

Optimization of Fermentation Parameters for Enhanced Bioethanol Production by Multistress-Tolerant Saccharomycodes ludwigii APRE2 Using Undetoxified Sugarcane Bagasse Hydrolysate

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Microbial Strain and Culture Conditions
2.3. Sugarcane Bagasse Hydrolysate (SCH) Preparation
2.4. Ethanol Production from Acid and Enzymatic Hydrolysate of Sugarcane Bagasse
2.5. Optimization Conditions for Ethanol Production from SBH
2.6. Analytical Methods and Statistical Analysis
3. Results and Discussion
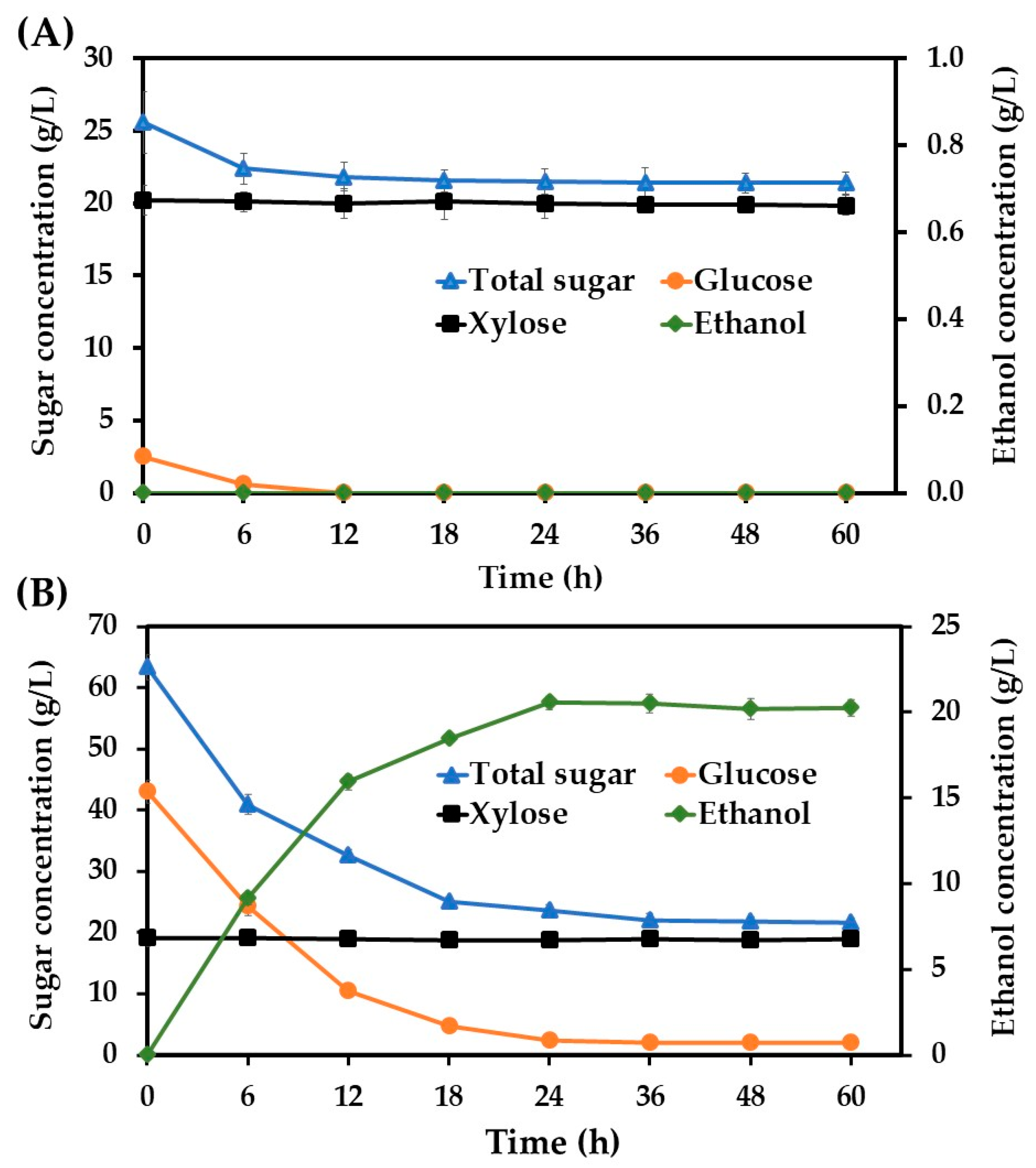
3.1. Ethanol Production Potential of S. ludwigii Using Acid and Enzymatic Sugarcane Bagasse Hydrolysate as Feedstock
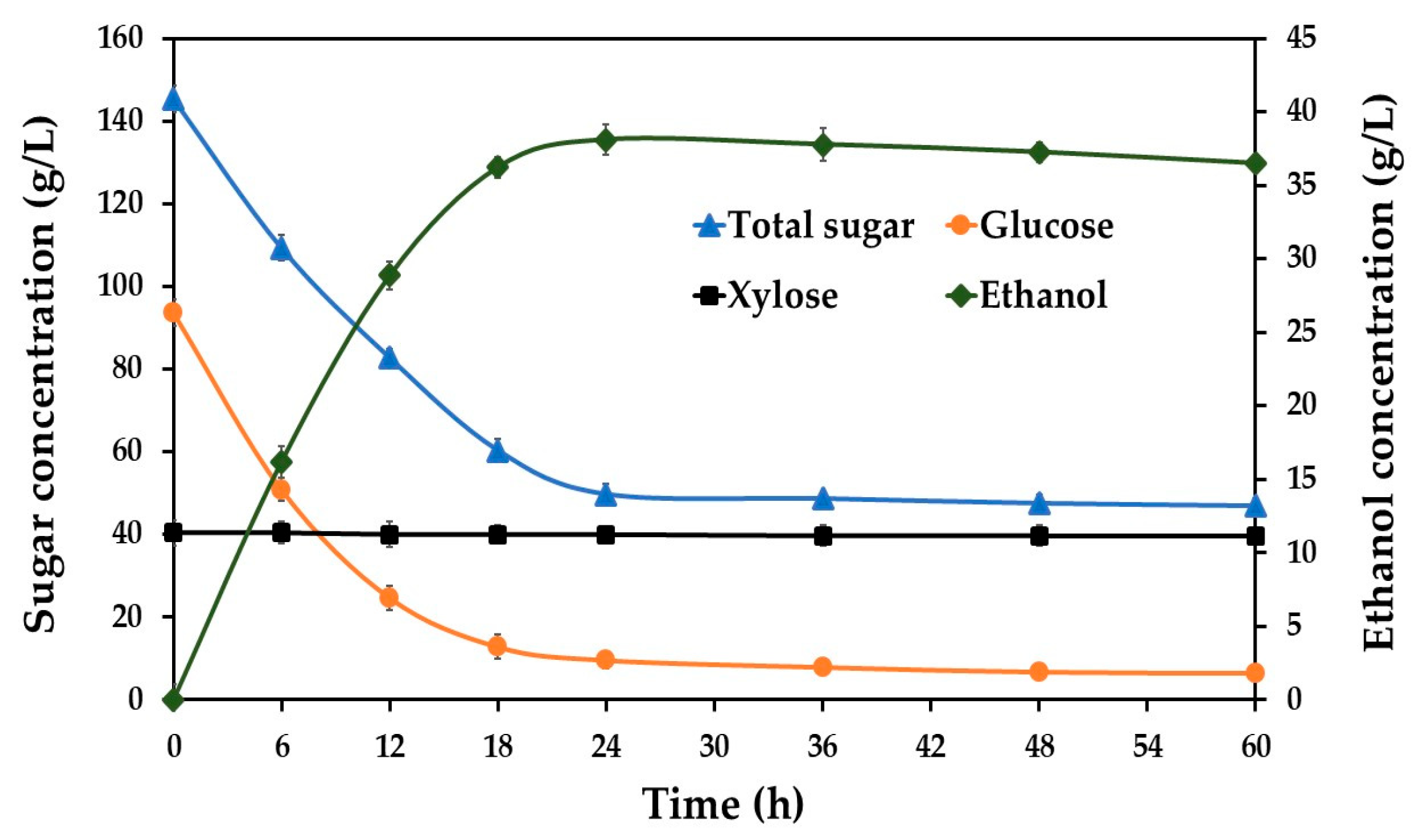
3.2. Optimization Conditions for Ethanol Production from SBH
0.24AE − 0.48BC + 0.21BD − 0.19BE − 0.15CD − 0.10CE − 0.003DE − 4.74A2 − 0.97B2 −
3.93C2 − 1.62D2 − 1.61E2

{kind=link}
{kind=link}
{kind=link}
| Microbe | Feedstock | Fermentation Parameter 1 | References | |||||
|---|---|---|---|---|---|---|---|---|
| S (g/L) | T (°C) | P (g/L) | Qp (g/L·h) | TY (%) | T (h) | |||
| P. kudriavzevii RZ8-1 | Sugarcane bagasse 2 | 85 | 37 | 35.51 | 1.48 | 81.75 | 24 | [17] |
| 85 | 40 | 33.84 | 1.41 | 77.91 | 24 | |||
| P. kudriavzevii RGB3.2 | Rice straw | 19.10 | 40 | 9.32 | 0.39 | 95.49 | 24 | [16] |
| K. marxianus RGB4.5 | Rice straw | 19.10 | 40 | 8.03 | 0.33 | 82.27 | 24 | |
| P. fermentans MTCC189 | Wheat straw | 172.00 | 30 | 92.00 | 0.55 | 62.1 | 168 | [68] |
| K. marxianus CECT10875 | Woody and herbaceous biomass | NR | 42 | 19.0 | NR | 71.2 | NR | [64] |
| S. cerevisiae | Coffee pulp | NR | 30 | 71.39 | 0.99 | NR | 72 | [69] |
| Wheat bran | NR | 30 | 68.91 | 0.96 | NR | 72 | ||
| S. cerevisiae | Pomelo peel waste | NR | 30 | 36.00 | 0.75 | 73.50 | 48 | [67] |
| S. cerevisiae TC-5 | Corncob residue 3 | NR | 40 | 31.96 | 0.22 | NR | 144 | [65] |
| S. cerevisiae PTCC5052 | Wheat straw | NR | 25 | 24.02 | 0.25 | NR | 96 | [66] |
| S. ludwigii APRE2 | Pineapple waste | 105.65 | 37 | 38.02 | 1.58 | 82.35 | 24 | [22] |
| S. ludwigii APRE2 | Sugarcane bagasse 4 | 143.95 | 37 | 38.11 | 1.59 | 88.24 | 24 | This study |
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. Available online: https://ourworldindata.org/grapher/sugar-cane-production?country=USA~BRA~IND~CHN~European+Union~THA (accessed on 19 April 2025).
- Junpen, A.; Pansuk, J.; Garavait, S. Estimation of reduced air emissions as a result of the implementation of the measure to reduce burned sugarcane in Thailand. Atmosphere 2020, 11, 366. [Google Scholar] [CrossRef]
- Pereira, S.C.; Maehara, L.; Machado, C.M.M.; Farinas, C.S. 2G ethanol from the whole sugarcane lignocellulosic biomass. Biotechnol. Biofuels 2015, 8, 44. [Google Scholar] [CrossRef] [PubMed]
- de Souza, R.F.R.; Dutra, E.D.; Leite, F.C.B.; Cadete, R.M.; Rosa, C.A.; Stambuk, B.U.; Stamford, T.L.M.; de Morais, M.A., Jr. Production of ethanol fuel from enzyme-treated sugarcane bagasse hydrolysate using d-xylose-fermenting wild yeast isolated from Brazilian biomes. 3 Biotech 2018, 8, 312. [Google Scholar]
- de Araujo Guilherme, A.; Dantas, P.V.F.; Padilha, C.E.A.; Dos Santos, E.S.; de Macedo, G.R. Ethanol production from sugarcane bagasse: Use of different fermentation strategies to enhance an environmental-friendly process. J. Environ. Manag. 2019, 234, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Chamnipa, N.; Klanrit, P.; Thanonkeo, S.; Thanonkeo, P. Sorbitol production from a mixture of sugarcane bagasse and cassava pulp hydrolysates using thermotolerant Zymomonas mobilis TISTR548. Ind. Crops Prod. 2022, 188, 115741. [Google Scholar] [CrossRef]
- Dolpatcha, S.; Phong, H.X.; Thanonkeo, S.; Klanrit, P.; Yamada, M.; Thanonkeo, P. Adaptive laboratory evolution under acetic acid stress enhances the multistress tolerance and ethanol production efficiency of Pichia kudriavzevii from lignocellulosic biomass. Sci. Rep. 2023, 13, 21000. [Google Scholar] [CrossRef]
- Yusuf, A.A.; Inambao, F.L. Bioethanol production techniques from lignocellulose biomass as alternative fuel: A review. Int. J. Mech. Eng. Technol. 2019, 10, 34–71. [Google Scholar]
- Dussan, K.J.; Silva, D.D.V.; Moraes, E.J.C.; Arruda, P.V.; Felipe, M.G.A. Dilute-acid hydrolysis of cellulose to glucose from sugarcane bagasse. Chem. Eng. Trans. 2014, 38, 433–438. [Google Scholar]
- Azhar, S.H.M.; Abdulla, R.; Jambo, S.A.; Marbawi, H.; Gansau, J.A.; Faik, A.A.M.; Rodrigues, K.F. Yeasts in sustainable bioethanol production: A review. Biochem. Biophys. Rep. 2017, 10, 52–61. [Google Scholar]
- Sritrakul, N.; Nitisinprasert, S.; Keawsompong, S. Evaluation of dilute acid pretreatment for bioethanol fermentation from sugarcane bagasse pith. Agric. Nat. Resour. 2017, 51, 512–519. [Google Scholar] [CrossRef]
- Phong, H.X.; Klanrit, P.; Dung, N.T.P.; Yamada, M.; Thanonkeo, P. Isolation and characterization of thermotolerant yeasts for the production of second-generation bioethanol. Ann. Microbiol. 2019, 69, 765–776. [Google Scholar] [CrossRef]
- Sunkar, B.; Bhukya, B. Bi-phasic hydrolysis of corncobs for the extraction of total sugars and ethanol production using inhibitor resistant and thermotolerant yeast, Pichia kudriavzevii. Biomass Bioenergy 2021, 153, 106230. [Google Scholar] [CrossRef]
- Chandel, A.K.; Kapoor, R.K.; Singh, A.; Kuhad, R.C. Detoxification of sugarcane bagasse hydrolysate improves ethanol production by Candida shehatae NCIM 3501. Bioresour. Technol. 2007, 98, 1947–1950. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.H.; Park, E.H.; Kim, M.D. Isolation of thermotolerant yeast Pichia kudriavzevii from nuruk. Food Sci. Biotechnol. 2017, 26, 1357–1362. [Google Scholar] [CrossRef] [PubMed]
- Avchar, R.; Lanjekar, V.; Kshirsagar, P.; Dhakephalkar, P.K.; Dagar, S.S.; Baghela, A. Buffalo rumen harbours diverse thermotolerant yeasts capable of producing second-generation bioethanol from lignocellulosic biomass. Renew. Energy 2021, 173, 795–807. [Google Scholar] [CrossRef]
- Chamnipa, N.; Thanonkeo, S.; Klanrit, P.; Thanonkeo, P. The potential of the newly isolated thermotolerant yeast Pichia kudriavzevii RZ8-1 for high-temperature ethanol production. Braz. J. Microbiol. 2018, 49, 378–391. [Google Scholar] [CrossRef]
- Aouine, M.; Elalami, D.; Koraichi, S.I.; Haggoud, A.; Barakat, A. Exploring natural fermented foods as a source for new efficient thermotolerant yeasts for the production of second-generation bioethanol. Energies 2022, 15, 4954. [Google Scholar] [CrossRef]
- Wang, N.; Zhang, P.; Zhou, X.; Zheng, J.; Ma, Y.; Liu, C.; Wu, T.; Li, H.; Wang, X.; Wang, H.; et al. Isolation, identification, and characterization of an acid-tolerant Pichia kudriavzevii and exploration of its acetic acid tolerance mechanism. Fermentation 2023, 9, 540. [Google Scholar] [CrossRef]
- Charoensopharat, K.; Thanonkeo, P.; Thanonkeo, S.; Yamada, M. Ethanol production from Jerusalem artichoke tubers at high temperature by newly isolated thermotolerant inulin-utilizing yeast Kluyveromyces marxianus using consolidated bioprocessing. Antonie Van Leeuwenhoek 2015, 108, 173–190. [Google Scholar] [CrossRef]
- Hemansi; Himanshu; Patel, A.K.; Saini, J.K.; Singhania, R.R. Development of multiple inhibitor tolerant yeast via adaptive laboratory evolution for sustainable bioethanol production. Bioresour. Technol. 2022, 344, 126247. [Google Scholar] [CrossRef]
- Pilap, W.; Thanonkeo, S.; Klanrit, P.; Thanonkeo, P. The potential of multistress tolerant yeast, Saccharomycodes ludwigii, for second-generation bioethanol production. Sci. Rep. 2022, 12, 22062. [Google Scholar] [CrossRef]
- Loureiro, V.; Malfeito-Ferreira, M. Spoilage yeasts in the wine industry. Int. J. Food Microbiol. 2003, 86, 23–50. [Google Scholar] [CrossRef] [PubMed]
- Thammasittirong, S.N.; Chamduang, T.; Phonrod, U.; Sriroth, K. Ethanol production potential of ethanol-tolerant Saccharomyces and non-Saccharomyces yeasts. Pol. J. Microbiol. 2012, 61, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Udomsaksakul, N.; Kodama, K.; Tanasupawat, S.; Savarajara, A. Diversity of ethanol fermenting yeasts in coconut inflorescence sap and their application potential. Sci. Asia 2018, 44, 371–381. [Google Scholar] [CrossRef]
- Vejarano, R. Saccharomycodes ludwigii, control and potential uses in winemaking processes. Fermentation 2018, 4, 71. [Google Scholar] [CrossRef]
- Nuanpeng, S.; Thanonkeo, S.; Yamada, M.; Thanonkeo, P. Ethanol production from sweet sorghum juice at high temperatures using a newly isolated thermotolerant yeast Saccharomyces cerevisiae DBKKU Y-53. Energies 2016, 9, 253. [Google Scholar] [CrossRef]
- Nuanpeng, S.; Thanonkeo, S.; Klanrit, P.; Yamada, M.; Thanonkeo, P. Optimization conditions for ethanol production from sweet sorghum juice by thermotolerant yeast Saccharomyces cerevisiae: Using a statistical experimental design. Fermentation 2023, 9, 450. [Google Scholar] [CrossRef]
- Phong, H.X.; Klanrit, P.; Dung, N.T.P.; Thanonkeo, S.; Yamada, M.; Thanonkeo, P. High-temperature ethanol fermentation from pineapple waste hydrolysate and gene expression analysis of thermotolerant yeast Saccharomyces cerevisiae. Sci. Rep. 2022, 12, 13965. [Google Scholar] [CrossRef]
- Nguyen, P.V.; Nguyen, K.H.V.; Nguyen, N.L.; Ho, X.T.T.; Truong, P.H.; Nguyen, K.C.T. Lychee-derived, thermotolerant yeasts for second-generation bioethanol production. Fermentation 2022, 8, 515. [Google Scholar] [CrossRef]
- Pereira, F.; Guimarães, P.M.R.; Teixeira, J.A.; Domingues, L. Optimization of low-cost medium for very high gravity ethanol fermentations by Saccharomyces cerevisiae using statistical experimental designs. Bioresour. Technol. 2010, 101, 7856–7863. [Google Scholar] [CrossRef]
- Mecozzi, M. Estimation of total carbohydrate amount in environmental samples by the phenol−sulphuric acid method assisted by multivariate calibration. Chemom. Intell. Lab. Syst. 2005, 79, 84–90. [Google Scholar] [CrossRef]
- Laopaiboon, L.; Nuanpeng, S.; Srinophakun, P.; Klanrit, P.; Laopaiboon, P. Ethanol production from sweet sorghum juice using very high gravity technology: Effects of carbon and nitrogen supplementations. Bioresour. Technol. 2009, 100, 4176–4182. [Google Scholar] [CrossRef]
- Jönsson, L.J.; Martín, C. Pretreatment of lignocellulose: Formation of inhibitory by-products and strategies for minimizing their effects. Bioresour. Technol. 2016, 199, 103–112. [Google Scholar] [CrossRef]
- Chen, H.; Liu, J.; Chang, X.; Chen, D.; Xue, Y.; Liu, P.; Lin, H.; Han, S. A review on the pretreatment of lignocellulose for high-value chemicals. Fuel Process. Technol. 2017, 160, 196–206. [Google Scholar] [CrossRef]
- Galbe, M.; Wallberg, O. Pretreatment for biorefineries: A review of common methods for efficient utilisation of lignocellulosic materials. Biotechnol. Biofuels 2019, 12, 294. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Tang, J.; Chen, J.; Zhang, Q. Quantitative structure-toxicity relationship analysis of combined toxic effects of lignocellulose-derived inhibitors on bioethanol production. Bioresour. Technol. 2019, 289, 121724. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Zhao, Y.; Chang, J.S.; Lee, D.J. Inhibitor formation and detoxification during lignocellulose biorefinery: A review. Bioresour. Technol. 2022, 361, 127666. [Google Scholar] [CrossRef]
- Kumar, V.; Patel, S.K.S.; Gupta, R.K.; Otari, S.V.; Gao, H.; Lee, J.K.; Zhang, L. Enhanced saccharification and fermentation of rice straw by reducing the concentration of phenolic compounds using an immobilized enzyme cocktail. Biotechnol. J. 2019, 14, 1800468. [Google Scholar] [CrossRef]
- Almeida, J.R.; Modig, T.; Petersson, A.; Hagerdal, B.H.; Liden, G.; Grauslund, F.G. Increased tolerance and conversion of inhibitors in lignocellulosic hydrolysates by Saccharomyces cerevisiae. J. Chem. Technol. Biotechnol. 2007, 82, 340–349. [Google Scholar] [CrossRef]
- Narendranath, N.V.; Thomas, K.C.; Ingledew, W.M. Effects of acetic acid and lactic acid on the growth of Saccharomyces cerevisiae in a minimal medium. J. Ind. Microbiol. Biotechnol. 2001, 26, 171–177. [Google Scholar] [CrossRef]
- Casey, E.; Sedlak, M.; Ho, H.W.Y.; Mosier, N.S. Effect of acetic acid and pH on the cofermentation of glucose and xylose to ethanol by a genetically engineered strain of Saccharomyces cerevisiae. FEMS Yeast Res. 2010, 10, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Caspeta, L.; Castillo, T.; Nielsen, J. Modifying yeast tolerance to inhibitory conditions of ethanol production processes. Front. Bioeng. Biotechnol. 2015, 3, 184. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.; Palmqvist, E.; Hahn-Hägerdal, B.; Tengborg, C.; Stenberg, K.; Zacchi, G.; Nilvebrant, N. The generation of fermentation inhibitors during dilute acid hydrolysis of softwood. Enzym. Microb. Technol. 1999, 24, 151–159. [Google Scholar] [CrossRef]
- Hasunuma, T.; Sanda, T.; Yamada, R.; Yoshimura, K.; Ishii, J.; Kondo, A. Metabolic pathway engineering based on metabolomics confers acetic acid and formic acid tolerance to a recombinant xylose-fermenting strain of Saccharomyces cerevisiae. Microb. Cell Factories 2011, 10, 2. [Google Scholar] [CrossRef]
- Lin, F.M.; Qiao, B.; Yuan, Y.J. Comparative proteomic analysis of tolerant and adaptation of ethanologenic Saccharomyces cerevisiae to furfural, a lignocellulosic inhibitory compound. Appl. Environ. Microbiol. 2009, 11, 3765–3776. [Google Scholar] [CrossRef]
- Janzowski, C.; Glaab, V.; Samimi, E.; Schlatter, J.; Eisenbrand, G. 5-hydroxymethylfurfural: Assessment of mutagenicity, DNA damaging potential and reactivity towards cellular glutathione. Food Chem. Toxicol. 2000, 9, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Modig, T.; Liden, G.; Taherzadeh, M.J. Inhibition effects of furfural on alcohol dehydrogenase, aldehyde dehydrogenase and pyruvate dehydrogenase. Biochem. J. 2002, 32, 379–385. [Google Scholar] [CrossRef]
- Allen, S.A.; Clark, W.; McCaffery, J.M.; Cai, Z.; Lanctot, A.; Slininger, P.J.; Liu, Z.L.; Gorsich, S.W. Furfural induces reactive oxygen species accumulation and cellular damage in Saccharomyces cerevisiae. Biotechnol. Biofuels 2010, 3, 2. [Google Scholar] [CrossRef]
- Klinke, H.B.; Thomsen, A.B.; Ahring, B.K. Inhibition of ethanol-producing yeast and bacteria by degradation products produced during pretreatment of biomass. Appl. Microbiol. Biotechnol. 2004, 66, 10–26. [Google Scholar] [CrossRef]
- Jönsson, L.J.; Alriksson, B.; Nilvebrant, N.O. Bioconversion of lignocellulose: Inhibitors and detoxification. Biotechnol. Biofuels 2013, 6, 16. [Google Scholar] [CrossRef]
- Limtong, S.; Sringiew, C.; Yongmanitchai, W. Production of fuel ethanol at high temperature from sugar cane juice by a newly isolated Kluyveromyces marxianus. Bioresour. Technol. 2007, 98, 3367–3374. [Google Scholar] [CrossRef] [PubMed]
- Pilap, W.; Thanonkeo, S.; Klanrit, P.; Thanonkeo, P. The potential of the newly isolated thermotolerant Kluyveromyces marxianus for high-temperature ethanol production using sweet sorghum juice. 3 Biotech 2018, 8, 126. [Google Scholar] [CrossRef]
- Bai, F.W.; Chen, L.J.; Zhang, Z.; Anderson, W.A.; Moo-Young, M. Continuous ethanol production and evaluation of yeast cell lysis and viability loss under very high gravity medium conditions. J. Biotechnol. 2004, 110, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Ozmihci, S.; Kargi, F. Comparison of yeast strains for batch ethanol fermentation of cheese-whey powder (CWP) solution. Lett. Appl. Microbiol. 2007, 44, 602–606. [Google Scholar] [CrossRef]
- Palmqvist, E.; Hahnhägerdal, B. Fermentation of lignocellulosic hydrolysates. I: Inhibition and detoxification. Bioresour. Technol. 2000, 74, 17–24. [Google Scholar] [CrossRef]
- Liu, Z.L.; Slininger, P.J.; Dien, B.S.; Berhow, M.A.; Kurtzman, C.P.; Gorsich, S.W. Adaptive response of yeasts to furfural and 5-hydroxymethylfurfural and new chemical evidence for HMF conversion to 2,5-bis-hydroxymethylfuran. J. Ind. Microbiol. Biotechnol. 2004, 31, 345–352. [Google Scholar] [CrossRef]
- Palmqvist, E.; Hahnhägerdal, B. Fermentation of lignocellulosic hydrolysates. II: Inhibitors and mechanisms of inhibition. Bioresour. Technol. 2000, 74, 25–33. [Google Scholar] [CrossRef]
- Li, B.; Liu, N.; Zhao, X. Response mechanisms of Saccharomyces cerevisiae to the stress factors present in lignocellulose hydrolysate and strategies for constructing robust strains. Biotechnol. Biofuels Bioprod. 2022, 15, 28. [Google Scholar] [CrossRef]
- Walker, G.M.; Basso, T.O. Mitigating stress in industrial yeasts. Fungal Biol. 2020, 124, 387–397. [Google Scholar] [CrossRef]
- Eardley, J.; Timson, D.J. Yeast cellular stress: Impacts on bioethanol production. Fermentation 2020, 6, 109. [Google Scholar] [CrossRef]
- Hohmann, S. Osmotic stress signaling and osmoadaptation in yeasts. Microbiol. Mol. Biol. Rev. 2002, 66, 300–372. [Google Scholar] [CrossRef] [PubMed]
- Mager, W.H.; de Boer, A.H.; Siderius, M.H.; Voss, H.-P. Cellular responses to oxidative and osmotic stress. Cell Stress Chaperones 2000, 5, 73. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, I.; Oliva, J.M.; Negro, M.J.; Manzanares, P.; Ballesteros, M. Enzymic hydrolysis of steam exploded herbaceous agricultural waste (Brassica carinata) at different particule sizes. Process Biochem. 2002, 38, 187–192. [Google Scholar] [CrossRef]
- Boonchuay, P.; Techapun, C.; Leksawasdi, N.; Seesuriyachan, P.; Hanmoungjai, P.; Watanabe, M.; Chaiyaso, T. Bioethanol production from cellulose-rich corncob residue by the thermotolerant Saccharomyces cerevisiae TC-5. J. Fungi. 2021, 7, 547. [Google Scholar] [CrossRef]
- Valamonfared, J.; Javanmard, A.S.; Ghaedi, M.; Bagherinasab, M. Bioethanol production using lignocellulosic materials and thermophilic microbial hydrolysis. Biomass Convers. Biorefinery 2024, 14, 16589–16601. [Google Scholar] [CrossRef]
- Huang, R.; Cao, M.; Guo, H.; Qi, W.; Su, R.; He, Z. Enhanced ethanol production from pomelo peel waste by integrated hydrothermal treatment, multienzyme formulation, and fed-batch operation. J. Agric. Food Chem. 2014, 62, 4643–4651. [Google Scholar] [CrossRef]
- Shrivastava, A.; Sharma, R.K. Conversion of lignocellulosic biomass: Production of bioethanol and bioelectricity using wheat straw hydrolysate in electrochemical bioreactor. Heliyon 2023, 9, e12951. [Google Scholar] [CrossRef]
- Alabdalall, A.H.; Almutari, A.A.; Aldakeel, S.A.; Albarrag, A.M.; Aldakheel, L.A.; Alsoufi, M.H.; Alfuraih, L.Y.; Elkomy, H.M. Bioethanol production from lignocellulosic biomass using Aspergillus niger and Aspergillus flavus hydrolysis enzymes through immobilized S. cerevisiae. Energies 2023, 16, 823. [Google Scholar] [CrossRef]
- Jeffries, T.W. Engineering yeasts for xylose metabolism. Curr. Opin. Biotechnol. 2006, 17, 320–326. [Google Scholar] [CrossRef]
- Kuhad, R.C.; Gupta, R.; Khasa, Y.P.; Singh, A.; Zhang, Y.H.P. Bioethanol production from pentose sugars: Current status and future prospects. Renew. Sustain. Energy Rev. 2011, 15, 4950–4962. [Google Scholar] [CrossRef]
- Avanthi, A.; Kumar, S.; Sherpa, K.C.; Banerjee, R. Bioconversion of hemicelluloses of lignocellulosic biomass to ethanol: An attempt to utilize pentose sugars. Biofuels 2017, 8, 431–444. [Google Scholar] [CrossRef]
- Patelski, A.M.; Kobalczyk, A.; Dziekońska-Kubczak, U.; Januszewicz, B.; Domański, J. Effect of ultrasound on fermentation of thick molasses worts by distiller’s yeast. Appl. Sci. 2025, 15, 3811. [Google Scholar] [CrossRef]
- Saini, P.; Beniwal, A.; Kokkiligadda, A.; Vij, S. Response and tolerance of yeast to changing environmental stress during ethanol fermentation. Process Biochem. 2018, 72, 1–12. [Google Scholar] [CrossRef]
- Takagi, H. Molecular mechanisms and highly functional development for stress tolerance of the yeast Saccharomyces cerevisiae. Biosci. Biotechnol. Biochem. 2021, 85, 1017–1037. [Google Scholar] [CrossRef]
- Postaru, M.; Tucaliuc, A.; Cascaval, D.; Galaction, A.I. Cellular stress impact on yeast activity in biotechnological processes-A short overview. Microorganisms 2023, 11, 2522. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Wang, W.B.; Wang, Y.T.; Zhao, X.Q. Regulatory mechanisms underlying yeast chemical stress response and development of robust strains for bioproduction. Curr. Opin. Biotechnol. 2024, 86, 103072. [Google Scholar] [CrossRef]
- Xu, J.R.; Mehmood, M.A.; Wang, L.; Ahmad, N.; Ma, H.J. Omics-based strategies to explore stress tolerance mechanisms of Saccharomyces cerevisiae for efficient fuel ethanol production. Front. Energy Res. 2022, 10, 884582. [Google Scholar] [CrossRef]
- Topaloğlu, A.; Esen, Ö.; Turanli-Yildiz, B.; Arslan, M.; Çakar, Z.P. From Saccharomyces cerevisiae to ethanol: Unlocking the power of evolutionary engineering in metabolic engineering applications. J. Fungi 2023, 9, 984. [Google Scholar] [CrossRef]
- Gasch, A.P.; Spellman, P.T.; Kao, C.M.; Carmel-Harel, O.; Eisen, M.B.; Storz, G.; Botstein, D.; Brown, P.O. Genomic expression programs in the response of yeast cells to environmental changes. Mol. Biol. Cell 2000, 11, 4241–4257. [Google Scholar] [CrossRef]
- Kubiak-Szymendera, M.; Pryszcz, L.P.; Bialas, W.; Celinska, E. Epigenetic response of Yarrowia lipolytica to stress: Tracking methylation level and search for methylation patterns via whole-genome sequencing. Microorganisms 2021, 9, 1798. [Google Scholar] [CrossRef]
- Vázquez, J.; Grillitsch, K.; Daum, G.; Mas, A.; Beltran, G.; Torija, M.J. The role of the membrane lipid composition in the oxidative stress tolerance of different wine yeasts. Food Microbiol. 2019, 78, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.A.; Bourbon-Melo, N.; Sá-Correia, I. The cell wall and the response and tolerance to stresses of biotechnological relevance in yeasts. Front. Microbiol. 2022, 13, 953479. [Google Scholar] [CrossRef] [PubMed]
- Gan, Y.; Qi, X.; Lin, Y.; Guo, Y.; Zhang, Y.; Wang, Q. A hierarchical transcriptional regulatory network required for long-term thermal stress tolerance in an industrial Saccharomyces cerevisiae strain. Front. Bioeng. Biotechnol. 2022, 9, 826238. [Google Scholar] [CrossRef] [PubMed]
- Özel, A.; Topaloğlu, A.; Esen, Ö.; Holyavkin, C.; Baysan, M.; Çakar, Z.P. Transcriptomic and physiological meta-analysis of multiple stress-resistant Saccharomyces cerevisiae strains. Stresses 2024, 4, 714–733. [Google Scholar] [CrossRef]
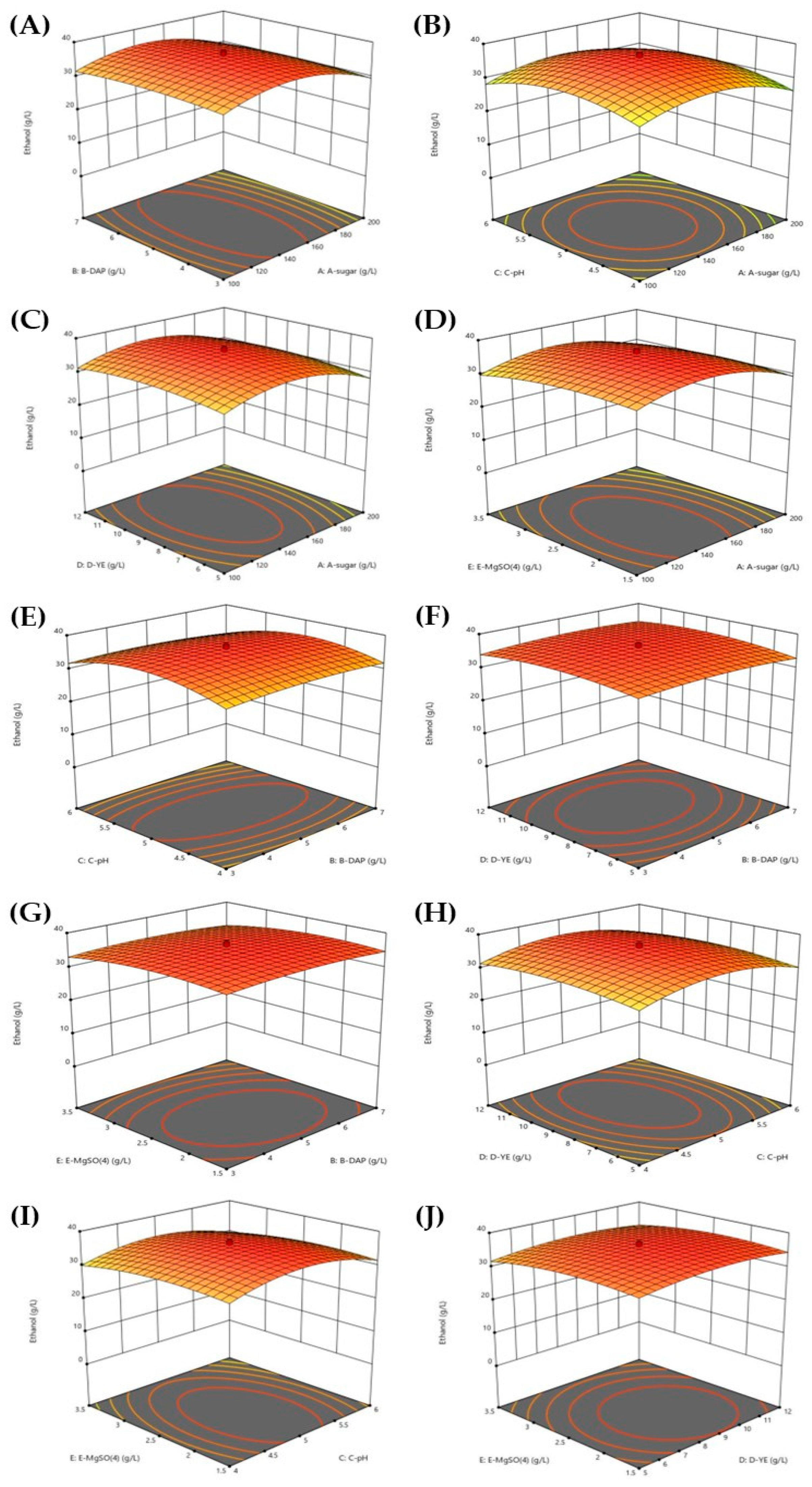
| Code | Variable | Unit | Level | ||||
|---|---|---|---|---|---|---|---|
| −2.37 | −1 | 0 | +1 | +2.37 | |||
| A | Sugar concentration | g/L | 31.08 | 100.00 | 150.00 | 200.00 | 268.92 |
| B | DAP | g/L | 0.24 | 3.00 | 5.00 | 7.00 | 9.76 |
| C | pH | - | 2.62 | 4.00 | 5.00 | 6.00 | 7.38 |
| D | Yeast extract | g/L | 0.18 | 5.00 | 8.50 | 12.00 | 16.82 |
| E | MgSO4 | g/L | 0.12 | 1.50 | 2.50 | 3.50 | 4.88 |
| Run | A-Sugar (g/L) | B-DAP (g/L) | C-pH | D-YE (g/L) | E-MgSO4 (g/L) | Ethanol Concentration (P, g/L) | |
|---|---|---|---|---|---|---|---|
| Predicted Value | Actual Value | ||||||
| 1 | 200.00 | 3.00 | 4.0 | 12.00 | 1.50 | 22.70 | 20.34 |
| 2 | 150.00 | 5.00 | 5.0 | 16.82 | 2.50 | 28.08 | 27.40 |
| 3 | 150.00 | 5.00 | 5.0 | 8.50 | 4.88 | 25.04 | 23.14 |
| 4 | 100.00 | 3.00 | 6.0 | 12.00 | 3.50 | 23.54 | 24.06 |
| 5 | 200.00 | 7.00 | 6.0 | 5.00 | 3.50 | 20.27 | 20.81 |
| 6 | 200.00 | 7.00 | 6.0 | 5.00 | 1.50 | 22.14 | 21.59 |
| 7 | 200.00 | 7.00 | 4.0 | 5.00 | 3.50 | 20.93 | 19.89 |
| 8 | 150.00 | 5.00 | 5.0 | 8.50 | 2.50 | 36.26 | 36.62 |
| 9 | 150.00 | 5.00 | 5.0 | 8.50 | 2.50 | 36.26 | 35.80 |
| 10 | 100.00 | 7.00 | 4.0 | 12.00 | 3.50 | 24.69 | 25.10 |
| 11 | 100.00 | 3.00 | 4.0 | 12.00 | 3.50 | 23.85 | 24.34 |
| 12 | 100.00 | 3.00 | 4.0 | 12.00 | 1.50 | 25.55 | 26.20 |
| 13 | 100.00 | 7.00 | 6.0 | 12.00 | 1.50 | 25.30 | 25.04 |
| 14 | 150.00 | 5.00 | 2.6 | 8.50 | 2.50 | 14.37 | 13.35 |
| 15 | 200.00 | 3.00 | 4.0 | 5.00 | 3.50 | 21.03 | 22.76 |
| 16 | 200.00 | 3.00 | 6.0 | 5.00 | 3.50 | 22.28 | 20.40 |
| 17 | 200.00 | 3.00 | 6.0 | 12.00 | 1.50 | 23.75 | 24.60 |
| 18 | 150.00 | 5.00 | 5.0 | 8.50 | 0.12 | 29.28 | 30.20 |
| 19 | 100.00 | 7.00 | 6.0 | 5.00 | 3.50 | 21.73 | 23.56 |
| 20 | 100.00 | 3.00 | 4.0 | 5.00 | 1.50 | 25.08 | 25.82 |
| 21 | 200.00 | 7.00 | 4.0 | 12.00 | 3.50 | 22.71 | 23.57 |
| 22 | 150.00 | 5.00 | 5.0 | 0.18 | 2.50 | 26.10 | 25.80 |
| 23 | 31.08 | 5.00 | 5.0 | 8.50 | 2.50 | 12.00 | 10.02 |
| 24 | 100.00 | 3.00 | 4.0 | 5.00 | 3.50 | 23.38 | 23.15 |
| 25 | 100.00 | 3.00 | 6.0 | 12.00 | 1.50 | 25.63 | 26.04 |
| 26 | 100.00 | 7.00 | 4.0 | 12.00 | 1.50 | 27.14 | 28.40 |
| 27 | 200.00 | 3.00 | 4.0 | 5.00 | 1.50 | 21.76 | 21.50 |
| 28 | 150.00 | 9.76 | 5.0 | 8.50 | 2.50 | 30.54 | 29.80 |
| 29 | 268.92 | 5.00 | 5.0 | 8.50 | 2.50 | 6.87 | 7.87 |
| 30 | 200.00 | 7.00 | 4.0 | 5.00 | 1.50 | 22.42 | 22.80 |
| 31 | 100.00 | 7.00 | 4.0 | 5.00 | 3.50 | 23.37 | 23.90 |
| 32 | 150.00 | 5.00 | 7.4 | 8.50 | 2.50 | 13.67 | 13.71 |
| 33 | 200.00 | 7.00 | 6.0 | 12.00 | 3.50 | 21.46 | 21.80 |
| 34 | 200.00 | 3.00 | 4.0 | 12.00 | 3.50 | 21.95 | 22.90 |
| 35 | 150.00 | 5.00 | 5.0 | 8.50 | 2.50 | 36.26 | 37.03 |
| 36 | 150.00 | 5.00 | 5.0 | 8.50 | 2.50 | 36.26 | 35.90 |
| 37 | 200.00 | 7.00 | 4.0 | 12.00 | 1.50 | 24.20 | 24.19 |
| 38 | 200.00 | 7.00 | 6.0 | 12.00 | 1.50 | 23.34 | 22.75 |
| 39 | 150.00 | 0.24 | 5.0 | 8.50 | 2.50 | 31.04 | 30.80 |
| 40 | 100.00 | 7.00 | 4.0 | 5.00 | 1.50 | 25.81 | 25.76 |
| 41 | 200.00 | 3.00 | 6.0 | 5.00 | 1.50 | 23.40 | 23.88 |
| 42 | 200.00 | 3.00 | 6.0 | 12.00 | 3.50 | 22.62 | 22.42 |
| 43 | 100.00 | 3.00 | 6.0 | 5.00 | 1.50 | 25.74 | 24.65 |
| 44 | 100.00 | 7.00 | 6.0 | 12.00 | 3.50 | 22.46 | 22.38 |
| 45 | 150.00 | 5.00 | 5.0 | 8.50 | 2.50 | 36.26 | 35.30 |
| 46 | 100.00 | 3.00 | 6.0 | 5.00 | 3.50 | 23.65 | 25.02 |
| 47 | 100.00 | 7.00 | 6.0 | 5.00 | 1.50 | 24.56 | 24.40 |
| Source | Sum Square | df | Mean Square | F-Value | p-Value * | Remark |
|---|---|---|---|---|---|---|
| Model | 1619.550 | 20 | 80.980 | 51.540 | <0.0001 | Significant |
| A | 50.420 | 1 | 50.420 | 32.090 | <0.0001 | |
| B | 0.471 | 1 | 0.471 | 0.300 | 0.5885 | |
| C | 0.935 | 1 | 0.935 | 0.595 | 0.4474 | |
| D | 7.520 | 1 | 7.520 | 4.790 | 0.0379 | |
| E | 34.560 | 1 | 34.560 | 22.000 | <0.0001 | |
| AB | 0.014 | 1 | 0.014 | 0.009 | 0.9266 | |
| AC | 1.910 | 1 | 1.910 | 1.220 | 0.2802 | |
| AD | 0.414 | 1 | 0.414 | 0.264 | 0.6120 | |
| AE | 1.850 | 1 | 1.850 | 1.180 | 0.2875 | |
| BC | 7.350 | 1 | 7.350 | 4.680 | 0.0399 | |
| BD | 1.440 | 1 | 1.440 | 0.920 | 0.3464 | |
| BE | 1.100 | 1 | 1.100 | 0.702 | 0.4098 | |
| CD | 0.684 | 1 | 0.684 | 0.436 | 0.5150 | |
| CE | 0.300 | 1 | 0.300 | 0.191 | 0.6656 | |
| DE | 0.0002 | 1 | 0.0002 | 0.000 | 0.9911 | |
| A2 | 1086.690 | 1 | 1086.690 | 691.660 | <0.0001 | |
| B2 | 45.170 | 1 | 45.170 | 28.750 | <0.0001 | |
| C2 | 746.940 | 1 | 746.940 | 475.410 | <0.0001 | |
| D2 | 126.960 | 1 | 126.960 | 80.810 | <0.0001 | |
| E2 | 125.030 | 1 | 125.030 | 79.580 | <0.0001 | |
| Residual | 40.850 | 26 | 1.570 | |||
| Lack of fit | 38.950 | 22 | 1.770 | 3.730 | 0.105 | Not significant |
| Pure error | 1.900 | 4 | 0.475 | |||
| Cor Total | 1660.400 | 46 | ||||
| R2 | 0.975 | |||||
| Adj. R2 | 0.957 | |||||
| CV (%) | 5.140 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klanrit, P.; Thanonkeo, S.; Pilap, W.; Apiraksakorn, J.; Fiala, K.; Leesing, R.; Yamada, M.; Thanonkeo, P. Optimization of Fermentation Parameters for Enhanced Bioethanol Production by Multistress-Tolerant Saccharomycodes ludwigii APRE2 Using Undetoxified Sugarcane Bagasse Hydrolysate. Energies 2025, 18, 3428. https://doi.org/10.3390/en18133428
Klanrit P, Thanonkeo S, Pilap W, Apiraksakorn J, Fiala K, Leesing R, Yamada M, Thanonkeo P. Optimization of Fermentation Parameters for Enhanced Bioethanol Production by Multistress-Tolerant Saccharomycodes ludwigii APRE2 Using Undetoxified Sugarcane Bagasse Hydrolysate. Energies. 2025; 18(13):3428. https://doi.org/10.3390/en18133428
Chicago/Turabian StyleKlanrit, Preekamol, Sudarat Thanonkeo, Warayutt Pilap, Jirawan Apiraksakorn, Khanittha Fiala, Ratanaporn Leesing, Mamoru Yamada, and Pornthap Thanonkeo. 2025. "Optimization of Fermentation Parameters for Enhanced Bioethanol Production by Multistress-Tolerant Saccharomycodes ludwigii APRE2 Using Undetoxified Sugarcane Bagasse Hydrolysate" Energies 18, no. 13: 3428. https://doi.org/10.3390/en18133428
APA StyleKlanrit, P., Thanonkeo, S., Pilap, W., Apiraksakorn, J., Fiala, K., Leesing, R., Yamada, M., & Thanonkeo, P. (2025). Optimization of Fermentation Parameters for Enhanced Bioethanol Production by Multistress-Tolerant Saccharomycodes ludwigii APRE2 Using Undetoxified Sugarcane Bagasse Hydrolysate. Energies, 18(13), 3428. https://doi.org/10.3390/en18133428