1. Introduction
The wear and corrosion of metal implant materials are the main risk factors for aseptic loosening and implant failure in orthopedic surgery. Although metal surfaces are protected by a passive oxygen layer, electrochemical reactions are still present at the implant site [
1]. Different components can cause galvanic effects resulting in oxidation and reduction reactions on the metal surfaces. As a consequence, a continuous exchange of electrons as well as ions between the metal and surrounding liquid is maintained [
2]. Head-neck taper corrosion is the most prominent example of this in orthopedic surgery, especially for large-diameter metal-on-metal bearings [
3], but also for metal-polyethylene bearings [
4]. Another type of corrosion is the intracellular reduction of wear particles in cells, especially in macrophages. When metal particles are taken up via active mechanisms or internalization, the degradation of these nanostructures is promoted by the lysosomal environment [
5]. Due to the acidic pH in the cellular components, the release of metal ions in different oxidation states is fostered [
6]. Besides the intracellular accumulation of these toxic substances, metal ions are released into the extracellular environment where other cell types can be directly influenced. However, osteolysis is not directly induced by the release of ions but rather the reaction of ions with biomolecules, which leads to higher bioactivity and destruction [
7]. A third mechanism of corrosion has been described as direct inflammatory cell-induced corrosion, which occurs on retrieved metal implants. Here, the direct cell attack and concomitant formation of a ruffled cell membrane have been assumed as a trigger for corrosive processes [
8].
Apart from titanium-aluminum-vanadium, cobalt-chromium-molybdenum (CoCr28Mo6) is the most prominent alloy in orthopedic surgery. While cobalt alloys are characterized by their high strength, fracture toughness, and excellent biocompatibility, the degradation of the surface due to corrosion, or a combination of corrosion and wear, can lead to harmful local and systemic side-effects in the human body [
2,
6,
9]. Cytotoxic effects of Co, Cr, Ti, and nickel (Ni), which present themselves mainly as apoptosis, necrosis, and inhibitory effects on the DNA repair mechanism, are caused by means of chromosomal damage and oxidative stress [
2,
10].
Despite these similar outcomes, different metal ions vary with regard to local and systemic effects. Cr ions are able to react with phosphate compounds to form stable chromium phosphates (CrPo
4) which then accumulate in the peri-implant tissue [
2,
6]. In contrast, Co ions can circulate within the body, affecting many organs and causing, e.g., neurological, cardiological, or endocrine symptoms [
2]. The varying behavior is also reflected in the different concentrations found locally as well as in the blood of patients suffering from aseptic loosening and metallosis. As blood is easily accessible, there is a sound base of data. Serum cobalt concentrations of 2–7 µg/L (0.002–0.007 ppm) or 10 µg/L (0.010 ppm) were stated respectively in a European multidisciplinary consensus statement [
11] or by the Mayo Clinic [
12] as indicative of metallic wear and implant loosening. The systemic concentrations reported in patients with acute cobalt poisoning after excessive wear particle release with fatal or near fatal outcomes were approximately a hundred times higher (serum levels of around 0.40–0.64 ppm cobalt and 0.05–0.08 ppm chromium) than the suggested threshold, with even higher values excreted in urine [
13,
14,
15,
16,
17]. While cobalt concentrations are higher than chromium concentrations in serum this is reversed in the periprosthetic tissue, partially due to the above mentioned reasons. Scharf et al. [
6] measured an average of 0.17 ppm for cobalt and 1.60 ppm for chromium in tissue surrounding of total hip implants with metal on metal (MoM) bearings in nine patients with revisions due to adverse local tissue reactions [
6]. In a more recent publication by Kuba et al. (2019) revision patients with long-term surviving implants displayed mean values of 6.52 ± 16.38 ppm for cobalt and 8.88 ± 26.88 ppm for chromium in periprosthetic tissue—concentrations in joint fluid were around 0.048 ppm for Co and 0.167 ppm for Cr [
18]. However, in the rare cases, where wear and loosening led to acute poisoning, local concentrations of 41 ppm Co [
16] up to 397.8 ppm Co and 236.0 ppm Cr [
19] in periprosthetic tissue, or 76 ppm Co and 126.5 ppm Cr in locally aspirated liquid [
14], were reported. These high local concentrations induce detrimental effects and an inflammatory response to metal ion intoxication can be locally found in the periprosthetic tissue. Tissue necrosis is associated with macrophage and lymphocyte infiltration [
6] and the secretion of a variety of pro-inflammatory mediators (e.g., interleukins (IL), tumor necrosis factor (TNF)), especially from macrophages, is induced [
7]. However, various studies have revealed that bone forming osteoblasts are also affected by metal ions, resulting in reduced proliferation and differentiation capacity as well as inducing pro-inflammatory cytokine release [
2,
20,
21]. In this context, it is interesting that mineralized bone tissue seems to be prone to accumulate metal debris as the concentrations of 38–413 ppm Co here were considerably higher than those reported above for soft tissue [
22].
In a previous in vitro study, we exposed human osteoblasts and macrophages to a mixture of Co and Cr ions (200 µg/L or 0.2 ppm) and analyzed their pro-osteolytic capacity in the cell cultures [
21]. Although we observed marginal detrimental effects in single cell cultures, both a clear impact on osteoblastic function and the initiation of pro-osteolytic events were detectable under co-culture conditions of human osteoblasts and macrophages. However, a limitation of this study was the missing knowledge of the proportion of either Co or Cr ions. As a result, proven effects could not be attributed to one metal ion species or another. Thus, for further understanding of differential effects of Co and Cr ions, we now exposed human osteoblasts to different concentrations of Co and Cr metal salts. Based on previous studies by Scharf et al. (2014) and Drynda et al. (2018) we used ion concentrations between 10 µM and 500 µM [
6,
23]. These concentrations correspond to 0.59–29.47 ppm Co and 0.52–26.00 ppm Cr and are similar to those reported in periprosthetic tissue by Kuba et al. (2019) [
18], but lower than those in bone tissue [
22]. However, due to the analytical methodology it is difficult to specify whether the determined metal concentration originated from metal particles or metal ions.
Additionally to Co and Cr, nickel ions—which are known to have a higher toxicity on osteoblastic viability and proliferation than Co and Cr [
2,
24]—were investigated since forced corrosion of CoCr28Mo6 particles resulted in considerable levels of Ni ions in the solution that exceeded the amount of molybdenum [
21]. Simultaneously, human osteoblasts were also treated with CoCr28Mo6 particles in order to clarify whether corrosion products exhibit similar or differing effects to abrasion particles in cells. Hence, the objective of this study was to assess the effects of relevant corrosion products on cell survival, bone remodeling, and the release of pro-osteolytic mediators in mature human osteoblasts. Therefore, short-term studies with different metal ion concentrations were carried out to determine the threshold at which metal salts are harmful for cell survival.
2. Materials and Methods
2.1. Preparation of Metal Salt Solutions
The following metal salts were purchased from Sigma-Aldrich (Sigma-Aldrich Chemie GmbH, Munich, Germany): Cobalt(II) chloride (purum p.a., anhydrous, purity ≥ 98.0% (KT)), Nickel(II) chloride (anhydrous, powder, purity 99.99% trace metals basis) and Chromium(III) chloride hexahydrate (purum p.a., purity ≥ 98.0% (RT)). Stock solutions of a concentration of 100 mM were produced by dissolving the appropriate amount of salt in Aqua ad iniectabilia (B. Braun Melsungen AG, Melsungen, Germany). Stock solutions were stored in Schott Duran
® laboratory glass bottles (Schott AG, Mainz, Germany) at 4 °C in the dark. Before the use in the experiments the chromium salt stock solution was equilibrated according to Drynda et al. (2018) for at least two weeks until the color of the solution changed completely from emerald green to purple, indicating the presence of the stable species of [Cr
3 + (H
2O)
6]Cl
3 [
23]. For cell experiments, the stock solutions were further diluted with cell culture media to expose cells to concentrations of 10 µM, 50 µM, 100 µM and 500 µM, respectively.
2.2. Isolation and Cultivation of Human Primary Osteoblasts
Isolation of human primary osteoblasts (n = 11; eight female donors: mean age 70 years ± 18 years; three male donors: mean age 70 years ± 13 years) was performed according to the protocol of Lochner et al. (2011) [
25]. The bone marrow was extracted under sterile conditions from femoral heads of patients undergoing primary hip replacements who had given written consent (Local Ethical Committee AZ: 2010-10). Extracted spongiosa was washed three times in phosphate-buffered saline (PBS, Biochrom AG, Berlin, Germany) followed by an enzymatic digestion with collagenase A and dispase II (both from Roche, Penzberg, Germany) at 37 °C. The material was filtered through a cell strainer (70 µm pores, BD Biosciences, Bedford, UK) and the isolated cell suspension was centrifuged at 118× g for 10 min. The cell sediment was re-suspended in cell culture medium. Human primary osteoblasts were cultivated in a special formulation of calcium-depleted Dulbecco’s modified Eagle medium (DMEM, Pan Biotech GmbH, Aidenbach, Germany) containing 10% fetal calf serum (Pan Biotech GmbH), 1% penicillin/streptomycin, 1% amphotericin b, 1% HEPES buffer, and the osteogenic additives L-ascorbate-2-phosphate (50 µg/mL), β-glycerophosphate (10 mM), as well as dexamethasone (100 nM) (all: Sigma-Aldrich, Munich, Germany) at 37 °C and 5% CO
2. These specific conditions, in particular the addition of osteogenic factors dexamethasone, β-glycerophosphate and L-ascorbate-2-phosphate, led to the differentiation of the isolated pre-osteoblasts into mature osteoblasts in standard cell culture flasks (polystyrene) [
26,
27]. The osteoblastic phenotype was analyzed by alkaline phosphatase staining (with fuchsin+substrate-chromogen; DAKO, Hamburg, Germany). Cells from passage 3 were harvested for subsequent cell culture experiments as follows. Cells were washed with PBS, trypsinized, and centrifuged at 118×
g. If not otherwise stated, 30,000 cells (in duplicate) were transferred into a well of a 24-well cell culture plate allowing cell adherence over 24 h at 37 °C and 5% CO
2. Afterwards, cells were exposed to different concentrations of metal salts. Untreated cells served as negative controls whereas osteoblasts treated with CoCr28Mo6 particles (particle concentration: 0.01 mg/mL) [
28,
29] were used as positive control. For the comparison of the influence of different culture media, matured cells from passage 3 were not only seeded in the above described Ca-depleted osteogenic medium but also in standard Dulbecco’s modified Eagle medium (Gibco™ DMEM Glutamax, ThermoFisher Scientific, Waltham, MA, USA) with osteogenic additives L-ascorbate-2-phosphate (50 µg/mL), β-glycerophosphate (10 mM) and dexamethasone (100 nM) (all: Sigma-Aldrich) before treatment.
2.3. Cellular Activity
The cell activity of ion-exposed human osteoblasts was determined after 48 h of exposure. Osteoblasts (10,000 cells per well, in duplicates) were seeded in black 96-well cell culture plates (Thermo Fisher Scientific Inc., Waltham, MA, USA). After 24 h of adherence under standard cell culture conditions, cells were treated with metal salts or particles. Supernatants were removed after 48 h and the water soluble tetrazolium salt (WST-1) assay (Roche, Penzberg, Germany) was performed according to the manufacturer’s recommendations. Cells were incubated with a defined volume of WST-1/medium (1:10 ratio) reagent for 30 min. Subsequently, supernatants were transferred in a 96-well cell culture plate and absorbance at 450 nm (reference wave length: 630 nm) was determined in a Tecan Infinite
® 200 Pro microplate reader (Tecan Group AG, Maennedorf, Switzerland). Afterwards, the same cells were used to quantify cell numbers using the CyQUANT
® NF Cell Proliferation Assay Kit (Invitrogen, Thermo Fisher Scientific Inc., Waltham, MA, USA) according to the recommendations of the manufacturer. This combined approach was possible as the CyQUANT
® kit measures the emission of fluorescence at 520 nm after the lysis of the cells and the subsequent binding of a proprietary green fluorescent dye, CyQUANT
® GR dye, to cellular nucleic acids. The fluorescence signal depends solely on the amount of DNA present in the sample. In order to relate the fluorescence signal to an actual cell number, a cell number calibration curve was prepared with defined cell numbers between 0 and 20,000 cells in duplicate prior to each experiment. The calibration curve was then used to determine the cell number of treated cells per well. Fluorescence intensity was measured with the Tecan Infinite
® 200 Pro (Tecan Group AG, Maennedorf, Switzerland) microplate reader (excitation: 485 nm, emission: 535 nm). Cellular activity was calculated by dividing WST-1 results by the respective cell number as measured by CyQuant (see
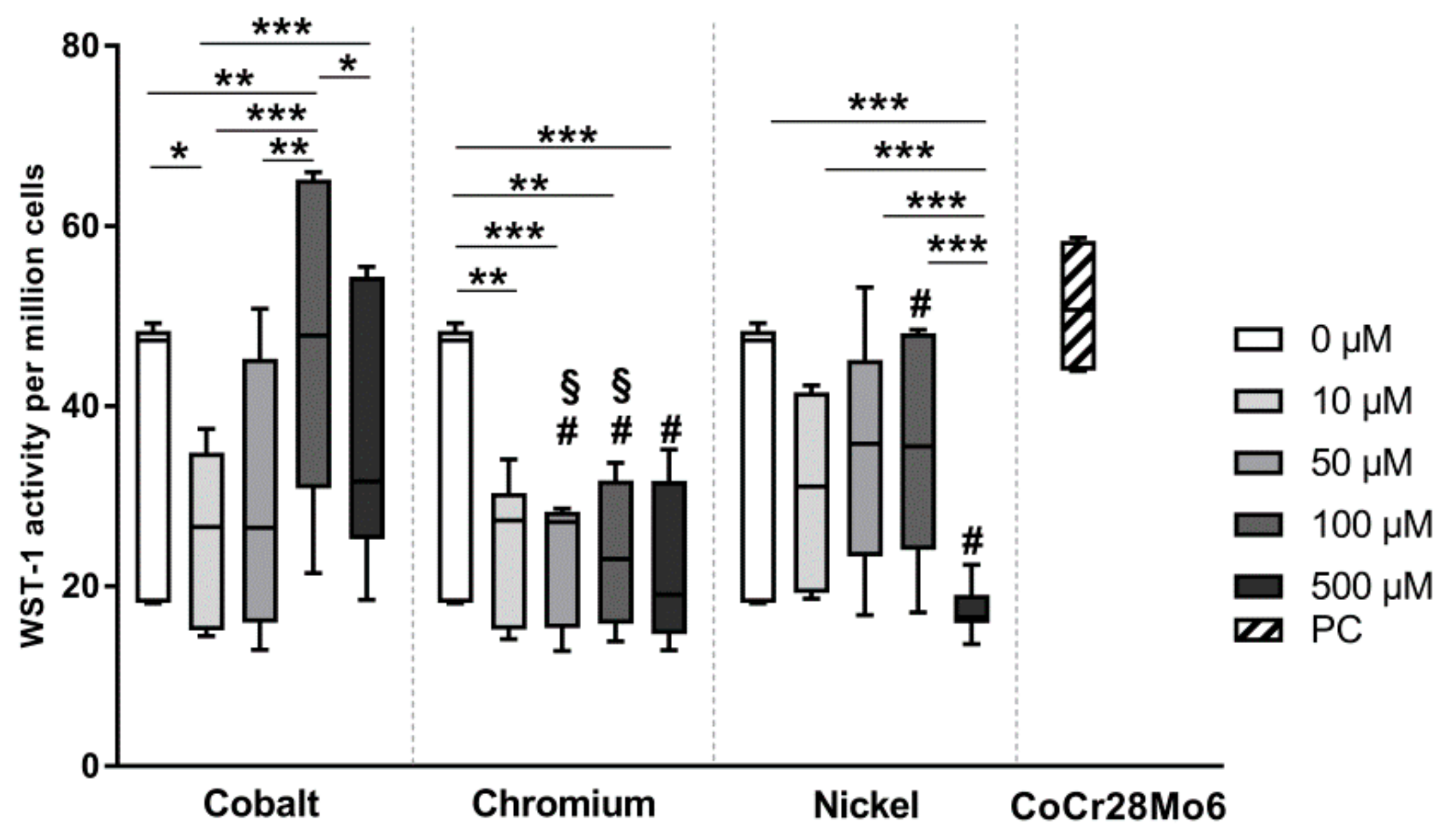
Supplementary Materials Figures S1 and S2). Since the activity values per cell were very small, results are presented as activity per one million cells for easier illustration of data.
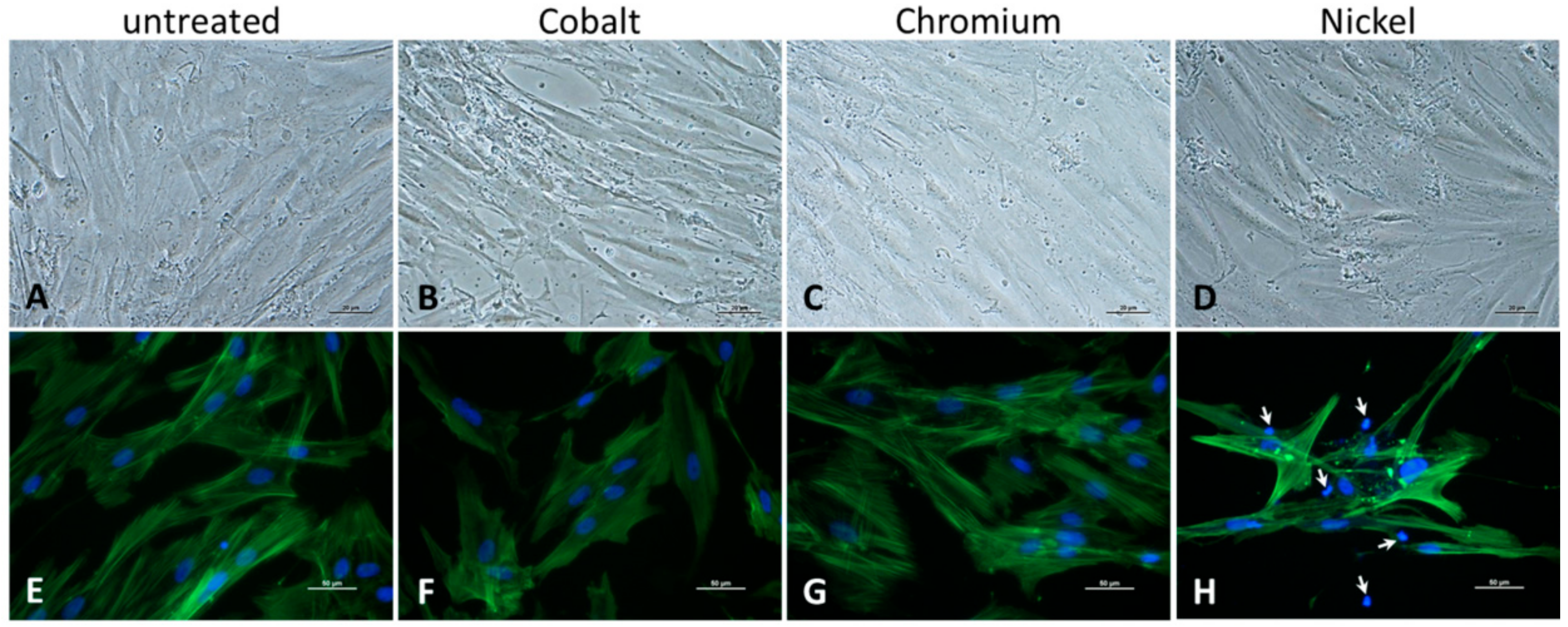
Additionally, microscopic examinations of cell cultures were carried out after 48 h of incubation. Morphology of human osteoblasts was documented via light microscopy using a 200× magnification (Nikon ECLIPSE TS100, Nikon GmbH, Duesseldorf, Germany). The formation of actin filaments within cell structures was visualized by actin staining and DAPI counterstain following the procedure described by Klinder et al. (2018) [
28]. Pictures for actin staining were taken with 400× magnification at 500 nm where actin stain fluoresced green. Cell nuclei were visualized with DAPI stain at 400 nm and showed a blue fluorescence. The respective pictures were taken from exactly the same spot and superimposed upon each other with the help of Adobe Photoshop CS6 image processing software Version 13.0.1 (Adobe Systems Software Ireland Ltd., Dublin, Ireland).
2.4. Gene Expression Analysis
RNA isolation was carried out using the peqGOLD Total RNA Kit (VWR International GmbH, Hanover, Germany) following the manufacturer’s protocol. RNA was eluted into a fresh sterile tube using RNase free water and RNA concentration was measured using the Tecan Infinite® 200 (Tecan Group AG, Maennedorf, Switzerland) microplate reader and NanoQuant Plate™ with RNase free water as blank. The purity of the isolated RNA was assessed and median ratios at 260/280 nm of 2.10 (2.10–2.11), 2.10 (1.78–2.14), 2.13 (2.04–2.19), 2.11(1.90–2.13) and 2.09 (2.08–2.11) were recorded for untreated, Co-treated, Cr-treated, Ni-treated and particle-treated samples, respectively. After RNA isolation, a reverse transcriptase polymerase chain reaction (RT-PCR) was used to transcribe the RNA into cDNA. Here, the High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA) was used. A master mix was prepared as described in the manufacturer’s protocol. The specific amount of each RNA sample containing 200 ng RNA was calculated using the results from concentration measurement and added up to 10 μL with RNase free water in PCR tubes. Subsequently, 10 μL of master mix was added and mixed well. The samples were placed in a thermocycler (Analytik Jena, Jena, Germany) and the following RT-PCR protocol was used: 10 min at 25 °C, 120 min at 37 °C, 15 s at 85 °C. Afterwards, samples were diluted in additional 20 μL RNase free water and stored at −20 °C.
To determine the expression level of differentiation- and inflammation-associated genes, the cDNA of treated and untreated cells was used to perform a semiquantitative real-time (q-PCR) with SybrGreen. For the PCR reaction, the 2× innuMIX qPCR MasterMix SyGreen (Analytik Jena, Jena, Germany) was used following the manufacturer’s protocol. To that master mix, 0.5 μL of forward and reverse primer (12 μM), respectively, as well as 3 μL of Aqua dest. were added. For each sample, 1 μL template cDNA (in duplicates) was pipetted onto the bottom of a 96-well PCR plate and filled up with 9 µL of the mentioned master mix. The used primer sequences of osteogenic (Col1A1, ALP) and pro-osteolytic mediators (IL-6, IL-8) are listed in
Table 1. Distilled water, instead of cDNA, served as negative control. The plate was sealed with adhesive foil and placed in the qTower 2.0 (Analytik Jena, Jena, Germany). Gene expression analysis was done under the following conditions: initial activation time of 2 min at 95 °C, 40 times of rotation of denaturation for 5 s at 95 °C and annealing/elongation for 25 s at 60–65 °C. A cycle of threshold (Ct) of 28 was set as limit. The relative expression of each gene compared to the housekeeping gene hypoxanthine guanine phosphoribosyl transferase (HPRT) was calculated using the equation: ΔCt = Ct
target − Ct
HPRT. The relative amount of target mRNA of cells treated with metal salts and controls was calculated using 2
(−ΔΔCt) with ΔΔCt
treatment = ΔCt
treated − ΔCt
control.
2.5. Protein Analysis
The protein contents of bone remodeling markers (pro-collagen type 1 (C1CP), osteoprotegerin (OPG), receptor activator of nuclear factor κb ligand (RANKL)) and pro-inflammatory mediators (interleukin (IL) 6 and 8) were determined in the supernatant of control and ion-exposed osteoblasts. For this purpose, the supernatants were collected and stored at −20 °C prior to quantification. C1CP was determined using the C1CP ELISA (Quidel, Marburg, Germany) according to the manufacturer’s recommendations. Absorbance was measured at 405 nm (reference wave length: 630 nm) using the Tecan Infinite® 200 Pro (Tecan Group AG, Maennedorf, Switzerland) microplate reader. A standard curve was prepared to calculate protein concentration in samples. OPG and RANKL were determined via LEGENDplex™ (BioLegend, San Diego, CA, USA) using fluorescence-labeled beads. These beads are conjugated with the specific antibody on its surface. The samples were incubated with the antibody-conjugated beads, thus forming capture bead-analyte-detection antibody sandwiches. Afterwards, streptavidin-phycoerythrin was added, which bound to the biotinylated detection antibodies, providing fluorescent signal intensities in proportion to the amount of bound analytes. Fluorescence intensities (excitation: 575 nm, emission: 660 nm) in samples were analyzed on a flow cytometer (FACSAria™ IIIu, BD Biosciences). The concentrations of OPG and RANKL were quantified using a standard curve generated in the same assay as well as LEGENDplexTM Data Analysis Software v8 (BioLegend, San Diego, CA, USA).
Soluble proteins of IL6 and IL8 were quantified via eBioscience™ Human IL-6/IL-8 ELISA Ready-SET-Go!™ Kits (both: ThermoFisher Scientific, Waltham, MA, USA) according to the instructions of the manufacturer. Absorbance was measured at 405 nm (reference wave length: 630 nm) using the Tecan Infinite® 200 Pro (Tecan Group AG, Maennedorf, Switzerland) microplate reader. Sample concentrations were calculated using a standard curve, respectively.
Finally, all protein contents within the samples were normalized to the overall protein content which was quantified by the Qubit Protein Assay Kit and Qubit 1.0 (both: Invitrogen) according to the manufacturer’s instructions.
2.6. Data Analysis and Illustration
Data analysis and illustration were performed by GraphPadPRISM v.7.02 (GraphPad Inc., San Diego, CA, USA). Results are shown as box plots. Boxes depict interquartile ranges, horizontal lines within boxes depict medians, and whiskers depict maximum and minimum values. For cell culture experiments, human osteoblasts were used in duplicates with a minimum of four independent donors. While gene expression results are depicted as percentage of 2(−ΔΔCt) for a better visualization of the changes with the untreated control (0 µM) set as 100%, the underlying statistical analysis was performed with the ΔCt values to allow the statistical comparison to the untreated control (0 µM). For the statistical analysis of protein data the values of the specific protein amount normalized to total protein content were used.
Statistical comparisons regarding the influence metal salts at different concentrations on cellular activity, gene expression, and protein synthesis were performed with repeated measures (RM) two-way analysis of variance (ANOVA) with “type of metal salt” and “concentration” as variables. Tukey’s multiple comparisons test was used for post hoc testing. Results after particle exposure (positive control) were compared to untreated samples (negative control) by either paired t-test or Wilcoxon matched-pairs test depending on normal distribution of data according to Shapiro-Wilk testing. The effects of the different cell culture media were analyzed with RM two-way ANOVA with “type of media” and “concentration” as variables. Post hoc testing was performed with Bonferroni’s multiple comparison test. Significances were set to a p-value less than 0.05. Further details of statistical tests are indicated in the results section and the figure legends.
4. Discussion
The corrosion of metal implants can lead to the release of free ions, which affect cellular behavior either locally or systemically. In this study, we analyzed the effects of different Co and Cr ions on osteoblastic survival, metabolism, and inflammation potential and compared the results with CoCr28Mo6 particle-exposed cells in order to assess the grade of toxicity. Since Ni is a common component in metal alloys, we additionally treated cells with Ni ions. The main purpose here was to compare the effects of different bivalent ions (Ni and Co) on the above mentioned cell processes.
For the experiments, we used Co(2+), Cr(3+) and Ni(2+) ions derived from metal salts. The tested concentrations were similar to those found in periprosthetic soft tissue [
18] but lower than those reported for mineralized bone tissue [
22]. The authors explained the relative high values in the mineralized bone tissue with the fact that the tissue was derived from the immediate vicinity, i.e., within a few millimeters, of the implant. This already highlights the difficulty to determine the relevant concentrations for in vitro-testing as these might differ depending on the distance from the implant but also depend on the conditions that aid corrosion. Also, when analyzing the in vivo concentrations, there is no differentiation whether the metals occur in form of ions or particles. Thus, our tested concentrations can only represent a rough estimate of the in vivo situation. Within the applied concentration range we were able to show that Co(2+) had a concentration-dependent influence on cell activity. Although the cell number remained (
Figure S1) almost stable, an increase of metabolic activity (
Figure S2) was initially detectable between 10 µM and 100 µM ion treatment. This effect can be explained by the fact that Co(2+) is able to surmount the cell membrane via membrane transport systems similar to Ca(2+). Intracellular, cobalt can then trigger hypoxia-like reactions in the cell, which lead to the upregulation of HIF-associated genes resulting in enhanced glycolytic reactions [
31]. However, the highest Co(2+) concentration (500 µM) seemed to be more cytotoxic, not least because of a decline in cell number (
Figure S1). The cytotoxicity of Co(2+) has been associated with enhanced oxidative stress, the formation of ROS, and DNA damage [
2,
32]. Consequentially, apoptosis or necrosis occurs [
2,
32] which was also detectable in our previous study [
21]. Additionally, the toxic effects might be further explained by the inhibition of Ca(2+) entry and Ca(2+) signaling, leading to the intracellular competition of Co(2+) and Ca(2+) with intracellular Ca(2+) binding proteins affecting cellular behavior [
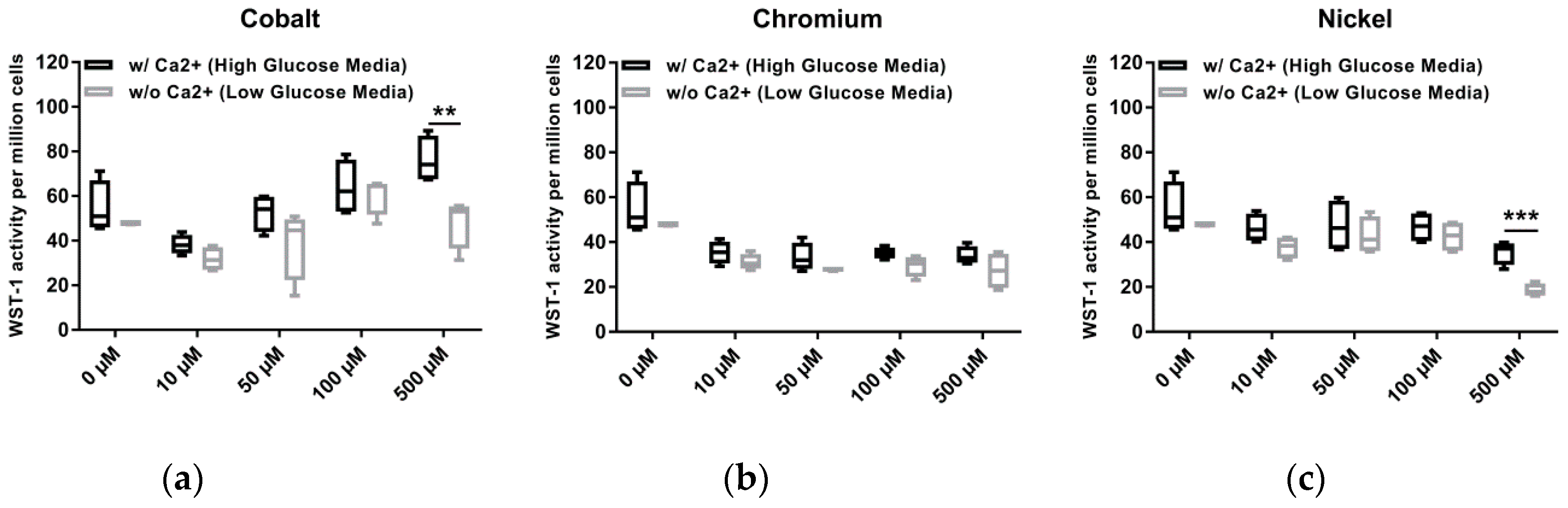
6]. The results of our comparative analysis of Ca-containing and –depleted media might reflect the effect described by Scharf et al. (2014) [
6]. Osteoblasts in Ca-containing medium showed increased metabolic activity compared to cells incubated in calcium-depleted medium after exposure to 500 µM Co(2+). We assume that the increased glucose content of the calcium-containing medium did not cause the increased metabolic activity, since no differences in cell activity were detected both in untreated cells and at the lower Co(2+) concentrations. As Co(2+) cannot be actively transported out of the cell, intracellular accumulation and cytotoxicity occurs [
31]. Thus, in consideration of our results we rather hypothesize that, despite the increased presence of Co(2+), the cells prefer the Ca influx in Ca-enriched medium, resulting in a reduced intracellular Co(2+) accumulation, fewer cytotoxic effects and restored/increased metabolic activity. Bivalent Ni ions might affect osteoblasts via the same mechanism, but are however more cytotoxic than Co(2+). This was reflected by our observation of reduced cell numbers, especially at 500 µM and a loss of the actin filament structure.
In comparison to bivalent metal ions, the cell number was not affected by Cr(3+). However, a decrease in cell metabolism was detectable without concentration-dependent differences. This effect can be explained by the fact that Cr(3+) ions cannot enter the cell membrane [
33] and therefore accumulate at the cell membrane [
34], which may have an effect on cell metabolism. On the other hand, Magone et al. (2015) described the possibility of Cr(3+) oxidation to Cr(4+) in order to enter the cell membrane [
7]. Intracellular, Cr(4+) conjugates with proteins resulting in reduction of Cr(4+) to Cr(3+) [
7,
32]. Although Shrivastava et al. (2002) mentioned that cell degeneration, DNA damage, and ROS accumulation is assigned to be the result of Cr(4+) exposure [
23], other studies have revealed a release of ROS in the presence of Cr(3+) not least because of the rapid Cr (4+) reduction [
6,
7,
32]. Additionally, cellular damage and the induction of apoptotic pathways by Cr(3+) were also described by Rudolf and Cervinka (2009) [
35].
Exposure to CoCr28Mo6 particles resulted in unaffected cell activity compared to untreated cells. Compared to ion exposure, particle treatment led to an activity level similar to 100 µM Co(2+) or to obviously enhanced levels compared to Cr(3+). One simple explanation for this result can be the differing mechanism for the uptake and intracellular availability of ions and particles which directly affect cellular activity. As mentioned before, ions can enter the cell membrane via different ion-mediated transporter systems while particles have to be recognized by receptors or actively incorporated via phagocytosis [
29,
32]. On the other hand, the corrosion of particles within the cell culture media, or by cell-mediated mechanisms including phagocytosis, cannot be excluded. Thus, the released ions from the particles would have the same effect as Co(2+) ions from the metal salts on triggering the previously mentioned hypoxia-like reactions. However, the corrosion-mediated release of ions in the cell culture medium has not been examined yet, but will be the subject of further investigations.
A limitation for the determination of metabolic activity via WST-1 might be that this assay is influenced by the presence of superoxide, which is associated with phagocytosis and oxidative burst [
36]. Therefore, the results for metabolic activity can also reflect the presence of ROS, as indeed WST-1 was used for the measurement of superoxide production [
36]. However, WST-1 was performed in ion-free media, so we assumed that most of the results were caused by the reduction of formazan via the mitochondrial respiratory chain. Future experiments will establish whether the changes observed in metabolic activity after particle and metal ion exposure are linked to ROS production in our cells.
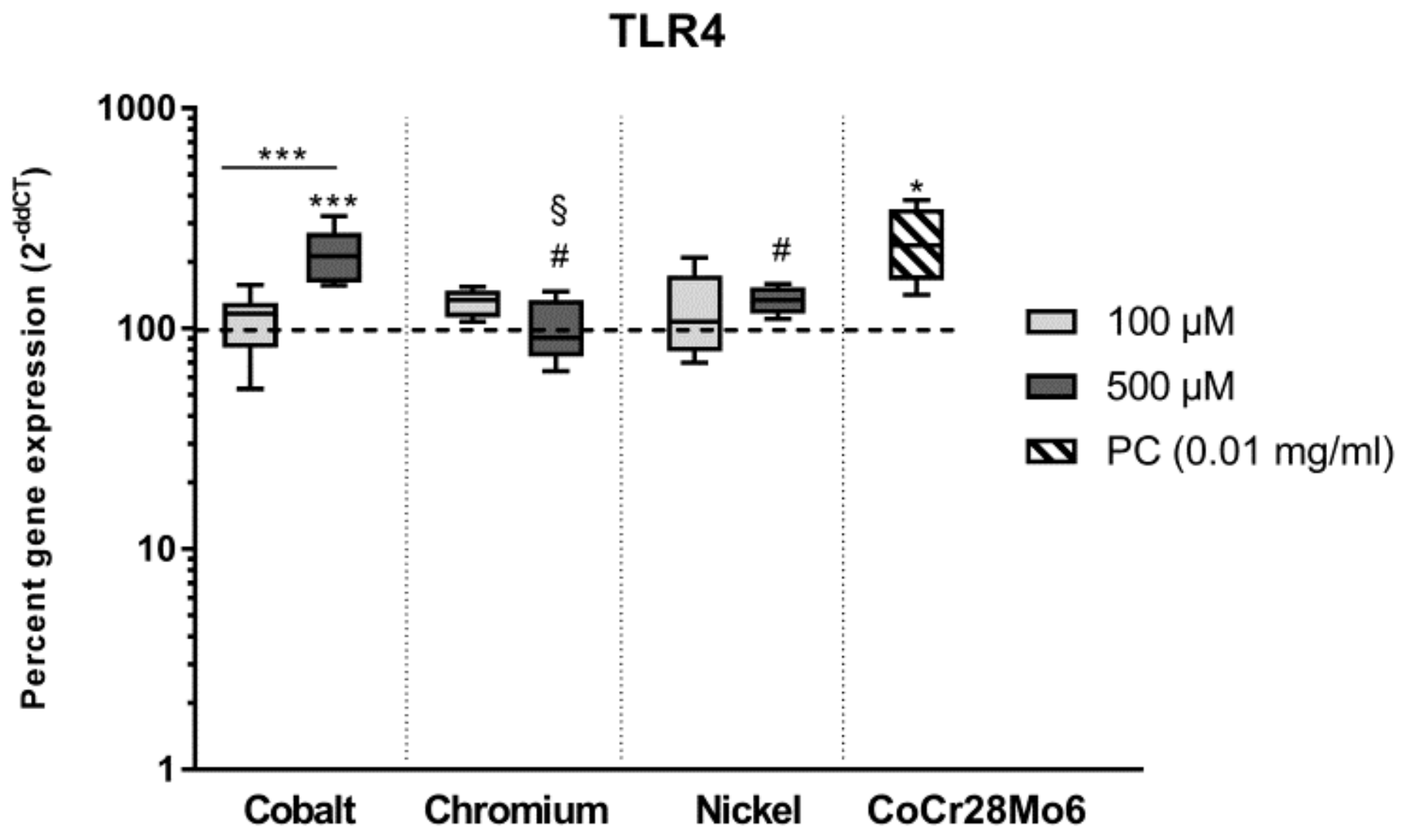
Intracellular downstream processes, including ROS production and oxidative burst, can be induced by a pathogen or substance binding on surface receptors. Here, toll-like receptors (TLR) recognize a wide spectrum of exogenous and endogenous danger signals [
30] and therefore play a central role in the induction of intracellular cascades, especially in macrophages. Since TLR 4 is known to be the most relevant receptor in aseptic implant loosening [
30,
37], we looked at TLR4 sensitization in human osteoblasts after exposure to metal salts and controls. We initially assumed that CoCr28Mo6 particles primarily induce TLR4 expression and further downstream processes [
38] while ions enter the cell membrane via channels and initiate immunological reactions. However, our results indicated that metal ions, especially Co(2+), have almost the same impact on
TLR4 gene expression as particles. It seems likely that the increased release of IL6 and IL8 can be directly linked to TLR4 activation. This is also supported by
TLR4 and IL6/IL8 results of Cr(3+) and Ni(2+) exposed cells. Lawrence and co-workers already showed, that Co(2+) activates TLR 4 which is in turn associated with enhanced chemokine release [
39,
40]. In contrast, Samelko et al. (2016) reported that CoCr particles did not preferentially activate TLR4 induced inflammation [
41]. This is supported by our previous [
28] as well as our current findings of reduced IL6 and IL8 secretion after particle exposure. Here, the release of pro-osteolytic mediators might be induced by other downstream mechanism such as inflammasome activation [
41,
42]. However, an impact of metal ions on inflammasome activation cannot be excluded as a strong link between ion channel expression and inflammasome activation has been described [
43]. Thus, to finally prove the link between our findings of ion-mediated IL6 and IL8 release and TLR4 activation, further studies have been carried out, especially for the quantification of TLR4 on the surface of osteoblastic cells.
Osteoblasts exposed to metal ions and CoCr28Mo6 particles showed reduced capacity of differentiation which was almost in accordance to our previous work [
21,
28]. Interestingly, Drynda et al. (2018) reported in their study that Cr(3+) instead of Co(2+) led to a concentration-dependent inhibition (10 μM to 250 μM) of osteoblastic mineralization [
22]. This fact could not be proven in our work, since we found significantly reduced osteogenic differentiation at higher Co(2+) concentration, whereas Cr(3+) had significantly less (Col1 protein) or no effects (
Col1A1, ALP). Our present data show that not only CoCr28Mo6 particles affect extracellular matrix production and hence bone formation processes, but additionally the presence of bivalent metal ions can inhibit collagen type 1 synthesis. Bone degradation and osteolysis can be further advanced by altered OPG/RANKL ratios as these affect osteoclastogenesis and bone resorption. Here, it should be noted that OPG prevents bone loss while RANKL promotes osteoclastic resorption [
7]. Our data indicated that mainly bivalent ions have a significant impact on OPG/RANKL ratios. Although OPG protein concentrations were clearly elevated compared to RANKL, concentration-dependent effects were found: while OPG was significantly downregulated at 500 µM, RANKL protein was significantly upregulated. As a result, OPG/RANKL ratio at 500 μM was only about one tenth of the ratio at 100 μM. This was in contrast to the study of Zijlstra et al. (2012) who revealed a time-dependent effect of Co and Cr ions on
OPG/RANKL mRNA ratio without concentration-dependent differences [
44]. However, they only calculated ratios via gene expression results and not on protein data, which is important to demonstrate osteolytic reactions in the periprosthetic tissue. In our study, we were not able to detect
OPG and
RANKL transcripts although we used clearly lower metal ion concentrations. Nonetheless, our protein data can provide important information about the effects of ion exposure on bone remodeling. In this context, co-cultures should be carried out in the future to prove the effects of osteoclastogenesis and further bone resorption as already shown in our previous study [
21].

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}