
Repopulating Decellularized Kidney Scaffolds: An Avenue for Ex Vivo Organ Generation



{kind=link}
Abstract
:1. Introduction: Bioartificial Kidneys—An Alternative to Dialysis?
1.1. Renal Functions and the Burden of Kidney Disease
1.2. Bioartificial Kidney Devices
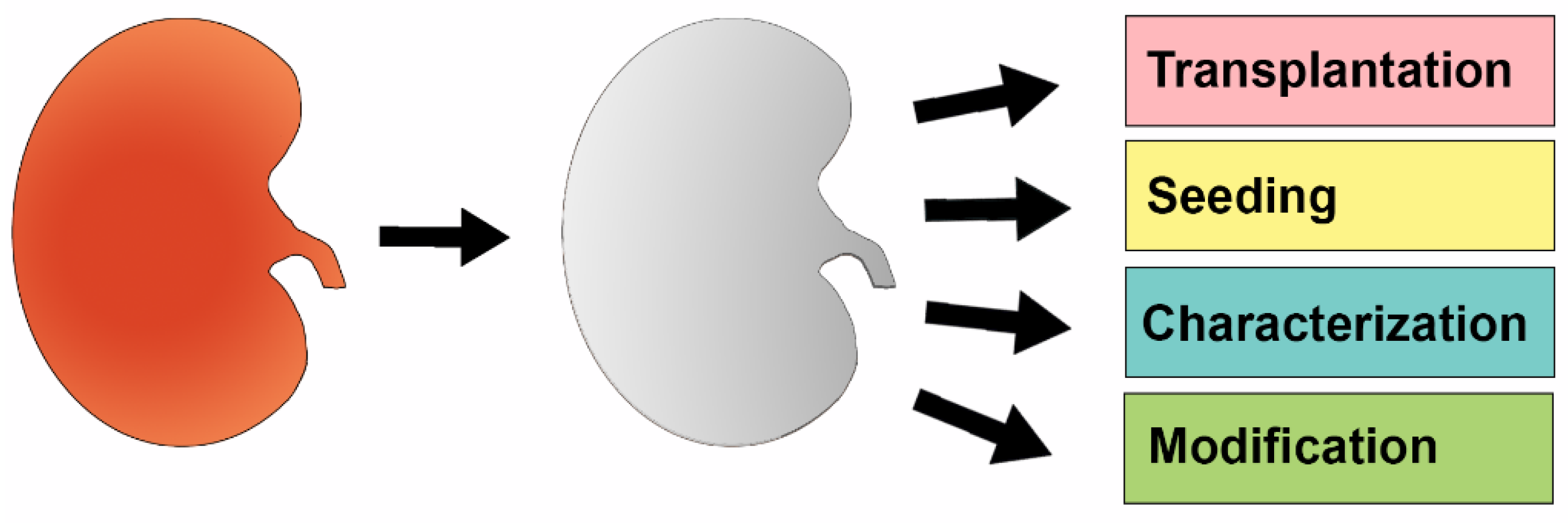
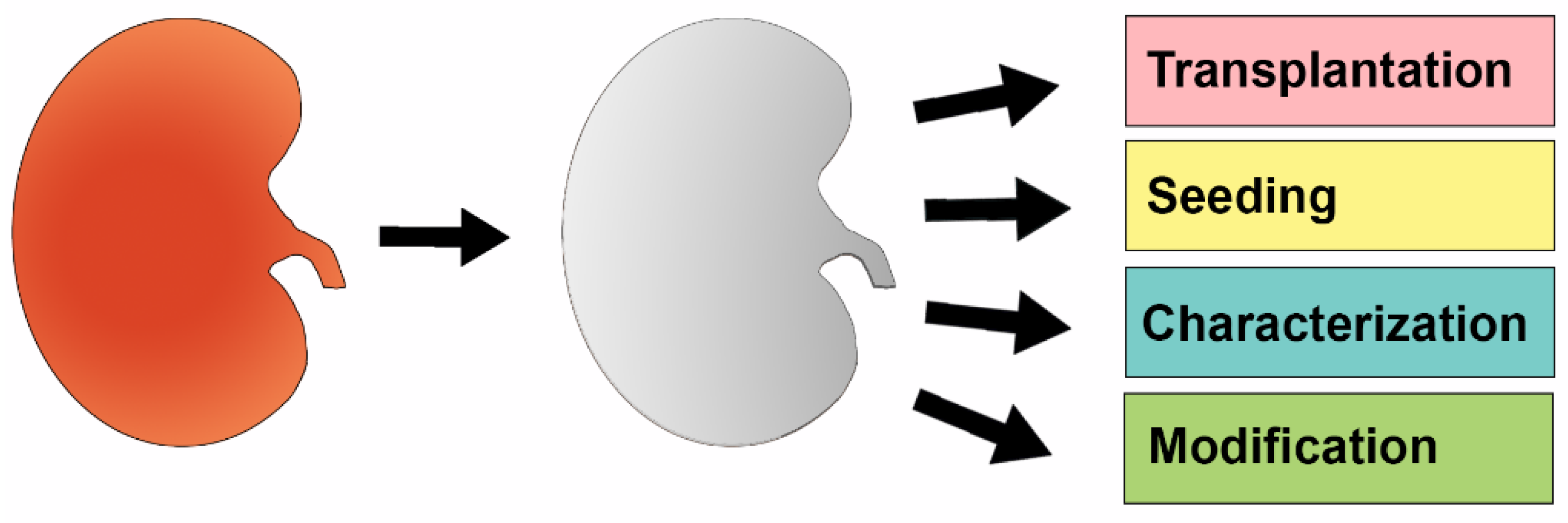
1.3. The Emergence of Techniques to Perform Cellular Seeding of Decellularized Organ Scaffolds
2. Decellularized Kidney Scaffolds—Cadaveric Organs Supporting Life
2.1. Establishment of Basic Parameters for Renal Decellularization and Preservation of the ECM
2.2. Reseeding of Decellularized Kidneys with Various Types of Stem Cells
2.3. Further Optimization of Reseeding the Decellularized Kidney
3. Acellular Environment—Environment Directed Differentiation
3.1. Assessment of ECM Reseeding in Different Kidney Regions: Mouse and Porcine Xenograft Studies
3.2. Assessments of ECM Composition and Xenograft Studies with Mammalian Kidneys and Human Stem Cells
4. Summary and Future Directions for the Nephrology Field
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AKI | acute kidney injury |
| BRECS | bioartifical renal epithelial cell system |
| CKD | chronic kidney disease |
| ECM | extracellular matrix |
| ES | embryonic stem |
| ESRD | end stage renal disease |
| GFP | green fluorescent protein |
| HES | human embryonic stem |
| HUVECs | human umbilical venous endothelial cells |
| ICU | intensive care unit |
| iPS | induced pluripotent stem |
| KMS | kidney-derived microscaffolds |
| MSC | mesenchymal stem cell |
| NKCs | neonatal kidney cells |
| PKD | polycystic kidney disease |
| RAD | renal assist device |
| SDS | sodium dodecyl sulfate |
| sGAG | sulfated glycosaminoglycan |
References
- Chawla, L.S.; Kimmel, P.L. Acute kidney injury and chronic kidney disease. Kidney Int. 2012, 82, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Chawla, L.S.; Amdur, R.L.; Amodeo, S.; Kimmel, P.L.; Palant, C.E. The severity of acute kidney injury predicts profession to chronic kidney disease. Kidney Int. 2011, 79, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Belayev, L.Y.; Palevsky, P.M. The link between acute kidney injury and chronic kidney disease. Curr. Opin. Nephrol. Hypertens. 2014, 23, 149–154. [Google Scholar] [CrossRef] [PubMed]
- McCampbell, K.K.; Wingert, R.A. Renal stem cells: fact or science fiction? Biochem. J. 2012, 444, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wingert, R.A. Regenerative medicine for the kidney: stem cell prospects and challenges. Clin. Tranls. Med. 2013, 2. [Google Scholar] [CrossRef] [PubMed]
- Poureetezadi, S.J.; Wingert, R.A. Congenital and acute kidney disease: translational research insights from zebrafish chemical genetics. Gen. Med. Los Angel 2013, 1. [Google Scholar] [CrossRef] [PubMed]
- Kroeger, P.T., Jr.; Wingert, R.A. Using zebrafish to study podocyte genesis during kidney development and regeneration. Genesis 2014, 52, 771–792. [Google Scholar] [CrossRef] [PubMed]
- McKee, R.A.; Wingert, R.A. Zebrafish renal pathology: Emerging models of acute kidney injury. Curr. Pathobiol. Rep. 2015, 3, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Casino, F.G. Dialysis dose quantification in critically ill patients. G. Ital. Nefrol. 2010, 27, 383–390. [Google Scholar] [PubMed]
- Halloran, P.F. Immunosuppressive drugs for kidney transplantation. N. Engl. J. Med. 2004, 351, 2715–2729. [Google Scholar] [CrossRef] [PubMed]
- Dantal, J.; Soulillou, J.P. Immunosuppressive drugs and the risk of cancer after organ transplantation. N. Eng. J. Med. 2005, 352, 1371–1373. [Google Scholar] [CrossRef] [PubMed]
- Improving Global Outcomes (KDIGO) Transplant Work Group. Special Issue: KDIGO clinical practice guidelines for the care of kidney transplant recipients. Am. J. Transplant. 2009, 9, S1–S155. [Google Scholar]
- Matas, A.J.; Smith, J.M.; Skeans, M.A.; Thompson, B.; Gustafson, S.K.; Stewart, D.E.; Cherikh, W.S.; Wainright, J.L.; Boyle, G.; Snyder, J.J.; Israni, A.K.; Kasiske, B.L. OPTN/SRTR 2013 Annual Data Report: Kidney. Am. J. Transplant. 2013, 15, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Grams, M.E.; Massie, A.B.; Schold, J.D.; Chen, B.P.; Segev, D.L. Trends in the inactive kidney transplant waitlist and implications for candidate survival. Am. J. Transplant. 2013, 13, 1012–1018. [Google Scholar] [CrossRef] [PubMed]
- Miyagawa, S.; Sawa, Y.; Kitagawa-Sakakida, S.; Taketani, S.; Kondoh, H.; Memon, I.A. Tissue cardiomyoplasty using bioengineered contractile cardiomyocyte sheets to repair damaged myocardium: their integration with recipient myocardium. Transplantation 2005, 80, 1586–1595. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, W.; Melnychenko, I.; Wasmeier, G.; Didie, M.; Naito, H.; Nixdorff, U.; Hess, A.; Budinsky, L.; Brune, K.; Michaelis, B.; et al. Engineered heart tissue grafts improve systolic and diastolic function in infarcted rat hearts. Nat. Med. 2006, 12, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.A.; Li, J.; Mathison, M.; Redkar, A.; Cui, J.; Chronos, N.A.F.; Matheny, R.G.; Badylak, S.F. Extracellular matrix scaffold for cardiac repair. Circulation 2005, 97, 452–458. [Google Scholar]
- MacKay, S.M.; Funke, A.J.; Buffington, D.A.; Humes, H.D. Tissue Engineering of a bioartifical renal tubule. ASAIO J. 1998, 44, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Nikolovski, J.; Gulari, E.; Humes, H.D. Design engineering of a bioartifical renal tubule cell therapy device. Cell. Transplant. 1999, 8, 351–364. [Google Scholar] [PubMed]
- Humes, H.D.; MacKay, S.M.; Funke, A.J.; Buffington, D.A. Tissue engineering of a bioartifical renal tubule assist device: In vitro transport and metabolic characteristics. Kidney Int. 1999, 55, 2502–2514. [Google Scholar] [CrossRef] [PubMed]
- Humes, H.D.; Buffington, D.A.; MacKay, S.M.; Funk, A.J.; Weitzel, W.F. Replacement of renal function in uremic animals with a tissue-engineered kidney. Nature Biotech. 1999, 17, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Humes, H.D.; Weitzel, W.F.; Bartlett, R.H.; Swaniker, F.C.; Paganini, E.P.; Luderer, J.R.; and Sobota, J. Initial clinical results of the bioartifical kidney containing human cells in ICU patients with acute renal failure. Kidney Int. 2004, 66, 1578–1588. [Google Scholar] [CrossRef] [PubMed]
- Tumlin, J.; Wali, R.; Williams, W.; Murray, P.; Tolwani, A.J.; Vinnikova, A.K.; Szerlip, H.M.; Ye, J.; Paganini, E.P.; Dworkin, L.; et al. Efficacy and safety of renal tubule cell therapy for acute renal failure. J. Am. Soc. Nephrol. 2008, 19, 1034–1040. [Google Scholar] [CrossRef] [PubMed]
- Buffington, D.A.; Pino, C.J.; Chen, L.; Westover, A.J.; Hageman, G.; Humes, H.D. Bioartifical Renal Epithelial cell system (BRECS): A compact, cryopreservable extracorporeal renal replacement device. Cell Med. 2012, 4, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Oo, Z.Y.; Kandasamy, K.; Tasnim, F.; Zink, D. A novel design of bioartifical kidneys with improved cell performance and haemocompatibility. J. Cell. Mol. Med. 2013, 17, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Jansen, J.; De Napoli, I.E.; Fedecostante, M.; Schophyizen, C.M.S.; Chevtchi, N.V.; Wilmer, M.J.; van Asbeck, A.H.; Croes, H.J.; Pertijs, J.C.; Wetzels, J.F.M.; et al. Human proximal tubule epithelial cells cultured on hollow fibers: Living membranes that actively transport organic cations. Sci. Rep. 2015, 5, 16702. [Google Scholar] [CrossRef] [PubMed]
- Ott, H.C.; Matthiesen, T.S.; Goh, S.; Black, L.D.; Kren, S.M.; Netoff, T.I.; Taylor, D.A. Perfusion-decellularized matrix: using nature’s platform to engineer a bioartifical heart. Nature Med. 2008, 14, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, K.H.; Batchelder, C.A.; Lee, C.I.; Tarantal, A.F. Decellularized rhesus monkey kidney as a three-dimensional scaffold for renal tissue engineering. Tissue Eng. 2010, 16, 2207–2216. [Google Scholar] [CrossRef] [PubMed]
- Ross, E.A.; Williams, M.J.; Hamazaki, T.; Terada, N.; Clapp, W.L.; Adin, C.; Ellison, G.W.; Jorgensen, M.; Batich, C.D. Embryonic stem cells proliferate and differentiate when seeded into kidney scaffolds. J. Am. Nephrol. 2009, 20, 2338–2347. [Google Scholar] [CrossRef] [PubMed]
- Ross, E.A.; Abrahamson, D.R.; St. John, P.L.; Clapp, W.L.; Williams, M.J.; Terada, N.; Hamazaki, T.; Ellison, G.W.; Batich, C.D. Mouse stem cells seeded into decellularized rat kidney scaffolds endothelialize and remodel basement membranes. Organogenesis 2012, 8, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Bonandrini, B.; Figliuzzi, M.; Papadimou, E.; Morigi, M.; Perico, N.; Casiraghi, F.; Sangalli, F.; Conti, S.; Benigni, A.; Remuzzi, A.; Remuzzi, G. Recellularization of well-preserved acellular kidney scaffold using embryonic stem cells. Tissue Eng. 2014, 20, 1486–1498. [Google Scholar] [CrossRef] [PubMed]
- Song, J.J.; Guyette, J.P.; Glipin, S.E.; Gonzalez, G.; Vacanti, J.P.; Ott, H.C. Regeneration and experimental orthotopic transplantation of a bioengineered kidney. Nat Med. 2013, 19, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Liu, S.; Sun, C.; Cheng, G.; Kong, F.; Luan, Y.; Xie, X.; Zhao, S.; Zhang, D.; Wang, J.; et al. The effective bioengineering method of implantation decellularized renal extracellular matrix scaffolds. Oncotarget 2015, 6, 36126–36138. [Google Scholar] [PubMed]
- Yu, Y.L.; Shao, Y.K.; Ding, Y.Q.; Lin, K.Z.; Chen, B.; Zhang, H.Z.; Zhao, L.N.; Wang, Z.B.; Zhang, J.S.; Tang, M.L.; Mei, J. Decellularized kidney scaffold-mediated renal regeneration. Biomaterials 2014, 35, 6822–6828. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Liu, S.; Liu, Y.; Sun, C.; Cheng, G.; Luan, Y.; Li, K.; Wang, G.; Xie, X.; Zhao, S. Porcine kidneys as a source of ECM scaffold for kidney regeneration. Mater. Sci. Eng. 2015, 56, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Vishwakarma, S.K.; Bhavani, P.G.; Bardia, A.; Abkari, A.; Murthy, G.S.N.; Venkateshwarulu, J.; Khan, A.A. Preparation of natural three-dimensional goat kidney scaffold for the development of bioartifical organ. Indian J. Nephrol. 2014, 24, 372–375. [Google Scholar] [PubMed]
- Orlando, G.; Booth, C.; Wang, Z.; Totonelli, G.; Ross, C.L.; Moran, E.; Salvatori, M.; Maghsoudlou, P.; Turmaie, M.; Delario, G.; et al. Discarded human kidneys as a source of ECM scaffold for kidney regeneration technologies. Biomaterials 2013, 34, 5915–5925. [Google Scholar] [CrossRef] [PubMed]
- Orlando, G.; Farney, A.C.; Iskandar, S.S.; Mirmalek-Sani, S.; Sullivan, D.C.; Moran, E.; AbouShwareb, T.; Paolo, D.C.; Wood, K.J.; Stratta, R.J.; et al. Production and implantation of renal extracellular matrix scaffolds from porcine kidneys as a platform for renal bioengineering investigations. Ann. Surg. 2012, 256, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Lefaucheur, C.; Loupy, A.; Hill, G.S.; Andrade, J.; Nochy, D.; Antoine, C.; Gautreau, C.; Charron, D.; Glotz, D.; Suberbielle-Boissel, C. Preexisting donor-specific HLA antibodies predict outcome in kidney transplantation. J. Am. Soc. Nephrol. 2010, 21, 1398–1406. [Google Scholar] [CrossRef] [PubMed]
- Morales, E.E.; Wingert, R.A. Renal stem reprogramming: prospects in regenerative medicine. World J. Stem Cells 2014, 6, 458–466. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, J.D.; Freytes, D.O.; Anandappa, A.; Oliver, J.A.; Vunjak-Novakovic, G. The regulation of growth and metabolism of kidney stem cell with regional specificity using extracellular matrix derived from kidney. Biomaterials 2013, 34, 9830–9841. [Google Scholar] [CrossRef] [PubMed]
- Finesilver, G.; Kahana, M.; Mitrani, E. Kidney-specific microscaffolds and kidney-derived serum-free condition media support in vitro expansion, differentiation, and organization of human embryonic stem cells. Tissue Eng. 2014, 20, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, K.H.; Lee, C.C.; Batchelder, C.A.; Tarantal, A.F. Tissue specificity of decellularized rhesus monkey kidney and lung scaffolds. PLoS ONE 2013, 8, e64134. [Google Scholar] [CrossRef] [PubMed]
- Takasato, M.; Er, P.X.; Chiu, H.S.; Maier, B.; Baillie, G.J.; Ferguson, C.; Parton, R.G.; Wolvetang, E.J.; Roost, M.S.; Chuva de Sousa Lopes, S.M.; Little, M.H. Kidney organoids from human iPS cells contain multiple lineages and model human nephrogenesis. Nat. Letter 2015, 526, 564–568. [Google Scholar] [CrossRef] [PubMed]
- Xinaris, C.; Benedetti, V.; Novelli, R.; Abbate, M.; Rizzo, P.; Conti, S.; Tomasoni, S.; Corna, D.; Pozzobon, M.; Cavallotti, D.; et al. Functional human podocytes generated in organoids from amniotic fluid stem cells. J. Am. Soc. Nephrol. 2015. pii: ASN.2015030316. [Google Scholar] [CrossRef] [PubMed]
- Morizane, R.; Lam, A.Q.; Freedman, B.S.; Kishi, S.; Valerius, M.T.; Bonventure, J.V. Nephron organoids derived from human pluripotent stem cells model kidney development and injury. Nat. Biotech. 2015, 33, 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- Hendry, C.E.; Vanslambrouck, J.M.; Ineson, J.; Suhaimi, N.; Takasato, M.; Rae, F.; Little, M.H. Direction transcriptional reprogramming of adult cells to embryonic nephron progenitors. J. Am. Soc. Nephrol. 2015, 24, 1424–1434. [Google Scholar] [CrossRef] [PubMed]
- Gough, N.R. Bioprinting cartilage scaffolds. Regen. Med. 2014, 7, ec347. [Google Scholar] [CrossRef]
- Pereira, R.F.; Bartolo, P.J. 3D bioprinting of photocrosslinkable hydrogel constructs. J. Appl. Polym. Sci. 2015, 132, 42458. [Google Scholar] [CrossRef]
- Nakamura, M.; Kobayashi, A.; Takagi, F.; Watanabe, A.; Hiruma, Y.; Ohuchi, K.; Iwasaki, Y.; Horie, M.; Morita, I.; Takatani, S. Biocompatible inkjet printing technique for designed seeding of individual living cells. Tissue Eng. 2005, 11, 1658–1666. [Google Scholar] [CrossRef] [PubMed]
- Kolesky, D.B.; Truby, R.L.; Gladman, S.; Busbee, T.A.; Homan, K.A.; Lewis, J.A. 3D bioprinting of vascularized, heterogenous cell-laden tissue constructs. Adv. Mater. 2014, 26, 3124–3130. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Chen, B.; Meng, Q.; Liu, S.; Zheng, X.; Zhang, C.; Wang, H.; Li, H.; Wang, N.; Dai, J. 3D bioprinting of BMSC-laden methacrylamide gelatin scaffolds with CBD-BMP2-collagen microfibers. Biofabrication 2015, 7, 044104. [Google Scholar] [CrossRef] [PubMed]
- Markstedt, K.; Mantas, A.; Tournier, I.; Avila, H.M.; Hagg, D.; Gatenholm, P. 3D bioprinting human chrondrocytes with nanocellulose-alginate bioink for cartilage tissue engineering applications. Biomacromolecules 2015, 16, 1489–1496. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.N.; Lancaster, K.Z.; Zhen, G.; He, J.; Gupta, M.K.; Kong, Y.L.; Engel, E.A.; Krick, K.D.; Ju, A.; Meng, F.; et al. 3D printed anatomical nerve regeneration pathways. Adv. Funct. Mater. 2015, 25, 6205–6217. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McKee, R.A.; Wingert, R.A. Repopulating Decellularized Kidney Scaffolds: An Avenue for Ex Vivo Organ Generation. Materials 2016, 9, 190. https://doi.org/10.3390/ma9030190
McKee RA, Wingert RA. Repopulating Decellularized Kidney Scaffolds: An Avenue for Ex Vivo Organ Generation. Materials. 2016; 9(3):190. https://doi.org/10.3390/ma9030190
Chicago/Turabian StyleMcKee, Robert A., and Rebecca A. Wingert. 2016. "Repopulating Decellularized Kidney Scaffolds: An Avenue for Ex Vivo Organ Generation" Materials 9, no. 3: 190. https://doi.org/10.3390/ma9030190
APA StyleMcKee, R. A., & Wingert, R. A. (2016). Repopulating Decellularized Kidney Scaffolds: An Avenue for Ex Vivo Organ Generation. Materials, 9(3), 190. https://doi.org/10.3390/ma9030190