
Development of a Two-Stage Bioprocess for the Production of Bioethanol from the Acid Hydrolysate of Brewer’s Spent Grain

 , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Analytical Methods
2.2.2. Yeasts Isolation from Biowaste and Rotten Wood
2.2.3. Single-Substrate Fermentation in a Batch Reactor
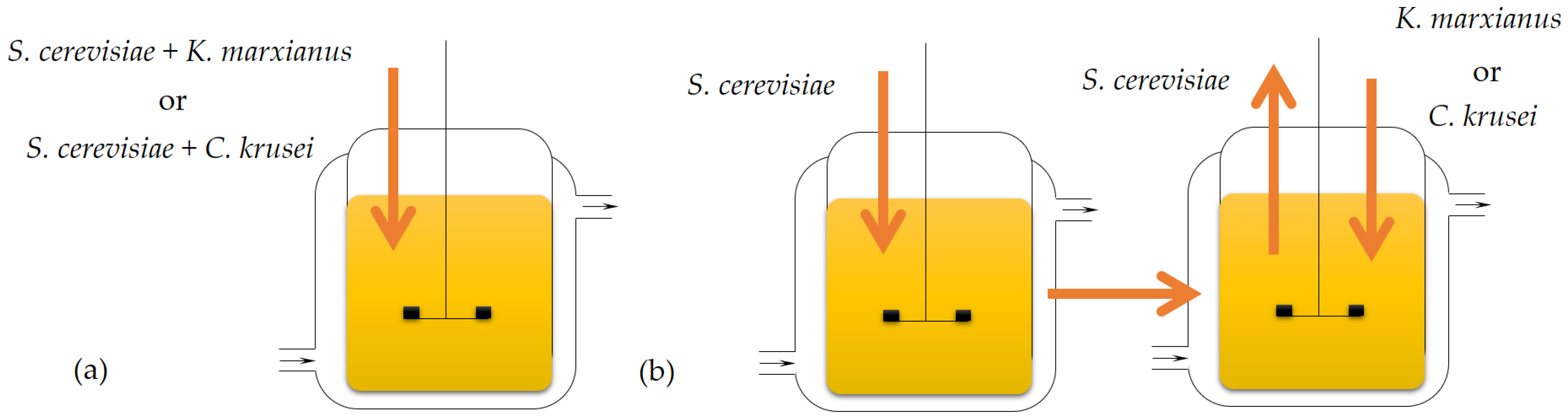
2.2.4. Two-Substrate Fermentation with K. marxianus and C. krusei Yeasts in a Batch Reactor
2.2.5. Two-Stage Fermentation of the Acidic Hydrolysate of BSG
2.2.6. Mathematical Modelling and Calculation of Bioprocess Efficiency Parameters
2.2.7. Data Processing
3. Results and Discussion
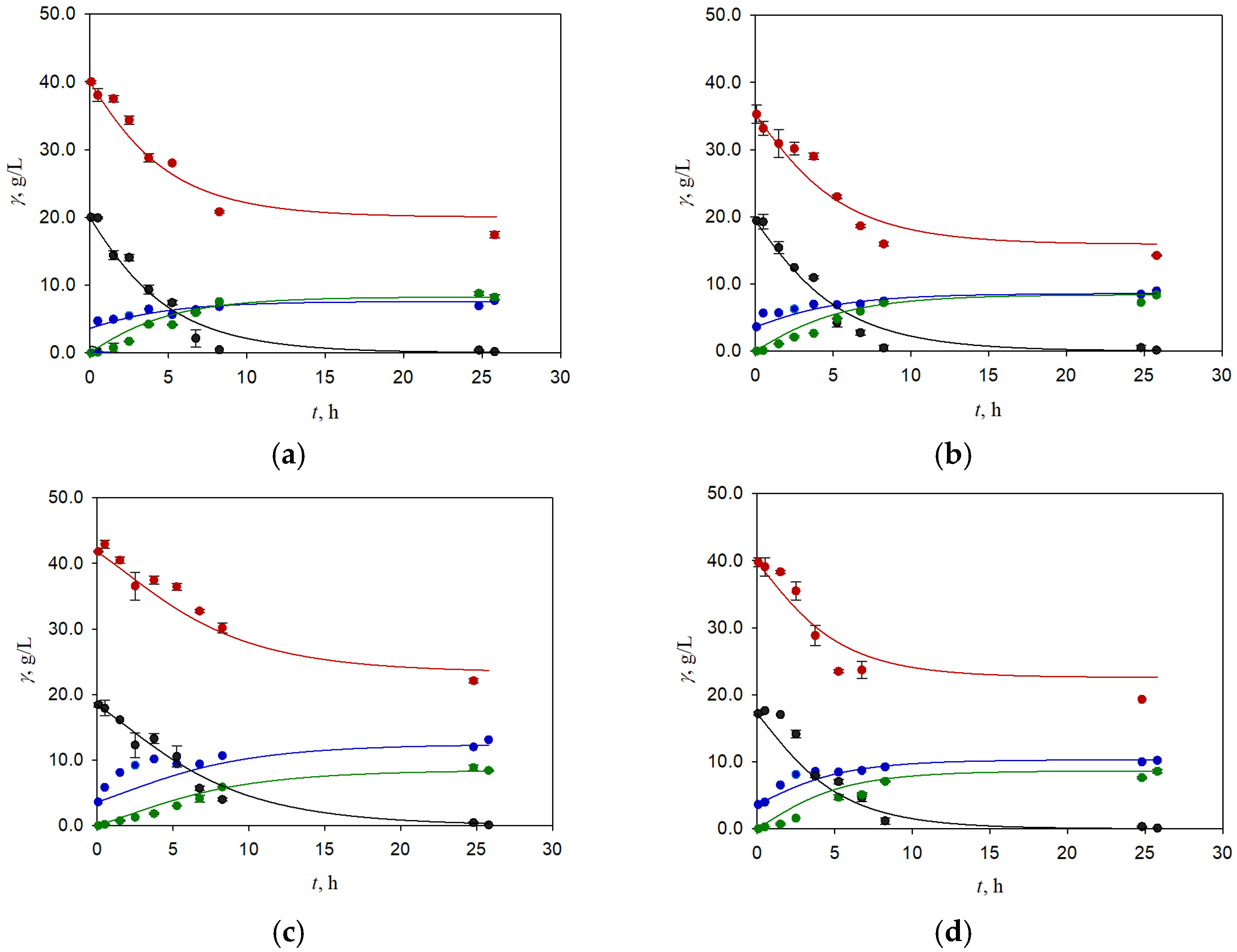
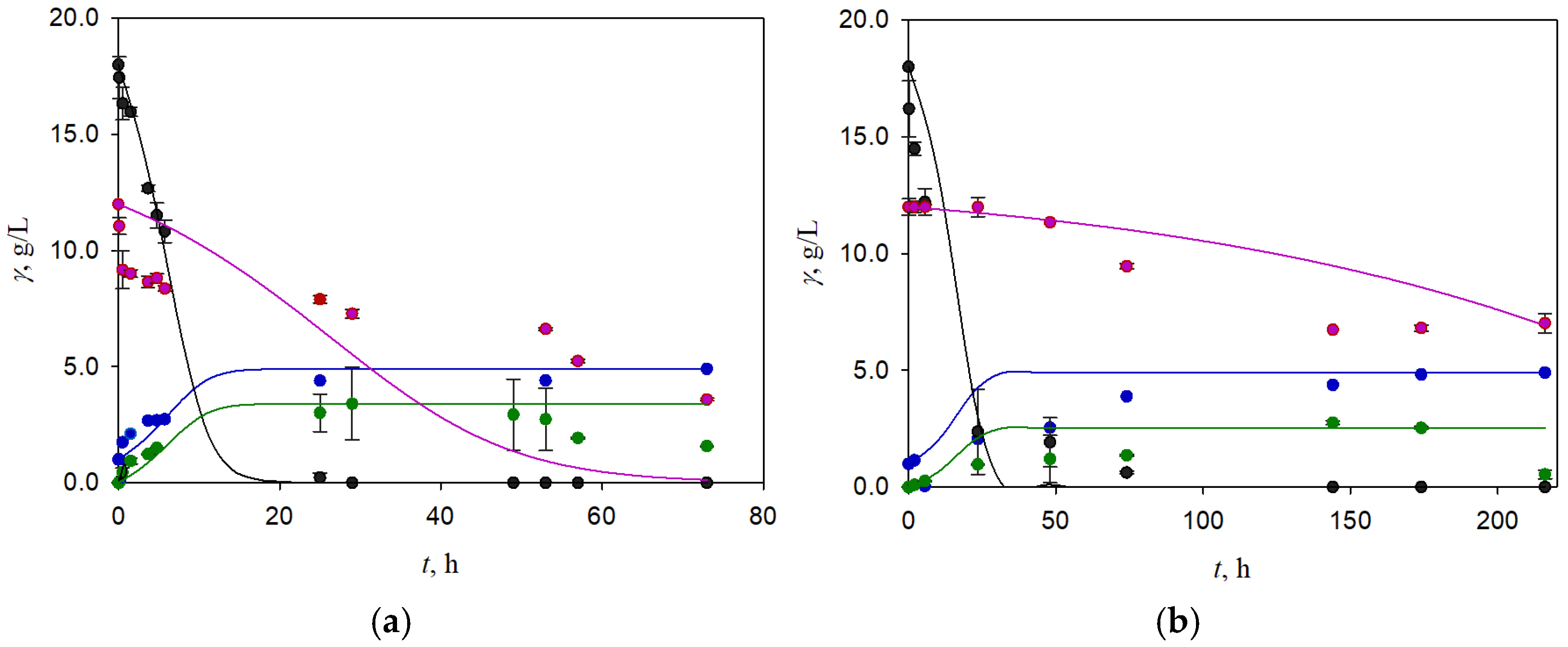
3.1. Batch Fermentation of Brewer’s Spent Grain Hydrolysate Obtained by Weak Acid Pretreatment
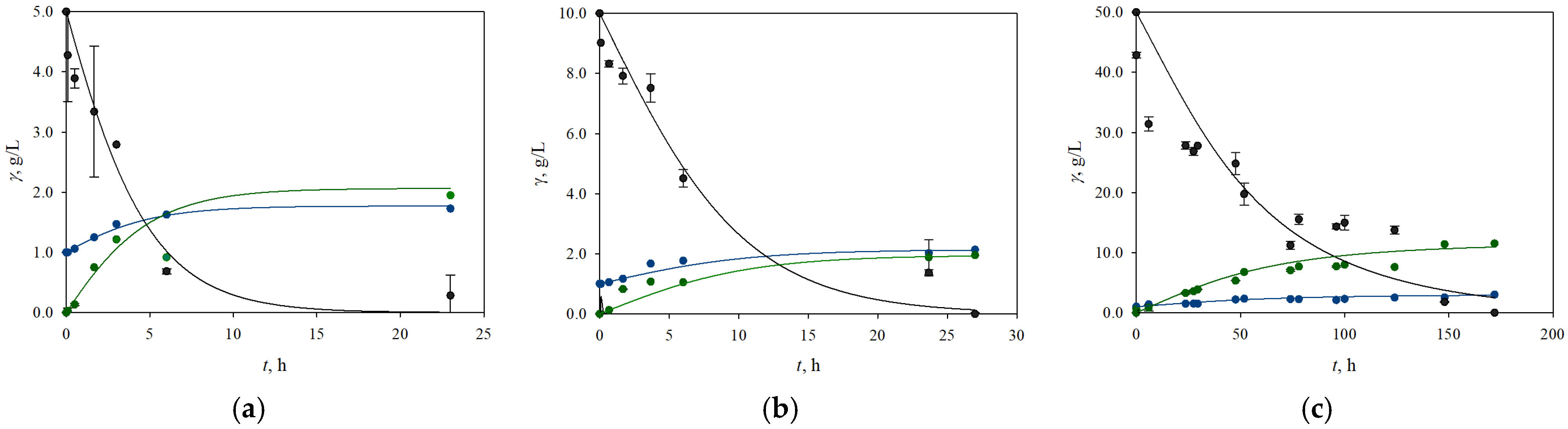
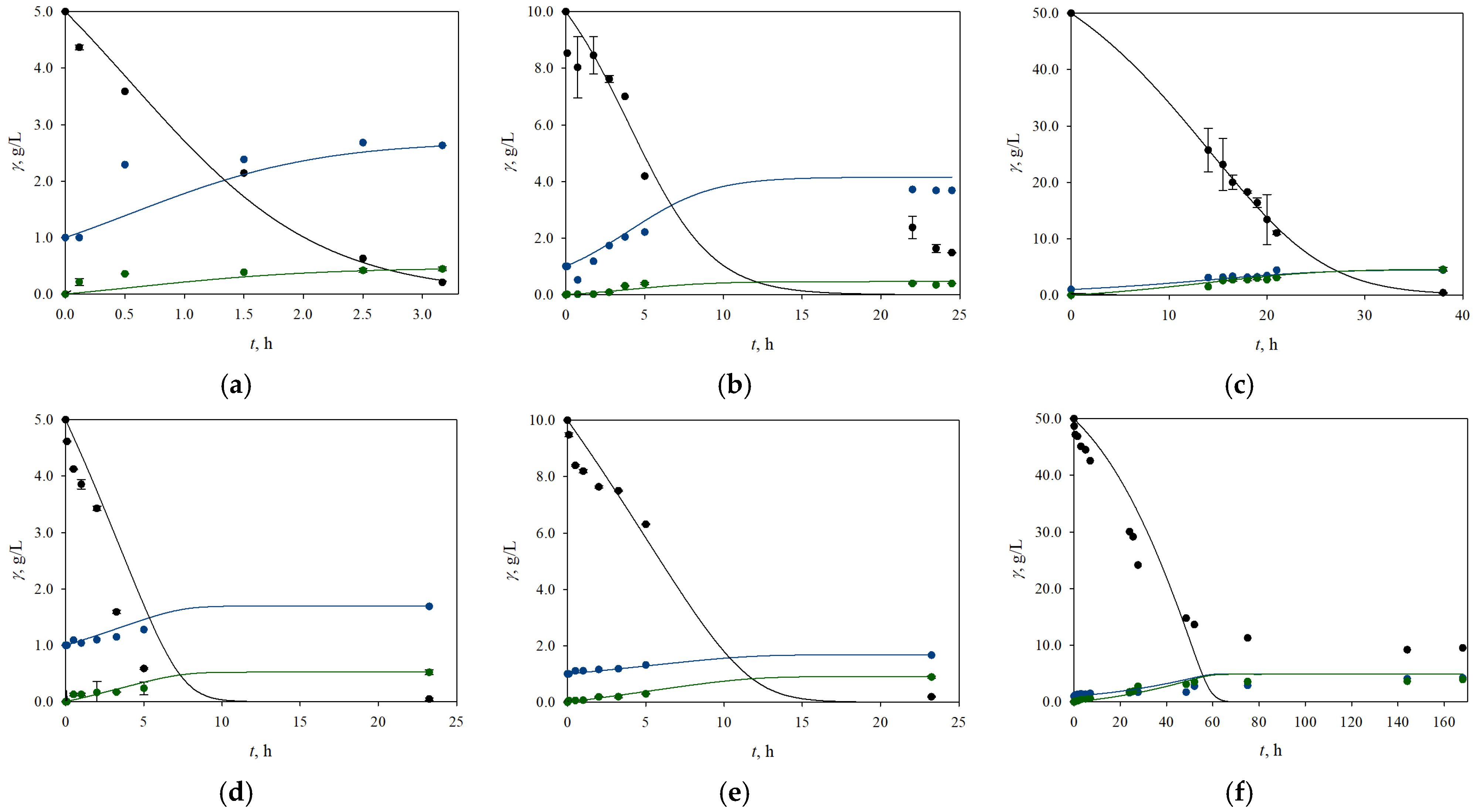
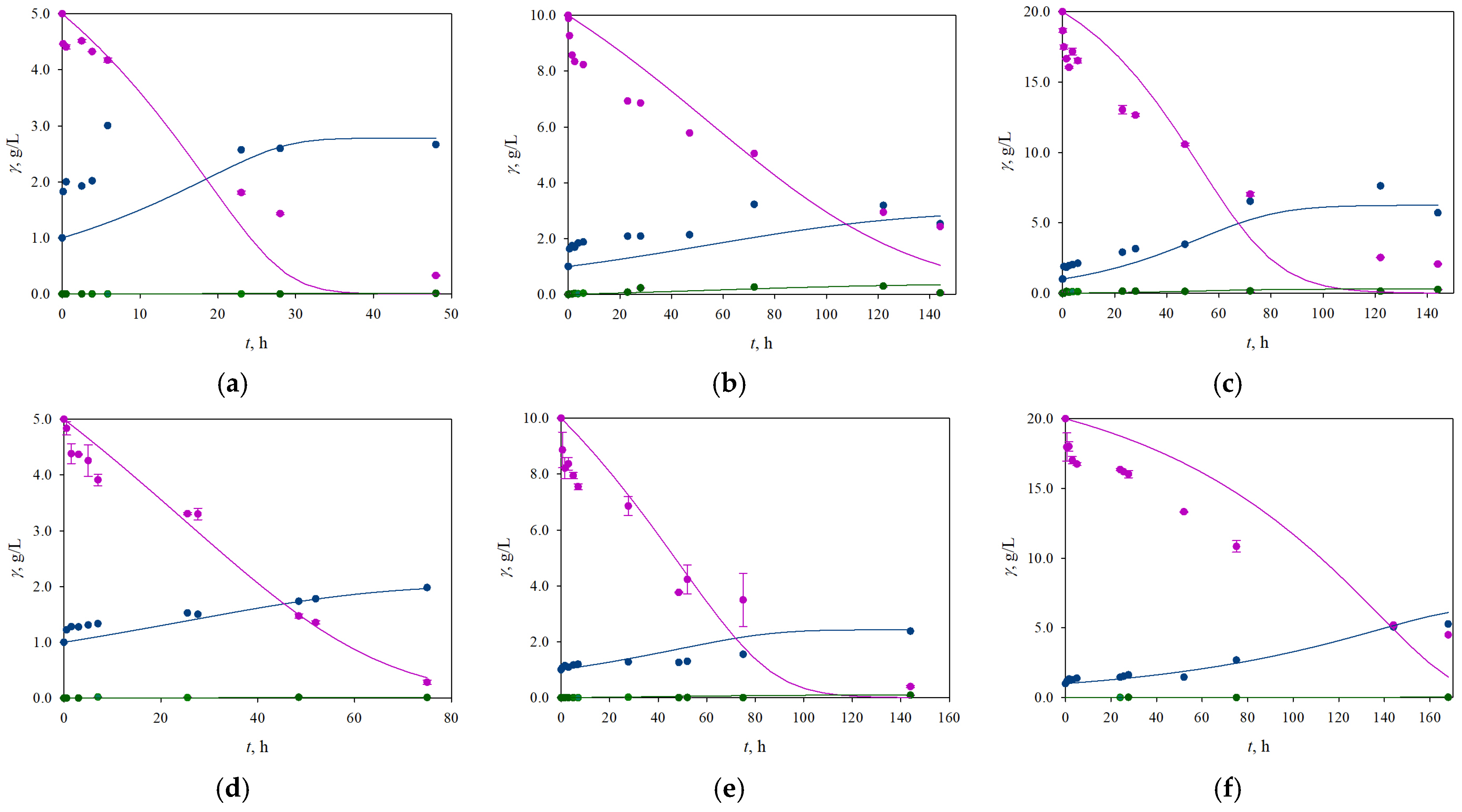
3.2. The Influence of Different Initial Concentrations of Glucose and Xylose on the Process Productivity and Growth of the Yeasts K. marxianus and C. krusei
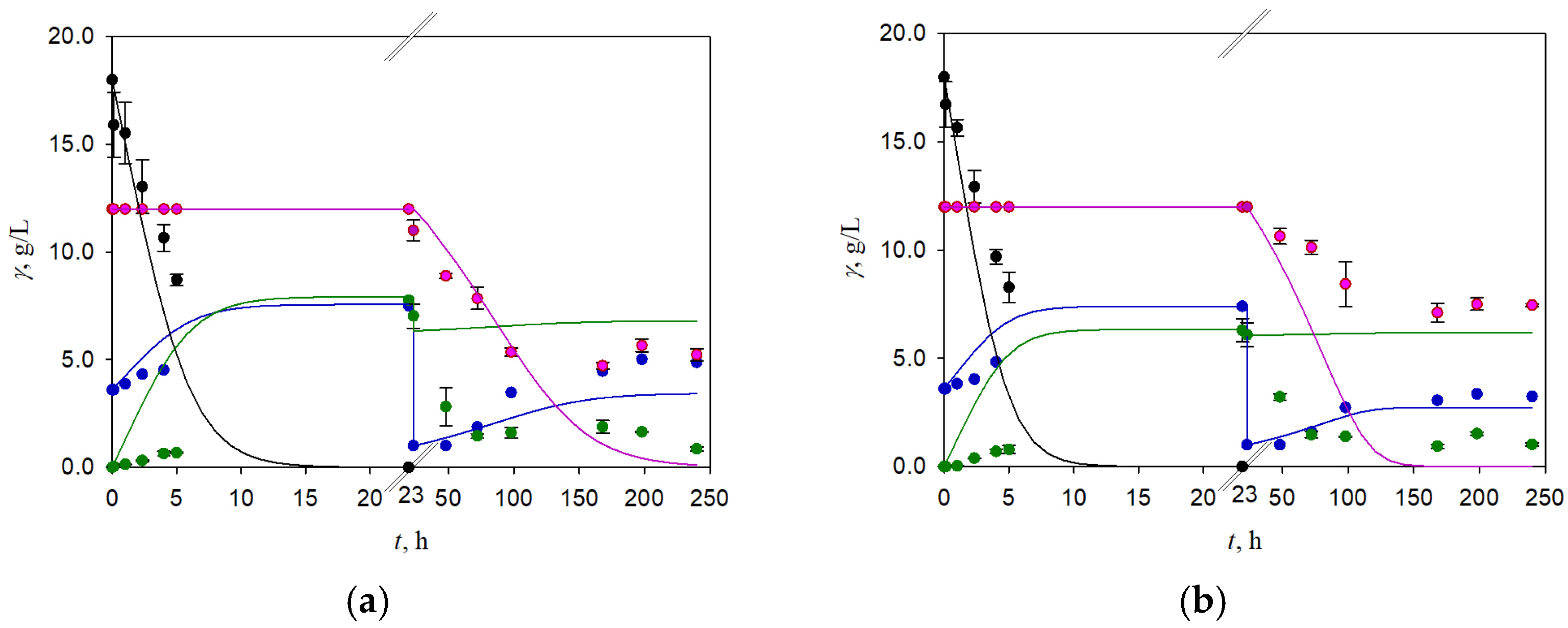
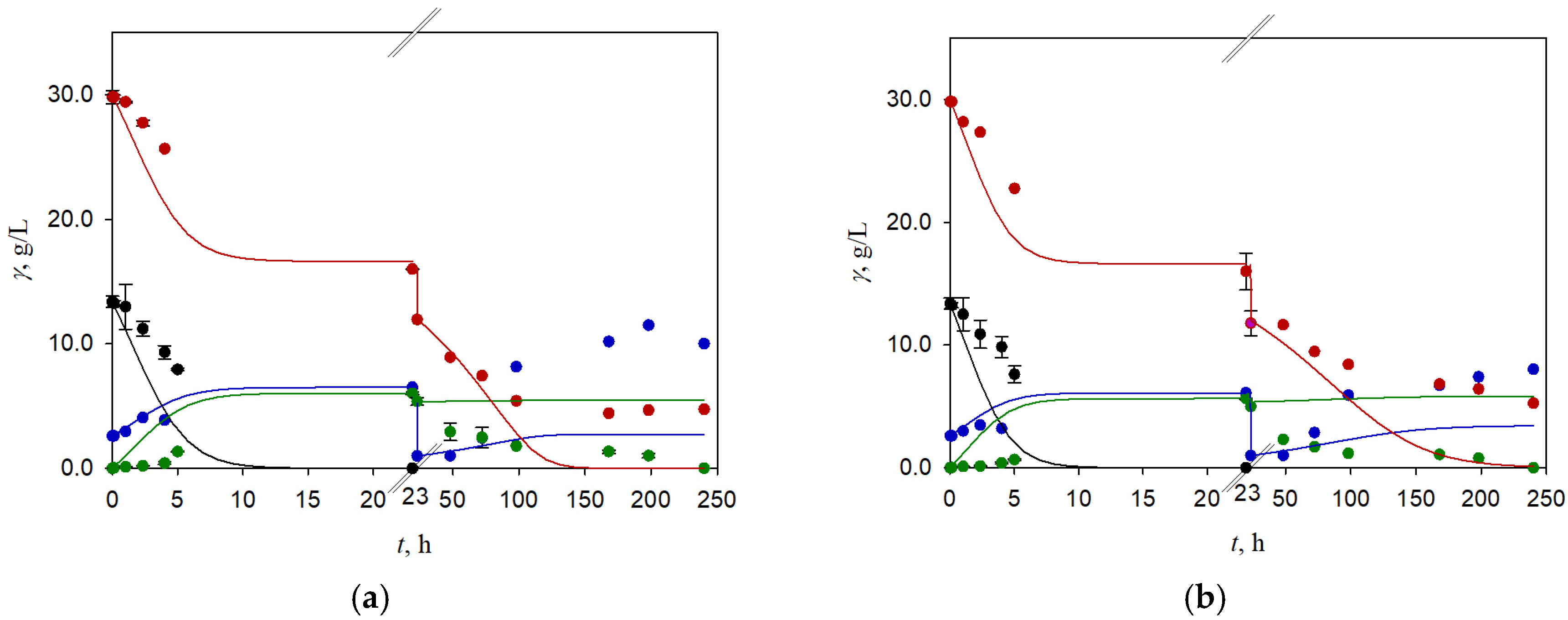
3.3. Two-Stage Fermentation of BSG
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zheng, J.; Negi, A.; Khomlaem, C.; Kim, B.S. Comparison of Bioethanol Production by Candida Molischiana and Saccharomyces Cerevisiae from Glucose, Cellobiose, and Cellulose. J. Microbiol. Biotechnol. 2019, 29, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Sharker, B.; Islam, M.A.; Hossain, M.A.A.; Ahmad, I.; Al Mamun, A.; Ghosh, S.; Rahman, A.; Hossain, M.S.; Ashik, M.A.; Hoque, M.R.; et al. Characterization of Lignin and Hemicellulose Degrading Bacteria Isolated from Cow Rumen and Forest Soil: Unveiling a Novel Enzymatic Model for Rice Straw Deconstruction. Sci. Total Environ. 2023, 904, 166704. [Google Scholar] [CrossRef] [PubMed]
- Merritt, H.; Barragán-Ocaña, A. The Impact of Market Factors on the Development of Eco-Friendly Energy Technologies: The Case of Bioethanol. Clean Technol. Environ. Policy 2021, 52, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Kazemi Shariat Panahi, H.; Dehhaghi, M.; Kinder, J.E.; Ezeji, T.C. A Review on Green Liquid Fuels for the Transportation Sector: A Prospect of Microbial Solutions to Climate Change. Biofuel Res. J. 2019, 6, 995–1024. [Google Scholar] [CrossRef]
- Novak, M.; Marđetko, N.; Trontel, A.; Pavlečić, M.; Kelemen, Z.; Perković, L.; Petravić Tominac, V.; Šantek, B. Development of an Integrated Bioprocess System for Bioethanol and Arabitol Production from Sugar Beet Cossettes. Food Technol. Biotechnol. 2024, 62, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Phitsuwan, P.; Sakka, K.; Ratanakhanokchai, K. Structural Changes and Enzymatic Response of Napier Grass (Pennisetum Purpureum) Stem Induced by Alkaline Pretreatment. Bioresour. Technol. 2016, 218, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Tursi, A. A Review on Biomass: Importance, Chemistry, Classification, and Conversion. Biofuel Res. J. 2019, 6, 962–979. [Google Scholar] [CrossRef]
- Rosado, M.J.; Marques, G.; Rencoret, J.; Gutiérrez, A.; del Río, J.C. Chemical Composition of Lipophilic Compounds from Rice (Oryza Sativa) Straw: An Attractive Feedstock for Obtaining Valuable Phytochemicals. Front. Plant. Sci. 2022, 13, 868319. [Google Scholar] [CrossRef] [PubMed]
- Didak Ljubas, B.; Novak, M.; Trontel, A.; Rajković, A.; Kelemen, Z.; Marđetko, N.; Grubišić, M.; Pavlečić, M.; Tominac, V.P.; Šantek, B. Production of Different Biochemicals by Paenibacillus Polymyxa DSM 742 From Pretreated Brewers’ Spent Grains. Front. Microbiol. 2022, 13, 812457. [Google Scholar] [CrossRef]
- Ravindran, R.; Jaiswal, S.; Abu-Ghannam, N.; Jaiswal, A.K. A Comparative Analysis of Pretreatment Strategies on the Properties and Hydrolysis of Brewers’ Spent Grain. Bioresour. Technol. 2018, 248, 272–279. [Google Scholar] [CrossRef]
- Pinheiro, T.; Coelho, E.; Romaní, A.; Domingues, L. Intensifying Ethanol Production from Brewer’s Spent Grain Waste: Use of Whole Slurry at High Solid Loadings. New Biotechnol. 2019, 53, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lynch, K.M.; Steffen, E.J.; Arendt, E.K. Brewers’ Spent Grain: A Review with an Emphasis on Food and Health. J. Inst. Brew. 2016, 122, 553–568. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, Z.; Huang, Y.; Wang, H.; Xiao, Q.; Ye, H.; Huang, X. Extraction and Preparation of Cellulose Nanocrystal from Brewer’s Spent Grain and Application in Pickering Emulsions. Bioact. Carbohydr. Diet. Fibre 2024, 31, 100418. [Google Scholar] [CrossRef]
- Vargas-Tah, A.; Moss-Acosta, C.L.; Trujillo-Martinez, B.; Tiessen, A.; Lozoya-Gloria, E.; Orencio-Trejo, M.; Gosset, G.; Martinez, A. Non-Severe Thermochemical Hydrolysis of Stover from White Corn and Sequential Enzymatic Saccharification and Fermentation to Ethanol. Bioresour. Technol. 2015, 198, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Parra-Ramírez, D.; Martinez, A.; Cardona, C.A. Technical and Economic Potential Evaluation of the Strain Escherichia Coli MS04 in the Ethanol Production from Glucose and Xylose. Biochem. Eng. J. 2018, 140, 123–129. [Google Scholar] [CrossRef]
- Wagner, E.; Sierra-Ibarra, E.; Rojas, N.L.; Martinez, A. One-Pot Bioethanol Production from Brewery Spent Grain Using the Ethanologenic Escherichia Coli MS04. Renew. Energy 2022, 189, 717–725. [Google Scholar] [CrossRef]
- Kręgiel, D.; Pawlikowska, E.; Antolak, H. Non-Conventional Yeasts in Fermentation Processes: Potentialities and Limitations. In Old Yeasts—New Questions; InTech: London, UK, 2017. [Google Scholar]
- Bergmann, J.C.; Trichez, D.; de Morais Junior, W.G.; Ramos, T.G.S.; Pacheco, T.F.; Carneiro, C.V.G.C.; Honorato, V.M.; Serra, L.A.; Almeida, J.R.M. Biotechnological Application of Non-Conventional Yeasts for Xylose Valorization. In Non-Conventional Yeasts: From Basic Research to Application; Springer International Publishing: Cham, Switzerland, 2019; pp. 23–74. [Google Scholar]
- Marđetko, N.; Novak, M.; Trontel, A.; Grubišić, M.; Galić, M.; Šantek, B. Bioethanol Production from Dilute-Acid Pre-Treated Wheat Straw Liquor Hydrolysate by Genetically Engineered Saccharomyces Cerevisiae. Chem. Biochem. Eng. Q 2018, 32, 483–499. [Google Scholar] [CrossRef]
- Tišma, M.; Jurić, A.; Bucić-Kojić, A.; Panjičko, M.; Planinić, M. Biovalorization of Brewers’ Spent Grain for the Production of Laccase and Polyphenols. J. Inst. Brew. 2018, 124, 182–186. [Google Scholar] [CrossRef]
- Rascón-Careaga, A.; Corella-Madueño, M.A.G.; Pérez-Martínez, C.J.; García-Rojas, A.M.; Souflé-Vásquez, S.Z.; García-Moroyoqui, M.T.; Córdoba-Beltrán, L.J.; Cáñez-Carrasco, M.G.; García-Alegría, A.M. Validation and Estimation of Uncertainty for a Glucose Determination Method GOD-PAP Using a Multi-Calibrator as Reference. Mapan J. Metrol. Soc. India 2021, 36, 269–278. [Google Scholar] [CrossRef]
- Du, R.; Guo, W.; Shen, Y.; Dai, J.; Zhang, H.; Fu, M.; Wang, X. In Situ Assay of the Reducing Sugars in Hydrophilic Natural Deep Eutectic Solvents by a Modified DNS Method. J. Mol. Liq. 2023, 385, 122286. [Google Scholar] [CrossRef]
- Šalić, A.; Faletar, P.; Zelić, B. NAD+ Regeneration in a Microreactor Using Permeabilized Baker’s Yeast Cells. Biochem. Eng. J. 2013, 77, 88–96. [Google Scholar] [CrossRef]
- Budžaki, S. Enzyme-Catalysed Biodiesel Production from Edible and Waste Cooking Oils. Chem. Biochem. Eng. Q 2015, 29, 329–333. [Google Scholar] [CrossRef]
- Sonnleitner, B.; Locher, G.; Fiechter, A. Biomass Determination. J. Biotechnol. 1992, 25, 5–22. [Google Scholar] [CrossRef] [PubMed]
- Foutch, G.L.; Johannes, A.H. Reactors in Process Engineering. In Encyclopedia of Physical Science and Technology; Elsevier: Amsterdam, The Netherlands, 2003; pp. 23–43. [Google Scholar]
- Ghose, T.K.; Tyagi, R.D. Rapid Ethanol Fermentation of Cellulose Hydrolysate. II. Product and Substrate Inhibition and Optimization of Fermentor Design. Biotechnol. Bioeng. 1979, 21, 1401–1420. [Google Scholar] [CrossRef]
- Mussatto, S.I.; Dragone, G.; Roberto, I.C. Brewers’ Spent Grain: Generation, Characteristics and Potential Applications. J. Cereal Sci. 2006, 43, 1–14. [Google Scholar] [CrossRef]
- Karlović, A.; Jurić, A.; Ćorić, N.; Habschied, K.; Krstanović, V.; Mastanjević, K. By-Products in the Malting and Brewing Industries—Re-Usage Possibilities. Fermentation 2020, 6, 82. [Google Scholar] [CrossRef]
- Kucharska, K.; Rybarczyk, P.; Hołowacz, I.; Łukajtis, R.; Glinka, M.; Kamiński, M. Pretreatment of Lignocellulosic Materials as Substrates for Fermentation Processes. Molecules 2018, 23, 2937. [Google Scholar] [CrossRef] [PubMed]
- Jönsson, L.J.; Alriksson, B.; Nilvebrant, N.O. Bioconversion of Lignocellulose: Inhibitors and Detoxification. Biotechnol. Biofuels 2013, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fathin Ruslan, N. Sustainable Bioethanol Production by Solid State Fermentation: A Systematic Review. Res. Sq. 2023. [Google Scholar] [CrossRef]
- Pavlečić, M.; Novak, M.; Trontel, A.; Marđetko, N.; Grubišić, M.; Ljubas, B.D.; Tominac, V.P.; Rakovac, R.Č.; Šantek, B. Mathematical Modelling of Bioethanol Production from Raw Sugar Beet Cossettes in a Horizontal Rotating Tubular Bioreactor. Fermentation 2022, 8, 13. [Google Scholar] [CrossRef]
- Graeme, M. Yeast Physiology and Biotechnology; John Wiley & Sons: Hoboken, NJ, USA, 1998; Volume 1. [Google Scholar]
- Tesfaw, A.; Assefa, F. Current Trends in Bioethanol Production by Saccharomyces Cerevisiae: Substrate, Inhibitor Reduction, Growth Variables, Coculture, and Immobilization. Int. Sch. Res. Notices 2014, 2014, 532852. [Google Scholar] [CrossRef] [PubMed]
- Xiros, C.; Topakas, E.; Katapodis, P.; Christakopoulos, P. Hydrolysis and Fermentation of Brewer’s Spent Grain by Neurospora Crassa. Bioresour. Technol. 2008, 99, 5427–5435. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, S.; Smart, K.A.; Cook, D.J. Optimisation of Alkaline Reagent Based Chemical Pre-Treatment of Brewers Spent Grains for Bioethanol Production. Ind. Crops Prod. 2014, 62, 219–227. [Google Scholar] [CrossRef]
- White, J.S.; Yohannan, B.K.; Walker, G.M. Bioconversion of Brewer’s Spent Grains to Bioethanol. FEMS Yeast Res. 2008, 8, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Xiros, C.; Christakopoulos, P. Enhanced Ethanol Production from Brewer’s Spent Grain by a Fusarium Oxysporum Consolidated System. Biotechnol. Biofuels 2009, 2, 4. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, S.; Smart, K.A.; Cook, D.J. A Comparison of Dilute Acid- and Alkali-Catalyzed Hydrothermal Pretreatments for Bioethanol Production from Brewers’ Spent Grains. J. Am. Soc. Brew. Chem. 2014, 72, 143–153. [Google Scholar] [CrossRef]
- Xiros, C.; Topakas, E.; Katapodis, P.; Christakopoulos, P. Evaluation of Fusarium Oxysporum as an Enzyme Factory for the Hydrolysis of Brewer’s Spent Grain with Improved Biodegradability for Ethanol Production. Ind. Crops Prod. 2008, 28, 213–224. [Google Scholar] [CrossRef]
- Wilkinson, S.; Smart, K.A.; Cook, D.J. Optimising the (Microwave) Hydrothermal Pretreatment of Brewers Spent Grains for Bioethanol Production. J. Fuels 2015, 2015, 369283. [Google Scholar] [CrossRef]
- Heredia-Olea, E.; Pérez-Carrillo, E.; Serna-Saldívar, S.O. Effect of Extrusion Conditions and Hydrolysis with Fiber-Degrading Enzymes on the Production of C5 and C6 Sugars from Brewers’ Spent Grain for Bioethanol Production. Biofuel Res. J. 2015, 2, 203–208. [Google Scholar] [CrossRef]
- Kemppainen, K.; Rommi, K.; Holopainen, U.; Kruus, K. Steam Explosion of Brewer’s Spent Grain Improves Enzymatic Digestibility of Carbohydrates and Affects Solubility and Stability of Proteins. Appl. Biochem. Biotechnol. 2016, 180, 94–108. [Google Scholar] [CrossRef]
- Khaleghian, H.; Karimi, K.; Behzad, T. Ethanol Production from Rice Straw by Sodium Carbonate Pretreatment and Mucor Hiemalis Fermentation. Ind. Crops Prod. 2015, 76, 1079–1085. [Google Scholar] [CrossRef]
- Kwak, S.; Jin, Y.-S. Production of Fuels and Chemicals from Xylose by Engineered Saccharomyces Cerevisiae: A Review and Perspective. Microb. Cell Fact. 2017, 16, 82. [Google Scholar] [CrossRef] [PubMed]
- Febrianti, F. Bioethanol Production from Tofu Waste by Simultaneous Saccharification and Fermentation (SSF) Using Microbial Consortium. Int. J. Technol. 2017, 8, 898–908. [Google Scholar] [CrossRef]
- Byadgi, S.A.; Kalburgi, P.B. Production of Bioethanol from Waste Newspaper. Procedia Environ. Sci. 2016, 35, 555–562. [Google Scholar] [CrossRef]
- Karim, A.; Gerliani, N.; Aïder, M. Kluyveromyces Marxianus: An Emerging Yeast Cell Factory for Applications in Food and Biotechnology. Int. J. Food Microbiol. 2020, 333, 108818. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, G.G.; Heinzle, E.; Wittmann, C.; Gombert, A.K. The Yeast Kluyveromyces Marxianus and Its Biotechnological Potential. Appl. Microbiol. Biotechnol. 2008, 79, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Zwirzitz, A.; Alteio, L.; Sulzenbacher, D.; Atanasoff, M.; Selg, M. Ethanol Production from Wheat Straw Hydrolysate by Issatchenkia Orientalis Isolated from Waste Cooking Oil. J. Fungi 2021, 7, 121. [Google Scholar] [CrossRef] [PubMed]
- Yadav, J.S.S.; Bezawada, J.; Yan, S.; Tyagi, R.D.; Surampalli, R.Y. Candida Krusei: Biotechnological Potentials and Concerns about Its Safety. Can. J. Microbiol. 2012, 58, 937–952. [Google Scholar] [CrossRef]
- Chang, Y.-H.; Chang, K.-S.; Chen, C.-Y.; Hsu, C.-L.; Chang, T.-C.; Jang, H.-D. Enhancement of the Efficiency of Bioethanol Production by Saccharomyces Cerevisiae via Gradually Batch-Wise and Fed-Batch Increasing the Glucose Concentration. Fermentation 2018, 4, 45. [Google Scholar] [CrossRef]
- Dasgupta, D.; Junghare, V.; Nautiyal, A.K.; Jana, A.; Hazra, S.; Ghosh, D. Xylitol Production from Lignocellulosic Pentosans: A Rational Strain Engineering Approach toward a Multiproduct Biorefinery. J. Agric. Food Chem. 2019, 67, 1173–1186. [Google Scholar] [CrossRef]
- Mo, W.; Wang, M.; Zhan, R.; Yu, Y.; He, Y.; Lu, H. Kluyveromyces Marxianus Developing Ethanol Tolerance during Adaptive Evolution with Significant Improvements of Multiple Pathways. Biotechnol. Biofuels 2019, 12, 63. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| YP, g/L | YX/S, g/g | YP/S, g/g | E, % | Pr, g/(L·h) | |
|---|---|---|---|---|---|
| NaOH | 8.192 ± 0.251 | 0.199 ± 0.015 | 0.413 ± 0.013 | 80.788 ± 2.472 | 0.318 ± 0.010 |
| activated charcoal + NaOH | 8.335 ± 0.015 | 0.272 ± 0.010 | 0.433 ± 0.001 | 84.617 ± 0.152 | 0.323 ± 0.001 |
| Ca(OH)2 | 8.403 ± 0.114 | 0.478 ± 0.001 | 0.459 ± 0.006 | 89.662 ± 1.124 | 0.326 ± 0.004 |
| activated charcoal + Ca(OH)2 | 8.576 ± 0.247 | 0.389 ± 0.002 | 0.502 ± 0.014 | 98.148 ± 2.829 | 0.332 ± 0.010 |
| Yeast | γG, g/L | YP, g/L | YP/S, g/g | E, % | Pr, g/(L·h) | KS, g/L | µmax, 1/h |
|---|---|---|---|---|---|---|---|
| 5 | 0.446 ± 0.031 | 0.093 ± 0.006 | 18.194 ± 1.278 | 0.141 ± 0.009 | 12.568 | 2.412 | |
| K. marxianus | 10 | 0.395 ± 0.001 | 0.046 ± 0.001 | 9.0704 ± 0.032 | 0.016 ± 0.001 | 12.568 | 0.520 |
| 50 | 4.546 ± 0.324 | 0.092 ± 0.007 | 17.926 ± 1.279 | 0.120 ± 0.008 | 12.568 | 0.097 | |
| 5 | 0.520 ± 0.043 | 0.105 ± 0.009 | 20.559 ± 1.702 | 0.022 ± 0.002 | 2.095 | 0.186 | |
| C. krusei | 10 | 0.891 ± 0.039 | 0.091 ± 0.004 | 17.801 ± 0.774 | 0.038 ± 0.002 | 2.095 | 0.064 |
| 50 | 3.936 ± 0.028 | 0.097 ± 0.001 | 19.060 ± 0.133 | 0.023 ± 0.001 | 2.095 | 0.033 |
| Sugar, g/L | γGL, g/L | γBT, g/L | |||
|---|---|---|---|---|---|
| K. marxianus | C. krusei | K. marxianus | C. krusei | ||
| Glucose | 5 | 0.631 ± 0.001 | 0.244 ± 0.023 | 0.054 ± 0.002 | 0.002 ± 0.001 |
| 10 | 1.133 ± 0.321 | 0.258 ± 0.043 | 0.087 ± 0.011 | 0.003 ± 0.001 | |
| 50 | 5.309 ± 0.481 | 0.657 ± 0.119 | 0.131 ± 0.008 | 0.380 ± 0.007 | |
| Xylose | 5 | 0.025 ± 0.016 | 0.029 ± 0.002 | 0.059 ± 0.043 | 0.379 ± 0.005 |
| 10 | 0.037 ± 0.005 | 0.031 ± 0.001 | 0.412 ± 0.018 | 0.598 ± 0.005 | |
| 20 | 0.348 ± 0.012 | 0.041 ± 0.002 | 0.380 ± 0.007 | 1.860 ± 0.044 | |
| Process | Substrate | Yeast | YP, g/L | YP/S, g/g | E, % | Pr, g/(L·h) |
|---|---|---|---|---|---|---|
| One-stage fermentation | 18 g/L glucose and 12 g/L xylose | K. marxianus | 3.403 ± 1.561 | 0.150 ± 0.069 | 29.365 ± 13.470 | 0.117 ± 0.054 |
| C. krusei | 2.754 ± 0.084 | 0.118 ± 0.004 | 23.214 ± 0.708 | 0.019 ± 0.001 | ||
| Two-stage fermentation | S. cerevisiae and K. marxianus | 7.751 ± 0.001 | 0.431 ± 0.001 | 84.431 ± 0.001 | 0.337 ± 0.001 | |
| S. cerevisiae and C. krusei | 6.293 ± 0.538 | 0.350 ± 0.030 | 68.555 ± 5.861 | 0.274 ± 0.023 | ||
| Two-stage fermentation | BSG | S. cerevisiae and K. marxianus | 6.024 ± 0.326 | 0.451 ± 0.024 | 88.350 ± 4.776 | 0.262 ± 0.014 |
| S. cerevisiae and C. krusei | 5.632 ± 0.193 | 0.421 ± 0.014 | 82.589 ± 2.830 | 0.245 ± 0.008 |
| Process | Substrate | Yeast | γGL, g/L | γBT, g/L |
|---|---|---|---|---|
| One-stage fermentation | 18 g/L glucose and 12 g/L xylose | K. marxianus | 2.948 ± 1.516 | 0.547 ± 0.367 |
| C. krusei | 0.360 ± 0.047 | 1.330 ± 0.011 | ||
| Two-stage fermentation | K. marxianus | 2.446 ± 0.504 | 0.608 ± 0.062 | |
| C. krusei | 1.739 ± 0.074 | 0.909 ± 0.308 | ||
| Two-stage fermentation | BSG | K. marxianus | 1.674 ± 0.283 | 1.710 ± 0.043 |
| C. krusei | 0.999 ± 0.262 | 1.019 ± 0.022 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vičević, R.; Božinović, M.; Zekić, N.; Novak, M.; Grgić, D.K.; Šalić, A.; Zelić, B. Development of a Two-Stage Bioprocess for the Production of Bioethanol from the Acid Hydrolysate of Brewer’s Spent Grain. Energies 2024, 17, 3975. https://doi.org/10.3390/en17163975
Vičević R, Božinović M, Zekić N, Novak M, Grgić DK, Šalić A, Zelić B. Development of a Two-Stage Bioprocess for the Production of Bioethanol from the Acid Hydrolysate of Brewer’s Spent Grain. Energies. 2024; 17(16):3975. https://doi.org/10.3390/en17163975
Chicago/Turabian StyleVičević, Renata, Marko Božinović, Nikolina Zekić, Mario Novak, Dajana Kučić Grgić, Anita Šalić, and Bruno Zelić. 2024. "Development of a Two-Stage Bioprocess for the Production of Bioethanol from the Acid Hydrolysate of Brewer’s Spent Grain" Energies 17, no. 16: 3975. https://doi.org/10.3390/en17163975
APA StyleVičević, R., Božinović, M., Zekić, N., Novak, M., Grgić, D. K., Šalić, A., & Zelić, B. (2024). Development of a Two-Stage Bioprocess for the Production of Bioethanol from the Acid Hydrolysate of Brewer’s Spent Grain. Energies, 17(16), 3975. https://doi.org/10.3390/en17163975