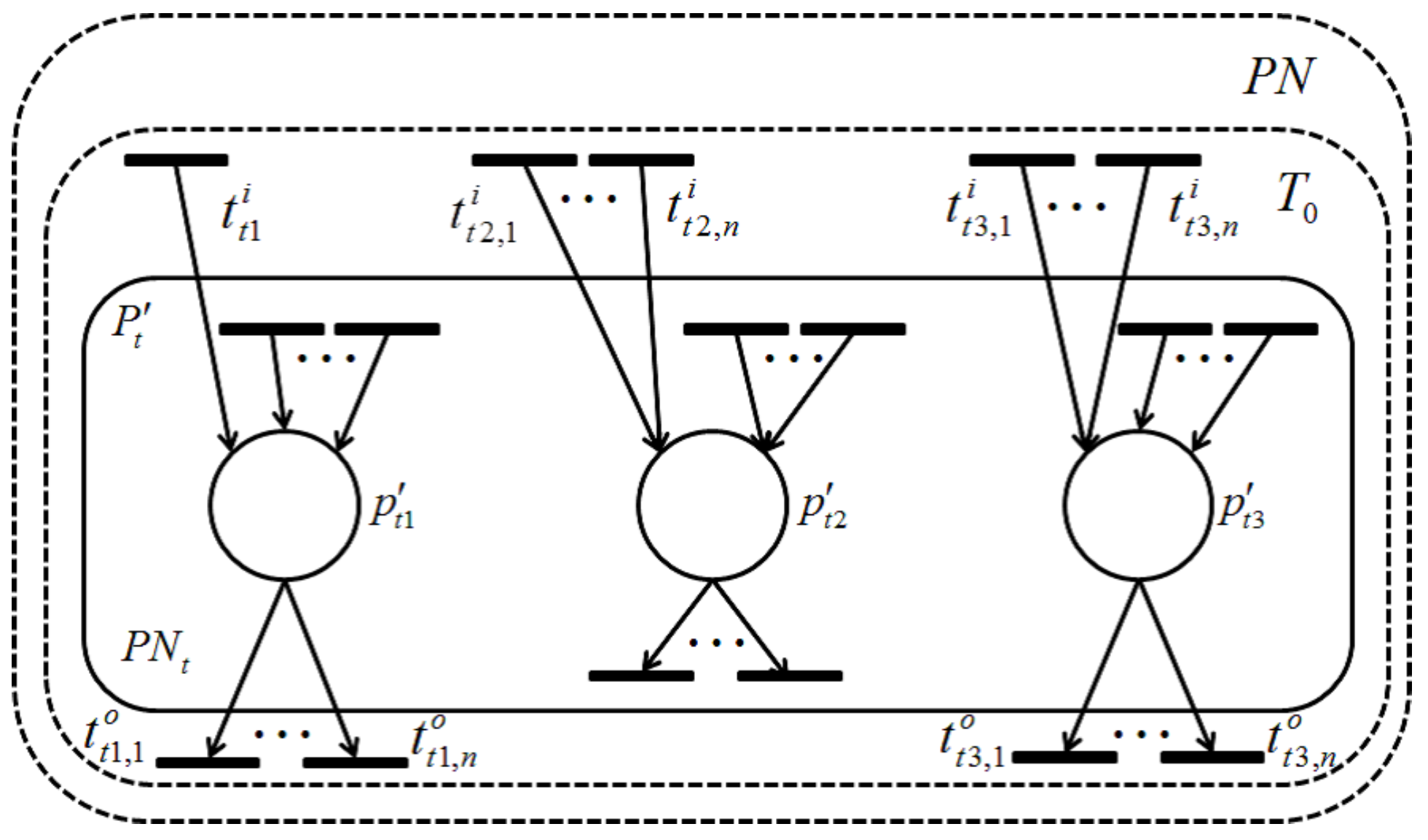
Definition 11. Let be a retention-free net, t be a transition in T and be a set of all transitions related to t.- (i)
Such a is called a t-dependent transition set.
- (ii)
A Petri net induced by is called a t-dependent subnet and is denoted by .
Note that, differing from
defined in Definition 10, the transitions included in
are not related to any transitions of
T –
owing to the fact that
is a maximum set of transitions with a dependency relation (shown in
Figure 11). The following lemma is immediate.
Lemma 2. Let and be the transition sets of t-dependent and t’-dependent, respectively. If , then .
Lemma 3. Let be a t-dependent transition set and be its t-dependent subnet of a retention-free Petri net . Then, is a connected subnet.
Proof. Assume that is not connected, i.e., consists of at least two divided nets, and . Let us see and its connection to the other part of . There exists a subset of places that includes all places connecting with the outside transitions (not included in ), for instance, . Thus, these transitions of are the only ones that connect to other parts of , and further, they are not related to any one of ( for and ); otherwise, . Because and are not connected, any transition of connecting to a transition of should pass through at least one of , which implies that the transitions of are not related to , and thus, this contradicts that is a t-dependent transition set. ☐
Theorem 2. Let t be a transition of a retention-free Petri net , be a t-dependent transition set, be a t-dependent subnet, be a net by shrinking and be a shrunken transition of . For , and are satisfied in . Further, if in , then in , and vice versa.
Proof. There exists a subset
of places that includes all places connecting with the outside transitions (not included in
), for example,
. Thus, these transitions of
are only those that connect
to other parts of
, and further, they are not related to any of
(
for
and
); otherwise,
. After shrinking
,
and
remain in
, and of course,
holds in
for
. Further, any transition
included in
connecting to
should pass through at least one of
, which means that
and
hold for
. Furthermore, shrinking
does not destroy the dependency relation for any transitions of
, as can be seen in
Figure 12 and
Figure 13 and, thus, for the transitions of
. Therefore, this theorem holds. ☐
According to Theorem 2, if we find all dependent subnets and shrink them individually into single transitions, then all of these transitions are representative of the equivalence classes because a dependency relation is an equivalence relation. The following corollary is immediate.
Corollary 1. Suppose a retention-free Petri net consists of k subnets, , each of which is a dependent subnet. Not depending on the order of dependent shrink for , a unique structure of a resultant net consisting of k transitions can be obtained.
From Lemma 1 and Theorem 2, we can simply obtain the following result.
Corollary 2. Iterating a dependent shrink for the cases shown in Figure 9 and Figure 10, a unique resultant net can be obtained. 5. Dependent Shrink Algorithm and Case Study
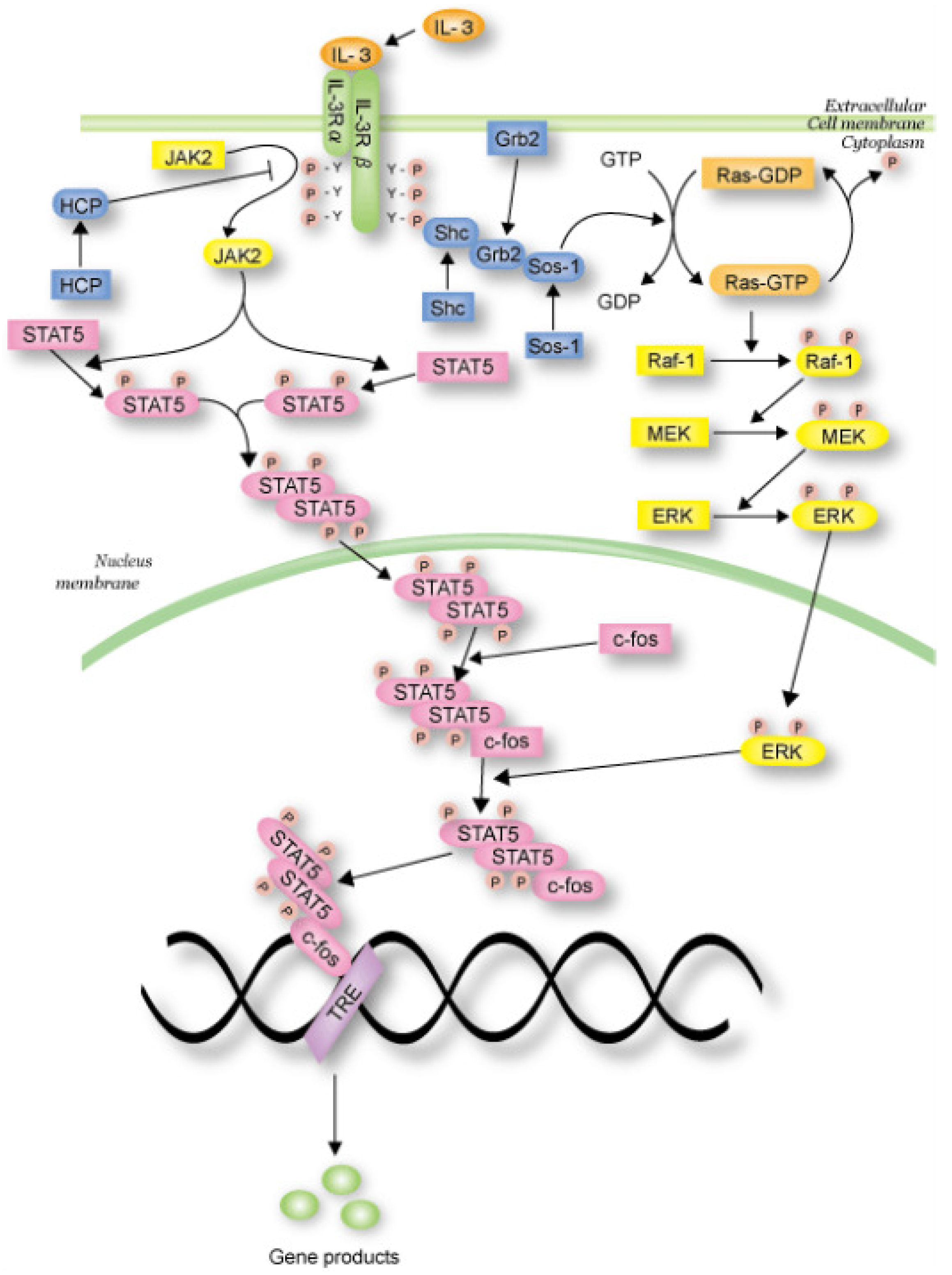
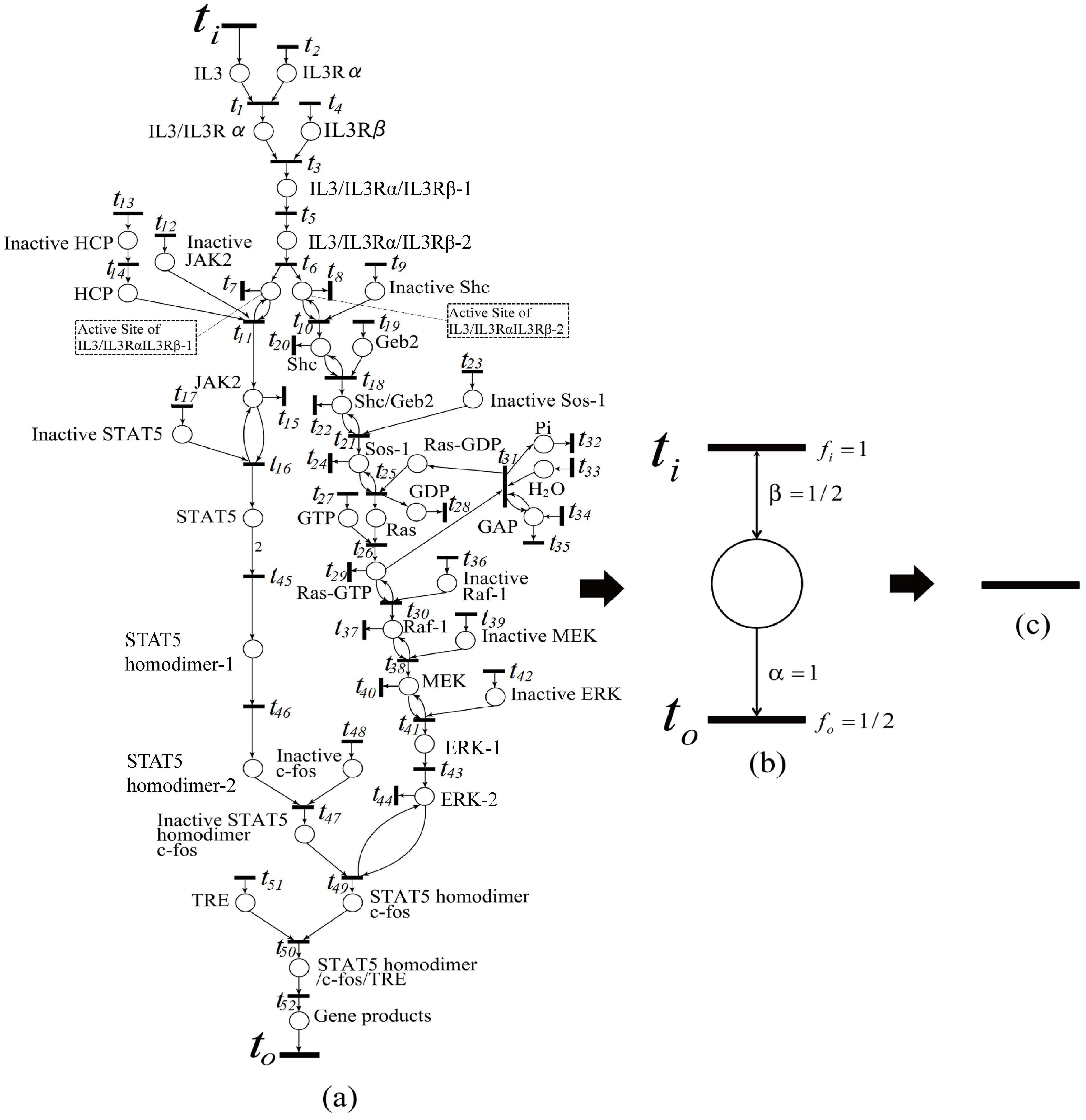
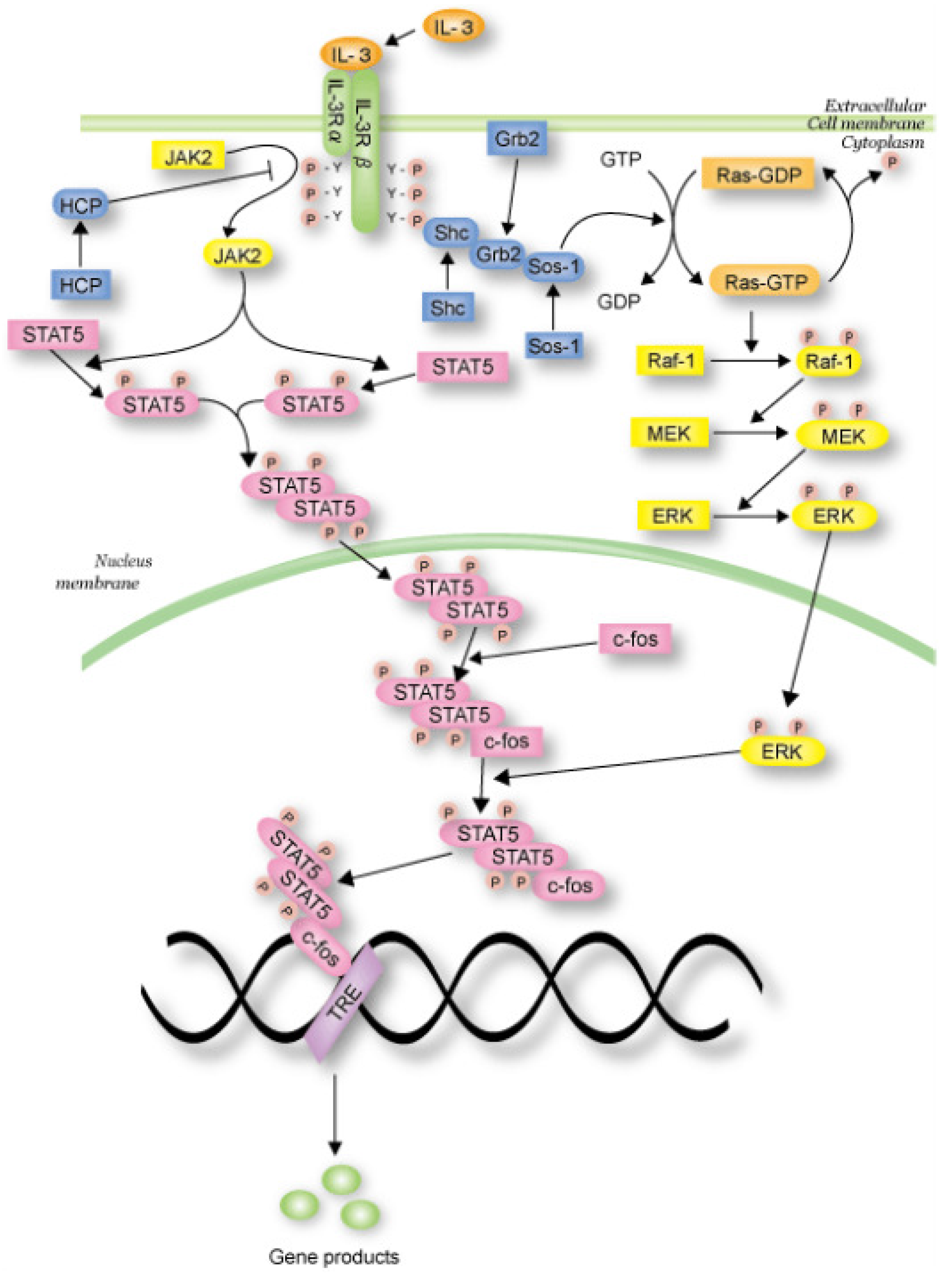
In this section, we propose the dependent shrink algorithm based on the discussion in the last section and apply this algorithm to the IL-3 signaling pathway Petri net model (shown in
Figure 15), which is transformed from the IL-3 phenomenon model (shown in
Figure 14) obtained from the website [
27], as a case study. Note that IL-3 is a glycoprotein and is known to be involved in the immune response [
28,
29,
30].
5.1. Outline of Shrink Process
The outline of the dependent shrink process can be briefly described as follows:
- Step 1:
Shrinking of self-loop structure:
A place randomly selected from a Petri net is stored in a queue after the conversion of the self-loops and the structures of conflict (as shown in
Figure 10).
- Step 2:
Shrink of conflict structure:
If a place picked up from the queue has a self-loop or a transition of one-input and one-output (as shown in
Figure 9), this place is shrunk.
- Step 3:
Changing weight of the input arc:
If a shrunk Petri net has a place with multiple input transitions, then enqueue and continue to Step 2. This procedure is repeated until the queue becomes empty or the number of places with multiple input transitions equals the length of the queue.
The variables used in the algorithm are as follows:
is a given signaling pathway Petri net model constituted by .
N is a variable that stores the Petri net after dependent shrink, constituted by and E.
Q is a queue.
X is a set of places initially set as a given place set, .
f is a flag by which a dependent shrink pattern is determined.
c is a counter, which counts the number of places in Q with multiple input transitions.
5.2. Dependent Shrink Algorithm
The following algorithm is used to repeatedly shrink the structures of
Figure 9 and
Figure 10 to find transitions with an interdependent firing frequency.
| Algorithm 1 Dependent Shrink |
| Input: |
| Output: Shrunk Petri net |
| Main |
| , , , |
| , , |
| while () |
| Pull an element x from |
| Enqueue() |
| Shrink1() |
| Shrink2() |
| Shrink1() |
| if () then |
| |
| Arcweight |
| if () then |
| |
| Arcweight() |
| Shrink2() |
| while |
| |
| if () then |
| |
| Enqueue |
| |
| else if () then |
| |
| |
| else if () then |
| |
| Enqueue(Q, x) |
| |
| if () then |
| Arcweight() |
| if () then |
| Break |
| Arcweight() |
| if () then |
| |
| |
| if ( then |
| |
| |
| else if () then |
| |
| else if () then |
| |
| else if () then |
| |
| |
| Choose . |
| |
| |
| |
| |
| |
| |
| |
| |
| else if () then |
| |
| Let be , (due to ). |
| |
| |
| |
| |
| |
| |
| |
| |
| |
| |
| |
The Algorithm 1 will terminate when all places are shrunk, i.e., a single transition remains, or becomes ones with multiple input transitions and a single output transition. The time complexity is . As a result, given with a retention-free Petri net, this algorithm gives a smaller shrunk net, in which a transition may represent a class of ones with a dependency relation.
5.3. Case Study
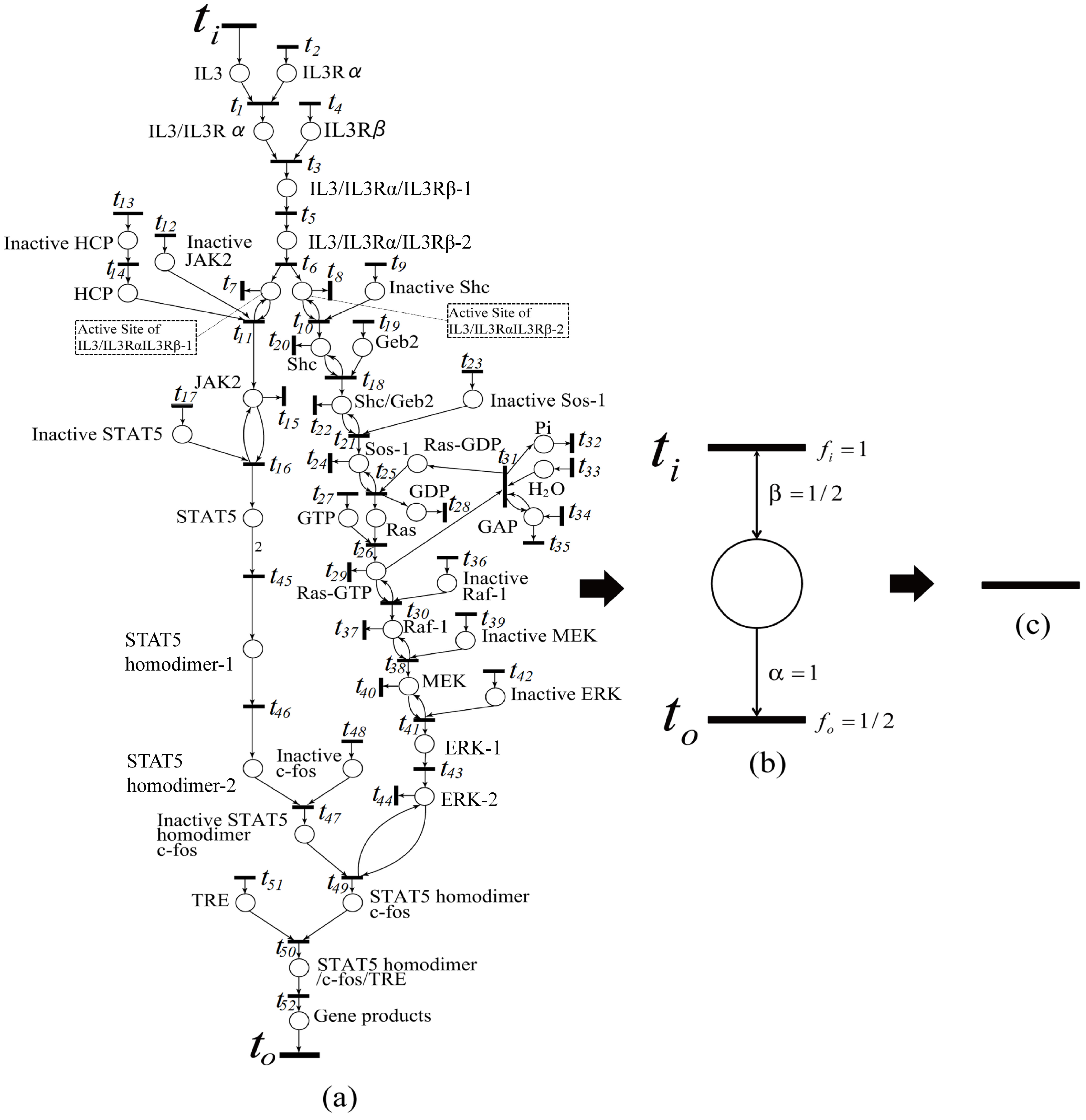
Here, we give an example showing an application of our proposed algorithm. Since the algorithm has not been implemented, here we manually apply it. The algorithm is applied to the IL-3 Petri net model (see
Figure 15a), obtained from the website [
27]. As a result, the original IL-3 Petri net model shown in
Figure 15a is shrunk into
Figure 15c. This means that the firing frequencies of all transitions in
Figure 15a are dependent on one another. In the intermediate shrunk net (see
Figure 15b), by assuming that the firing frequency
of the input transition
is one, and the weights of the input and output arcs are
and one, respectively, then the firing frequency
of output transition
is
. In this way, we can calculate all firing frequencies in the IL-3 Petri net model if the firing frequency of one transition is known in this model.
That is, all of the reaction speed data of the IL-3 Petri net model are dependent on one another, and thus, if one reaction speed datum is available, all of the others can be derived from it.
We have two ways to determine reaction speed data in biological pathway, i.e., firing frequencies. One is to find firing frequency relationships between transitions from the intermediate shrunk nets generated during the execution of the Algorithm 1, in turn from the final net to the original net.
In the intermediate shrunk net (see
Figure 15b), by assuming the firing frequency
of the input transition
to be available (say one), then the firing frequency
of output transition
is
. Recursively, we can derive all of the firing frequencies in the IL-3 Petri net model.
Another way is to establish a series of linear equations for all of the places according to retention-freeness, Equation (
8), and to solve these equations to finally get the solutions with the form
. Here,
are the firing frequencies corresponding to non-valid and valid reaction speed data.
Suppose
is only one transition whose corresponding speed data are valid; we can derive the firing frequencies for all of the other transitions of the IL-3 Petri net model in any one of two ways as follows:
In addition to IL-3, we also apply our algorithm to the endocytosis Petri net model and give the shrunk result as can be found in the
Supplementary Materials, in which, IL-3 and endocytosis respectively consist of 43 places, 54 transitions and 109 arcs and 24 places, 34 transitions and 70 arcs. Moreover, the System Biology Markup Language (SBML) [
31] and Petri Net Markup Language (PNML) [
32] files of those Petri net models can also be obtained from the
Supplementary Materials.
5.4. Discussion on the Resultant Structure from the Algorithm
As mentioned, when the Algorithm 1 terminates, there are two possible cases: (1) the resultant net is a single transition; (2) the resultant net is such that all places possess a single output transition and multiple input transitions.
Case (1) indicates that all transitions of the given net satisfy a dependency relation, and they comprise an equivalence class, that is, if the firing frequency of any one of the transitions is decided, then the frequencies of all of the others are automatically decided as well.
Case (2) may be further divided into: (i) the remaining net consisting of only such transitions that do not satisfy dependency relations with one another; and (ii) some of the transitions satisfying a dependency relation, but not having been shrunk into a single transition.
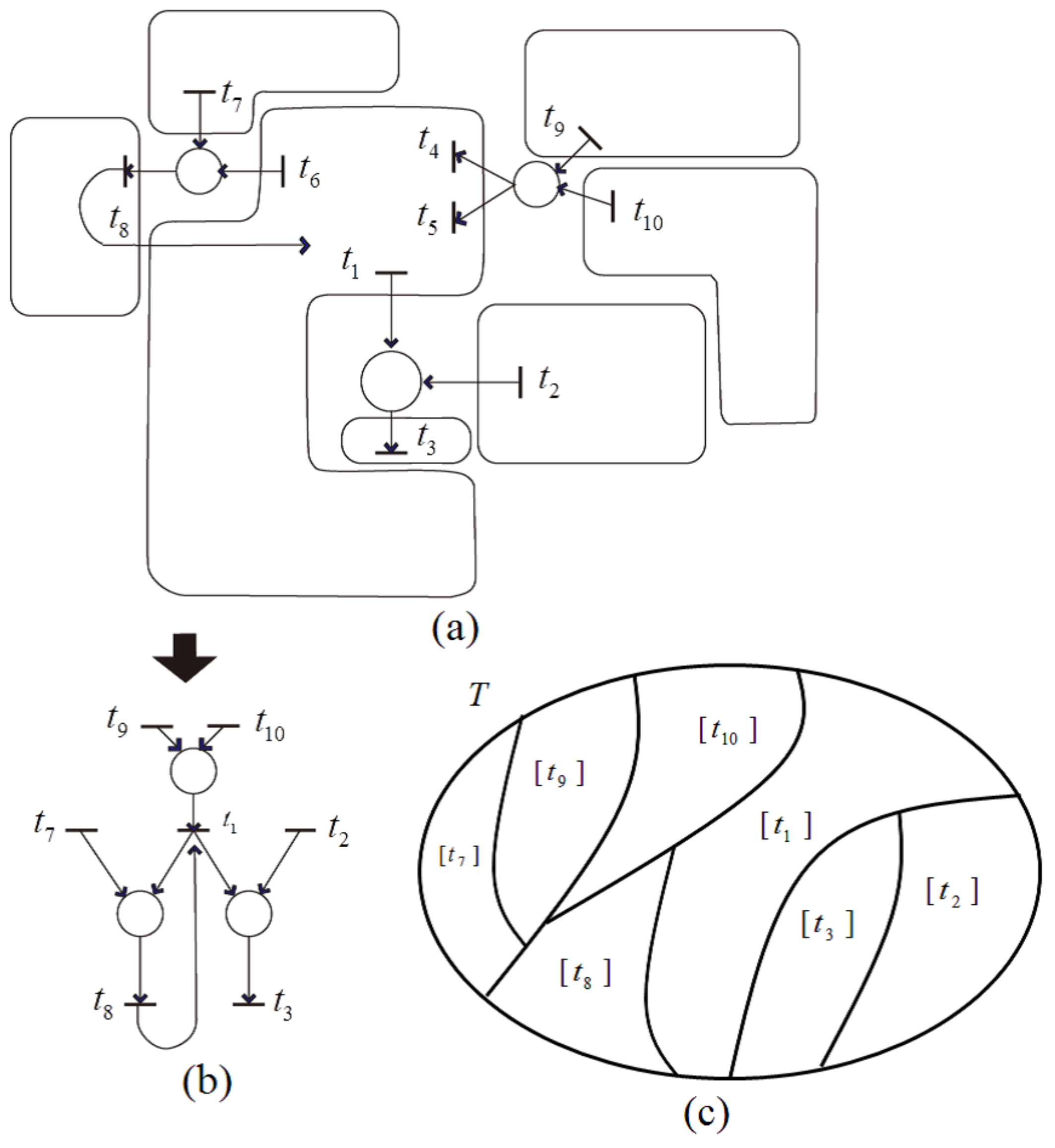
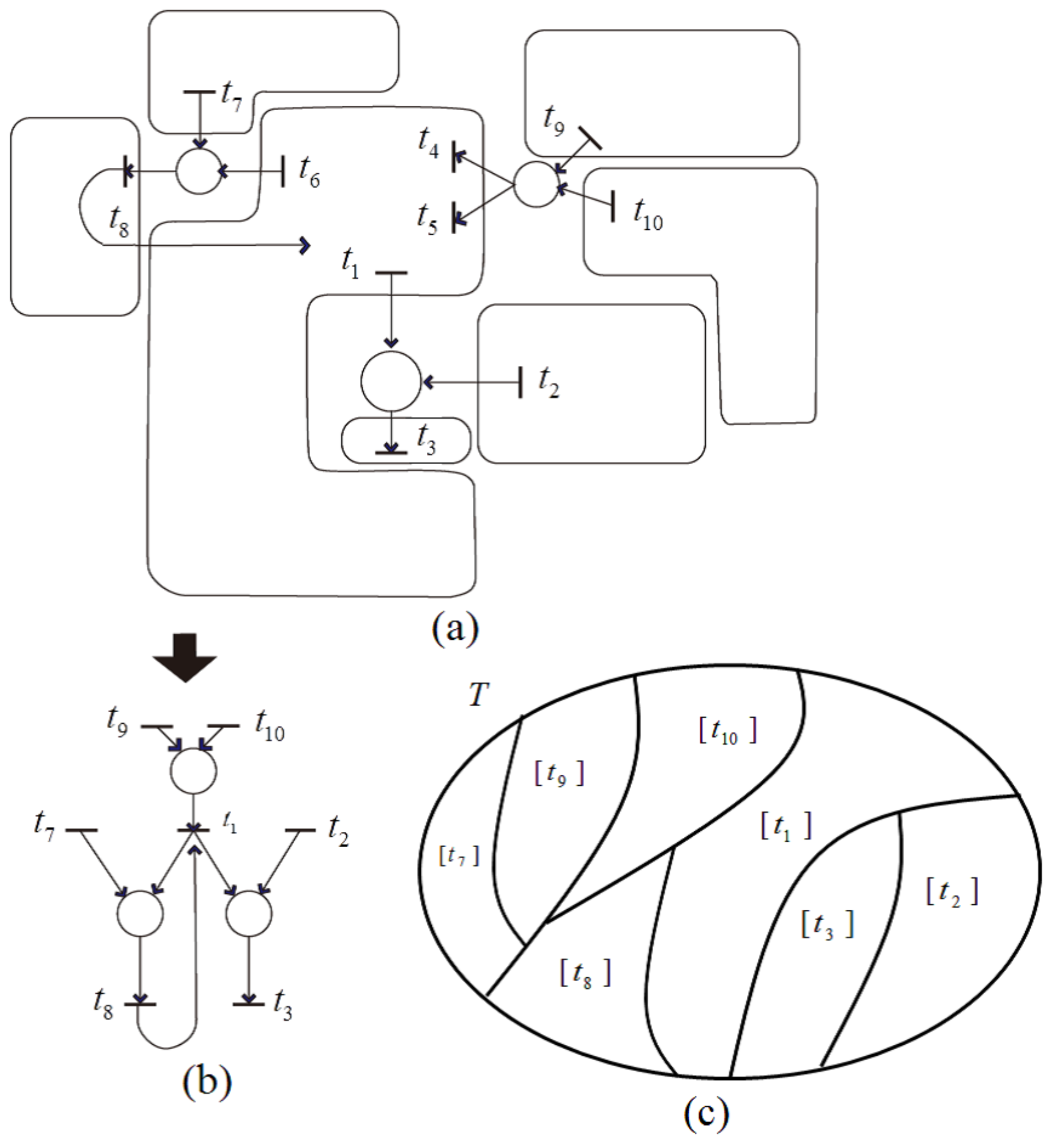
Case (2) (i) indicates that all transitions in the remaining net are independent, and each is a representative of an equivalence class, as can be illustrated in
Figure 16. In this
Figure 16a, each transition set is rounded by a line, and the transitions included in each set satisfy a dependency relation with one another. Applying dependent shrink, the transformed net is obtained as shown in
Figure 16b. Further, the equivalence classes of
represented by
are shown in
Figure 16c.
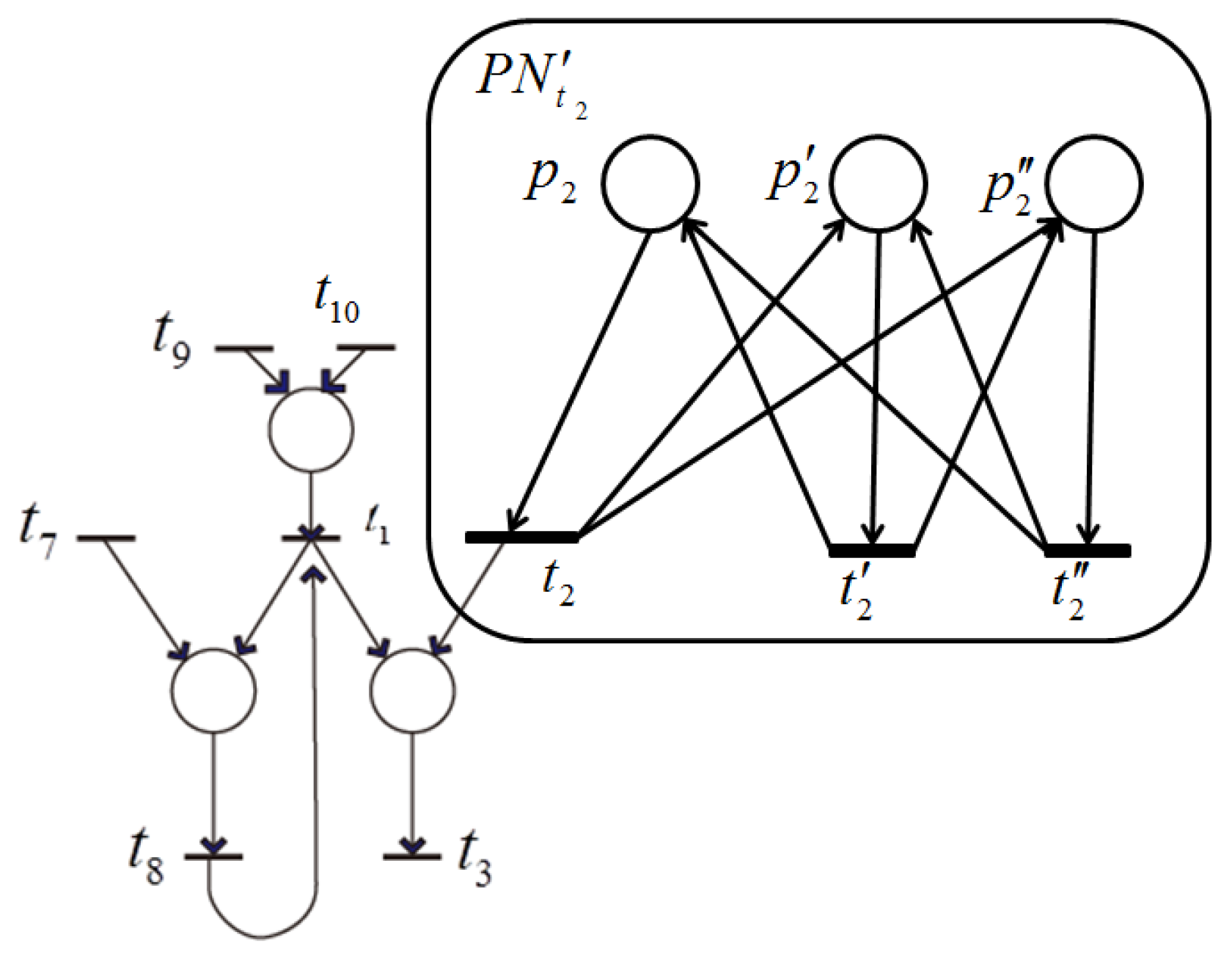
Case (2) (ii) implies that the remaining net includes a subnet that consists of transitions that all have a dependent relation, but cannot be shrunk into a single transition by shrinking the single-input and single-output structure (
Figure 7) or the conflict structure (
Figure 8).
, shown in
Figure 17, is such a subnet, where each transition has at least one input place with one output, which means
in
. To find such a subnet, we should establish a series of linear equations for all places according to the retention-freeness in which the input and output token flows are equal and solve these equations. Because
in
, the number of equations is more than that of the variables (firing frequencies of the transitions). Thus, if there are exactly
linear independent equations, then all of the transitions satisfy a dependency relation. A detailed discussion of this remains as a future issue.
6. Conclusions
Despite the recent rapid progress in high throughput measurements of biological data, it is still difficult to gather all reaction speed data in the biological pathways. Particularly for signaling pathways, owing to their complex structure, including protein formations and enzyme reactions, computational methods are essential to estimate as much reaction data as possible from biologically-valid data. In this paper, we presented an algorithm to shrink the transitions in a timed Petri net model into a smaller number of transitions based on their dependency with respect to the firing frequency and, equally, their amount of delay.
We have first introduced a new concept, called the dependency relation, over a set of transitions of a Petri net model representing a signaling pathway, which is in fact an equivalence relation over transitions. Through a structural analysis, we obtained several properties, one important result of which is that shrinking a connected subnet, induced by transitions with a dependency relation, into a transition never affects other parts of a Petri net. Based on these results, we designed an Algorithm 1 with time complexity to reduce the Petri net size by applying a dependent shrink process for two elementary structures, i.e., a “single-input and single-output structure” and a “conflict structure”, in order to efficiently find a transition set with a dependency relation. A case study was conducted by applying our algorithm to the signaling pathway Petri net model of IL-3, and as a result, IL-3 finally becomes a single transition, which means that all reaction speeds of IL-3 are dependent on one another and that measuring one reaction speed can help in deriving all other speeds.
Our proposed method can be applied not only to signaling pathways, but also to other biological pathways in which retention-freeness holds, for example to metabolic pathways. If, in a timed Petri net model constructed from such a biological pathway, there exists a transition violating retention-freeness, then some discrepancy should happen in the biological pathway, i.e., missing interaction and/or extra interaction, which will be caused by mutations or alternations to the pathway. Such a biological inconsistency is expected to be found with our technique.
We have also pointed out that there may remain some subnets with a dependency relation in the resultant net after applying Algorithm 1 because only a dependent shrink of a “single-input and single-output structure” and a “conflict structure” are adopted. We indicated that this can be resolved by solving a series of linear equations for such a subnet without taking up too much computation time because the size of the subnet should be much smaller compared with the original net. As future work, we are planning to: (1) develop a method to find such a subnet that includes only the places possessing a single-output and multiple-input transitions with a dependency relation; (2) design an algorithm to find the transitions with a dependency relation from the solution of a series of the prior mentioned linear equations; (3) extend our method to a stochastic Petri net by taking into account probabilistic firing delay times.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
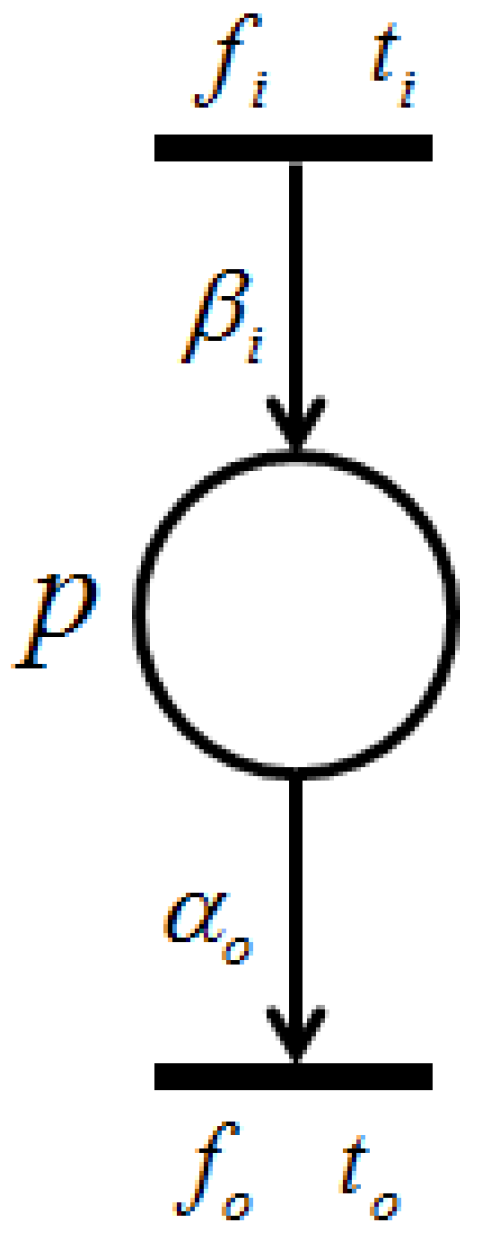{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}