Proteomic Analysis of Porcine-Derived Collagen Membrane and Matrix



Abstract
1. Introduction
2. Materials and Methods
2.1. Preparing Collagen Membranes for Analysis
2.2. Peptide Separation
2.3. Mass Spectrometry (MS) Analysis
2.4. Data Availability
3. Results
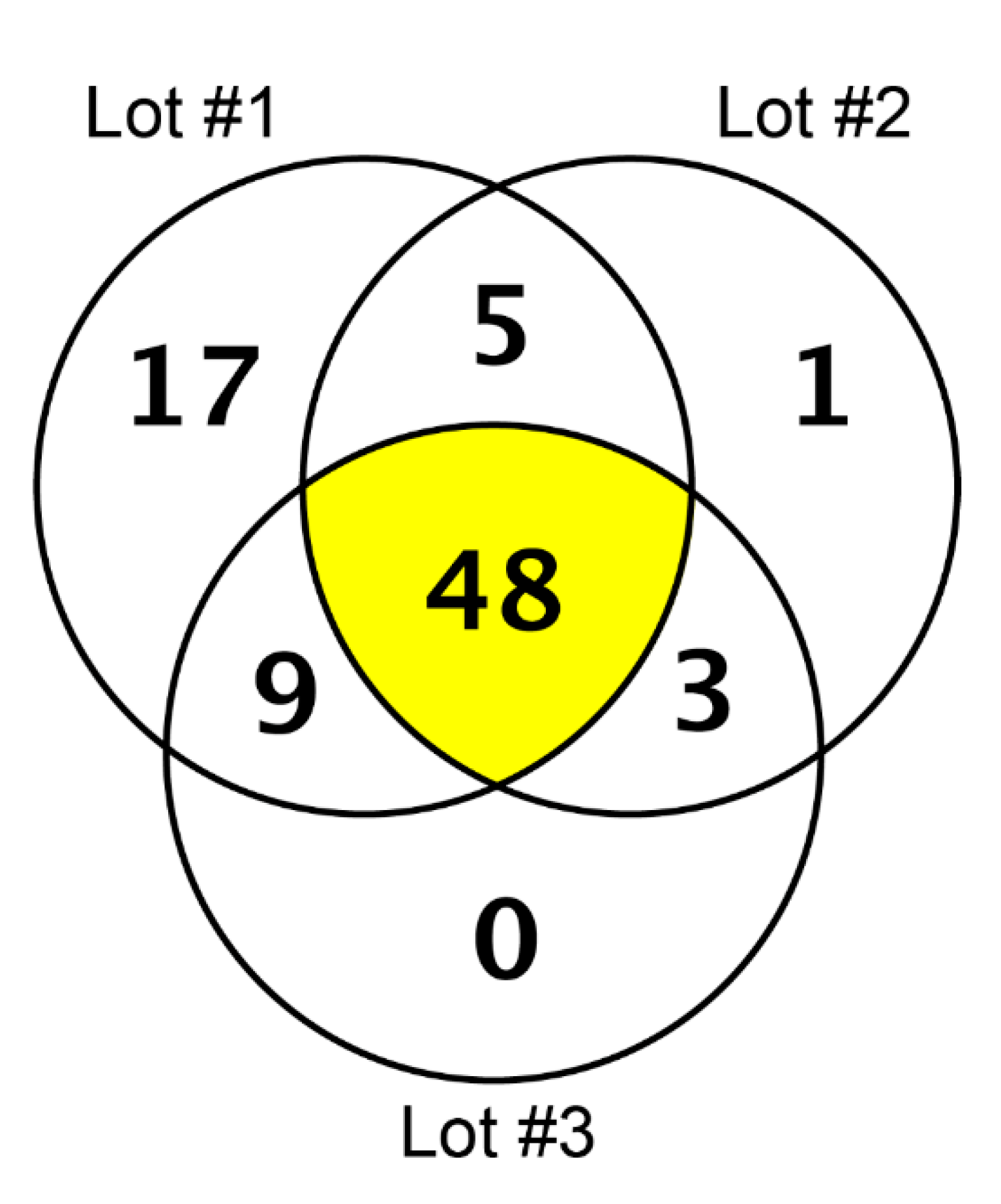
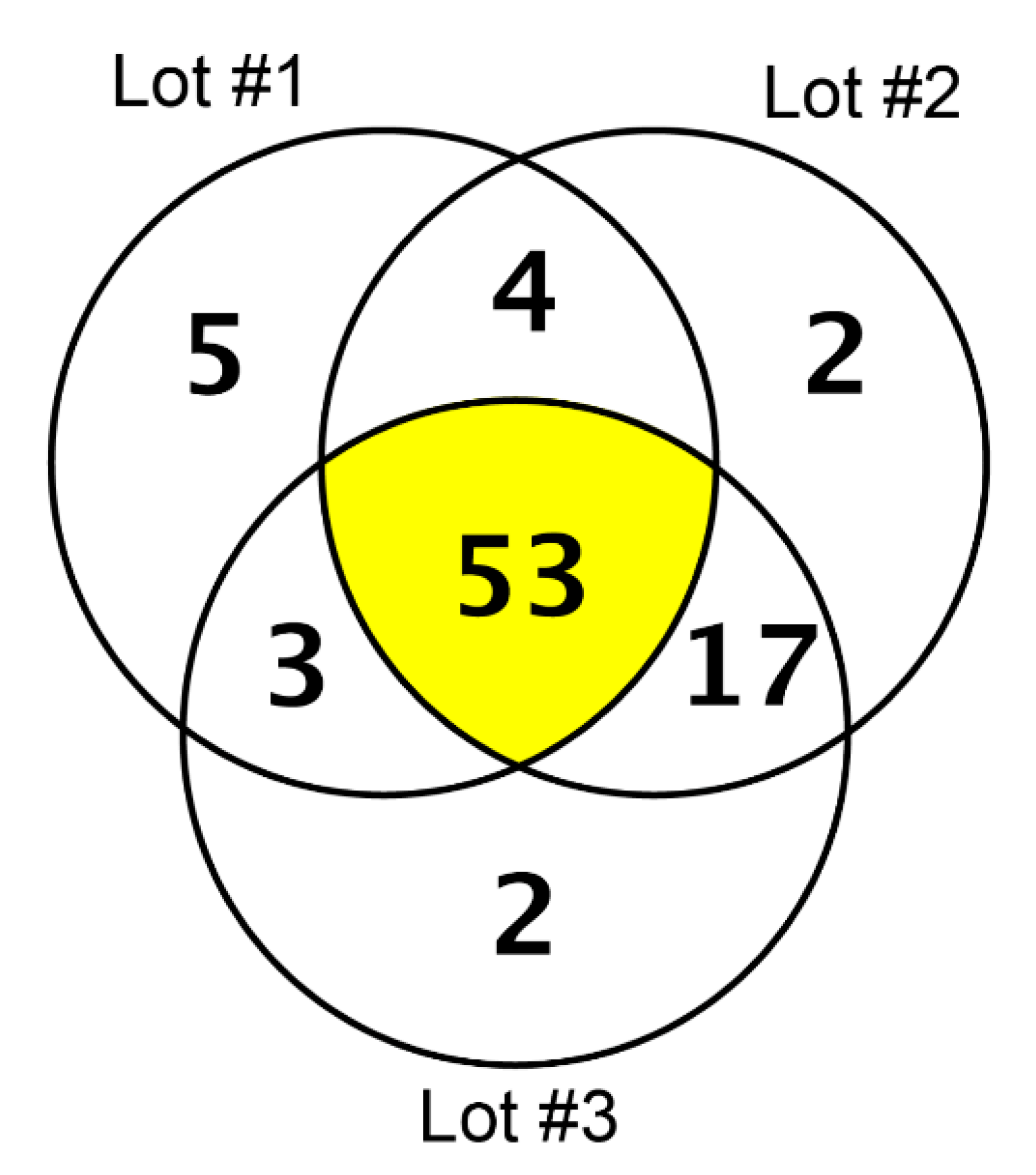
3.1. Proteins Present in Collagen Membrane and Matrix
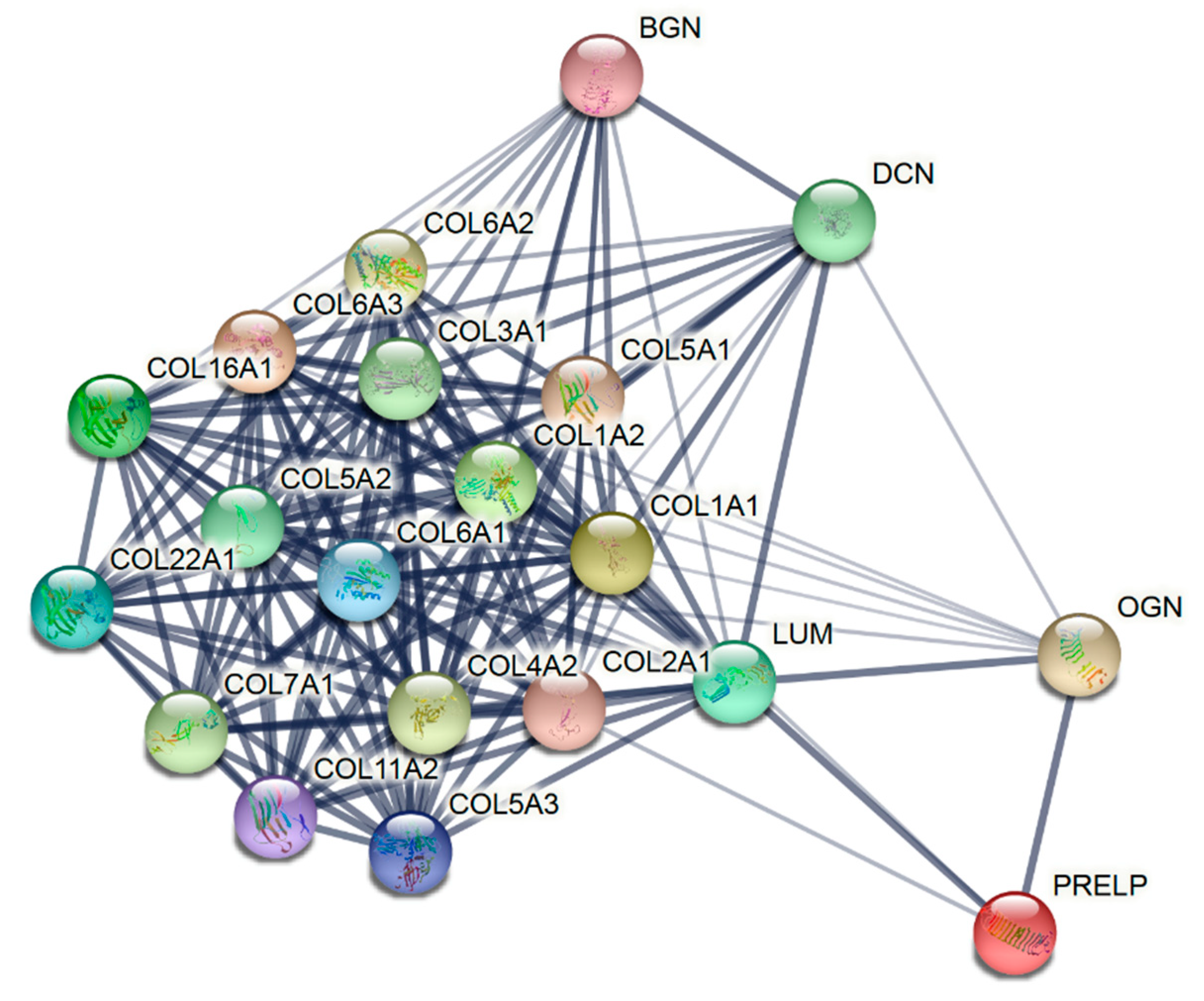
3.2. Proteins Present Exclusively in Collagen Membrane and Matrix
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dahlin, C.; Linde, A.; Gottlow, J.; Nyman, S. Healing of bone defects by guided tissue regeneration. Plast. Reconstr. Surg. 1988, 81, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Schenk, R.K.; Buser, D.; Hardwick, W.R.; Dahlin, C. Healing pattern of bone regeneration in membrane-protected defects: A histologic study in the canine mandible. Int. J. Oral Maxillofac. Implant. 1994, 9, 13–29. [Google Scholar]
- Cucchi, A.; Chierico, A.; Fontana, F.; Mazzocco, F.; Cinquegrana, C.; Belleggia, F.; Rossetti, P.; Soardi, C.M.; Todisco, M.; Luongo, R.; et al. Statements and Recommendations for Guided Bone Regeneration: Consensus Report of the Guided Bone Regeneration Symposium Held in Bologna, October 15 to 16, 2016. Implant Dent. 2019, 28, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Turri, A.; Elgali, I.; Vazirisani, F.; Johansson, A.; Emanuelsson, L.; Dahlin, C.; Thomsen, P.; Omar, O. Guided bone regeneration is promoted by the molecular events in the membrane compartment. Biomaterials 2016, 84, 167–183. [Google Scholar] [CrossRef]
- Omar, O.; Elgali, I.; Dahlin, C.; Thomsen, P. Barrier membranes: More than the barrier effect? J. Clin. Periodontol. 2019, 46 (Suppl. S21), 103–123. [Google Scholar] [CrossRef]
- Elgali, I.; Omar, O.; Dahlin, C.; Thomsen, P. Guided bone regeneration: Materials and biological mechanisms revisited. Eur. J. Oral Sci. 2017, 125, 315–337. [Google Scholar] [CrossRef]
- Taguchi, Y.; Amizuka, N.; Nakadate, M.; Ohnishi, H.; Fujii, N.; Oda, K.; Nomura, S.; Maeda, T. A histological evaluation for guided bone regeneration induced by a collagenous membrane. Biomaterials 2005, 26, 6158–6166. [Google Scholar] [CrossRef]
- You, P.; Liu, Y.; Wang, X.; Li, B.; Wu, W.; Tang, L. Acellular pericardium: A naturally hierarchical, osteoconductive, and osteoinductive biomaterial for guided bone regeneration. J. Biomed. Mater. Res. 2020, 1–14. [Google Scholar] [CrossRef]
- Zhao, S.; Pinholt, E.M.; Madsen, J.E.; Donath, K. Histological evaluation of different biodegradable and non-biodegradable membranes implanted subcutaneously in rats. J. Craniomaxillofac. Surg. 2000, 28, 116–122. [Google Scholar] [CrossRef]
- Ghanaati, S.; Schlee, M.; Webber, M.J.; Willershausen, I.; Barbeck, M.; Balic, E.; Gorlach, C.; Stupp, S.I.; Sader, R.A.; Kirkpatrick, C.J. Evaluation of the tissue reaction to a new bilayered collagen matrix in vivo and its translation to the clinic. Biomed. Mater. 2011, 6, 015010. [Google Scholar] [CrossRef]
- Calciolari, E.; Ravanetti, F.; Strange, A.; Mardas, N.; Bozec, L.; Cacchioli, A.; Kostomitsopoulos, N.; Donos, N. Degradation pattern of a porcine collagen membrane in an in vivo model of guided bone regeneration. J. Periodontal. Res. 2018, 53, 430–439. [Google Scholar] [CrossRef] [PubMed]
- El-Jawhari, J.J.; Moisley, K.; Jones, E.; Giannoudis, P.V. A crosslinked collagen membrane versus a non-crosslinked bilayer collagen membrane for supporting osteogenic functions of human bone marrow-multipotent stromal cells. Eur. Cell. Mater. 2019, 37, 292–309. [Google Scholar] [CrossRef] [PubMed]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell. Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef]
- Witz, C.A.; Montoya-Rodriguez, I.A.; Cho, S.; Centonze, V.E.; Bonewald, L.F.; Schenken, R.S. Composition of the extracellular matrix of the peritoneum. J. Soc. Gynecol. Investig. 2001, 8, 299–304. [Google Scholar] [CrossRef]
- Schlegel, A.K.; Mohler, H.; Busch, F.; Mehl, A. Preclinical and clinical studies of a collagen membrane (Bio-Gide). Biomaterials 1997, 18, 535–538. [Google Scholar] [CrossRef]
- Schmitt, C.M.; Moest, T.; Lutz, R.; Wehrhan, F.; Neukam, F.W.; Schlegel, K.A. Long-term outcomes after vestibuloplasty with a porcine collagen matrix (Mucograft((R))) versus the free gingival graft: A comparative prospective clinical trial. Clin. Oral Implant. Res. 2016, 27, e125–e133. [Google Scholar] [CrossRef]
- Theocharidis, G.; Drymoussi, Z.; Kao, A.P.; Barber, A.H.; Lee, D.A.; Braun, K.M.; Connelly, J.T. Type VI Collagen Regulates Dermal Matrix Assembly and Fibroblast Motility. J. Investig. Dermatol. 2016, 136, 74–83. [Google Scholar] [CrossRef]
- Hill, D.S.; Robinson, N.D.; Caley, M.P.; Chen, M.; O’Toole, E.A.; Armstrong, J.L.; Przyborski, S.; Lovat, P.E. A Novel Fully Humanized 3D Skin Equivalent to Model Early Melanoma Invasion. Mol. Cancer. Ther. 2015, 14, 2665–2673. [Google Scholar] [CrossRef] [PubMed]
- Debeer, S.; Le Luduec, J.B.; Kaiserlian, D.; Laurent, P.; Nicolas, J.F.; Dubois, B.; Kanitakis, J. Comparative histology and immunohistochemistry of porcine versus human skin. Eur. J. Dermatol. 2013, 23, 456–466. [Google Scholar] [CrossRef]
- Agarwal, P.; Zwolanek, D.; Keene, D.R.; Schulz, J.N.; Blumbach, K.; Heinegard, D.; Zaucke, F.; Paulsson, M.; Krieg, T.; Koch, M.; et al. Collagen XII and XIV, new partners of cartilage oligomeric matrix protein in the skin extracellular matrix suprastructure. J. Biol. Chem. 2012, 287, 22549–22559. [Google Scholar] [CrossRef]
- Pang, X.; Dong, N.; Zheng, Z. Small Leucine-Rich Proteoglycans in Skin Wound Healing. Front. Pharmacol. 2019, 10, 1649. [Google Scholar] [CrossRef] [PubMed]
- Matsushima, N.; Ohyanagi, T.; Tanaka, T.; Kretsinger, R.H. Super-motifs and evolution of tandem leucine-rich repeats within the small proteoglycans—Biglycan, decorin, lumican, fibromodulin, PRELP, keratocan, osteoadherin, epiphycan, and osteoglycin. Proteins 2000, 38, 210–225. [Google Scholar] [CrossRef]
- Hildebrand, A.; Romaris, M.; Rasmussen, L.M.; Heinegard, D.; Twardzik, D.R.; Border, W.A.; Ruoslahti, E. Interaction of the small interstitial proteoglycans biglycan, decorin and fibromodulin with transforming growth factor beta. Biochem. J. 1994, 302, 527–534. [Google Scholar] [CrossRef]
- Yung, S.; Thomas, G.J.; Stylianou, E.; Williams, J.D.; Coles, G.A.; Davies, M. Source of peritoneal proteoglycans. Human peritoneal mesothelial cells synthesize and secrete mainly small dermatan sulfate proteoglycans. Am. J. Pathol. 1995, 146, 520–529. [Google Scholar] [PubMed]
- Strauss, F.J.; Stähli, A.; Beer, L.; Mitulović, G.; Gilmozzi, V.; Haspel, N.; Schwab, G.; Gruber, R. Acid bone lysate activates TGFβ signalling in human oral fibroblasts. Sci. Rep. 2018, 8, 16065. [Google Scholar] [CrossRef]
- Toth, G.; Panic-Jankovic, T.; Mitulovic, G. Pillar array columns for peptide separations in nanoscale reversed-phase chromatography. J. Chromatogr. A 2019, 1603, 426–432. [Google Scholar] [CrossRef]
- Panic-Jankovic, T.; Mitulovic, G. Human chorionic gonadotrophin pharmaceutical formulations of urinary origin display high levels of contaminant proteins-A label-free quantitation proteomics study. Electrophoresis 2019, 40, 1622–1629. [Google Scholar] [CrossRef]
- Kotowski, U.; Erović, B.M.; Schnöll, J.; Stanek, V.; Janik, S.; Steurer, M.; Mitulović, G. Quantitative proteome analysis of Merkel cell carcinoma cell lines using SILAC. Clin. Proteomics 2019, 16, 42. [Google Scholar] [CrossRef]
- Bekes, K.; Mitulović, G.; Meißner, N.; Resch, U.; Gruber, R. Saliva proteomic patterns in patients with molar incisor hypomineralization. Sci. Rep. 2020, 10, 7560. [Google Scholar] [CrossRef]
- Othman, Z.; Cillero Pastor, B.; van Rijt, S.; Habibovic, P. Understanding interactions between biomaterials and biological systems using proteomics. Biomaterials 2018, 167, 191–204. [Google Scholar] [CrossRef]
- Abdallah, M.N.; Tran, S.D.; Abughanam, G.; Laurenti, M.; Zuanazzi, D.; Mezour, M.A.; Xiao, Y.; Cerruti, M.; Siqueira, W.L.; Tamimi, F. Biomaterial surface proteomic signature determines interaction with epithelial cells. Acta Biomater. 2017, 54, 150–163. [Google Scholar] [CrossRef] [PubMed]
- Bi, H.; Ye, K.; Jin, S. Proteomic analysis of decellularized pancreatic matrix identifies collagen V as a critical regulator for islet organogenesis from human pluripotent stem cells. Biomaterials 2020, 233, 119673. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, H.; Lu, Q.; Chen, D.; Zhou, M.; Kuang, Y.; Ying, S.; Song, J. Proteomics and N-glycoproteomics analysis of an extracellular matrix-based scaffold-human treated dentin matrix. J. Tissue Eng. Regen. Med. 2019, 13, 1164–1177. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Uygun, B.E.; Geerts, S.; Ozer, S.; Scalf, M.; Gilpin, S.E.; Ott, H.C.; Yarmush, M.L.; Smith, L.M.; Welham, N.V.; et al. Proteomic analysis of naturally-sourced biological scaffolds. Biomaterials 2016, 75, 37–46. [Google Scholar] [CrossRef]
- Al-Maawi, S.; Herrera-Vizcaino, C.; Orlowska, A.; Willershausen, I.; Sader, R.; Miron, R.J.; Choukroun, J.; Ghanaati, S. Biologization of Collagen-Based Biomaterials Using Liquid-Platelet-Rich Fibrin: New Insights into Clinically Applicable Tissue Engineering. Materials 2019, 12, 3993. [Google Scholar] [CrossRef]
- Schöbinger, M.; Klein, O.J.; Mitulović, G. Low-Temperature Mobile Phase for Peptide Trapping at Elevated Separation Temperature Prior to Nano RP-HPLC–MS/MS. Separations 2016, 3, 6. [Google Scholar] [CrossRef]
- Swissprot Sus Scrofa Database. Available online: https://www.uniprot.org/proteomes/UP000008227 (accessed on 13 November 2020).
- ELIXIR Core Data Resources-STRING Homepage. Available online: https://string-db.org (accessed on 13 November 2020).
- Perez-Riverol, Y.; Xu, Q.W.; Wang, R.; Uszkoreit, J.; Griss, J.; Sanchez, A.; Reisinger, F.; Csordas, A.; Ternent, T.; Del-Toro, N.; et al. PRIDE Inspector Toolsuite: Moving Toward a Universal Visualization Tool for Proteomics Data Standard Formats and Quality Assessment of ProteomeXchange Datasets. Mol. Cell Proteomics 2016, 15, 305–317. [Google Scholar] [CrossRef]
- Perez-Riverol, Y. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
- Caballe-Serrano, J.; Sawada, K.; Miron, R.J.; Bosshardt, D.D.; Buser, D.; Gruber, R. Collagen barrier membranes adsorb growth factors liberated from autogenous bone chips. Clin. Oral Implants Res. 2017, 28, 236–241. [Google Scholar] [CrossRef]
- Di Summa, F.; Kargarpour, Z.; Nasirzade, J.; Stahli, A.; Mitulovic, G.; Panic-Jankovic, T.; Koller, V.; Kaltenbach, C.; Muller, H.; Panahipour, L.; et al. TGFbeta activity released from platelet-rich fibrin adsorbs to titanium surface and collagen membranes. Sci. Rep. 2020, 10, 10203. [Google Scholar] [CrossRef]
- Geistlich, P.; Eckmayer, Z.; Boyne, P. Resorbable collagen membrane for use in guided tissue regeneration. U.S. Patent 5,837,278, 17 November 1998. [Google Scholar]
- Crapo, P.M.; Gilbert, T.W.; Badylak, S.F. An overview of tissue and whole organ decellularization processes. Biomaterials 2011, 32, 3233–3243. [Google Scholar] [CrossRef] [PubMed]
- Tazelaar, H.D.; Byrne, G.W.; McGregor, C.G. Comparison of Gal and non-Gal-mediated cardiac xenograft rejection. Transplantation 2011, 91, 968–975. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.C.; Kuo, Y.J.; Sun, F.W.; Chen, C.H.; Chiang, C.J.; Weng, P.W.; Tsuang, Y.H.; Huang, Y.Y. Optimized decellularization protocol including alpha-Gal epitope reduction for fabrication of an acellular porcine annulus fibrosus scaffold. Cell Tissue Bank 2017, 18, 383–396. [Google Scholar] [CrossRef]
- Allan, B.; Ruan, R.; Landao-Bassonga, E.; Gillman, N.; Wang, T.; Gao, J.; Ruan, Y.; Xu, Y.; Lee, C.; Goonewardene, M.; et al. Collagen Membrane for Guided Bone Regeneration in Dental and Orthopedic Applications. Tissue Eng. Part. A 2020. submitted for publication. [Google Scholar] [CrossRef]
- Griffiths, L.G.; Choe, L.H.; Reardon, K.F.; Dow, S.W.; Christopher Orton, E. Immunoproteomic identification of bovine pericardium xenoantigens. Biomaterials 2008, 29, 3514–3520. [Google Scholar] [CrossRef]
- Burlak, C.; Wang, Z.Y.; Chihara, R.K.; Lutz, A.J.; Wang, Y.; Estrada, J.L.; Tector, A.J. Identification of human preformed antibody targets in GTKO pigs. Xenotransplantation 2012, 19, 92–101. [Google Scholar] [CrossRef]
- Cescon, M.; Gattazzo, F.; Chen, P.; Bonaldo, P. Collagen VI at a glance. J. Cell. Sci. 2015, 128, 3525–3531. [Google Scholar] [CrossRef] [PubMed]
- Kuchler, U.; Rybaczek, T.; Dobask, T.; Heimel, P.; Tangl, S.; Klehm, J.; Menzel, M.; Gruber, R. Bone-conditioned medium modulates the osteoconductive properties of collagen membranes in a rat calvaria defect model. Clin. Oral Implants Res. 2018, 29, 381–388. [Google Scholar] [CrossRef]
- Strauss, F.J.; Kuchler, U.; Kobatake, R.; Heimel, P.; Tangl, S.; Gruber, R. Acid bone lysates reduce bone regeneration in rat calvaria defects. J. Biomed. Mater. Res. 2020, 1–7. [Google Scholar] [CrossRef]
- Wiesmann, H.P.; Meyer, U.; Plate, U.; Hohling, H.J. Aspects of collagen mineralization in hard tissue formation. Int. Rev. Cytol. 2005, 242, 121–156. [Google Scholar]
- Ricard-Blum, S.; Salza, R. Matricryptins and matrikines: Biologically active fragments of the extracellular matrix. Exp. Dermatol. 2014, 23, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Holstein, J.H.; Karabin-Kehl, B.; Scheuer, C.; Garcia, P.; Histing, T.; Meier, C.; Benninger, E.; Menger, M.D.; Pohlemann, T. Endostatin inhibits Callus remodeling during fracture healing in mice. J. Orthop. Res. 2013, 31, 1579–1584. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Matsumoto, E.; Higashimaki, Y.; Katagiri, T.; Sugimoto, T.; Seino, S.; Kaji, H. Role of osteoglycin in the linkage between muscle and bone. J. Biol. Chem. 2012, 287, 11616–11628. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.H.; Park, H.J.; Jeong, J.H.; Kim, Y.J.; Cha, D.W.; Kwon, D.K.; Lee, S.H.; Cho, J.Y. The role of asporin in mineralization of human dental pulp stem cells. J. Cell. Physiol. 2011, 226, 1676–1682. [Google Scholar] [CrossRef] [PubMed]
- Haruyama, N.; Sreenath, T.L.; Suzuki, S.; Yao, X.; Wang, Z.; Wang, Y.; Honeycutt, C.; Iozzo, R.V.; Young, M.F.; Kulkarni, A.B. Genetic evidence for key roles of decorin and biglycan in dentin mineralization. Matrix Biol. 2009, 28, 129–136. [Google Scholar] [CrossRef]
- Parisuthiman, D.; Mochida, Y.; Duarte, W.R.; Yamauchi, M. Biglycan modulates osteoblast differentiation and matrix mineralization. J. Bone Miner. Res. 2005, 20, 1878–1886. [Google Scholar] [CrossRef]
- Spinell, T.; Saliter, J.; Hackl, B.; Unger, K.; Hickel, R.; Folwaczny, M. In-vitro cytocompatibility and growth factor content of GBR/GTR membranes. Dent. Mater. 2019, 35, 963–969. [Google Scholar] [CrossRef]
- Panahipour, L.; Kargarpour, Z.; Luza, B.; Lee, J.S.; Gruber, R. TGF-beta Activity Related to the Use of Collagen Membranes: In Vitro Bioassays. Int. J. Mol. Sci. 2020, 21, 6636. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene ID | Protein Name | Gene Symbol |
|---|---|---|
| Q6QAQ1 | Actin, cytoplasmic 1 | ACTB |
| F1SJB5 | Annexin | ANXA1 |
| A0A287A531 | Annexin | ANXA5 |
| F1S2B6 | Biglycan | BGN |
| F1RYI8 | Collagen alpha-1(III) chain precursor | COL3A1 |
| F1S021 | Collagen alpha-1(V) chain precursor | COL5A1 |
| I3L781 | Collagen alpha-2(I) chain precursor | COL1A2 |
| A0A287BF88 | Collagen alpha-2(V) chain precursor | COL5A2 |
| F1S3G7 | Collagen alpha-3(V) chain precursor | COL5A3 |
| A0A287A1S6 | Collagen type I alpha 1 chain | COL1A1 |
| A0A286ZWS8 | Collagen type II alpha 1 chain | COL2A1 |
| F1RLL9 | Collagen type IV alpha 2 chain | COL4A2 |
| A0A287B5M9 | Collagen type VI alpha 1 chain | COL6A1 |
| I3LQ84 | Collagen type VI alpha 2 chain | COL6A2 |
| I3LUR7 | Collagen type VI alpha 3 chain | COL6A3 |
| F1SKM1 | Collagen type VII alpha 1 chain | COL7A1 |
| K7GME7 | Collagen type XI alpha 2 chain | COL11A2 |
| A0A286ZHY0 | Collagen type XVI alpha 1 chain | COL16A1 |
| F1RSI7 | Collagen type XXII alpha 1 chain | COL22A1 |
| Q9XSD9 | Decorin | DCN |
| I3LC73 | Fatty acid synthase | FASN |
| F1RII7 | Hemoglobin subunit beta | HBB |
| A0A287B959 | Histone H2A | HIST1H2AC |
| F2Z584 | Histone H2B | HIST1H2BD |
| P62802 | Histone H4 | no symbol for pig |
| A0A286ZZ03 | Ig-like and FN type III dom.-cont. protein 1 | IGFN1 |
| F1SQ09 | Lumican precursor | LUM |
| A0A287AB52 | Microfibril associated protein 4 | MFAP4 |
| A0A0H5ANC0 | Mimecan precursor | OGN |
| F1SSA6 | Myosin motor domain-containing protein | MYH10 |
| F1S6B4 | Prol. arg. rich end leuc. rich repeat protein | PRELP |
| I3LSK9 | Retrotransposon Gag like 9 | RTL9 |
| A0A287BAY9 | Serum albumin | ALB |
| A0A287B5W2 | Trypsinogen precursor | LOC100302368 |
| Q2XVP4 | Tubulin alpha-1B chain | TUBA1B |
| Q767L7 | Tubulin beta chain | TUBB |
| P02543 | Vimentin | VIM |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-S.; Mitulović, G.; Panahipour, L.; Gruber, R. Proteomic Analysis of Porcine-Derived Collagen Membrane and Matrix. Materials 2020, 13, 5187. https://doi.org/10.3390/ma13225187
Lee J-S, Mitulović G, Panahipour L, Gruber R. Proteomic Analysis of Porcine-Derived Collagen Membrane and Matrix. Materials. 2020; 13(22):5187. https://doi.org/10.3390/ma13225187
Chicago/Turabian StyleLee, Jung-Seok, Goran Mitulović, Layla Panahipour, and Reinhard Gruber. 2020. "Proteomic Analysis of Porcine-Derived Collagen Membrane and Matrix" Materials 13, no. 22: 5187. https://doi.org/10.3390/ma13225187
APA StyleLee, J.-S., Mitulović, G., Panahipour, L., & Gruber, R. (2020). Proteomic Analysis of Porcine-Derived Collagen Membrane and Matrix. Materials, 13(22), 5187. https://doi.org/10.3390/ma13225187