
Bioenergy, Biofuels, Lipids and Pigments—Research Trends in the Use of Microalgae Grown in Photobioreactors



Abstract
:1. Introduction
2. Materials and Methods
3. Results
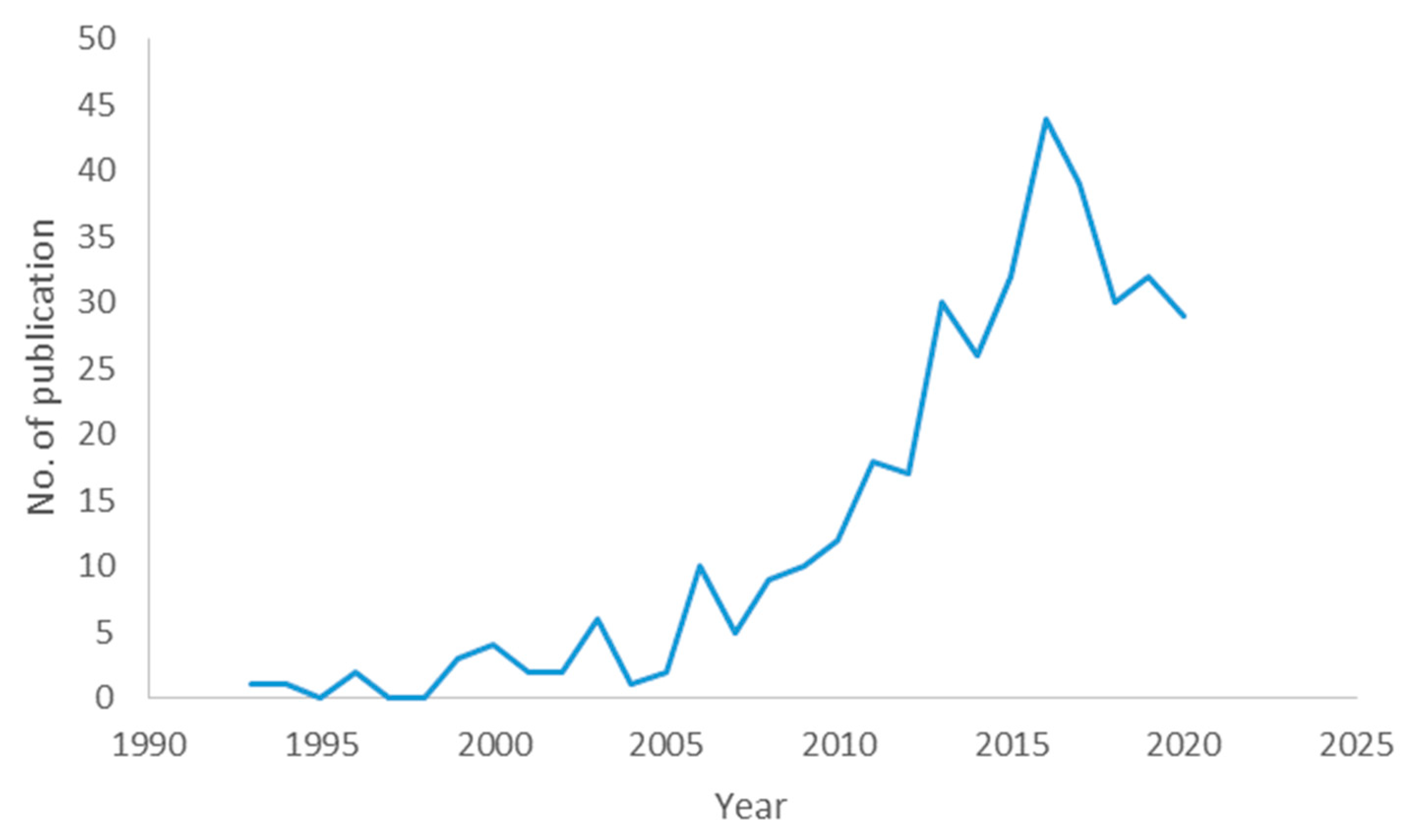
3.1. Descriptive Analysis
3.2. Scientometric Analysis
4. Content Analysis
- Techno-economic profitability of biofuel, bioenergy and pigment production in microalgae biorefineries.
- Influence of photobioreactor design and process parameters on the efficiency of microalgae cultivation.
- Strategies for increasing the amount of obtained lipids and obtaining biodiesel in the cultivation of Chlorella microalgae.
- Industrial production of astaxanthin using Haematococcus microalgae.
- Productivity of biomass and the use of alternative carbon sources in microalgae farming.
- Influence of light and carbon dioxide conversion on biomass efficiency.
- Heterotrophy.
4.1. Techno-Economic Profitability of the Production of Biofuels, Bioenergy and Pigments in a Microalgae Biorefinery
4.2. The Influence of Photobioreactor Design and Process Parameters on the Efficiency of Microalgae Cultivation
4.2.1. Photobioreactors
4.2.2. Parameters of Microalgae Cultivation
4.2.3. Culture Strategies
4.2.4. Light
4.3. Strategies for Increasing the Amount of Obtained Lipids and Obtaining Biodiesel in the Cultivation of Chlorella Microalgae
4.4. Industrial Production of Astaxanthin Using Haematococcus Microalgae
4.5. Increasing the Productivity of Biomass and the Use of Alternative Carbon Sources in Microalgae Farming
4.6. Influence of Light and Carbon Dioxide Conversion on Biomass Efficiency
4.7. Heterotrophy
5. Future Research
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Nomenclature
| EUR | European Union currency (Euro) |
| BBM | Bold’s Basal Medium |
| BG | Blue-Green Medium |
| CFD | Computational fluid dynamics |
| CO2 | Carbon dioxide |
| COD | Chemical oxygen demand |
| DHA | Docosahexaenoic acid |
| EPA | Eicosapentaenoic acid |
| EPS | Extracellular polysaccharides |
| EU | European Union |
| GRAS | Generally Recognized As Safe |
| IPC | Integrated permeate channel |
| kLa | Volumetric mass transfer coefficient |
| L | Liters |
| L/D | Light/dark cycle |
| LCA | Life cycle assessment |
| LED | Light-emitting diode |
| LNG | Liquefied natural gas |
| N/P | Nitrate-to-phosphate ratio |
| NPV | Net present value |
| NTG | N-methyl-N′-nitro-N-nitrosoguanidine |
| O2 | Oxygen |
| ORP | Open raceway pond |
| PBR | Photobioreactor |
| PE | Photosynthetic efficiency |
| pH | Hydrogen ion concentration |
| PMMA | Poly(methyl methacrylate) |
| PUFA | Polyunsaturated fatty acid |
| PV | Photovoltaic |
| PVC | Poly(vinyl chloride) |
| sp. | Species |
| TAG | Triacylglycerol |
| USD | United States currency (dollar) |
| UV | Ultraviolet |
| vvm | Volume per volume per minute |
References
- Pignolet, O.; Jubeau, S.; Vaca-Garcia, C.; Michaud, P. Highly valuable microalgae: Biochemical and topological aspects. J. Ind. Microbiol. Biotechnol. 2013, 40, 781–796. [Google Scholar] [CrossRef] [Green Version]
- Benedetti, M.; Vecchi, V.; Barera, S.; Dall’Osto, L. Biomass from microalgae: The potential of domestication towards sustainable biofactories. Microb. Cell Fact. 2018, 17, 173. [Google Scholar] [CrossRef] [Green Version]
- Ambati, R.R.; Gogisetty, D.; Aswathanarayana, R.G.; Ravi, S.; Bikkina, P.N.; Bo, L.; Yuepeng, S. Industrial potential of carotenoid pigments from microalgae: Current trends and future prospects. Crit. Rev. Food Sci. Nutr. 2018, 59, 1880–1902. [Google Scholar] [CrossRef]
- Han, S.-F.; Jin, W.-B.; Tu, R.-J.; Wu, W. Biofuel production from microalgae as feedstock: Current status and potential. Crit. Rev. Biotechnol. 2015, 35, 255–268. [Google Scholar] [CrossRef]
- Benavente-Valdés, J.R.; Aguilar, C.; Contreras-Esquivel, J.C.; Méndez-Zavala, A.; Montañez, J. Strategies to enhance the production of photosynthetic pigments and lipids in chlorophycae species. Biotechnol. Rep. 2016, 10, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Yuan, S.; Chen, R.; Ochieng, R.M. Sustainable production of energy from microalgae: Review of culturing systems, economics, and modelling. J. Renew. Sustain. Energy 2015, 7, 012701. [Google Scholar] [CrossRef]
- Liu, J.; Sun, Z.; Gerken, H.; Liu, Z.; Jiang, Y.; Chen, F. Chlorella zofingiensis as an Alternative Microalgal Producer of Astaxanthin: Biology and Industrial Potential. Mar. Drugs 2014, 12, 3487–3515. [Google Scholar] [CrossRef] [Green Version]
- Skjånes, K.; Rebours, C.; Lindblad, P. Potential for green microalgae to produce hydrogen, pharmaceuticals and other high value products in a combined process. Crit. Rev. Biotechnol. 2013, 33, 172–215. [Google Scholar] [CrossRef] [Green Version]
- Schulze, P.S.; Barreira, L.A.; Pereira, H.G.; Perales, J.A.; Varela, J.C. Light emitting diodes (LEDs) applied to microalgal production. Trends Biotechnol. 2014, 32, 422–430. [Google Scholar] [CrossRef]
- Wichuk, K.; Brynjolfsson, S.; Fu, W. Biotechnological production of value-added carotenoids from microalgae. Bioengineered 2014, 5, 204–208. [Google Scholar] [CrossRef] [Green Version]
- Bona, D.; Papurello, D.; Flaim, G.; Cerasino, L.; Biasioli, F.; Silvestri, S. Management of Digestate and Exhausts from Solid Oxide Fuel Cells Produced in the Dry Anaerobic Digestion Pilot Plant: Microalgae Cultivation Approach. Waste Biomass Valorization 2020, 11, 6499–6514. [Google Scholar] [CrossRef]
- Santarelli, M.; Briesemeister, L.; Gandiglio, M.; Herrmann, S.; Kuczynski, P.; Kupecki, J.; Lanzini, A.; Llovell, F.; Papurello, D.; Spliethoff, H.; et al. Carbon recovery and re-utilization (CRR) from the exhaust of a solid oxide fuel cell (SOFC): Analysis through a proof-of-concept. J. CO2 Util. 2017, 18, 206–221. [Google Scholar] [CrossRef]
- Llamas, B.; Suárez-Rodríguez, M.C.; González-López, C.V.; Mora, P.; Acién, F.G. Techno-economic analysis of microalgae related processes for CO2 bio-fixation. Algal Res. 2021, 57, 102339. [Google Scholar] [CrossRef]
- Kiran, B.; Kumar, R.; Deshmukh, D. Perspectives of microalgal biofuels as a renewable source of energy. Energy Convers. Manag. 2014, 88, 1228–1244. [Google Scholar] [CrossRef]
- Medipally, S.R.; Yusoff, F.; Banerjee, S.; Shariff, M. Microalgae as Sustainable Renewable Energy Feedstock for Biofuel Production. BioMed Res. Int. 2015, 2015, 519513. [Google Scholar] [CrossRef]
- Kothari, R.; Pandey, A.K.; Ahmad, S.; Kumar, A.; Pathak, V.V.; Tyagi, V.V. Microalgal cultivation for value-added products: A critical enviro-economical assessment. 3 Biotech 2017, 7, 243. [Google Scholar] [CrossRef]
- Tao, J.; Ge, Y.; Liang, R.; Sun, Y.; Cheng, Z.; Yan, B.; Chen, G. Technologies integration towards bio-fuels production: A state-of-the-art review. Appl. Energy Combust. Sci. 2022, 10, 100070. [Google Scholar] [CrossRef]
- Saei, A.A.; Ghanbari, P.; Barzegari, A. Haematococcus as a promising cell factory to produce recombinant pharmaceutical proteins. Mol. Biol. Rep. 2012, 39, 9931–9939. [Google Scholar] [CrossRef]
- Solovchenko, A.; Chekanov, K. Production of carotenoids using microalgae cultivated in photobioreactors. In Production of Biomass and Bioactive Compounds Using Bioreactor Technology; Springer: Dordrecht, The Netherlands, 2014; pp. 63–91. ISBN 9789401792233. [Google Scholar]
- Goli, A.; Shamiri, A.; Talaiekhozani, A.; Eshtiaghi, N.; Aghamohammadi, N.; Aroua, M.K. An overview of biological processes and their potential for CO2 capture. J. Environ. Manag. 2016, 183, 41–58. [Google Scholar] [CrossRef]
- Acién Fernández, F.G.; Fernández Sevilla, J.M.; Molina Grima, E. Photobioreactors for the production of microalgae. Rev. Environ. Sci. Biotechnol. 2013, 12, 131–151. [Google Scholar] [CrossRef]
- Chang, J.-S.; Show, P.-L.; Ling, T.-C.; Chen, C.-Y.; Ho, S.-H.; Tan, C.-H.; Nagarajan, D.; Phong, W.-N. Photobioreactors. In Current Developments in Biotechnology and Bioengineering: Bioprocesses, Bioreactors and Controls; Elsevier: Amsterdam, The Netherlands, 2017; pp. 313–352. [Google Scholar] [CrossRef]
- Shin, Y.S.; Choi, H.I.; Choi, J.W.; Lee, J.S.; Sung, Y.J.; Sim, S.J. Multilateral approach on enhancing economic viability of lipid production from microalgae: A review. Bioresour. Technol. 2018, 258, 335–344. [Google Scholar] [CrossRef]
- Koller, M. Design of Closed Photobioreactors for Algal Cultivation. In Algal Biorefineries: Volume 2: Products and Refinery Design; Springer International Publishing: Berlin/Heidelberg, Germany, 2015; pp. 133–186. [Google Scholar]
- Khan, A.; Hassan, M.K.; Paltrinieri, A.; Dreassi, A.; Bahoo, S. A bibliometric review of takaful literature. Int. Rev. Econ. Financ. 2020, 69, 389–405. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. Int. J. Surg. 2010, 8, 336–341. [Google Scholar] [CrossRef] [Green Version]
- Terán-Yépez, E.; Marín-Carrillo, G.M.; Casado-Belmonte, M.D.P.; Capobianco-Uriarte, M.D.L.M. Sustainable entrepreneurship: Review of its evolution and new trends. J. Clean. Prod. 2020, 252, 119742. [Google Scholar] [CrossRef]
- Wilson, A. Embracing Bayes Factors for Key Item Analysis in Corpus Linguistics. In New Approaches to the Study of Linguistic Variability, Language Competence and Language Awareness in Europe; Peter Lang: Frankfurt, Germany, 2013; pp. 3–11. [Google Scholar]
- Sarkar, S.; Maiti, J. Machine learning in occupational accident analysis: A review using science mapping approach with citation network analysis. Saf. Sci. 2020, 131, 104900. [Google Scholar] [CrossRef]
- van Eck, N.J.; Waltman, L. Software survey: VOSviewer, a computer program for bibliometric mapping. Scientometrics 2010, 84, 523–538. [Google Scholar] [CrossRef] [Green Version]
- van Eck, N.J.; Waltman, L. Text mining and visualization using VOSviewer. ISSI Newsl. 2011, 7, 50–54. [Google Scholar]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef]
- Brennan, L.; Owende, P. Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sustain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Yeh, K.-L.; Aisyah, R.; Lee, D.-J.; Chang, J.-S. Cultivation, photobioreactor design and harvesting of microalgae for biodiesel production: A critical review. Bioresour. Technol. 2011, 102, 71–81. [Google Scholar] [CrossRef]
- Ugwu, C.; Aoyagi, H.; Uchiyama, H. Photobioreactors for mass cultivation of algae. Bioresour. Technol. 2008, 99, 4021–4028. [Google Scholar] [CrossRef]
- Greenwell, H.C.; Laurens, L.M.L.; Shields, R.; Lovitt, R.; Flynn, K. Placing microalgae on the biofuels priority list: A review of the technological challenges. J. R. Soc. Interface 2010, 7, 703–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posten, C. Design principles of photo-bioreactors for cultivation of microalgae. Eng. Life Sci. 2009, 9, 165–177. [Google Scholar] [CrossRef]
- Borowitzka, M.A. Commercial production of microalgae: Ponds, tanks, tubes and fermenters. J. Biotechnol. 1999, 70, 313–321. [Google Scholar] [CrossRef]
- Del Campo, J.A.; García-González, M.; Guerrero, M.G. Outdoor cultivation of microalgae for carotenoid production: Current state and perspectives. Appl. Microbiol. Biotechnol. 2007, 74, 1163–1174. [Google Scholar] [CrossRef]
- Kumar, K.; Dasgupta, C.N.; Nayak, B.; Lindblad, P.; Das, D. Development of suitable photobioreactors for CO2 sequestration addressing global warming using green algae and cyanobacteria. Bioresour. Technol. 2011, 102, 4945–4953. [Google Scholar] [CrossRef]
- Suali, E.; Sarbatly, R. Conversion of microalgae to biofuel. Renew. Sustain. Energy Rev. 2012, 16, 4316–4342. [Google Scholar] [CrossRef]
- Carvalho, A.; Silva, S.; Baptista, J.; Malcata, F.X. Light requirements in microalgal photobioreactors: An overview of biophotonic aspects. Appl. Microbiol. Biotechnol. 2011, 89, 1275–1288. [Google Scholar] [CrossRef]
- Lee, Y.-K. Microalgal mass culture systems and methods: Their limitation and potential. J. Appl. Phycol. 2001, 13, 307–315. [Google Scholar] [CrossRef]
- Janssen, M.; Tramper, J.; Mur, L.R. Enclosed outdoor photobioreactors: Light regime, photosynthetic efficiency, scale-up, and future prospects. Biotechnol. Bioeng. 2002, 81, 193–210. [Google Scholar] [CrossRef]
- Apt, K.E.; Behrens, P.W. Commercial developments in microalgal biotechnology. J. Phycol. 1999, 35, 215–226. [Google Scholar] [CrossRef]
- Zhao, G.; Yu, J.; Jiang, F.; Zhang, X.; Tan, T. The effect of different trophic modes on lipid accumulation of Scenedesmus quadricauda. Bioresour. Technol. 2012, 114, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Jeffryes, C.; Rosenberger, J.; Rorrer, G.L. Fed-batch cultivation and bioprocess modeling of Cyclotella sp. for enhanced fatty acid production by controlled silicon limitation. Algal Res. 2013, 2, 16–27. [Google Scholar] [CrossRef]
- CYoo, C.; Choi, G.-G.; Kim, S.-C.; Oh, H.-M. Ettlia sp. YC001 showing high growth rate and lipid content under high CO2. Bioresour. Technol. 2013, 127, 482–488. [Google Scholar] [CrossRef]
- Song, L.; Qin, J.G.; Su, S.; Xu, J.; Clarke, S.; Shan, Y. Micronutrient Requirements for Growth and Hydrocarbon Production in the Oil Producing Green Alga Botryococcus braunii (Chlorophyta). PLoS ONE 2012, 7, e41459. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.W.; Moon, M.; Park, W.-K.; Yoo, G.; Choi, Y.-E.; Yang, J.-W. Energy-efficient cultivation of Chlamydomonas reinhardtii for lipid accumulation under flashing illumination conditions. Biotechnol. Bioprocess Eng. 2014, 19, 150–158. [Google Scholar] [CrossRef]
- YDasan, Y.K.; Lam, M.K.; Yusup, S.; Lim, J.W.; Show, P.L.; Tan, I.S.; Lee, K.T. Cultivation of Chlorella vulgaris using sequential-flow bubble column photobioreactor: A stress-inducing strategy for lipid accumulation and carbon dioxide fixation. J. CO2 Util. 2020, 41, 101226. [Google Scholar] [CrossRef]
- Hossain, N.; Zaini, J.; Mahlia, T.M.I. Life cycle assessment, energy balance and sensitivity analysis of bioethanol production from microalgae in a tropical country. Renew. Sustain. Energy Rev. 2019, 115, 109371. [Google Scholar] [CrossRef]
- Hossain, N.; Mahlia, T.M.I.; Zaini, J.; Saidur, R. Techno-economics and Sensitivity Analysis of Microalgae as Commercial Feedstock for Bioethanol Production. Environ. Prog. Sustain. Energy 2019, 38, 13157. [Google Scholar] [CrossRef] [Green Version]
- Lubián, L.M.; Montero, O.; Moreno-Garrido, I.; Huertas, E.; Sobrino, C.; Gonzalez-Delvalle, M.A.; Parés, G. Nannochloropsis (Eustigmatophyceae) as source of commercially valuable pigments. J. Appl. Phycol. 2000, 12, 249–255. [Google Scholar] [CrossRef]
- Katsuda, T.; Shimahara, K.; Shiraishi, H.; Yamagami, K.; Ranjbar, R.; Katoh, S. Effect of flashing light from blue light emitting diodes on cell growth and astaxanthin production of Haematococcus pluvialis. J. Biosci. Bioeng. 2006, 102, 442–446. [Google Scholar] [CrossRef] [PubMed]
- JMasojídek, J.; Torzillo, G.; Kopecký, J.; Koblížek, M.; Nidiaci, L.; Komenda, J.; Lukavská, A.; Sacchi, A. Changes in chlorophyll fluorescence quenching and pigment composition in the green alga Chlorococcum sp. grown under nitrogen deficiency and salinity stress. J. Appl. Phycol. 2000, 12, 417–426. [Google Scholar] [CrossRef]
- Wannachod, T.; Wannasutthiwat, S.; Powtongsook, S.; Nootong, K. Photoautotrophic cultivating options of freshwater green microalgal Chlorococcum humicola for biomass and carotenoid production. Prep. Biochem. Biotechnol. 2018, 48, 335–342. [Google Scholar] [CrossRef]
- Carbone, D.A.; Olivieri, G.; Pollio, A.; Melkonian, M. Biomass and phycobiliprotein production of Galdieria sulphuraria, immobilized on a twin-layer porous substrate photobioreactor. Appl. Microbiol. Biotechnol. 2020, 104, 3109–3119. [Google Scholar] [CrossRef] [PubMed]
- Pérez-López, P.; González-García, S.; Ulloa, R.G.; Sineiro, J.; Feijoo, G.; Moreira, M.T. Life cycle assessment of the production of bioactive compounds from Tetraselmis suecica at pilot scale. J. Clean. Prod. 2014, 64, 323–331. [Google Scholar] [CrossRef]
- Moroni, M.; Lorino, S.; Cicci, A.; Bravi, M. Design and Bench-Scale Hydrodynamic Testing of Thin-Layer Wavy Photobioreactors. Water 2019, 11, 1521. [Google Scholar] [CrossRef] [Green Version]
- Christiansen, K.L.; Raman, D.R.; Anex, R.P. Predicting cost growth and performance of first-generation algal production systems. Energy Policy 2012, 51, 382–391. [Google Scholar] [CrossRef]
- Razon, L.F.; Tan, R.R. Net energy analysis of the production of biodiesel and biogas from the microalgae: Haematococcus pluvialis and Nannochloropsis. Appl. Energy 2011, 88, 3507–3514. [Google Scholar] [CrossRef]
- Thomassen, G.; Vila, U.E.; Van Dael, M.; Lemmens, B.; Van Passel, S. A techno-economic assessment of an algal-based biorefinery. Clean Technol. Environ. Policy 2016, 18, 1849–1862. [Google Scholar] [CrossRef]
- Monari, C.; Righi, S.; Olsen, S.I. Greenhouse gas emissions and energy balance of biodiesel production from microalgae cultivated in photobioreactors in Denmark: A life-cycle modeling. J. Clean. Prod. 2016, 112, 4084–4092. [Google Scholar] [CrossRef] [Green Version]
- Mirón, A.S.; Garcıía, M.-C.C.; Camacho, F.G.; Grima, E.M.; Chisti, Y. Growth and biochemical characterization of microalgal biomass produced in bubble column and airlift photobioreactors: Studies in fed-batch culture. Enzym. Microb. Technol. 2002, 31, 1015–1023. [Google Scholar] [CrossRef]
- Deprá, M.C.; Mérida, L.G.; de Menezes, C.R.; Zepka, L.Q.; Jacob-Lopes, E. A new hybrid photobioreactor design for microalgae culture. Chem. Eng. Res. Des. 2019, 144, 1–10. [Google Scholar] [CrossRef]
- Salmean, C.; Bonilla, S.; Azimi, Y.; Aitchison, J.S.; Allen, D.G. Design and testing of an externally-coupled planar waveguide photobioreactor. Algal Res. 2019, 44, 101684. [Google Scholar] [CrossRef]
- Raha, H.E.; Shafii, M.B.; Roshandel, R. Energy efficient cultivation of microalgae using phosphorescence materials and mirrors. Sustain. Cities Soc. 2018, 41, 449–454. [Google Scholar] [CrossRef]
- Masojídek, J.; Sergejevová, M.; Rottnerová, K.; Jirka, V.; Korecko, J.; Kopecky, J.; Zaťková, I.; Torzillo, G.; Stys, D. A two-stage solar photobioreactor for cultivation of microalgae based on solar concentrators. J. Appl. Phycol. 2009, 21, 55–63. [Google Scholar] [CrossRef]
- Pham, H.-M.; Kwak, H.S.; Hong, M.-E.; Lee, J.; Chang, W.S.; Sim, S.J. Development of an X-Shape airlift photobioreactor for increasing algal biomass and biodiesel production. Bioresour. Technol. 2017, 239, 211–218. [Google Scholar] [CrossRef]
- Kwak, H.S.; Kim, J.Y.H.; Sim, S.J. A microscale approach for simple and rapid monitoring of cell growth and lipid accumulation in Neochloris oleoabundans. Bioprocess Biosyst. Eng. 2015, 38, 2035–2043. [Google Scholar] [CrossRef]
- Jeffryes, C.; Severi, V.; Delhaye, A.; Urbain, B.; Grama, B.S.; Agathos, S.N. Energy conversion in an internally illuminated annular-plate airlift photobioreactor. Eng. Life Sci. 2016, 16, 348–354. [Google Scholar] [CrossRef]
- Rebolledo-Oyarce, J.; Mejía-López, J.; García, G.; Rodríguez-Córdova, L.; Sáez-Navarrete, C. Novel photobioreactor design for the culture of Dunaliella tertiolecta—Impact of color in the growth of microalgae. Bioresour. Technol. 2019, 289, 121645. [Google Scholar] [CrossRef]
- López, M.G.-M.; Sanchez, E.D.R.; López, J.C.; Fernandez, F.G.A.; Sevilla, J.M.F.; Rivas, J.; Guerrero, M.; Grima, E.M. Comparative analysis of the outdoor culture of Haematococcus pluvialis in tubular and bubble column photobioreactors. J. Biotechnol. 2006, 123, 329–342. [Google Scholar] [CrossRef]
- Kumar, K.; Das, D. Growth characteristics of Chlorella sorokiniana in airlift and bubble column photobioreactors. Bioresour. Technol. 2012, 116, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Imamoglu, E. Fotobiyoreaktörlerin geometrik konfigürasyonlarının sucul bentik diatom Cylindrotheca closterium üremesi üzerine etkisi. Turk. J. Biochem. 2016, 41, 105–111. [Google Scholar] [CrossRef]
- Powtongsook, S.; Nootong, K. Photoautotrophic cultivation of Chlorococcum humicola in stirred tank and airlift photobioreactors under different light settings and light supplying strategies for biomass and carotenoid production. J. Chem. Technol. Biotechnol. 2019, 94, 3084–3092. [Google Scholar] [CrossRef]
- Suh, I.S.; Joo, H.-N.; Lee, C.-G. A novel double-layered photobioreactor for simultaneous Haematococcus pluvialis cell growth and astaxanthin accumulation. J. Biotechnol. 2006, 125, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Cui, X.; Feng, Y.; Jing, G.; Kang, L.; Luo, M. Experimental study on microalgae cultivation in novel photobioreactor of concentric double tubes with aeration pores along tube length direction. Int. J. Green Energy 2017, 14, 1269–1276. [Google Scholar] [CrossRef]
- Issarapayup, K.; Powtongsook, S.; Pavasant, P. Flat panel airlift photobioreactors for cultivation of vegetative cells of microalga Haematococcus pluvialis. J. Biotechnol. 2009, 142, 227–232. [Google Scholar] [CrossRef]
- Hincapie, E.; Stuart, B.J. Design, Construction, and Validation of an Internally Lit Air-Lift Photobioreactor for Growing Algae. Front. Energy Res. 2015, 2, 65. [Google Scholar] [CrossRef] [Green Version]
- Borowitzka, M.A.; Vonshak, A. Scaling up microalgal cultures to commercial scale. Eur. J. Phycol. 2017, 52, 407–418. [Google Scholar] [CrossRef]
- Chandra, T.S.; Aditi, S.; Kumar, M.M.; Mukherji, S.; Modak, J.; Chauhan, V.S.; Sarada, R.; Mudliar, S.N. Growth and biochemical characteristics of an indigenous freshwater microalga, Scenedesmus obtusus, cultivated in an airlift photobioreactor: Effect of reactor hydrodynamics, light intensity, and photoperiod. Bioprocess Biosyst. Eng. 2017, 40, 1057–1068. [Google Scholar] [CrossRef]
- Mehariya, S.; Sharma, N.; Iovine, A.; Casella, P.; Marino, T.; LaRocca, V.; Molino, A.; Musmarra, D. An Integrated Strategy for Nutraceuticals from Haematoccus pluvialis: From Cultivation to Extraction. Antioxidants 2020, 9, 825. [Google Scholar] [CrossRef]
- Narala, R.R.; Garg, S.; Sharma, K.K.; Thomas-Hall, S.R.; Deme, M.; Li, Y.; Schenk, P.M. Comparison of Microalgae Cultivation in Photobioreactor, Open Raceway Pond, and a Two-Stage Hybrid System. Front. Energy Res. 2016, 4, 29. [Google Scholar] [CrossRef]
- Guedes, A.C.; Amaro, H.M.; Pereira, R.D.; Malcata, F.X. Effects of temperature and pH on growth and antioxidant content of the microalga Scenedesmus obliquus. Biotechnol. Prog. 2011, 27, 1218–1224. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Slavens, S.; Crunkleton, D.W.; Johannes, T.W. Interactive effect of light quality and temperature on Chlamydomonas reinhardtii growth kinetics and lipid synthesis. Algal Res. 2021, 53, 102127. [Google Scholar] [CrossRef]
- Gonçalves, V.D.; Fagundes-Klen, M.R.; Trigueros, D.E.G.; Kroumov, A.D.; Módenes, A.N. Statistical and optimization strategies to carotenoids production by Tetradesmus acuminatus (LC192133.1) cultivated in photobioreactors. Biochem. Eng. J. 2019, 152, 107351. [Google Scholar] [CrossRef]
- Coulombier, N.; Nicolau, E.; Le Déan, L.; Barthelemy, V.; Schreiber, N.; Brun, P.; Lebouvier, N.; Jauffrais, T. Effects of Nitrogen Availability on the Antioxidant Activity and Carotenoid Content of the Microalgae Nephroselmis sp. Mar. Drugs 2020, 18, 453. [Google Scholar] [CrossRef]
- Xie, Y.; Ho, S.-H.; Chen, C.-N.N.; Chen, C.-Y.; Ng, I.-S.; Jing, K.-J.; Chang, J.-S.; Lu, Y. Phototrophic cultivation of a thermo-tolerant Desmodesmus sp. for lutein production: Effects of nitrate concentration, light intensity and fed-batch operation. Bioresour. Technol. 2013, 144, 435–444. [Google Scholar] [CrossRef]
- Morales-Sánchez, D.; Schulze, P.S.; Kiron, V.; Wijffels, R.H. Production of carbohydrates, lipids and polyunsaturated fatty acids (PUFA) by the polar marine microalga Chlamydomonas malina RCC2488. Algal Res. 2020, 50, 102016. [Google Scholar] [CrossRef]
- Xia, S.; Wan, L.; Li, A.; Sang, M.; Zhang, C. Effects of nutrients and light intensity on the growth and biochemical composition of a marine microalga Odontella aurita. Chin. J. Oceanol. Limnol. 2013, 31, 1163–1173. [Google Scholar] [CrossRef]
- Liu, J.; Sommerfeld, M.; Hu, Q. Screening and characterization of Isochrysis strains and optimization of culture conditions for docosahexaenoic acid production. Appl. Microbiol. Biotechnol. 2013, 97, 4785–4798. [Google Scholar] [CrossRef]
- Pedro, A.S.; González-López, C.; Acién, F.; Molina-Grima, E. Outdoor pilot-scale production of Nannochloropsis gaditana: Influence of culture parameters and lipid production rates in tubular photobioreactors. Bioresour. Technol. 2014, 169, 667–676. [Google Scholar] [CrossRef]
- Van Wagenen, J.; De Francisci, D.; Angelidaki, I. Comparison of mixotrophic to cyclic autotrophic/heterotrophic growth strategies to optimize productivity of Chlorella sorokiniana. J. Appl. Phycol. 2015, 27, 1775–1782. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, L.; Nagarajan, N.G.; Karuppan, M. Enhanced biomass and lutein production by mixotrophic cultivation of Scenedesmus sp. using crude glycerol in an airlift photobioreactor. Biochem. Eng. J. 2020, 161, 107684. [Google Scholar] [CrossRef]
- Schüler, L.M.; Gangadhar, K.N.; Duarte, P.; Placines, C.; Molina-Márquez, A.M.; Léon-Bañares, R.; Sousa, V.S.; Varela, J.; Barreira, L. Improvement of carotenoid extraction from a recently isolated, robust microalga, Tetraselmis sp. CTP4 (chlorophyta). Bioprocess Biosyst. Eng. 2020, 43, 785–796. [Google Scholar] [CrossRef]
- Zhang, J.; Müller, B.S.F.; Tyre, K.N.; Hersh, H.L.; Bai, F.; Hu, Y.; Resende, M.F.R.J.; Rathinasabapathi, B.; Settles, A.M. Competitive Growth Assay of Mutagenized Chlamydomonas reinhardtii Compatible With the International Space Station Veggie Plant Growth Chamber. Front. Plant Sci. 2020, 11, 631. [Google Scholar] [CrossRef]
- Fernandes, B.D.; Dragone, G.M.; Teixeira, J.A.; Vicente, A.A. Light Regime Characterization in an Airlift Photobioreactor for Production of Microalgae with High Starch Content. Appl. Biochem. Biotechnol. 2010, 161, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Sforza, E.; Simionato, D.; Giacometti, G.M.; Bertucco, A.; Morosinotto, T. Adjusted Light and Dark Cycles Can Optimize Photosynthetic Efficiency in Algae Growing in Photobioreactors. PLoS ONE 2012, 7, e38975. [Google Scholar] [CrossRef]
- Goold, H.D.; Cuiné, S.; Légeret, B.; Liang, Y.; Brugière, S.; Auroy, P.; Javot, H.; Tardif, M.; Jones, B.; Beisson, F.; et al. Saturating light induces sustained accumulation of oil in plastidal lipid droplets in Chlamydomonas reinhardtii. Plant Physiol. 2016, 171, 2406–2417. [Google Scholar] [CrossRef] [Green Version]
- Fu, W.; Guðmundsson, Ó.; Paglia, G.; Herjólfsson, G.; Andrésson, Ó.S.; Palsson, B.Ø.; Brynjólfsson, S. Enhancement of carotenoid biosynthesis in the green microalga Dunaliella salina with light-emitting diodes and adaptive laboratory evolution. Appl. Microbiol. Biotechnol. 2013, 97, 2395–2403. [Google Scholar] [CrossRef] [Green Version]
- Kula, M.; Rys, M.; Możdżeń, K.; Skoczowski, A. Metabolic activity, the chemical composition of biomass and photosynthetic activity of Chlorella vulgarisunder different light spectra in photobioreactors. Eng. Life Sci. 2013, 14, 57–67. [Google Scholar] [CrossRef]
- Li, Y.; Li, L.; Liu, J.; Qin, R. Light absorption and growth response of Dunaliella under different light qualities. J. Appl. Phycol. 2020, 32, 1041–1052. [Google Scholar] [CrossRef]
- Kim, Z.-H.; Park, Y.-S.; Ryu, Y.-J.; Lee, C.-G. Enhancing biomass and fatty acid productivity of Tetraselmis sp. in bubble column photobioreactors by modifying light quality using light filters. Biotechnol. Bioprocess Eng. 2017, 22, 397–404. [Google Scholar] [CrossRef]
- Markou, G. Effect of Various Colors of Light-Emitting Diodes (LEDs) on the Biomass Composition of Arthrospira platensis Cultivated in Semi-continuous Mode. Appl. Biochem. Biotechnol. 2014, 172, 2758–2768. [Google Scholar] [CrossRef]
- Ooms, M.D.; Graham, P.J.; Nguyen, B.; Sargent, E.H.; Sinton, D. Light dilution via wavelength management for efficient high-density photobioreactors. Biotechnol. Bioeng. 2017, 114, 1160–1169. [Google Scholar] [CrossRef]
- Dineshkumar, R.; Subramanian, G.; Dash, S.K.; Sen, R. Development of an optimal light-feeding strategy coupled with semi-continuous reactor operation for simultaneous improvement of microalgal photosynthetic efficiency, lutein production and CO2 sequestration. Biochem. Eng. J. 2016, 113, 47–56. [Google Scholar] [CrossRef]
- Abu-Ghosh, S.; Fixler, D.; Dubinsky, Z.; Iluz, D. Flashing light in microalgae biotechnology. Bioresour. Technol. 2016, 203, 357–363. [Google Scholar] [CrossRef]
- Glemser, M.; Heining, M.; Schmidt, J.; Becker, A.; Garbe, D.; Buchholz, R.; Brück, T. Application of light-emitting diodes (LEDs) in cultivation of phototrophic microalgae: Current state and perspectives. Appl. Microbiol. Biotechnol. 2016, 100, 1077–1088. [Google Scholar] [CrossRef]
- Jain, A.; Voulis, N.; Jung, E.E.; Doud, D.F.R.; Miller, W.B.; Angenent, L.T.; Erickson, D. Optimal Intensity and Biomass Density for Biofuel Production in a Thin-Light-Path Photobioreactor. Environ. Sci. Technol. 2015, 49, 6327–6334. [Google Scholar] [CrossRef]
- Sun, Z.; Zhou, Z.-G.; Gerken, H.; Chen, F.; Liu, J. Screening and characterization of oleaginous Chlorella strains and exploration of photoautotrophic Chlorella protothecoides for oil production. Bioresour. Technol. 2015, 184, 53–62. [Google Scholar] [CrossRef]
- Liao, Q.; Sun, Y.; Huang, Y.; Xia, A.; Fu, Q.; Zhu, X. Simultaneous enhancement of Chlorella vulgaris growth and lipid accumulation through the synergy effect between light and nitrate in a planar waveguide flat-plate photobioreactor. Bioresour. Technol. 2017, 243, 528–538. [Google Scholar] [CrossRef]
- Wen, X.; Geng, Y.; Li, Y. Enhanced lipid production in Chlorella pyrenoidosa by continuous culture. Bioresour. Technol. 2014, 161, 297–303. [Google Scholar] [CrossRef]
- Kim, N.J.; Suh, I.S.; Hur, B.K.; Lee, C.G. Simple monodimensional model for linear growth rate of photosynthetic microorganisms in flat-plate photobioreactor. J. Microbiol. Biotechnol. 2002, 12, 962–971. [Google Scholar]
- Guil-Guerrero, J.; Rebolloso-Fuentes, M. Nutrient Composition of Chlorella spp. and Monodus subterraneus Cultured in a Bubble Column Bioreactor. Food Biotechnol. 2008, 22, 218–233. [Google Scholar] [CrossRef]
- Heinrich, J.M.; Irazoqui, H.A. Kinetic Model of Photoautotrophic Growth of Chlorella sp. Microalga, Isolated from the Setúbal Lagoon. Photochem. Photobiol. 2015, 91, 1095–1102. [Google Scholar] [CrossRef]
- Zemke, P.E.; Sommerfeld, M.R.; Hu, Q. Assessment of key biological and engineering design parameters for production of Chlorella zofingiensis (Chlorophyceae) in outdoor photobioreactors. Appl. Microbiol. Biotechnol. 2013, 97, 5645–5655. [Google Scholar] [CrossRef]
- Wong, Y.K.; Ho, K.C.; Tsang, Y.F.; Wang, L.; Yung, K.K.L. Cultivation of Chlorella vulgaris in Column Photobioreactor for Biomass Production and Lipid Accumulation. Water Environ. Res. 2016, 88, 40–46. [Google Scholar] [CrossRef]
- Huo, S.; Wang, Z.; Zhu, S.; Shu, Q.; Zhu, L.; Qin, L.; Zhou, W.; Feng, P.; Zhu, F.; Yuan, Z.; et al. Biomass Accumulation of Chlorella zofingiensis G1 Cultures Grown Outdoors in Photobioreactors. Front. Energy Res. 2018, 6, 21. [Google Scholar] [CrossRef] [Green Version]
- RKatiyar, R.; Gurjar, B.R.; Kumar, A.; Bharti, R.K.; Biswas, S.; Pruthi, V. A novel approach using low-cost Citrus limetta waste for mixotrophic cultivation of oleaginous microalgae to augment automotive quality biodiesel production. Environ. Sci. Pollut. Res. 2019, 26, 16115–16124. [Google Scholar] [CrossRef]
- Huang, Y.; Sun, Y.; Liao, Q.; Fu, Q.; Xia, A.; Zhu, X. Improvement on light penetrability and microalgae biomass production by periodically pre-harvesting Chlorella vulgaris cells with culture medium recycling. Bioresour. Technol. 2016, 216, 669–676. [Google Scholar] [CrossRef]
- Zhang, Z.; Huang, J.J.; Sun, D.; Lee, Y.; Chen, F. Two-step cultivation for production of astaxanthin in Chlorella zofingiensis using a patented energy-free rotating floating photobioreactor (RFP). Bioresour. Technol. 2017, 224, 515–522. [Google Scholar] [CrossRef]
- Imaizumi, Y.; Nagao, N.; Yusoff, F.M.; Taguchi, S.; Toda, T. Estimation of optimum specific light intensity per cell on a high-cell-density continuous culture of Chlorella zofingiensis not limited by nutrients or CO2. Bioresour. Technol. 2014, 162, 53–59. [Google Scholar] [CrossRef]
- Kim, U.; Cho, D.-H.; Heo, J.; Yun, J.-H.; Choi, D.-Y.; Cho, K.; Kim, H.-S. Two-stage cultivation strategy for the improvement of pigment productivity from high-density heterotrophic algal cultures. Bioresour. Technol. 2020, 302, 122840. [Google Scholar] [CrossRef]
- Patidar, S.K.; Mitra, M.; Goel, S.; Mishra, S. Effect of carbon supply mode on biomass and lipid in CSMCRI’s Chlorella variabilis (ATCC 12198). Biomass Bioenergy 2016, 86, 1–10. [Google Scholar] [CrossRef]
- Al Ketife, A.M.; Judd, S.; Znad, H. Optimization of cultivation conditions for combined nutrient removal and CO2 fixation in a batch photobioreactor. J. Chem. Technol. Biotechnol. 2016, 92, 1085–1093. [Google Scholar] [CrossRef] [Green Version]
- Sforza, E.; Cipriani, R.; Morosinotto, T.; Bertucco, A.; Giacometti, G.M. Excess CO2 supply inhibits mixotrophic growth of Chlorella protothecoides and Nannochloropsis salina. Bioresour. Technol. 2012, 104, 523–529. [Google Scholar] [CrossRef]
- Sun, Z.; Dou, X.; Wu, J.; He, B.; Wang, Y.; Chen, Y.-F. Enhanced lipid accumulation of photoautotrophic microalgae by high-dose CO2 mimics a heterotrophic characterization. World J. Microbiol. Biotechnol. 2016, 32, 9. [Google Scholar] [CrossRef]
- Grama, B.S.; Chader, S.; Khelifi, D.; Agathos, S.N.; Jeffryes, C. Induction of canthaxanthin production in a Dactylococcus microalga isolated from the Algerian Sahara. Bioresour. Technol. 2014, 151, 297–305. [Google Scholar] [CrossRef]
- Di Caprio, F.; Altimari, P.; Pagnanelli, F. Effect of Ca2+ concentration on Scenedesmus sp. growth in heterotrophic and photoautotrophic cultivation. New Biotechnol. 2018, 40, 228–235. [Google Scholar] [CrossRef]
- Schultze, L.K.P.; Simon, M.-V.; Li, T.; Langenbach, D.; Podola, B.; Melkonian, M. High light and carbon dioxide optimize surface productivity in a Twin-Layer biofilm photobioreactor. Algal Res. 2015, 8, 37–44. [Google Scholar] [CrossRef]
- Praveenkumar, R.; Kim, B.; Lee, J.; Vijayan, D.; Lee, K.; Nam, B.; Jeon, S.G.; Kim, D.-M.; Oh, Y.-K. Mild pressure induces rapid accumulation of neutral lipid (triacylglycerol) in Chlorella spp. Bioresour. Technol. 2016, 220, 661–665. [Google Scholar] [CrossRef]
- Kuo, C.-M.; Lin, T.-H.; Yang, Y.-C.; Zhang, W.-X.; Lai, J.-T.; Wu, H.-T.; Chang, J.-S.; Lin, C.-S. Ability of an alkali-tolerant mutant strain of the microalga Chlorella sp. AT1 to capture carbon dioxide for increasing carbon dioxide utilization efficiency. Bioresour. Technol. 2017, 244, 243–251. [Google Scholar] [CrossRef]
- Kim, Z.-H.; Kim, S.-H.; Lee, H.-S.; Lee, C.-G. Enhanced production of astaxanthin by flashing light using Haematococcus pluvialis. Enzym. Microb. Technol. 2006, 39, 414–419. [Google Scholar] [CrossRef]
- Schulze, P.S.; Brindley, C.; Fernández, J.M.; Rautenberger, R.; Pereira, H.; Wijffels, R.H.; Kiron, V. Flashing light does not improve photosynthetic performance and growth of green microalgae. Bioresour. Technol. Rep. 2020, 9, 100367. [Google Scholar] [CrossRef]
- Abiusi, F.; Sampietro, G.; Marturano, G.; Biondi, N.; Rodolfi, L.; D’Ottavio, M.; Tredici, M.R. Growth, photosynthetic efficiency, and biochemical composition of Tetraselmis suecica F&M-M33 grown with LEDs of different colors. Biotechnol. Bioeng. 2014, 111, 956–964. [Google Scholar] [CrossRef]
- Toledo-Cervantes, A.; Morales, M.; Novelo, E.; Revah, S. Carbon dioxide fixation and lipid storage by Scenedesmus obtusiusculus. Bioresour. Technol. 2013, 130, 652–658. [Google Scholar] [CrossRef]
- Chiu, S.-Y.; Tsai, M.-T.; Kao, C.-Y.; Ong, S.-C.; Lin, C.-S. The air-lift photobioreactors with flow patterning for high-density cultures of microalgae and carbon dioxide removal. Eng. Life Sci. 2009, 9, 254–260. [Google Scholar] [CrossRef]
- Tababa, H.G.; Hirabayashi, S.; Inubushi, K. Media optimization of Parietochloris incisa for arachidonic acid accumulation in an outdoor vertical tubular photobioreactor. J. Appl. Phycol. 2012, 24, 887–895. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Yadav, K.; Singh, R.S. Biofixation of carbon dioxide using mixed culture of microalgae. Indian J. Biotechnol. 2015, 14, 228–232. [Google Scholar]
- Katiyar, R.; Gurjar, B.; Bharti, R.K.; Kumar, A.; Biswas, S.; Pruthi, V. Heterotrophic cultivation of microalgae in photobioreactor using low cost crude glycerol for enhanced biodiesel production. Renew. Energy 2017, 113, 1359–1365. [Google Scholar] [CrossRef]
- Ogbonna, J.C.; Tanaka, H. Light requirement and photosynthetic cell cultivation—Development of processes for efficient light utilization in photobioreactors. J. Appl. Phycol. 2000, 12, 207–218. [Google Scholar] [CrossRef]
- Papazi, A.; Makridis, P.; Divanach, P.; Kotzabasis, K. Bioenergetic changes in the microalgal photosynthetic apparatus by extremely high CO2 concentrations induce an intense biomass production. Physiol. Plant. 2008, 132, 338–349. [Google Scholar] [CrossRef]
- CGrünewald, C.F.; Garcés, E.; Alacid, E.; Rossi, S.; Camp, J. Biomass and Lipid Production of Dinoflagellates and Raphidophytes in Indoor and Outdoor Photobioreactors. Mar. Biotechnol. 2013, 15, 37–47. [Google Scholar] [CrossRef]
- Fan, J.; Huang, J.; Li, Y.; Han, F.; Wang, J.; Li, X.; Wang, W.; Li, S. Sequential heterotrophy–dilution–photoinduction cultivation for efficient microalgal biomass and lipid production. Bioresour. Technol. 2012, 112, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Feng, P.; Deng, Z.; Hu, Z.; Fan, L. Lipid accumulation and growth of Chlorella zofingiensis in flat plate photobioreactors outdoors. Bioresour. Technol. 2011, 102, 10577–10584. [Google Scholar] [CrossRef]
- Feng, P.; Deng, Z.; Fan, L.; Hu, Z. Lipid accumulation and growth characteristics of Chlorella zofingiensis under different nitrate and phosphate concentrations. J. Biosci. Bioeng. 2012, 114, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.E.; Chang, W.S.; Patel, A.K.; Oh, M.S.; Lee, J.J.; Sim, S.J. Microalgal-Based Carbon Sequestration by Converting LNG-Fired Waste CO2 into Red Gold Astaxanthin: The Potential Applicability. Energies 2019, 12, 1718. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Zhang, W.; Zhang, X.; Amendola, P.; Hu, Q.; Chen, Y. Characterization of dissolved organic matters responsible for ultrafiltration membrane fouling in algal harvesting. Algal Res. 2013, 2, 223–229. [Google Scholar] [CrossRef]
- Řezanka, T.; Nedbalová, L.; Lukavský, J.; Střížek, A.; Sigler, K. Pilot cultivation of the green alga Monoraphidium sp. producing a high content of polyunsaturated fatty acids in a low-temperature environment. Algal Res. 2017, 22, 160–165. [Google Scholar] [CrossRef]
- Porcelli, R.; Dotto, F.; Pezzolesi, L.; Marazza, D.; Greggio, N.; Righi, S. Comparative life cycle assessment of microalgae cultivation for non-energy purposes using different carbon dioxide sources. Sci. Total Environ. 2020, 721, 137714. [Google Scholar] [CrossRef]
- Fuentes-Grünewald, C.; Bayliss, C.; Fonlut, F.; Chapuli, E. Long-term dinoflagellate culture performance in a commercial photobioreactor: Amphidinium carterae case. Bioresour. Technol. 2016, 218, 533–540. [Google Scholar] [CrossRef] [Green Version]
- Mohler, D.T.; Wilson, M.H.; Fan, Z.; Groppo, J.G.; Crocker, M. Beneficial Reuse of Industrial CO2 Emissions Using a Microalgae Photobioreactor: Waste Heat Utilization Assessment. Energies 2019, 12, 2634. [Google Scholar] [CrossRef] [Green Version]
- Bernard, O. Hurdles and challenges for modelling and control of microalgae for CO2 mitigation and biofuel production. J. Process Control 2011, 21, 1378–1389. [Google Scholar] [CrossRef]
- Chang, H.-X.; Huang, Y.; Fu, Q.; Liao, Q.; Zhu, X. Kinetic characteristics and modeling of microalgae Chlorella vulgaris growth and CO2 biofixation considering the coupled effects of light intensity and dissolved inorganic carbon. Bioresour. Technol. 2016, 206, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Chanona, E.A.D.R.; Liu, J.; Wagner, J.L.; Zhang, D.; Meng, Y.; Xue, S.; Shah, N. Dynamic modeling of green algae cultivation in a photobioreactor for sustainable biodiesel production. Biotechnol. Bioeng. 2018, 115, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Katiyar, R.; Bharti, R.K.; Gurjar, B.; Kumar, A.; Biswas, S.; Pruthi, V. Utilization of de-oiled algal biomass for enhancing vehicular quality biodiesel production from Chlorella sp. in mixotrophic cultivation systems. Renew. Energy 2018, 122, 80–88. [Google Scholar] [CrossRef]
- Zhao, Y.; Qiao, T.; Han, B.; Dong, X.; Geng, S.; Ning, D.; Ma, T.; Yu, X. Simultaneous enhancement of biomass and lipid production of Monoraphidium sp. QLZ-3 in a photobioreactor by using walnut shell extracts. Energy Convers. Manag. 2020, 204, 112326. [Google Scholar] [CrossRef]
- Do, T.-T.; Ong, B.-N.; Tran, M.-L.N.; Nguyen, D.; Melkonian, M.; Tran, H.-D. Biomass and Astaxanthin Productivities of Haematococcus pluvialis in an Angled Twin-Layer Porous Substrate Photobioreactor: Effect of Inoculum Density and Storage Time. Biology 2019, 8, 68. [Google Scholar] [CrossRef] [Green Version]
- Mazumdar, N.; Novis, P.M.; Visnovsky, G.; Gostomski, P. Effect of culturing parameters on the vegetative growth of Haematococcus alpinus (strain lcr-cc-261f) and modeling of its growth kinetics. J. Phycol. 2019, 55, 1071–1081. [Google Scholar] [CrossRef]
- Deniz, I. Scaling-up of Haematococcus pluvialis production in stirred tank photobioreactor. Bioresour. Technol. 2020, 310, 123434. [Google Scholar] [CrossRef]
- Fujii, K.; Nakajima, H.; Anno, Y. Potential of Monoraphidium sp. GK12 for energy-saving astaxanthin production. J. Chem. Technol. Biotechnol. 2008, 83, 1578–1584. [Google Scholar] [CrossRef]
- Yoo, J.J.; Choi, S.P.; Kim, J.Y.H.; Chang, W.S.; Sim, S.J. Development of thin-film photo-bioreactor and its application to outdoor culture of microalgae. Bioprocess Biosyst. Eng. 2013, 36, 729–736. [Google Scholar] [CrossRef]
- Zhu, C.; Zhai, X.; Wang, J.; Han, D.; Li, Y.; Xi, Y.; Tang, Y.; Chi, Z. Large-scale cultivation of Spirulina in a floating horizontal photobioreactor without aeration or an agitation device. Appl. Microbiol. Biotechnol. 2018, 102, 8979–8987. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Goh, Q.Y.; Tan, W.; Hossain, I.; Chen, W.N.; Lau, R. Lumostatic strategy for microalgae cultivation utilizing image analysis and chlorophyll α content as design parameters. Bioresour. Technol. 2011, 102, 6005–6012. [Google Scholar] [CrossRef]
- Ifrim, G.A.; Titica, M.; Boillereaux, L.; Caraman, S. Feedback linearizing control of light-to-microalgae ratio in artificially lighted photobioreactors. IFAC Proc. Vol. 2013, 46, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Imaizumi, Y.; Nagao, N.; Yusoff, F.; Kurosawa, N.; Kawasaki, N.; Toda, T. Lumostatic operation controlled by the optimum light intensity per dry weight for the effective production of Chlorella zofingiensis in the high cell density continuous culture. Algal Res. 2016, 20, 110–117. [Google Scholar] [CrossRef]
- Tran, N.-P.; Park, J.-K.; Kim, Z.-H.; Lee, C.-G. Influence of sodium orthovanadate on the production of astaxanthin from green algae Haematococcus lacustris. Biotechnol. Bioprocess Eng. 2009, 14, 322–329. [Google Scholar] [CrossRef]
- Tran, N.-P.; Park, J.-K.; Lee, C.-G. Proteomics analysis of proteins in green alga Haematococcus lacustris (Chlorophyceae) expressed under combined stress of nitrogen starvation and high irradiance. Enzym. Microb. Technol. 2009, 45, 241–246. [Google Scholar] [CrossRef]
- Lee, H.-S.; Kim, Z.-H.; Park, H.; Lee, C.-G. Specific light uptake rates can enhance astaxanthin productivity in Haematococcus lacustris. Bioprocess Biosyst. Eng. 2016, 39, 815–823. [Google Scholar] [CrossRef]
- Wu, K.; Ying, K.; Liu, L.; Zhou, J.; Cai, Z. High irradiance compensated with CO2 enhances the efficiency of Haematococcus lacustris growth. Biotechnol. Rep. 2020, 26, e00444. [Google Scholar] [CrossRef]
- Yoo, J.J.; Choi, S.P.; Kim, B.W.; Sim, S.J. Optimal design of scalable photo-bioreactor for phototropic culturing of Haematococcus pluvialis. Bioprocess Biosyst. Eng. 2012, 35, 309–315. [Google Scholar] [CrossRef]
- Sun, H.; Kong, Q.; Geng, Z.; Duan, L.; Yang, M.; Guan, B. Enhancement of cell biomass and cell activity of astaxanthin-rich Haematococcus pluvialis. Bioresour. Technol. 2015, 186, 67–73. [Google Scholar] [CrossRef]
- Ranjbar, R.; Inoue, R.; Shiraishi, H.; Katsuda, T.; Katoh, S. High efficiency production of astaxanthin by autotrophic cultivation of Haematococcus pluvialis in a bubble column photobioreactor. Biochem. Eng. J. 2008, 39, 575–580. [Google Scholar] [CrossRef]
- Sheng, B.; Fan, F.; Huang, J.; Bai, W.; Wang, J.; Li, S.; Li, W.; Wan, M.; Li, Y. Investigation on models for light distribution of Haematococcus pluvialis during astaxanthin accumulation stage with an application case. Algal Res. 2018, 33, 182–189. [Google Scholar] [CrossRef]
- Wan, M.; Zhang, Z.; Wang, J.; Huang, J.; Fan, J.; Yu, A.; Wang, W.; Li, Y. Sequential Heterotrophy–Dilution–Photoinduction Cultivation of Haematococcus pluvialis for efficient production of astaxanthin. Bioresour. Technol. 2015, 198, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, B.; Hu, Q.; Sommerfeld, M.; Li, Y.; Han, D. A new paradigm for producing astaxanthin from the unicellular green alga Haematococcus pluvialis. Biotechnol. Bioeng. 2016, 113, 2088–2099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.-E. Multistage Operation of Airlift Photobioreactor for Increased Production of Astaxanthin from Haematococcus pluvialis. J. Microbiol. Biotechnol. 2011, 21, 1081–1087. [Google Scholar] [CrossRef] [Green Version]
- Pérez-López, P.; González-García, S.; Jeffryes, C.; Agathos, S.N.; McHugh, E.; Walsh, D.; Murray, P.; Moane, S.; Feijoo, G.; Moreira, M.T. Life cycle assessment of the production of the red antioxidant carotenoid astaxanthin by microalgae: From lab to pilot scale. J. Clean. Prod. 2014, 64, 332–344. [Google Scholar] [CrossRef] [Green Version]
- Haque, F.; Dutta, A.; Thimmanagari, M.; Chiang, Y.W. Integrated Haematococcus pluvialis biomass production and nutrient removal using bioethanol plant waste effluent. Process Saf. Environ. Prot. 2017, 111, 128–137. [Google Scholar] [CrossRef]
- Onorato, C.; Rösch, C. Comparative life cycle assessment of astaxanthin production with Haematococcus pluvialis in different photobioreactor technologies. Algal Res. 2020, 50, 102005. [Google Scholar] [CrossRef]
- Yang, Z.; Cheng, J.; Li, K.; Zhou, J.; Cen, K. Optimizing gas transfer to improve growth rate of Haematococcus pluvialis in a raceway pond with chute and oscillating baffles. Bioresour. Technol. 2016, 214, 276–283. [Google Scholar] [CrossRef]
- Lee, H.S.; Kim, Z.H.; Jung, S.E.; Kim, J.D.; Lee, C.G. Specific light uptake rate can be served as a scale-up parameter in photobioreactor operations. J. Microbiol. Biotechnol. 2006, 16, 1890–1896. [Google Scholar]
- Wang, J.; Sommerfeld, M.R.; Lu, C.; Hu, Q. Combined effect of initial biomass density and nitrogen concentration on growth and astaxanthin production of Haematococcus pluvialis (Chlorophyta) in outdoor cultivation. Algae 2013, 28, 193–202. [Google Scholar] [CrossRef]
- Choi, Y.Y.; Joun, J.M.; Lee, J.; Hong, M.E.; Pham, H.-M.; Chang, W.S.; Sim, S.J. Development of large-scale and economic pH control system for outdoor cultivation of microalgae Haematococcus pluvialis using industrial flue gas. Bioresour. Technol. 2017, 244, 1235–1244. [Google Scholar] [CrossRef]
- Ranjbar, R.; Inoue, R.; Katsuda, T.; Yamaji, H.; Katoh, S. High efficiency production of astaxanthin in an airlift photobioreactor. J. Biosci. Bioeng. 2008, 106, 204–207. [Google Scholar] [CrossRef] [PubMed]
- Vega-Estrada, J.; Montes-Horcasitas, M.C.; Domínguez-Bocanegra, A.R.; Cañizares-Villanueva, R.O. Haematococcus pluvialis cultivation in split-cylinder internal-loop airlift photobioreactor under aeration conditions avoiding cell damage. Appl. Microbiol. Biotechnol. 2005, 68, 31–35. [Google Scholar] [CrossRef]
- Kim, Z.-H.; Park, H.; Lee, H.-S.; Lee, C.-G. Enhancing Photon Utilization Efficiency for Astaxanthin Production from Haematococcus lacustris Using a Split-Column Photobioreactor. J. Microbiol. Biotechnol. 2016, 26, 1285–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torzillo, G.; Goksan, T.; Faraloni, C.; Kopecky, J.; Masojídek, J. Interplay between photochemical activities and pigment composition in an outdoor culture of Haematococcus pluvialis during the shift from the green to red stage. J. Appl. Phycol. 2003, 15, 127–136. [Google Scholar] [CrossRef]
- Wang, F.; Gao, B.; Wu, M.; Huang, L.; Zhang, C. A novel strategy for the hyper-production of astaxanthin from the newly isolated microalga Haematococcus pluvialis JNU35. Algal Res. 2019, 39, 101466. [Google Scholar] [CrossRef]
- Kiperstok, A.; Sebestyén, P.; Podola, B.; Melkonian, M. Biofilm cultivation of Haematococcus pluvialis enables a highly productive one-phase process for astaxanthin production using high light intensities. Algal Res. 2017, 21, 213–222. [Google Scholar] [CrossRef]
- Imamoglu, E.; Dalay, M.C.; Sukan, F.V. Semi-continuous Cultivation of Haematococcus pluvialis for Commercial Production. Appl. Biochem. Biotechnol. 2010, 160, 764–772. [Google Scholar] [CrossRef]
- Fujita, T.; Aoyagi, H.; Ogbonna, J.C.; Tanaka, H. Effect of mixed organic substrate on α-tocopherol production by Euglena gracilis in photoheterotrophic culture. Appl. Microbiol. Biotechnol. 2008, 79, 371–378. [Google Scholar] [CrossRef]
- Kim, Z.-H.; Park, H.; Hong, S.-J.; Lim, S.-M.; Lee, C.-G. Development of a floating photobioreactor with internal partitions for efficient utilization of ocean wave into improved mass transfer and algal culture mixing. Bioprocess Biosyst. Eng. 2016, 39, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Giannelli, L.; Yamada, H.; Katsuda, T.; Yamaji, H. Effects of temperature on the astaxanthin productivity and light harvesting characteristics of the green alga Haematococcus pluvialis. J. Biosci. Bioeng. 2015, 119, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.Y.; Geng, Y.H.; Li, Z.K.; Hu, H.J.; Li, Y.G. Production of astaxanthin from Haematococcus in open pond by two-stage growth one-step process. Aquaculture 2009, 295, 275–281. [Google Scholar] [CrossRef]
- Choi, S.-L.; Suh, I.S.; Lee, C.-G. Lumostatic operation of bubble column photobioreactors for Haematococcus pluvialis cultures using a specific light uptake rate as a control parameter. Enzym. Microb. Technol. 2003, 33, 403–409. [Google Scholar] [CrossRef]
- Gao, X.; Wang, X.; Li, H.; Roje, S.; Sablani, S.S.; Chen, S. Parameterization of a light distribution model for green cell growth of microalgae: Haematococcus pluvialis cultured under red LED lights. Algal Res. 2017, 23, 20–27. [Google Scholar] [CrossRef]
- Jin, H.; Zhang, H.; Zhou, Z.; Li, K.; Hou, G.; Xu, Q.; Chuai, W.; Zhang, C.; Han, D.; Hu, Q. Ultrahigh-cell-density heterotrophic cultivation of the unicellular green microalga Scenedesmus acuminatus and application of the cells to photoautotrophic culture enhance biomass and lipid production. Biotechnol. Bioeng. 2020, 117, 96–108. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.-H.; Lee, Y.-K. Two-step process for ketocarotenoid production by a green alga, Chlorococcum sp. strain MA-1. Appl. Microbiol. Biotechnol. 2001, 55, 537–540. [Google Scholar] [CrossRef]
- Fu, W.; Gudmundsson, O.; Feist, A.M.; Herjolfsson, G.; Brynjolfsson, S.; Palsson, B. Maximizing biomass productivity and cell density of Chlorella vulgaris by using light-emitting diode-based photobioreactor. J. Biotechnol. 2012, 161, 242–249. [Google Scholar] [CrossRef]
- Wu, L.; Li, Z.; Song, Y. Hydrodynamic conditions in designed spiral photobioreactors. Bioresour. Technol. 2010, 101, 298–303. [Google Scholar] [CrossRef]
- Liang, F. Growth Rate and Biomass Productivity of Chlorella as Affected by Culture Depth and Cell Density in an Open Circular Photobioreactor. J. Microbiol. Biotechnol. 2013, 23, 539–544. [Google Scholar] [CrossRef] [Green Version]
- Grünewald, C.F.; Bayliss, C.; Zanain, M.; Pooley, C.; Scolamacchia, M.; Silkina, A. Evaluation of batch and semi-continuous culture of Porphyridium purpureum in a photobioreactor in high latitudes using Fourier Transform Infrared spectroscopy for monitoring biomass composition and metabolites production. Bioresour. Technol. 2015, 189, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Altunoz, M.; Pirrotta, O.; Forti, L.; Allesina, G.; Pedrazzi, S.; Obali, O.; Tartarini, P.; Arru, L. Combined effects of LED lights and chicken manure on Neochloris oleoabundans growth. Bioresour. Technol. 2017, 244, 1261–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohrabi, D.; Jazini, M.H.; Shariati, M. Mixotrophic Cultivation of Dunaliella salina on Crude Glycerol Obtained from Calcinated Fatty Acid Production Process. Russ. J. Mar. Biol. 2019, 45, 470–480. [Google Scholar] [CrossRef]
- Arashiro, L.T.; Boto-Ordóñez, M.; Van Hulle, S.W.; Ferrer, I.; Garfí, M.; Rousseau, D.P. Natural pigments from microalgae grown in industrial wastewater. Bioresour. Technol. 2020, 303, 122894. [Google Scholar] [CrossRef] [Green Version]
- Aray-Andrade, M.; Moreira, C.; Santander, V.; Mendoza, L.; Bermúdez, R. Characterization of three algal strains used as a tertiary treatment for rural wastewater of ecuadorian littoral. Eur. Biomass Conf. Exhib. Proc. 2019, 241–248. [Google Scholar] [CrossRef]
- García, J.; Ortiz, A.; Álvarez, E.; Belohlav, V.; García-Galán, M.J.; Díez-Montero, R.; Álvarez, J.A.; Uggetti, E. Nutrient removal from agricultural run-off in demonstrative full scale tubular photobioreactors for microalgae growth. Ecol. Eng. 2018, 120, 513–521. [Google Scholar] [CrossRef] [Green Version]
- de Mendonça, H.V.; Ometto, J.P.H.B.; Otenio, M.H.; Marques, I.P.R.; dos Reis, A.J.D. Microalgae-mediated bioremediation and valorization of cattle wastewater previously digested in a hybrid anaerobic reactor using a photobioreactor: Comparison between batch and continuous operation. Sci. Total Environ. 2018, 633, 1–11. [Google Scholar] [CrossRef] [Green Version]
- García-Galán, M.J.; Monllor-Alcaraz, L.S.; Postigo, C.; Uggetti, E.; de Alda, M.L.; Díez-Montero, R.; García, J. Microalgae-based bioremediation of water contaminated by pesticides in peri-urban agricultural areas. Environ. Pollut. 2020, 265, 114579. [Google Scholar] [CrossRef]
- Arashiro, L.T.; Ferrer, I.; Pániker, C.C.; Gómez-Pinchetti, J.L.; Rousseau, D.P.L.; Van Hulle, S.W.H.; Garfí, M. Natural Pigments and Biogas Recovery from Microalgae Grown in Wastewater. ACS Sustain. Chem. Eng. 2020, 8, 10691–10701. [Google Scholar] [CrossRef]
- Cardoso, L.G.; Duarte, J.H.; Costa, J.A.V.; Assis, D.; Lemos, P.V.F.; Druzian, J.I.; de Souza, C.O.; Nunes, I.L.; Chinalia, F.A. Spirulina sp. as a Bioremediation Agent for Aquaculture Wastewater: Production of High Added Value Compounds and Estimation of Theoretical Biodiesel. Bioenergy Res. 2021, 14, 254–264. [Google Scholar] [CrossRef]
- Rooke, J.C.; Léonard, A.; Su, B.-L. Targeting photobioreactors: Immobilisation of cyanobacteria within porous silica gel using biocompatible methods. J. Mater. Chem. 2008, 18, 1333–1341. [Google Scholar] [CrossRef]
- Liu, T.; Wang, J.; Hu, Q.; Cheng, P.; Ji, B.; Liu, J.; Chen, Y.; Zhang, W.; Chen, X.; Chen, L.; et al. Attached cultivation technology of microalgae for efficient biomass feedstock production. Bioresour. Technol. 2013, 127, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Wang, J.; Zhang, W.; Liu, J.; Wang, H.; Gao, L.; Liu, T. An applicable nitrogen supply strategy for attached cultivation of Aucutodesmus obliquus. J. Appl. Phycol. 2014, 26, 173–180. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, J.; Wang, J.; Liu, T. Attached cultivation of Haematococcus pluvialis for astaxanthin production. Bioresour. Technol. 2014, 158, 329–335. [Google Scholar] [CrossRef]
- Piltz, B.; Melkonian, M. Immobilized microalgae for nutrient recovery from source-separated human urine. J. Appl. Phycol. 2018, 30, 421–429. [Google Scholar] [CrossRef]
- Xie, X.; Huang, A.; Gu, W.; Zang, Z.; Pan, G.; Gao, S.; He, L.; Zhang, B.; Niu, J.; Lin, A.; et al. Photorespiration participates in the assimilation of acetate in Chlorella sorokiniana under high light. New Phytol. 2016, 209, 987–998. [Google Scholar] [CrossRef] [Green Version]
- Li, F.-F.; Yang, Z.-H.; Zeng, R.; Yang, G.; Chang, X.; Yan, J.-B.; Hou, Y.-L. Microalgae Capture of CO2 from Actual Flue Gas Discharged from a Combustion Chamber. Ind. Eng. Chem. Res. 2011, 50, 6496–6502. [Google Scholar] [CrossRef]
- Sforza, E.; Gris, B.; de Farias Silva, C.; Morosinotto, T.; Bertucco, A. Effects of light on cultivation of Scenedesmus obliquus in batch and continuous flat plate photobioreactor. Chem. Eng. Trans. 2014, 38, 211–216. [Google Scholar] [CrossRef]
- Lee, J.Y.; Hong, M.-E.; Chang, W.S.; Sim, S.J. Enhanced carbon dioxide fixation of Haematococcus pluvialis using sequential operating system in tubular photobioreactors. Process Biochem. 2015, 50, 1091–1096. [Google Scholar] [CrossRef]
- Guyon, J.-B.; Vergé, V.; Schatt, P.; Lozano, J.-C.; Liennard, M.; Bouget, F.-Y. Comparative Analysis of Culture Conditions for the Optimization of Carotenoid Production in Several Strains of the Picoeukaryote Ostreococcus. Mar. Drugs 2018, 16, 76. [Google Scholar] [CrossRef] [Green Version]
- Morales, M.; Sánchez, L.; Revah, S. The impact of environmental factors on carbon dioxide fixation by microalgae. FEMS Microbiol. Lett. 2018, 365, fnx262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masojídek, J.; Kopecká, J.; Koblížek, M.; Torzillo, G. The Xanthophyll Cycle in Green Algae (Chlorophyta): Its Role in the Photosynthetic Apparatus. Plant Biol. 2004, 6, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Huntley, M.E.; Johnson, Z.I.; Brown, S.L.; Sills, D.L.; Gerber, L.; Archibald, I.; Machesky, S.C.; Granados, J.; Beal, C.; Greene, C.H. Demonstrated large-scale production of marine microalgae for fuels and feed. Algal Res. 2015, 10, 249–265. [Google Scholar] [CrossRef] [Green Version]
- Kunjapur, A.M.; Eldridge, R.B. Photobioreactor Design for Commercial Biofuel Production from Microalgae. Ind. Eng. Chem. Res. 2010, 49, 3516–3526. [Google Scholar] [CrossRef]
- Krug, L.; Erlacher, A.; Berg, G.; Cernava, T. A novel, nature-based alternative for photobioreactor decontaminations. Sci. Rep. 2019, 9, 2864. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author | Number of Articles | Citations | Total Link Strength | Normalized Citations | Average Year | Average Citation |
|---|---|---|---|---|---|---|
| Chang J.-S. | 6 | 1317 | 0 | 14.33 | 2014.83 | 219.50 |
| Wang J. | 9 | 519 | 15 | 13.26 | 2015.00 | 56.67 |
| Liu J. | 8 | 414 | 11 | 11.89 | 2015.50 | 51.75 |
| Sim S.J. | 10 | 227 | 0 | 11.50 | 2015.10 | 22.70 |
| Hu Q. | 7 | 349 | 8 | 10.66 | 2014.71 | 49.86 |
| Li Y. | 9 | 240 | 6 | 10.42 | 2015.89 | 26.67 |
| Lee C.-G. | 17 | 613 | 22 | 9.79 | 2010.35 | 36.06 |
| Zhang W. | 5 | 456 | 12 | 9.05 | 2013.40 | 91.20 |
| Liu T. | 5 | 428 | 12 | 8.87 | 2013.80 | 85.60 |
| Lee H.-S. | 5 | 105 | 12 | 7.34 | 2010.00 | 21.00 |
| Melkonian M. | 6 | 123 | 0 | 7.34 | 2017.67 | 20.50 |
| Morosinotto T. | 5 | 422 | 0 | 7.27 | 2013.00 | 84.40 |
| Kim Z.-H. | 11 | 167 | 22 | 5.70 | 2012.91 | 15.18 |
| Sun Z. | 6 | 184 | 2 | 5.69 | 2014.83 | 30.67 |
| Kumar A. | 5 | 82 | 0 | 5.47 | 2017.60 | 16.40 |
| Katsuda T. | 5 | 167 | 0 | 3.82 | 2010.40 | 33.40 |
| Guerrero M.G. | 5 | 673 | 0 | 3.59 | 2007.40 | 134.60 |
| Park H. | 6 | 61 | 14 | 2.75 | 2016.33 | 10.17 |
| Jeffryes C. | 5 | 58 | 0 | 1.65 | 2015.40 | 11.60 |
| Powtongsook S. | 5 | 73 | 0 | 1.47 | 2014.40 | 14.04 |
| Authors | Year | Cited by | Normalized Citations | % of Total Sum of Citations (n = 26,015) |
|---|---|---|---|---|
| Chisti Y. [32] | 2007 | 6137 | 472.08 | 23.59 |
| Brennan L., Owende P. [33] | 2010 | 2695 | 269.50 | 10.36 |
| Chen C.-Y., Yeh K.-L., Aisyah R., Lee D.-J., Chang J.-S. [34] | 2011 | 1088 | 120.89 | 4.18 |
| Ugwu C.U., Aoyagi H., Uchiyama H. [35] | 2008 | 660 | 55.00 | 2.54 |
| Greenwell H.C., Laurens L.M.L., Shields R.J., Lovitt R.W., Flynn K.J. [36] | 2010 | 527 | 52.70 | 2.03 |
| Posten C. [37] | 2009 | 412 | 37.45 | 1.58 |
| Borowitzka, M.A. [38] | 1999 | 743 | 35.38 | 2.86 |
| Del Campo J.A., García-González M., Guerrero M.G. [39] | 2007 | 434 | 33.38 | 1.67 |
| Kumar K., Dasgupta C.N., Nayak B., Lindblad P., Das D. [40] | 2011 | 287 | 31.89 | 1.10 |
| Suali E., Sarbatly R. [41] | 2012 | 239 | 29.88 | 0.92 |
| Carvalho A.P., Silva S.O., Baptista J.M., Malcata F.X. [42] | 2011 | 229 | 25.44 | 0.88 |
| Lee Y.-K. [43] | 2001 | 355 | 18.68 | 1.36 |
| Janssen M., Tramper J., Mur L.R., Wijffels R.H. [44] | 2003 | 310 | 18.24 | 1.19 |
| Apt K.E., Behrens P.W. [45] | 1999 | 268 | 12.76 | 1.03 |
| Sum of citations of top 14 documents | 11,426 | 43.92 | ||
| Rest of the documents | 14,589 | 56.08 | ||
| Country | Documents | Total Citations | Normalized Citations | Average Year | Average Citation | Average Normalized Citations |
|---|---|---|---|---|---|---|
| China | 64 | 2351 | 76.46 | 2015.34 | 36.73 | 1.19 |
| United States | 40 | 2566 | 40.61 | 2013.97 | 64.15 | 1.02 |
| Australia | 20 | 478 | 29.26 | 2015.25 | 23.90 | 1.46 |
| Spain | 28 | 1773 | 29.07 | 2012.46 | 63.32 | 1.04 |
| South Korea | 33 | 911 | 27.14 | 2012.79 | 27.61 | 0.82 |
| Italy | 22 | 830 | 25.82 | 2016.50 | 37.73 | 1.17 |
| India | 25 | 776 | 23.63 | 2016.76 | 31.04 | 0.95 |
| Germany | 18 | 787 | 17.83 | 2015.56 | 43.72 | 0.99 |
| Japan | 15 | 1195 | 17.31 | 2010.80 | 79.67 | 1.15 |
| Portugal | 12 | 614 | 15.29 | 2015.58 | 51.17 | 1.27 |
| Taiwan | 7 | 1387 | 15.07 | 2014.00 | 198.14 | 2.15 |
| United Kingdom | 11 | 728 | 13.89 | 2016.73 | 66.18 | 1.26 |
| Netherlands | 7 | 458 | 10.00 | 2016.29 | 65.43 | 1.43 |
| France | 12 | 589 | 9.72 | 2013.33 | 49.08 | 0.81 |
| New Zealand | 6 | 6517 | 9.23 | 2008.33 | 1086.17 | 1.54 |
| Belgium | 10 | 190 | 6.62 | 2015.20 | 19.00 | 0.66 |
| Thailand | 6 | 101 | 5.40 | 2015.17 | 16.83 | 0.90 |
| Malaysia | 10 | 481 | 4.83 | 2016.30 | 48.10 | 1.80 |
| Iran | 5 | 53 | 4.83 | 2018.20 | 10.60 | 0.97 |
| Turkey | 7 | 48 | 3.59 | 2016.43 | 6.86 | 0.51 |
| Brazil | 7 | 52 | 3.34 | 2017.00 | 7.43 | 0.48 |
| Canada | 6 | 52 | 2.72 | 2016.50 | 8.67 | 0.45 |
| Authors’ Keyword | Occurren-ces | Total Link Strength | Average Year | Average Citation | Average Normalized Citation | Cluster |
|---|---|---|---|---|---|---|
| irradiance | 7 | 6 | 2013.14 | 25.29 | 2.24 | 4 |
| photosynthetic efficiency | 6 | 5 | 2013.33 | 103.00 | 2.24 | 5 |
| astaxanthin | 52 | 51 | 2011.65 | 40.29 | 1.78 | 4 |
| attached cultivation | 5 | 4 | 2014.60 | 64.00 | 1.78 | 5 |
| harvesting | 5 | 5 | 2015.60 | 224.40 | 1.74 | 3 |
| bioenergy | 7 | 6 | 2013.86 | 402.86 | 1.73 | 1 |
| lipids | 18 | 18 | 2015.78 | 40.11 | 1.59 | 1 |
| pigments | 6 | 5 | 2013.00 | 58.00 | 1.57 | 1 |
| stress | 5 | 5 | 2012.80 | 63.60 | 1.45 | 6 |
| heterotrophy | 6 | 5 | 2014.67 | 73.17 | 1.44 | 7 |
| Nannochloropsis | 6 | 6 | 2013.00 | 79.17 | 1.34 | 1 |
| algae | 17 | 16 | 2015.00 | 87.41 | 1.29 | 2 |
| photosynthesis | 9 | 8 | 2016.00 | 52.78 | 1.28 | 1 |
| biodiesel | 30 | 29 | 2013.27 | 273.73 | 1.25 | 3 |
| Haematococcus | 11 | 10 | 2010.36 | 45.09 | 1.25 | 4 |
| microalgae | 145 | 118 | 2014.55 | 116.26 | 1.25 | 5 |
| biomass | 26 | 24 | 2014.35 | 71.50 | 1.23 | 1 |
| biorefineries | 9 | 9 | 2015.67 | 80.11 | 1.19 | 1 |
| biofuels | 35 | 35 | 2014.46 | 235.17 | 1.16 | 1 |
| Haematococcus pluvialis | 47 | 43 | 2012.87 | 34.32 | 1.15 | 4 |
| photobioreactors | 132 | 115 | 2013.35 | 131.33 | 1.15 | 5 |
| flashing light | 6 | 6 | 2013.83 | 29.33 | 1.12 | 1 |
| continuous culture | 5 | 5 | 2011.82 | 24.00 | 1.08 | 4 |
| cyanobacteria | 10 | 9 | 2014.90 | 52.10 | 1.08 | 5 |
| light intensity | 6 | 6 | 2015.67 | 46.17 | 1.08 | 2 |
| microalgal cultivations | 7 | 6 | 2013.71 | 55.71 | 1.05 | 3 |
| scale up | 11 | 10 | 2014.73 | 36.91 | 1.02 | 4 |
| wastewater treatment | 7 | 7 | 2018.00 | 20.57 | 1.02 | 5 |
| Chlorella zofingiensis | 10 | 8 | 2014.00 | 52.10 | 1.01 | 3 |
| lumostatic operation | 5 | 5 | 2009.40 | 26.20 | 0.95 | 4 |
| productivity | 7 | 7 | 2014.86 | 25.10 | 0.95 | 5 |
| fatty acids | 9 | 8 | 2014.78 | 30.67 | 0.82 | 6 |
| outdoor culture | 6 | 6 | 2008.83 | 71.17 | 0.81 | 3 |
| carotenoids | 21 | 20 | 2012.62 | 27.62 | 0.73 | 6 |
| CO2 | 16 | 15 | 2015.44 | 24.56 | 0.69 | 6 |
| tubular photobioreactors | 7 | 6 | 2009.71 | 30.14 | 0.68 | 2 |
| lutein | 16 | 13 | 2015.12 | 36.94 | 0.62 | 2 |
| Chlorella | 16 | 15 | 2011.81 | 69.69 | 0.60 | 3 |
| flue gases | 5 | 5 | 2016.20 | 10.40 | 0.60 | 2 |
| lipid content | 5 | 4 | 2016.20 | 12.40 | 0.46 | 3 |
| growth rate | 6 | 6 | 2014.50 | 8.67 | 0.30 | 6 |
| airlift reactor | 5 | 5 | 2013.40 | 20.40 | 0.23 | 2 |
| light | 5 | 5 | 2017.40 | 63.60 | 0.19 | 6 |
| Chlamydomonas | 6 | 6 | 2013.17 | 18.00 | 0.15 | 2 |
| Description | Costs |
|---|---|
| Techno-economical assessment of bioethanol commercialization from Chlorella vulgaris in Brunei Darussalam. Two types of cultivation systems, namely, closed-system (photobioreactor—PBR) and open-pond approaches, were anticipated for an approximate total biomass of 220 t year−1 on 6 ha coastal areas. The biomass productivity was 56 t·ha−1 for the PBR and 28 t·ha−1 for the pond annually. The plant output was 58.90 m3·ha−1 for the PBR and 24.9 m3·ha−1 for the pond annually. The total bioethanol output of the plant was 57,087.58 gal·year−1, along with the value-added by-products (crude bioliquid and slurry cake). | The total production cost of this project was USD 2.22 million for bioethanol from microalgae, and the total bioethanol selling price was USD 2.22–2.87 million, along with a by-product sale price of USD 1.6 million [52,53]. |
| Three production technologies were investigated: (1) open raceway ponds (ORPs), (2) tubular photobioreactors (PBRs) and (3) systems coupling photobioreactors to open raceway ponds. | The greatest cost growth (1.5–1.8) was estimated for PBR systems, while the lowest cost growth (1.2–1.4) was estimated for the ORP systems and coupled systems. Plant performance ranged from 13% to 40% of nameplate capacity [61]. |
| Assumption: to leave scale out of the analysis to avoid disproportionate comparisons. | There was nearly a 25-fold difference in total cost per hectare between the ORP and PBR/PBR–ORP systems modeled, even with similar cost growth. If the algae were only 30% oil, the unit cost growth factor for algal oil would be somewhere between 10 and 46 times the proposed price [62]. |
| Four production pathways, ranging from a base case with commercial technologies to an improved case with innovative technologies, were studied. All region-specific parameters were adapted to Belgian conditions. | The lowest carotenoid price for which the process had a positive NPV in the different scenarios, keeping all other parameters constant, was EUR 1059 per kg β-carotene (base scenario), EUR 657 per kg β-carotene (intermediate scenario), EUR 1379 per kg β-carotene (advanced scenario) and EUR 4725 per kg astaxanthin (alternative scenario). The most profitable scenario was the scenario that uses a specialized membrane for medium recycling and an open-pond algae cultivation [63]. |
| Type of Reactor | Description |
|---|---|
| Closed continuous-run tubular loop bioreactors | Connected with pipes with a diameter usually less than 0.08 m [65]. |
| Vertical column reactors | Column diameter > 0.1 m [65]. |
| Outdoor bubble columns and airlift bioreactors | 0.19 m column diameter, 2 m tall, 0.06 m3 working volume; outdoor bubble column and airlift photobioreactors [65]. |
| Panel photobioreactor | C-shaped flat vessel of various widths and heights (usually less than 1 m) and a thickness of 1–5 cm, made of transparent materials. PBR flat panels are positioned vertically or tilted towards the sun [44]. |
| Hybrid reactors | Thin-film PBRs combined with bubble columns [66]. |
| PBRs with internal lighting | Lighting in the form of submerged fluorescent lamps, LEDs, optical fibers or waveguides is a solution to the shading of the deeper layers of microalgae cultures [67]. |
| The phosphorescent materials inside the PBR emit light in the dark phase, extending the microalgae culture’s illumination time. | The phosphorescent materials inside the PBR increased cells’ specific growth rate and dry mass by about 9% and 24%, respectively, due to the light emission of these materials in the dark phase [68]. |
| Tubular PBR using dual Fresnel linear lenses | Lenses concentrated sunlight on the surface of glass culture tubes placed on a movable frame installed on a vertical wall and under a sloping roof window (42° angle). An automatic tracking system kept the glass cultivation tubes in the focus of the collector lenses as the sun changed its position. The use of collectors on vertical walls was beneficial in spring and autumn when the angle of inclination of the sun was small. In spring, the total radiation energy collected by both types of panels was similar. In summer, (sloping) roof collectors delivered, on average, 4–12 times higher radiation intensity on the pipe surfaces than vertical collectors [69]. |
| Unusual geometric arrangements of 5-L X-shaped and H-shaped photobioreactors with two aeration bubblers and four serially arranged and connected columns with one bubbler in each column were tested | Increases in biomass and lipid production and high content of induced monounsaturated fatty acids were observed [70]. |
| Transparent and gas-permeable micro-reactors made of poly(dimethylsiloxane) | There are special chambers and microchannels that allow obtaining the height of the culture substrate at a level of 3.5 mm [71]. |
| Description | Achievements |
|---|---|
| Biomass production in three types of vertical reactors with identical dimensions: bubble column, split-cylinder airlift device and a draft-tube airlift bioreactor. | The results proved that cells in all types of PBR were subjected to identical values of mean intensity of irradiation [65]. |
| Compared light gradients and mixed light/dark (L/D) cycles and the performance of an aeration column, tubular reactor and flatbed reactor. | The light regimes and productivity in aerated and airlift column reactors were similar to each other [44]. |
| Compared the results of H. pluvialis cultures in column and tube reactors in order to find the best reactor for outdoor production of astaxanthin. | Better biomass and astaxanthin productivity results were obtained in tubular photobioreactors [74]. |
| Compared the mixing time, volumetric mass transfer coefficient kLa and the microalgae growth profile for an aerated column and airlift photobioreactor. | Better cell growth efficiency was observed in the airlift PBR, in which microalgae cells spent only 2 s in the riser (dark zone) and the remaining 10 s (or 84% of the time) in the downcomer (light zone) in a 12 s mixing cycle [75]. |
| Studies of the influence of the geometric shape of photobioreactors on the cultivation of Cylindrotheca closterium diatoms. A bag photobioreactor made of polyethylene, a flat one made of plexiglass and a bubble made of glass were tested experimentally. | The highest values of cell and chlorophyll-a concentration, specific growth rate and doubling time were achieved in a bag photobioreactor [76]. |
| Comparison of airlift photobioreactors and an agitated photobioreactor with the same working volume and operating conditions. | Due to the higher liquid height and different fluid flow patterns, culture in airlift photobioreactors resulted in a higher concentration of biomass and carotenoids at the end of the culture than when using an agitated photobioreactor with the same working volume and operating conditions [77]. |
| Proposed a hybrid photobioreactor made of an opaque, columnar bubble reactor connected to a lighting platform built of eight open tubular structures arranged in a linear manner, with an inclination of 9°. The lighting platform was connected by a flow pump to the bubble column. | The hybrid PBR requires a much smaller area, ensuring the appropriate surface-to-volume ratio, which is extremely important, especially when scaling the solution to industrial conditions [66]. |
| Study on the concentration of light in a PBR. They used waveguides illuminated by external light using three different specially designed and 3D-printed types of paraboloid mirrors to cultivate algal biofilm. | The productivity of algae biofilm biomass on the waveguide was 2.5 times higher when using mirrored concentrators [67]. |
| Successfully tested a novel double-layered column photobioreactor consisting of two glass tubes, allowing the simultaneous growth of H. pluvialis microalgae cells and astaxanthin accumulation. An inner tube was destined for the vegetative growth of cells, which received light energy reduced by mutual shading by cells in the outer mantle. | An excessive amount of light reached the outer layer, which increased the accumulation of astaxanthin in the cells [78]. |
| Study of a photobioreactor built from one tube inside the other but arranged horizontally, with aeration holes in the inner tube for both aeration and mixing of the culture. | The new photobioreactor assured the mitigation of changes in pH and dissolved oxygen along with the extension of the cultivation time, which resulted in obtaining high biomass productivity [79]. |
| Proposed new construction of a flat airlift photobioreactor, the volume scaling of which consisted of increasing its length to deal with the problem of losing hydrodynamic properties with increasing culture scale. | The 90 L PBR performed just as well as the 17 L one, only with a slightly slower growth rate. The results of the economic analysis showed that although the operation of the 90 L PBR was associated with higher utility costs, the unit cost of production was lower due to the large number of cells grown in one batch. In smaller systems, it was necessary to repeat the process several times to obtain the same number of cells [80]. |
| Description of the Culture Conditions or Parameters | Effect on Biomass Growth |
|---|---|
| The growth kinetics of microalgae and lipid synthesis for various temperatures and light colors were tested. | Red-orange light at a temperature of 24 °C obtained a 38% higher biomass productivity than blue light. In contrast, blue light at 32 °C was 13% more productive than red-orange light. The accumulation of lipids was favored by red-orange light and a temperature of 30–32 °C [87]. |
| Tetradesmus acuminatus culture conducted in a flat-plate PBR was assessed. The increase in biomass and its relationship with the maximization of carotenoid production (astaxanthin and β-carotene) under the influence of light intensity, photoperiod, pH, NaCl and nitrogen concentration were studied. | The increase in light intensity and irradiation time favored the growth of microalgae biomass and resulted in the highest concentration of carotenoids. The maximum production of carotenoids in mg g−1 was observed at 85 μmol m−2 s−1 light intensity and a light/dark photoperiod of 15.8:8.2 h, and in mg L−1, it was at 595 μmol m−2 s−1 light intensity for 24 h [88]. |
| The correlation between aeration and lighting was also experimentally verified in Scenedesmus obtusus microalgae culture conducted in an airlift photobioreactor with different inlet gas flow rates, different light intensities and different light/dark cycles. | Higher inlet gas flow rates (0.88 and 1.17 vvm) resulted in higher biomass productivity. The maximum biomass productivity of 0.07 g L−1 day−1 was achieved with an inlet gas flow rate of 3 Lpm. The maximum biomass efficiency of 0.103 g L−1 day−1 was achieved at an illuminance of 150 µmol m−2 s−1 in continuous light [83]. |
| Investigated the effect and optimized the nitrogen availability in combination with high light intensity on antioxidant activity and carotenoid content in Nephroselmis sp. These microalgae produce sifonaxanthin—a rare pigment for biotechnology applications. | Under the conditions of unrestricted access to nitrogen, scientists found a 3-fold increase in the content and productivity of primary carotenoids and a 2.4-fold increase in antioxidant activity. Nitrogen availability had no effect on the content of lutein and β-carotene. The experiments were carried out at a constant temperature of 26.5 ± 0.3 °C, a regulated pH level of 7.75 ± 0.04 and a light intensity of 600 µmol m−2 s−1 [89]. |
| The ability to produce lutein by thermotolerant strains of Desmodesmus sp. —the influence of substrate composition, nitrate concentration and light intensity. | The best cell growth and lutein production were achieved with a light intensity of 600 µmol m−2 s−1 and an initial nitrate concentration of 8.8 mM. The highest productivity (3.56 ± 0.10 mg L−1 d−1) and content (5.05 ± 0.20 mg g−1) of lutein were obtained in batch culture with a pulsatile feed of 2.2 mM nitrate when its content was almost exhausted [90]. |
| Research with the arctic microalga Chlamydomonas malina RCC2488 at a temperature of 8 °C to produce carbohydrates, lipids and polyunsaturated fatty acids (PUFA). This strain tolerates a wide range of salinity and high light intensity with a high content of produced lipids. | The highest biomass (527 mg L−1 d−1), lipid (161.3 mg L−1 d−1) and polyunsaturated fatty acid (PUFA; 85.4 mg L−1 d−1) productivities were obtained at a salinity of 17.5, light intensity of 250 μmol m−2 s−1 and nitrogen-replete conditions. Nitrogen deprivation induced the accumulation of carbohydrates in cells (up to 49.5% w/w) at the expense of proteins, but without compromising lipid biosynthesis [91]. |
| The effect of nutrients and light intensity on the growth and biochemical composition of the marine microalga Odontella auryta grown in columnar and flat photobioreactors. | The optimal composition of the medium for the photoautotrophic cultivation of O. aurite was found, and carbohydrates, consisting mainly of β-1,3-glucan, were found to be the main storage materials produced under stress conditions. The production of lipids and eicosapentaenoic acid (EPA) was low. The highest biomass increase was obtained at a light intensity of 300 μmol m−2 s−1) and a light path length of 3 cm. The biomass production of 3.8 g L−1 obtained in the 3 cm light path photobioreactor was 46% greater than that obtained with a 6 cm light path and 110% greater than with a 12 cm light path reactor [92]. |
| Investigated the effects of different light intensities, forms and concentrations of nitrogen, phosphorus and salinity on the growth and production of docosahexaenoic acid (DHA) in a bubble column photobioreactor (PBR). Tested 19 natural Isochrysis strains. | A record high DHA productivity was achieved, amounting to 13.4 mg·L−1·d−1. The optimal light intensity for DHA production was 60–90 µmol m−2 s−1, and the optimal phosphorus concentration was 4.5 mg L−1. Of the three light paths of 1.9, 3.8 and 7.6 cm, the 3.8 cm PBR yielded the highest volumetric biomass productivity, 0.54 ± 0.05 (g L−1 d−1) [93]. |
| Light Conditions | Achievement |
|---|---|
| Use of fiber optic spectrometer to determine the quantitative and qualitative light characteristics of the airlift photobioreactor downcomer. | The circular geometry allows for more efficient light penetration and illumination of a larger part of the PBR interior compared to the flat geometry. Limiting light availability depends on the applied wavelength, cell concentration, PBR geometry and the penetration distance of light [99]. |
| The effects of light quality and the delivery strategy on the growth and production of carotenoids by Chlorococcum humicola. | For the one- and two-stage strategy, among white, red and blue light, the highest carotenoid productivity was obtained with two-stage lighting. During the growth period, white light at an intensity of 5000 lux was supplied, followed by a combination of white (100,000 lux) and blue (5000 lux) light in the stationary phase [77]. |
| The effect of different light intensities under continuous lighting as well as alternating light and dark cycles at different frequencies. | Microalgae Nannochloropsis salina can use even very intense light efficiently, provided that dark cycles occur. If the alternation of light and dark is not optimal, the algae undergo radiation damage, and the photosynthetic efficiency drops significantly. This shows how important it is to optimize mixing in a photobioreactor to ensure efficient use of light energy by microalgae [100]. |
| Experiments with a culture of Chlamydomonas reinhardtii with high light saturation with unlimited access to CO2 in turbidostatic photobioreactors. | Significant lipid accumulation can also occur under maximum growth conditions, without slowing down nitrogen starvation as a stress factor [101]. |
| The effects of cultivating microalgae under fluorescent light and LED diodes in six different colors: blue, purple, orange, white, green and red. | Monochromatic blue light can substitute for polychromatic fluorescent light, and the colors generating the greatest amount of biomass are blue and red, as they are absorbed by chlorophyll a and b contained in Dunaliella tertiolecta cells [73]. |
| Research on the growth rate, biomass yield and biosynthesis of carotenoids in the microalgae Dunaliella salina using red LEDs with a wavelength of 660 nm and blue LEDs with a wavelength of 470 nm and a narrow output spectrum (20 nm band at half the peak height). | The results showed that increasing the red light intensity to 170 μE/m2/s did not increase the accumulation of carotenoids. On the other hand, combining a red LED (75%) with a blue LED (25%) made it possible to increase carotenoids with the total photon flux. Additional blue light instead of red led to increased accumulation of β-carotene and lutein [102]. |
| Three typical lighting systems were compared. | The influence of white light, blue-red LED light and blue-red LED light with far-red light on metabolic activity, chemical composition and yield of Chlorella vulgaris microalgae was analyzed. Classic fluorescent lamps turned out to be a much better source of light for the cultivation of C. vulgaris than LED sources emitting narrow bands of radiation in the blue, red and far-red ranges. Fluorescent light most effectively stimulated biomass growth and had the least impact on the physiological conditions of the culture. The fluorescent lighting was located between the reactor and the external coil, and the LED lighting only shone on the coil that covered the reactor [103]. |
| Cultivation under blue, red or white light provided by light-emitting diodes and white light provided by fluorescent lamps to assess the growth characteristics of Dunaliella sp. in the green phase. | It was confirmed that for a batch culture run in PBRs with different LED lights, red and blue dichromatic lighting was more appropriate than monochrome, red or blue light. A stage one light delivery strategy was proposed for faster cell growth [104]. |
| They used natural sunlight and wrapped the photobioreactors with filters of red, blue and red + blue light in the culture of Tetraselmis sp. in a bubble column photobioreactor outside. | The highest biomass and fatty acid productivity were obtained with a red light filter. Increased biodiesel production from algae can be achieved without artificial light sources, thanks to appropriate light filters transmitting selected wavelengths from solar radiation [105]. |
| Maximizing the use of light and thus reducing the production costs of microalgae biomass. | They positioned mirrors around a column photobioreactor with internal lighting to increase the light intensity by reflecting the outgoing light rays back into the photobioreactor. The mirrors increased the light intensity by about 1.7 times without any additional energy consumption, and the specific growth rate and dry weight of C. vulgaris microalgae cells increased by about 25% and 91%, respectively, compared to the reference culture [68]. |
| Investigated the influence of different colors of light-emitting diodes on the biomass composition of Arthrospira platensis (Spirulina) microalgae. | Different enzymatic steps are required to synthesize carbohydrates, lipids or proteins. Each of these stages uses a certain amount of energy. Hence, some colors that do not provide a sufficient number of photons (energy) allow for efficient synthesis of only less energy-consuming compounds [106]. |
| Investigated high-density cultures. | Red light, which is heavily absorbed, is not necessarily the best wavelength for growing algae with monochrome lighting. Green light, which is less absorbed and can be scattered over a larger area of the photobioreactor, will be a better solution [107]. |
| Experimenting with linear and exponential light intensity strategies in microalgae culture found that in the photoautotrophic mode, the light energy required by microalgae varies with the growth of the biomass. | In the early stages of growth, less light is needed to penetrate well through the algae culture, while moderately dense cultures require higher light intensities to counteract cell shading and enhance photosynthesis [108]. |
| Description of the Reactor | Achievement |
|---|---|
| Transparent and gas-permeable micro-reactors made of Poly(dimethylsiloxane), 3.5 mm in height. | The system provides high-throughput screening for growth and analysis of useful products such as lipids, carotenoids and polymers [71]. |
| Small prototype reactors, acrylic tubes with a diameter of 1″, filled to 2 or 6 cm, which gave a total volume of 30 to 50 mL. | Verification of the possibility of exploiting mixotrophy (organic substrates as carbon source) in combination with excess CO2 in cultivation of Chlorella protothecoides and Nannochloropsis salina [107]. |
| Multichannel photobioreactor made of 24 stands of 0.1 L bottles. | Pigment production from heterotrophically cultivated Chlorella sp. HS2 was optimized by regulating a suite of environmental conditions using the mcPBR system [125]. |
| Laboratory scale—volume of 0.25 L to 0.5 L. | Impact of environmental conditions on biomass productivity and carbon dioxide fixation [126]. |
| Photobioreactor with 350 mL cylindrical glass columns (ID = 4 cm), each with a 250 mL working volume. | Identified the optimum conditions and combined nutrient abatement from wastewater with CO2 fixation from flue gases [127]. |
| Laboratory scale—volume of 0.25 L. | Characterization of secondary carotenoid production from Dactylococcus dissociatus MT1 isolated from the Sahara Desert of Algeria [128]. |
| Laboratory scale—volume of 0.5 L. | High-dose CO2 significantly enhanced the light energy conversion and storage into lipids in the cultivation of Chlorella sorokiniana CS-1 [129]. |
| Laboratory scale—volume of 0.5 L. | The effect of light quality on cell size and cell cycle, growth rate, productivity, PE and biomass composition of Tetraselmis suecica F&M-M33 [130]. |
| Laboratory scale—volume of 0.25 L to 0.5 L. | Techno-economic feasibility of the cultivation process was based on measurements of long-term, sustained production at a demonstration scale [131]. |
| Poly (ethylene terephthalate) cylindrical photobioreactors with an inner diameter of 80 mm and total volume of the PBR of 500 mL. | The effect of various colors of LED on the content of the main pigments of Arthrospira platensis, such as phycocyanin, chlorophyll and total carotenoids, and the contents of the proteins, carbohydrates and lipids in semi-continuous cultures [106]. |
| Twin-layer biofilm photobioreactor with a volume of 800 mL. | The effect of light intensities and different concentrations of additional CO2 on biomass productivity and total biofilm dry weight of the green algae Halochlorella rubescens [132]. |
| Bubble column photobioreactor with a volume of 1 L. | Mild-pressure-induced physical stress to promote rapid TAG accumulation in microalgae [133]. |
| Column-type glass-fabricated PBRs (ϕ 6 cm × 80 cm high) with 1 L of working volume. | Growth and CO2 utilization efficiency of the Chlorella sp. AT1 with intermittent CO2 aeration in a semi-continuous long-term culture [134]. |
| Bubble column photobioreactor with a volume of 2 L. | Flashing light as a light source in microalgae cultivation [135,136]. |
| 2 L flat-panel photobioreactors. | The effect of light quality on cell size and cell cycle, growth rate, productivity, PE and biomass composition of Tetraselmis suecica F&M-M33, with the objective of improving biomass quality and value for aquaculture and decrease production costs [137]. |
| 2 L airlift PBR. | Development of an optimal light-feeding strategy coupled with semi-continuous mode of reactor operation that resulted in greater lutein productivity, photosynthetic efficiency and CO2 fixation rate of Chlorella minutissima [108]. |
| Reactors with a capacity of 3 L. | Optimization of a suite of environmental conditions for pigment production from heterotrophically cultivated Chlorella sp. HS2 [125]. |
| Bubble column photobioreactors in batch operation mode 3 L. | Study of three key factors influencing cell growth of isolated microalga Scenedesmus obtusiusculus: CO2, aeration and light intensity and the accumulation of lipids assessed under nitrogen limitation [138]. |
| Air-lift-type photobioreactor with a porous centric tube with a volume of 4 L and bubble column and centric-tube photobioreactors. | Comparison of culturing microalgae at high density [139]. |
| Vertical tubular photobioreactors with a working volume of 4 L. | Research on the optimization of the Parietochloris incisa substrate for the accumulation of arachidonic acid [140]. |
| Bubble column photobioreactor with a volume of 4.3 L. | Study of the medium’s composition in microalgae cultures of Chlorella sp. and Monodus subterraneus [116]. |
| Airlift photobioreactor having a working volume of 5 L. | Cultivated a mixed culture of microalgae in an airlift photobioreactor in batch mode and studied the effect of varying CO2 concentrations and the combined CO2 and air flow rate, changes in pH with time at different CO2 concentrations and the effect of CO2 concentration on CO2 the biofixation rate [141]. |
| Reactors with a capacity of 7 L. | Evaluation of the recycling of low-cost crude glycerol as a feedstock for heterotrophic cultivation of microalgae [142]. |
| Prototype photobioreactors of various designs—volumes of 2.5, 5 and 20 L. | Effect of photobioreactor scale on cell growth and carbon dioxide fixation by Chlorella sorokiniana [143]. |
| Double-walled polyethylene 20 L bags. | Control the tolerance of the unicellular green microalga Chlorella minutissima under extreme carbon dioxide concentrations, natural temperature and high-light conditions [144]. |
| A 35 L bubble column photobioreactor | Experiments aimed at increasing the production of biomass and lipids by two autochthonous dinoflagellates (Alexandrium minutumand and Karlodinium veneficum) and one raphidophyte (Heterosigma akashiwo) [145]. |
| A 50 L reactor cooperating with a 60 L open pool. | Developing a novel culture model for algal biomass and lipid production, namely, sequential heterotrophy–dilution–photoinduction [146]. |
| Tanks with a volume of 60 L. | The tolerance to shear stress and the biochemical characteristics of the Phaeodactylum tricornutum microalgae in a column aerated photobioreactor and two versions of airlift photobioreactors with a split-cylinder airlift device and a concentric draft-tube airlift vessel [65]. |
| Flat photobioreactors with a volume of 60 L. | The level of lipid accumulation and the growth characteristics of Chlorella zofingiensis microalgae were checked at different concentrations of nitrates and phosphates [147,148]. |
| Pilot photobioreactors with a volume of 100 L. | Identification of the potential application of the biomass derived from microalgae cultivation with the use of industrial flue gas in the animal feed industry [149]. |
| Ultrafiltration membrane fouling in a collection of algae from a 120 L tank was investigated. | Characterization of algal-related organic matter and evaluation of the impacts on ultrafiltration membrane fouling [150]. |
| Photobioreactors with a volume of 150 L. | Pilot-scale outdoor cultivation of newly isolated strain Monoraphidium sp. CCALA 1094 with low requirements for cultivation temperature and a minimal requirement for light [151]. |
| Pilot photobioreactors with a volume of 250 L. | The use of the waste CO2 recovered from a biogas upgrading process, in place of synthetic commercial CO2, in the cultivation of microalgae [152]. |
| Photobioreactors with a much larger volume of 320 L or 540 L. | Culture performance of the dinoflagellate Amphidinium carterae in controlled conditions, in a closed system (PBR), in a large scale (>500 L), long-term culture (>8 months) grown in semi-continuous mode [153]. |
| A pilot-scale photobioreactor, an open pond and a hybrid two-stage system. | The cultivation of microalgae for lipid targets [85]. |
| The tubular photobioreactor was 1200 L, and the open raceway pond had a volume of 1000 L. | Comparison of microalgae cultivation systems, including open pond, closed PBR and hybrid cultivation [85]. |
| Pilot photobioreactors with a volume of 1136 L. | Possibility of using waste heat from a combustion power plant to heat algae cultures in colder climates [154]. |
| Type of the Reactor | Description |
|---|---|
| Classic mechanical stirring reactors | The use of mechanical agitation photobioreactors to cultivate microalgae is not the best choice. High shear rates around the impellers of the agitators damage the cells of microorganisms, which are characterized by poor mechanical strength [162,174,175]. |
| Various diameters of bubble columns | Higher volume-averaged light intensity, lower shear stress, higher astaxanthin content and higher mass concentration [168]. Higher biomass concentration and a longer light path will increase the extent of light attenuation [176]. |
| Column aeration photobioreactors | The substrate circulates therein, rising in the central area and falling in the area close to the wall. In this way, the cells are ensured cycles of high light intensity, being close to the wall and lower illumination inside the column. At the same time, the absorption of carbon dioxide, which is a carbon source in photoautotrophic cultures, and the removal of photosynthetically produced oxygen, which inhibits photosynthesis, take place [171,177,178]. |
| Classic airlift reactors | A very good solution for conducting photoautotrophic cultures [161,179,180,181,182]. |
| Raceway pond with a gutter and oscillating partitions | It uses movable baffles to oscillate in the flowing substrate and fixed prismatic baffles to generate a vortex, which were developed to enhance the mixing efficiency of H. pluvialis cultures. The results showed that applying the new solutions could improve the mass transfer coefficient and shorten the mixing time, as well as the growth rate of microalgae, in the raceway pond [183]. |
| Two-layer, slightly inclined (15°) photobioreactor with a porous substrate | Used to conduct immobilized microalgae cultures to obtain astaxanthin. The advantage of this solution is its simple structure, easy operation and technically simple harvesting of biomass, which translates into saving water, energy and culture time [160]. |
| Horizontal, floating photobioreactor | Without aeration or a mixing device [165]. |
| Reactor Type | Achievement |
|---|---|
| V-shaped bottom slope photobioreactor, air volume flow (vvm), reactor height/diameter ratio and air bubble diameter. | They optimized the main factors related to substrate mixing in an aerated column reactor [173]. |
| Tested the influence of the shape of the photobioreactor (flat panel, horizontal tubular and vertical tubular) on the efficiency of microalgae cultivation. | They tested various sparger shapes (ball, cube and cylinder) made of various materials (glass, plastic, steel and rubber) to determine mixing efficiency. Additionally, they investigated the effect of the light path length on the biomass efficiency by changing the photobioreactor diameter (5, 10, 15 and 20 cm) [164]. |
| Effect of split-cylinder. | They used a specially placed partition forming an inner loop inside the cylinder [188]. |
| To increase the illumination efficiency in split-column photobioreactors. | They used a photobioreactor, which was built of two interconnected bubble columns of different sizes with continuous circulation of the culture medium from one column to the other. Light was delivered to the smaller column only, allowing astaxanthin to be induced more efficiently by improving light distribution [189]. |
| Comparison of bubble column photobioreactors with airlift. | The results clearly spoke in favor of the airlift. Higher cell density was obtained in the growth phase. Regular light/dark cycles and downstream laminar flow positively affected astaxanthin accumulation, increasing it by 16% compared to the aerated column [187]. |
| Material | Ref. |
|---|---|
| Glass laboratory vessels and photobioreactors made of glass tubes. | [185,190,191] |
| Transparent plastics, such as a twin-layer photobioreactor made of a PMMA acrylic tube set on a polyvinyl chloride (PVC) base. | [192] |
| Plexiglass—a vertical panel-type 1.5 L photobioreactor. | [193] |
| Polypropylene film, which is transparent and has a very high durability (6 L column photobioreactors). | [174] |
| Polyethylene—angled twin-layer porous substrate photobioreactor (10 L bags). | [160] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borowiak, D.; Krzywonos, M. Bioenergy, Biofuels, Lipids and Pigments—Research Trends in the Use of Microalgae Grown in Photobioreactors. Energies 2022, 15, 5357. https://doi.org/10.3390/en15155357
Borowiak D, Krzywonos M. Bioenergy, Biofuels, Lipids and Pigments—Research Trends in the Use of Microalgae Grown in Photobioreactors. Energies. 2022; 15(15):5357. https://doi.org/10.3390/en15155357
Chicago/Turabian StyleBorowiak, Daniel, and Małgorzata Krzywonos. 2022. "Bioenergy, Biofuels, Lipids and Pigments—Research Trends in the Use of Microalgae Grown in Photobioreactors" Energies 15, no. 15: 5357. https://doi.org/10.3390/en15155357
APA StyleBorowiak, D., & Krzywonos, M. (2022). Bioenergy, Biofuels, Lipids and Pigments—Research Trends in the Use of Microalgae Grown in Photobioreactors. Energies, 15(15), 5357. https://doi.org/10.3390/en15155357