Sargassum Inundations in Turks and Caicos: Methane Potential and Proximate, Ultimate, Lipid, Amino Acid, Metal and Metalloid Analyses




Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation
2.1.1. Sample Collection
2.1.2. Freeze-Drying
2.2. Compositional Analyses
2.2.1. Moisture
2.2.2. Ash
2.2.3. Salt
2.2.4. Carbon, Hydrogen, Nitrogen Sulphur (CHNS)
2.2.5. ‘Heavy Metals’
2.2.6. Phenolic Content
2.2.7. Amino Acids
2.2.8. Fatty Acids
2.2.9. Total Lipid
2.2.10. Fibre
2.2.11. Higher Heating Value
2.3. Methane Potential
2.3.1. Theoretical Methane Potential
2.3.2. Methane Potential Determination
- (a)
- Fresh Mixed Sample (A)
- (b)
- Freeze-Dried Mixed Sample (A)
- (c)
- S. natans VIII (B)
- (d)
- S. natans I (C)
- (e)
- S. fluitans (D)
- (f)
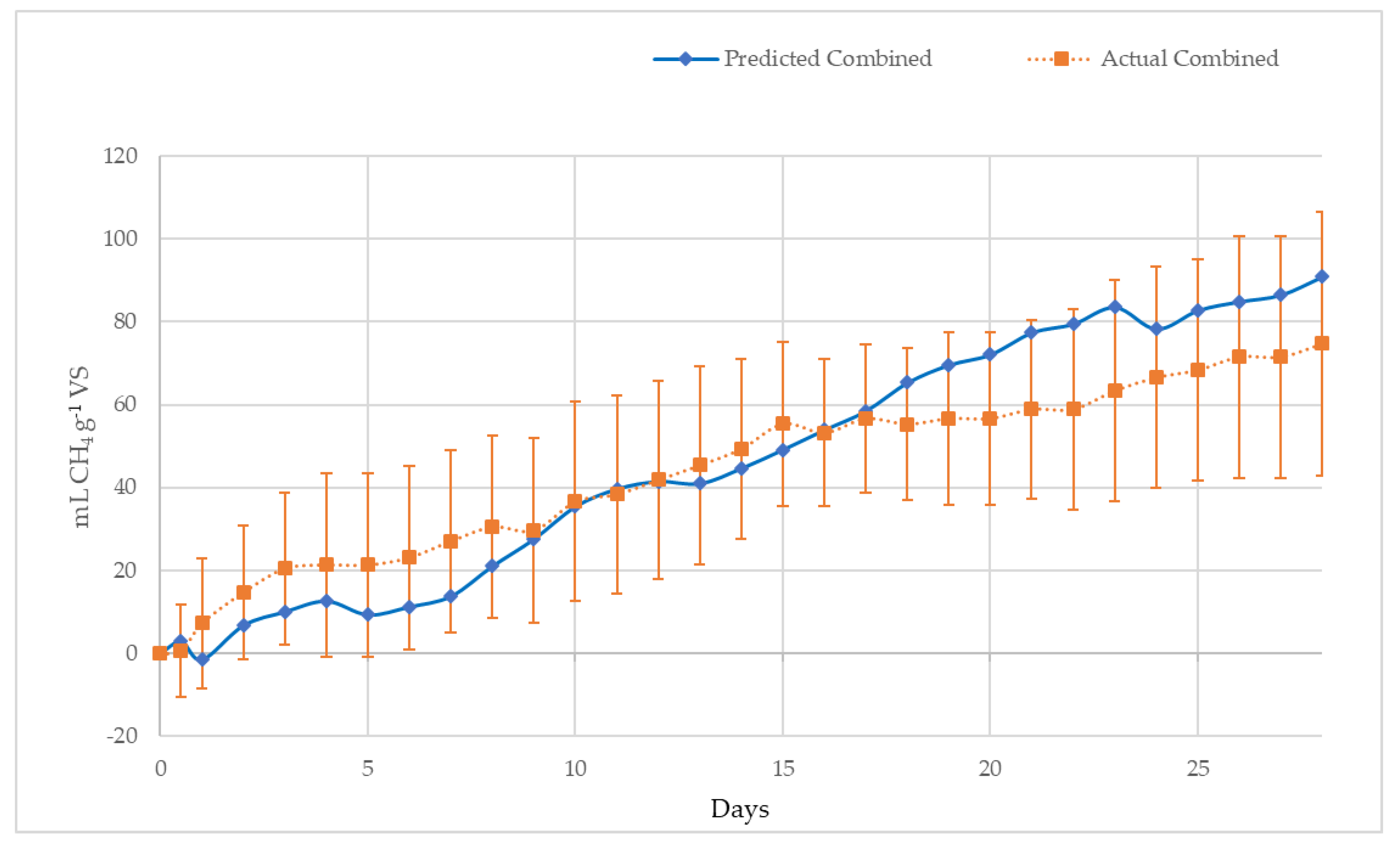
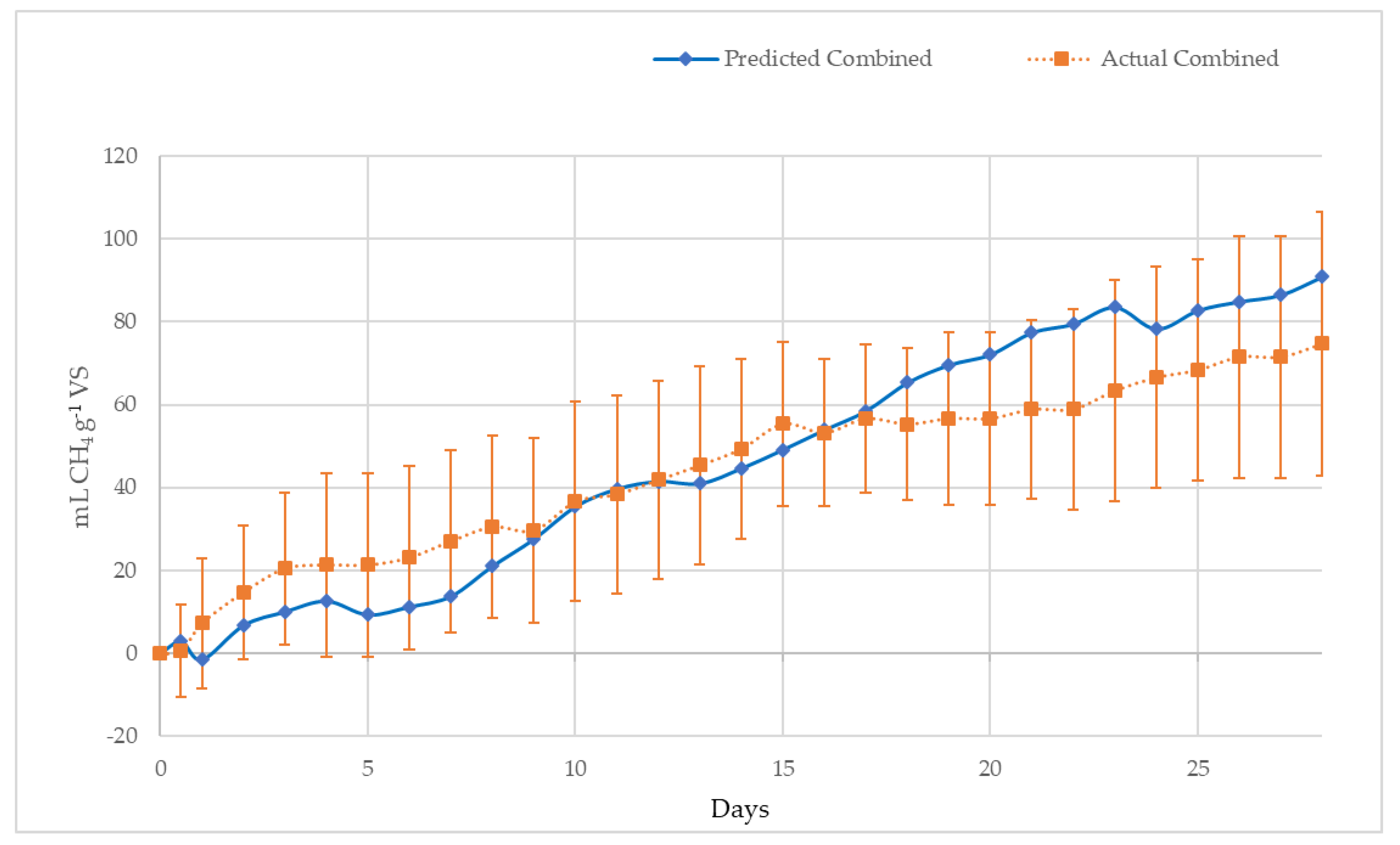
- A mixture based on the volatile solids (VS) percentage of 1.0% S. natans VIII (B), 49.3% S. natans I (C) and 49.7% S. fluitans (D), the ratio of the three pelagic Sargassum species found previously in Shark Bay [61]. The ratio was selected to examine the potential synergistic and antagonistic effects of a mixture of species on methane yield. However, the composition of floating Sargassum mats and beach inundations can vary widely [62].
2.4. Statistical Analysis
3. Results
3.1. Compositional Analysis
3.1.1. Moisture, Ash, Total Solids and Volatile Solids
3.1.2. Salt and XRD analysis
3.1.3. CHNS
3.1.4. ‘Heavy Metals’
3.1.5. Phenols
3.1.6. Amino Acids
3.1.7. Fatty Acids
3.1.8. Total lipid and Fibre
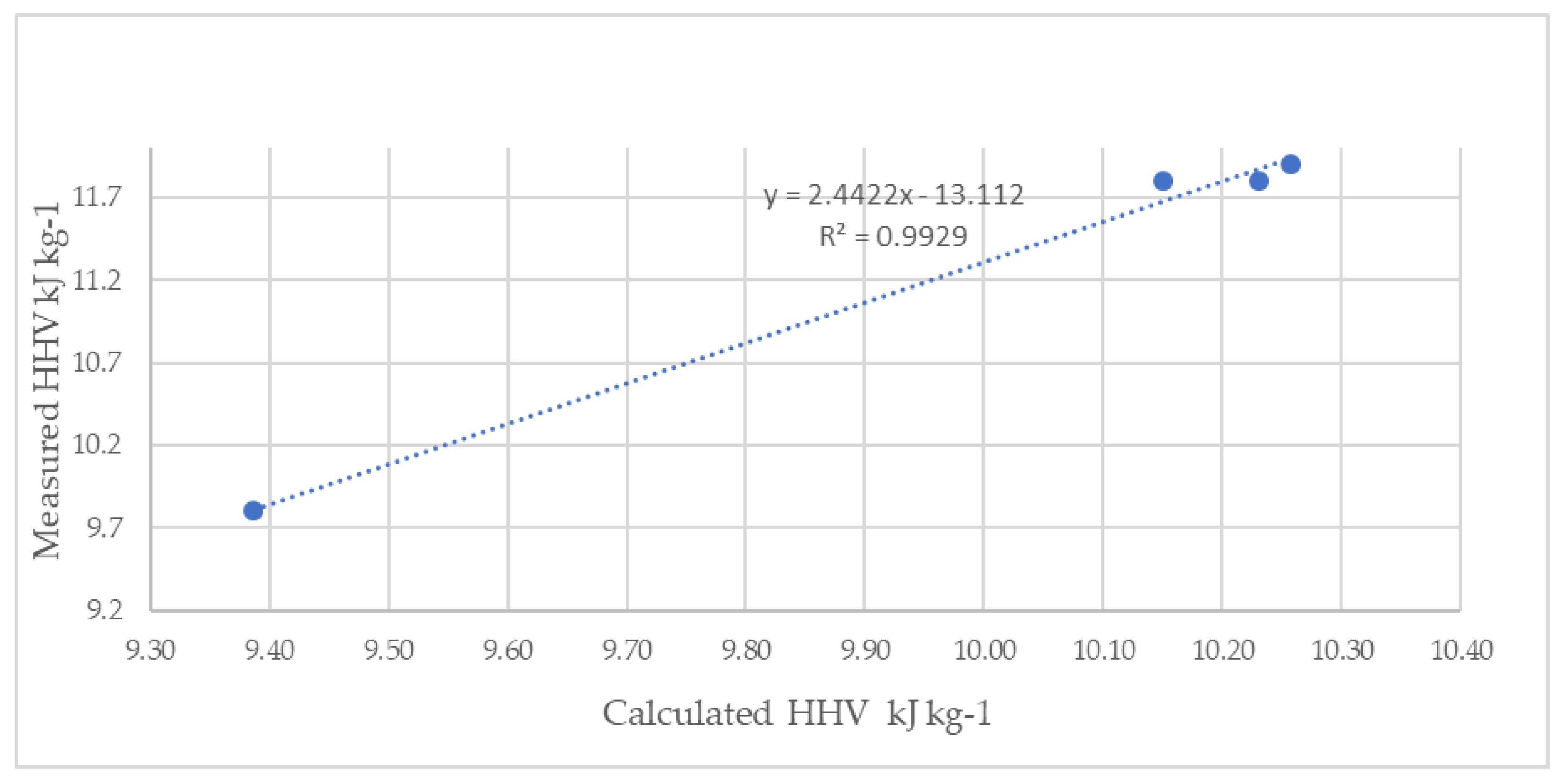
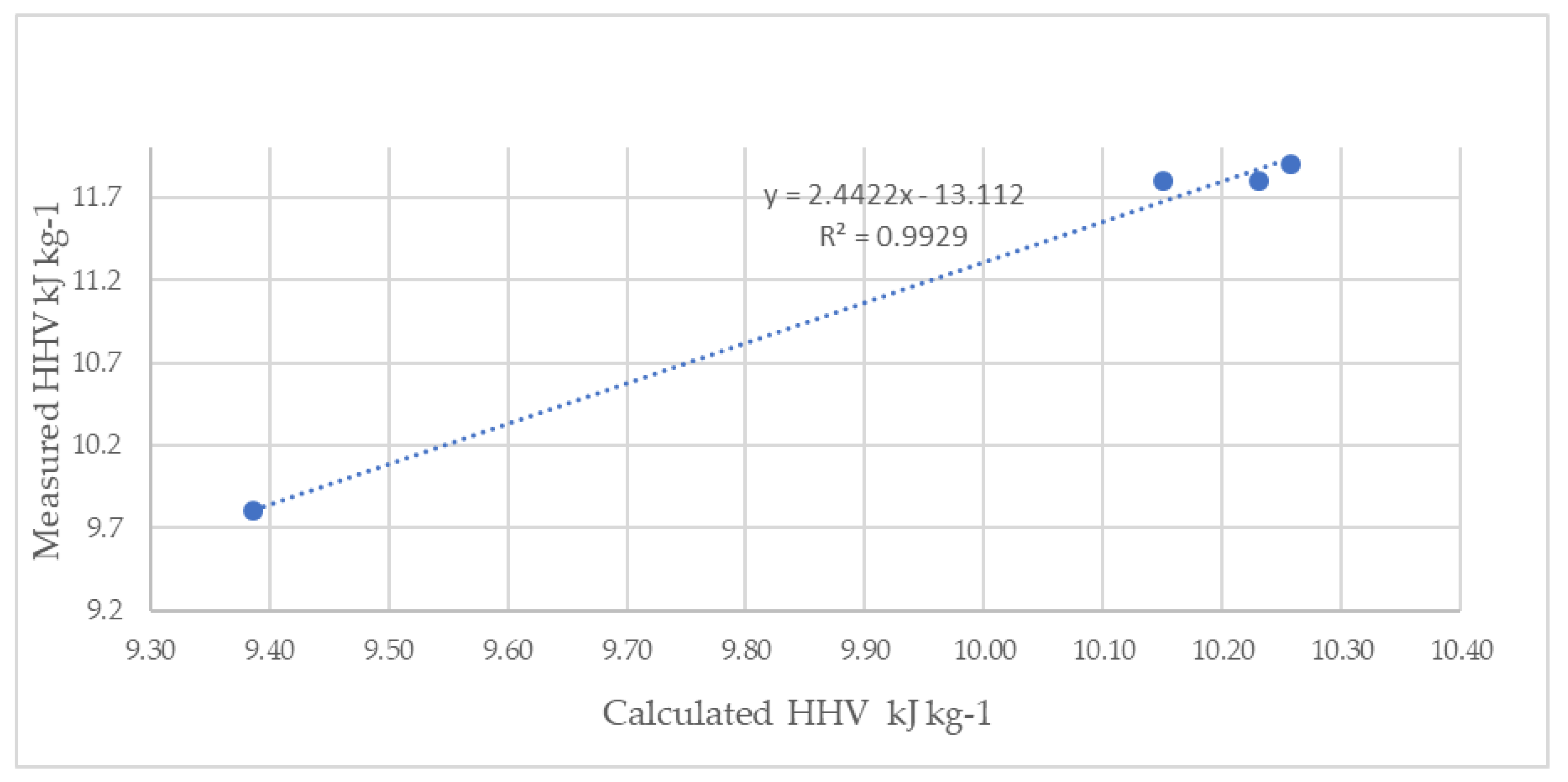
3.1.9. Higher Heating Value
3.2. Methane Potential
3.2.1. Theoretical Methane Potential of S. Natans VIII, S. Natans I and S. Fluitans
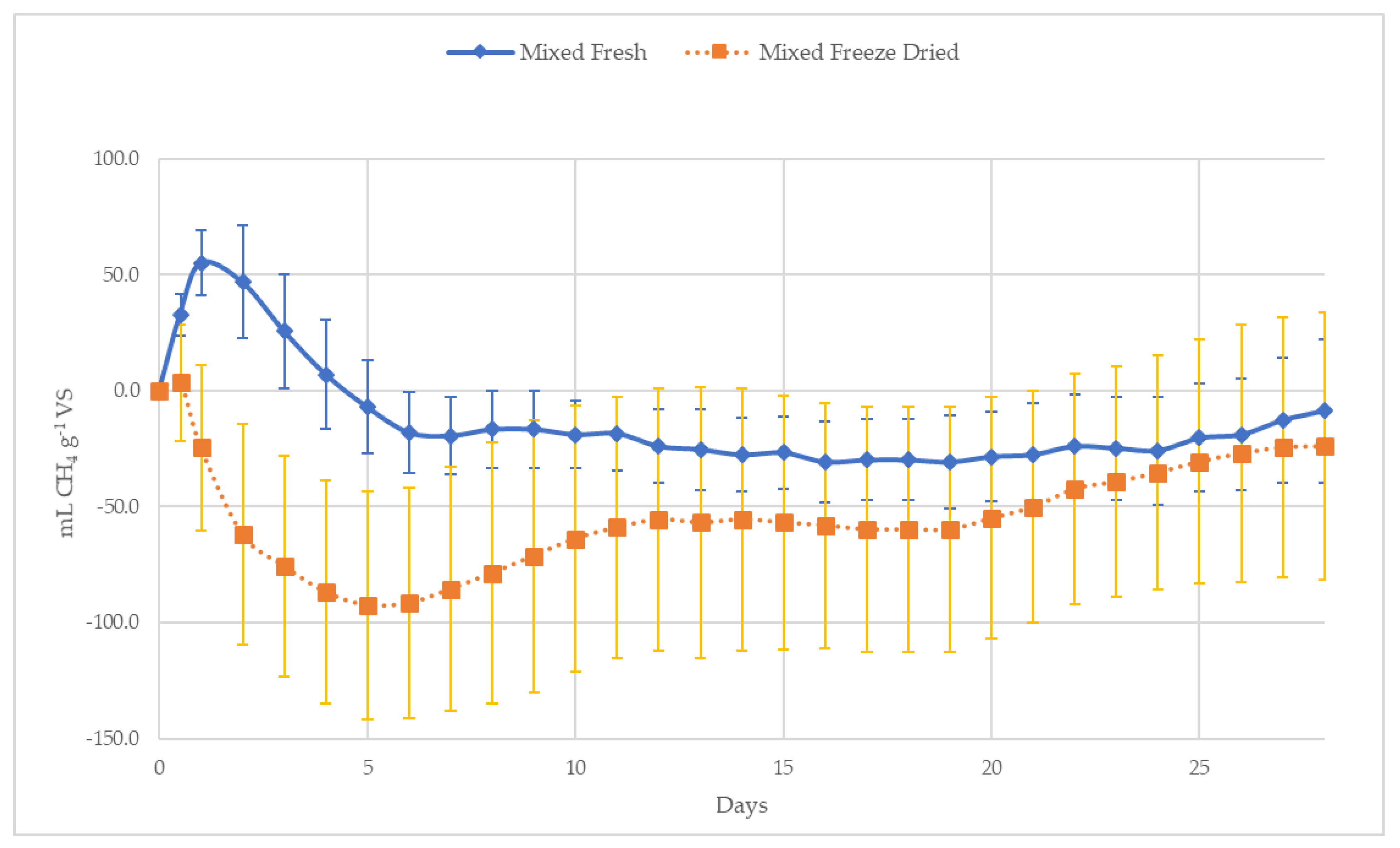
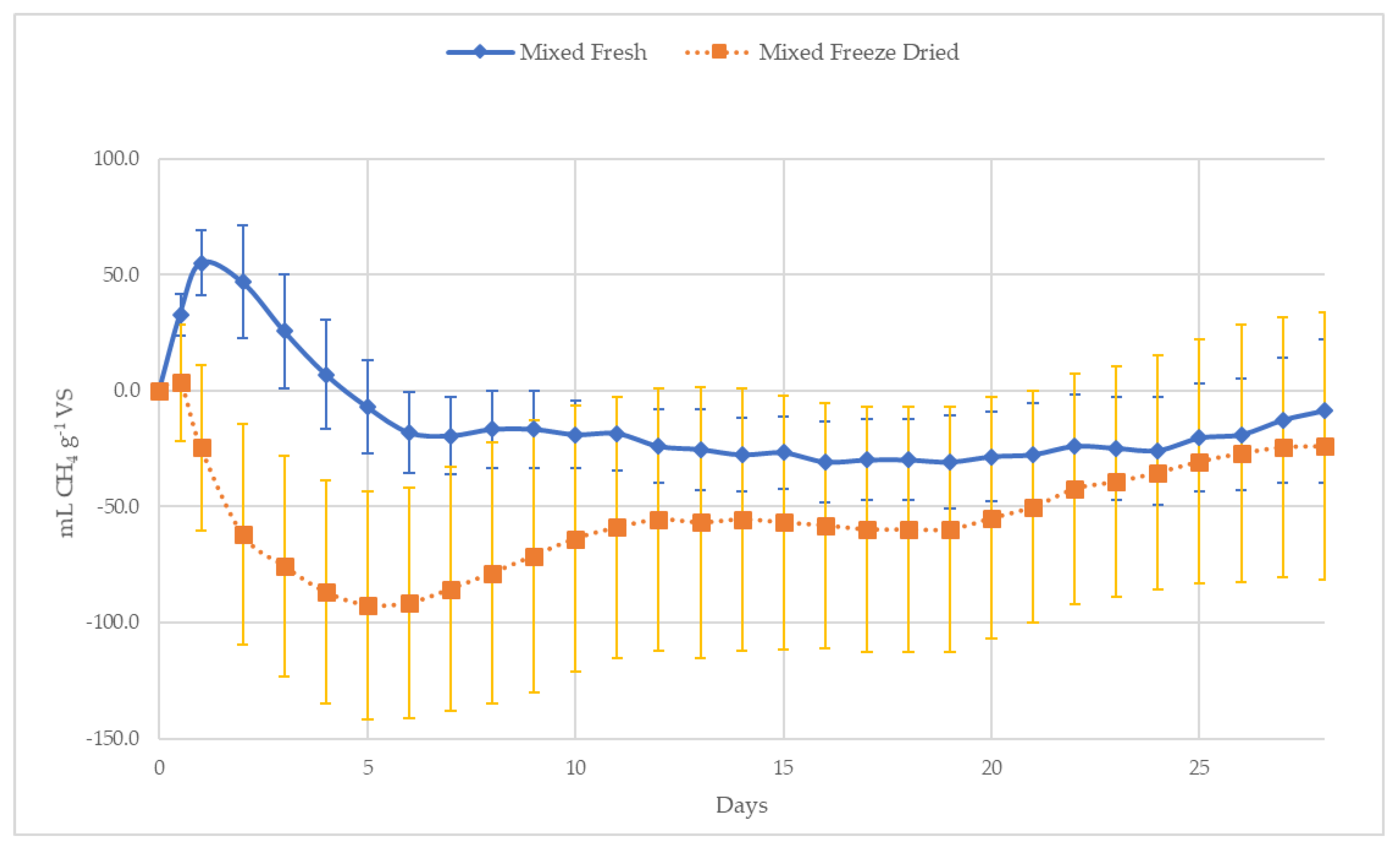
3.2.2. Fresh versus Freeze-Dried
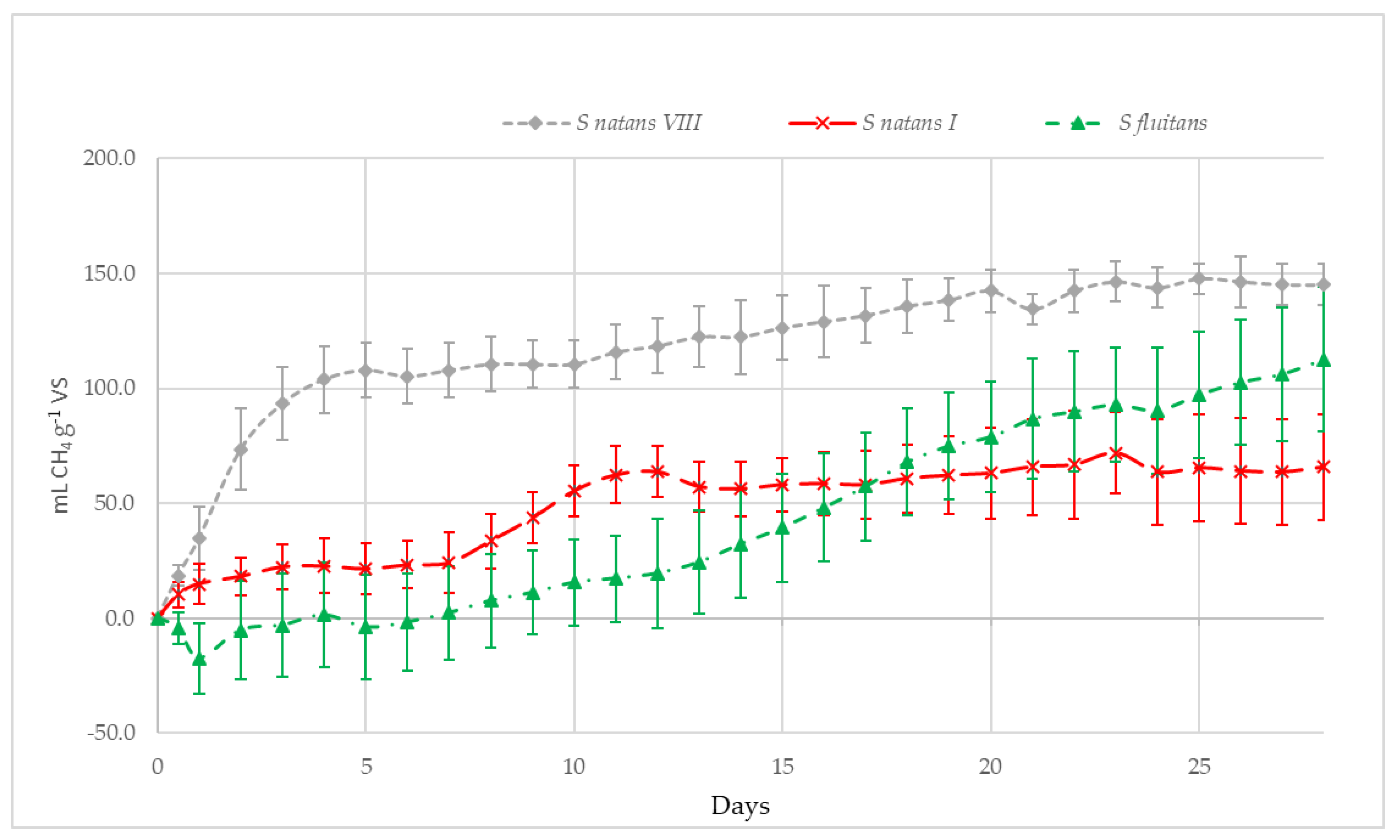
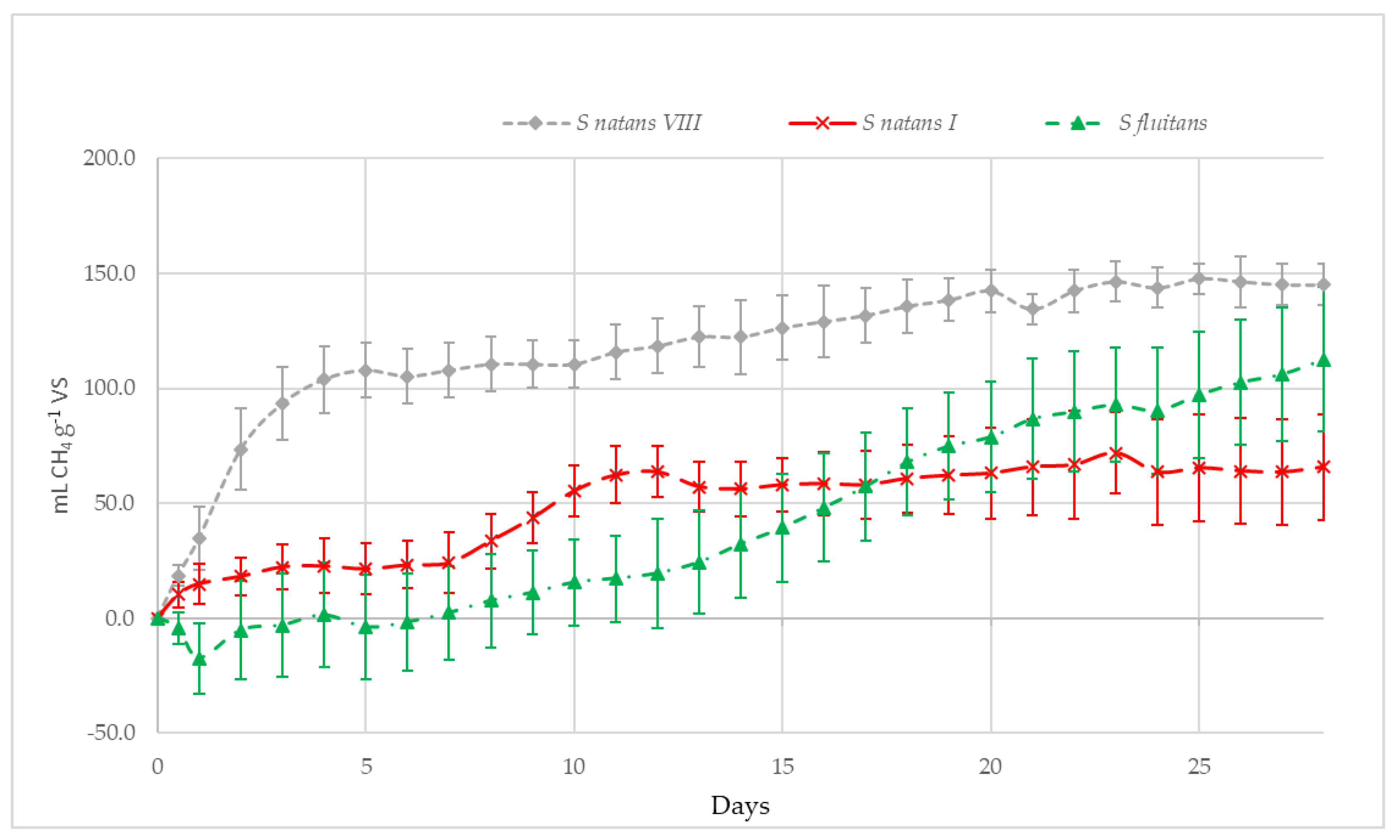
3.2.3. Methane Potential of S. Natans VIII, S. Natans I and S. Fluitans
3.2.4. Methane Potential of S. Natans VIII, S. Natans I and S. Fluitans
4. Discussion
4.1. Composition of Sargassum
4.1.1. Minerals and Metals
4.1.2. Phenols
4.1.3. Sulphur and Carbon
4.1.4. Protein and Amino Acids
4.1.5. Fibre
4.1.6. Lipid and Fatty Acids
4.1.7. Higher Heating Value
4.2. Methane Potential
4.2.1. Fresh Versus Freeze Dried
4.2.2. Methane Potential of S. Natans VIII, S. Natans I, S. Fluitans and ‘Mixed Sargassum’ mats
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smetacek, V.; Zingone, A. Green and golden seaweed tides on the rise. Nature 2013, 504, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Milledge, J.J.; Harvey, P. Golden tides: Problem or golden opportunity? The valorisation of sargassum from beach inundations. J. Mar. Sci. Eng. 2016, 4, 60. [Google Scholar] [CrossRef]
- Van Tussenbroek, B.I.; Hernandez Arana, H.A.; Rodriguez-Martinez, R.E.; Espinoza-Avalos, J.; Canizales-Flores, H.M.; Gonzalez-Godoy, C.E.; Barba-Santos, M.G.; Vega-Zepeda, A.; Collado-Vides, L. Severe impacts of brown tides caused by Sargassum spp. on near-shore caribbean seagrass communities. Mar. Pollut. Bull. 2017, 122, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Burrowes, R.; Wabnitz, C.; Eyzaguirre, J. The Great Sargassum Disaster of 2018. Available online: https://essa.com/the-great-sargassum-disaster-of-2018/ (accessed on 25 March 2019).
- Langin, K. Seaweed masses assault caribbean islands. Science 2018, 360, 1157–1158. [Google Scholar] [CrossRef] [PubMed]
- Thompson, T.M.; Young, B.R.; Baroutian, S. Pelagic sargassum for energy and fertiliser production in the caribbean: A case study on barbados. Renew. Sustain. Energy Rev. 2020, 118, 109564. [Google Scholar] [CrossRef]
- Maurer, A.S.; De Neef, E.; Stapleton, S. Sargassum accumulation may spell trouble for nesting sea turtles. Front. Ecol. Environ. 2015, 13, 394–395. [Google Scholar] [CrossRef] [Green Version]
- Resiere, D.; Valentino, R.; Nevière, R.; Banydeen, R.; Gueye, P.; Florentin, J.; Cabié, A.; Lebrun, T.; Mégarbane, B.; Guerrier, G.; et al. Sargassum seaweed on Caribbean islands: An international public health concern. Lancet 2018, 392, 2691. [Google Scholar] [CrossRef] [Green Version]
- Willoughby, S. Sargassum and the Fishing Industry. In Proceedings of the Sargassum Symposium, UWI, Cave Hill, London, UK, 17 August 2015. [Google Scholar]
- World Travel & Tourism Council. Caribbean Resiliencea and Recovery: Minimising the Impact of the 2017 Hurricane Season on the Caribbean’s Tourism Sector; WTTC: London, UK, 2017. [Google Scholar]
- Anon. Caribbean-Bound Tourists Cancel Holidays Due to Foul-Smelling Seaweed. The Guardian. 10 August 2015. Available online: https://www.leonetwork.org/en/posts/show/7BE7F4EC-F482-4925-BD7D-1252030F54F3 (accessed on 4 March 2016).
- Jessop, D. Sargassum a Continuing Challenge for Tourism. In The Caribbean Council Research & Analysis; The Caribbean Council: London, UK, 2019; Volume 22. [Google Scholar]
- Tourism Down 30–35% Due to Sargassum: Playa Del Carmen Mayor-Elect. Mexico News Daily. 4 August 2018. Available online: http://sargassummonitoring.com/author/denji/page/2/ (accessed on 11 December 2019).
- Beckles, H. Greetings from the vice-chancellor. In Proceedings of the Sargassum Symposium, UWI, Cave Hill, London, UK, 17 August 2015. [Google Scholar]
- Khan, A. Region Needs US$120 m to Fight Seaweed. Trinidad Daily Express. 18 August 2015. Available online: https://mexiconewsdaily.com/news/sargassum-a-potential-disaster-for-beaches/ (accessed on 3 March 2016).
- Sargassum a Potential Natural Disaster for Caribbean beaches: Researchers. Mexico News Daily. 3 August 2018. Available online: https://www.google.com.hk/url?sa=t&rct=j&q=&esrc=s&source=web&cd=12&ved=2ahUKEwiO4_K6up7oAhWaZt4KHe7fDhQQFjALegQIAhAB&url=https%3A%2F%2Fwww.soundingsonline.com%2Ffeatures%2Fsargassum-blooms-grow-monstrous-and-researchers-say-more-will-come&usg=AOvVaw3nYzoFw6ZtI2oRYwtPOU5s (accessed on 11 December 2019).
- De Schaun, K. The Summer of seaweed. In Proceedings of the Sustainability Webinar Series, Online, 21 July 2015. [Google Scholar]
- Caribbean Sea Commission. In Proceedings of the Challenges, Dialogue and Cooperation towards the Sustainability of the Caribbean Sea, Port of Spain, Spain, 23–24 November 2015.
- Sembera, J.A.; Meier, E.J.; Waliczek, T.M. Composting as an alternative management strategy for sargassum drifts on coastlines. HortTechnology 2018, 28, 80–84. [Google Scholar] [CrossRef]
- Laffoley, D.D.A.; Roe, H.S.J.; Angel, M.V.; Ardron, J.; Bates, N.R.; Boyd, L.L.; Brooke, S.; Buck, K.N.; Carlson, C.A.; Causey, B.; et al. The Protection And Management of The Sargasso Sea: The Golden Floating Rainforest of The Atlantic Ocean: Summary Science And Supporting Evidence Case; Sargasso Sea Alliance: Bermuda, UK, 2011. [Google Scholar]
- Murphy, F.; Devlin, G.; Deverell, R.; McDonnell, K. Biofuel production in ireland—An approach to 2020 targets with a focus on algal biomass. Energies 2013, 6, 6391–6412. [Google Scholar] [CrossRef]
- Milledge, J.J.; Nielsen, B.V.; Maneein, S.; Harvey, P.J. A brief review of anaerobic digestion of algae for bioenergy. Energies 2019, 12, 1166. [Google Scholar] [CrossRef] [Green Version]
- Milledge, J.J.; Staple, A.; Harvey, P. Slow pyrolysis as a method for the destruction of japanese wireweed, sargassum muticum. Environ. Nat. Resour. Res. 2015, 5, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Milledge, J.J.; Smith, B.; Dyer, P.; Harvey, P. Macroalgae-derived biofuel: A review of methods of energy extraction from seaweed biomass. Energies 2014, 7, 7194–7222. [Google Scholar] [CrossRef]
- Barbot, Y.; Al-Ghaili, H.; Benz, R. A review on the valorization of macroalgal wastes for biomethane production. Mar. Drugs 2016, 14, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aresta, M.; Dibenedetto, A.; Barberio, G. Utilization of macro-algae for enhanced CO2 fixation and biofuels production: Development of a computing software for an LCA study. Fuel Process. Technol. 2005, 86, 1679–1693. [Google Scholar] [CrossRef]
- Sutherland, A.; Varela, J. Comparison of various microbial inocula for the efficient anaerobic digestion of Laminaria hyperborea. BMC Biotechnol. 2014, 14, 7. [Google Scholar] [CrossRef]
- Allen, E.; Wall, D.M.; Herrmann, C.; Xia, A.; Murphy, J.D. What is the gross energy yield of third generation gaseous biofuel sourced from seaweed? Energy 2015, 81, 352–360. [Google Scholar] [CrossRef]
- Huesemann, M.; Roesjadi, G.; Benemann, J.; Metting, F.B. Biofuels from microalgae and seaweeds. In Biomass to Biofuels; Blackwell Publishing Ltd.: Oxford, UK, 2010; pp. 165–184. [Google Scholar] [CrossRef]
- Jard, G.; Marfaing, H.; Carrere, H.; Delgenes, J.P.; Steyer, J.P.; Dumas, C. French Brittany macroalgae screening: Composition and methane potential for potential alternative sources of energy and products. Bioresour. Technol. 2013, 144, 492–498. [Google Scholar] [CrossRef]
- Soto, M.; Vazquez, M.A.; de Vega, A.; Vilarino, J.M.; Fernandez, G.; de Vicente, M.E. Methane potential and anaerobic treatment feasibility of Sargassum muticum. Bioresour. Technol. 2015, 189, 53–61. [Google Scholar] [CrossRef]
- Milledge, J.J.; Harvey, P.J. Ensilage and anaerobic digestion of Sargassum muticum. J. Appl. Phycol. 2016, 28, 3021–3030. [Google Scholar] [CrossRef]
- Milledge, J.; Nielsen, B.; Sadek, M.; Harvey, P. Effect of freshwater washing pretreatment on Sargassum muticum as a feedstock for biogas production. Energies 2018, 11, 1771. [Google Scholar] [CrossRef] [Green Version]
- Morrison, M.; Gray, D. Anaerobic Digestion Economic Feasibility Study: Generating Energy From Waste, Sewage and Sargassum Seaweed in the OECS; The Caribbean Council: London, UK, 2017. [Google Scholar]
- Carro, L.; Barriada, J.L.; Herrero, R.; de Vicente, M.E.S. Interaction of heavy metals with Ca-pretreated Sargassum muticum algal biomass: Characterization as a cation exchange process. Chem. Eng. J. 2015, 264, 181–187. [Google Scholar] [CrossRef]
- Besada, V.; Andrade, J.M.; Schultze, F.; González, J.J. Heavy metals in edible seaweeds commercialised for human consumption. J. Mar. Syst. 2009, 75, 305–313. [Google Scholar] [CrossRef]
- Khaled, A.; Hessein, A.; Abdel-Halim, A.M.; Morsy, F.M. Distribution of heavy metals in seaweeds collected along marsa-matrouh beaches, egyptian mediterranean sea. Egypt. J. Aquat. Res. 2014, 40, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, E.J.; Smith, K.L. Plastics on sargasso sea-surface. Science 1972, 175, 1240. [Google Scholar] [CrossRef]
- UNEP. Sargassum White Paper—Sargassum Outbreak in the Caribbean: Challenges, Opportunities and Regional Situation; Scientific and Technical Advisory Committee (STAC) to the Protocol Concerning Specially Protected Areas and Wildlife (SPAW) in the Wider Caribbean Region: Panama City, FL, USA, 2018. [Google Scholar]
- Davis, T.A.; Volesky, B.; Vieira, R. Sargassum seaweed as biosorbent for heavy metals. Water Res. 2000, 34, 4270–4278. [Google Scholar] [CrossRef] [Green Version]
- Schmuck, A.M.; Lavers, J.L.; Stuckenbrock, S.; Sharp, P.B.; Bond, A.L. Geophysical features influence the accumulation of beach debris on caribbean islands. Mar. Pollut. Bull. 2017, 121, 45–51. [Google Scholar] [CrossRef]
- Nkoa, R. Agricultural benefits and environmental risks of soil fertilization with anaerobic digestates: A review. Agron. Sustain. Dev. 2014, 34, 473–492. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Martínez, R.E.; Roy, P.D.; Torrescano-Valle, N.; Cabanillas-Terán, N.; Carrillo-Domínguez, S.; Collado-Vides, L.; García-Sánchez, M.; van Tussenbroek, B.I. Element concentrations in pelagic Sargassum along the Mexican Caribbean coast in 2018-2019. PeerJ 2020, 8, e8667. [Google Scholar] [CrossRef] [Green Version]
- Cameron, C.M.; Gatewood, J.B. Beyond Sun, Sand and Sea: The Emergent Tourism Programme in the Turks and Caicos Islands. J. Herit. Tour. 2008, 3, 55–73. [Google Scholar] [CrossRef]
- Statistics Department|turks and Caicos Islands Government. Turks and Caicos Islands Percentage Contribution of Gross Domestic Product by Economic Activity in Constant Prices: 2012–2018; Department, S., Ed.; Turks and Caicos Islands Government: Turks and Caicos Islands, UK, 2019.
- Anon. Turks and Caicos. Available online: https://lovinturksandcaicos.com/turks-and-caicos-government-news/the-tcig-has-an-important-message-about-the-seaweed-collecting-on-our-shores (accessed on 29 January 2018).
- Solid Biofuels. Determination of Moisture Content. Oven dry Method. Total Moisture. Simplified Method. In BS EN 14774-2:2009; BSI: London, UK, 2009.
- Solid Biofuels -Determination of Ash Content. In BS EN 14775:2009; BSI: London, UK, 2009.
- Pearson, D. Laboratory Techniques in Food Analysis; Butterworth: London, UK, 1973. [Google Scholar]
- Skoog, D.A. Fundamentals of Analytical Chemistry, 8th ed.; Thomson/Brooks/Cole: London, UK, 2004. [Google Scholar]
- Matanjun, P.; Mohamed, S.; Mustapha, N.M.; Muhammad, K.; Ming, C.H. Antioxidant activities and phenolics content of eight species of seaweeds from north Borneo. J. Appl. Phycol. 2008, 20, 367. [Google Scholar] [CrossRef]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AOAC. Method 960.52. In Official Methods of Analysis of AOAC International. Association; AOAC, Ed.; AOAC International: Gaithersburg, MD, USA, 1997. [Google Scholar]
- Determination of the Gross Heat of Combustion (Calorific Value). In BS EN ISO 1716:2010; BSI: London, UK, 2010.
- Online Combustion Handbook. Method from Combustion File 24; International Flame Research Foundation: Ijmuiden, The Netherlands, 2004.
- Heaven, S.; Milledge, J.; Zhang, Y. Comments on Anaerobic digestion of microalgae as a necessary step to make microalgal biodiesel sustainable. Biotechnol. Adv. 2011, 29, 164–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Symons, G.E.; Buswell, A.M. The methane fermentation of carbohydrates. J. Am. Chem. Soc. 1933, 55, 2028–2036. [Google Scholar] [CrossRef]
- Buswell, A.M.; Mueller, H.F. Mechanism of methane fermentation. Ind. Eng. Chem. 1952, 44, 550–552. [Google Scholar] [CrossRef]
- Tabassum, M.R.; Xia, A.; Murphy, J.D. Comparison of pre-treatments to reduce salinity and enhance biomethane yields of Laminaria digitata harvested in different seasons. Energy 2017, 140, 546–551. [Google Scholar] [CrossRef]
- Maneein, S.; Milledge, J.J.; Nielsen, B.V.; Harvey, P.J. A review of seaweed pre-treatment methods for enhanced biofuel production by anaerobic digestion or fermentation. Fermentation 2018, 4, 100. [Google Scholar] [CrossRef] [Green Version]
- Myers, S. Distribution and Composition of Sargassum Spp. Washed Ashore in the Turks and Caicos Islands MSc; University of Greenwich: London, UK, 2019. [Google Scholar]
- Schell, J.M.; Goodwin, D.S.; Siuda, A.N.S. Recent sargassum inundation events in the caribbean shipboard observations reveal dominance of a previously rare form. Oceanography 2015, 28, 8–10. [Google Scholar] [CrossRef] [Green Version]
- Sahena, F.; Zaidul, I.S.M.; Jinap, S.; Saari, N.; Jahurul, H.A.; Abbas, K.A.; Norulaini, N.A. Pufas in fish: Extraction, fractionation, importance in health. Compr. Rev. Food Sci. Food Saf. 2009, 8, 59–74. [Google Scholar] [CrossRef]
- Tiwari, B.; Troy, D. (Eds.) Seaweed Sustainability: Food and Non-Food Applications, 1st ed.; Academic Press: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Pereira, L. A review of the nutrient composition of selected edible seaweeds. In Seaweed; Pomin, V.H., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2011. [Google Scholar]
- Kraan, S. Mass-cultivation of carbohydrate rich macroalgae, a possible solution for sustainable biofuel production. Mitig. Adapt. Strateg. Glob. Chang. 2013, 18, 27–46. [Google Scholar] [CrossRef]
- Critchley, A.T.; Devisscher, P.R.M.; Nienhuis, P.H. Canopy characteristics of the brown alga Sargassum-muticum (fucales, phaeophyta) in lake grevelingen, southwest Netherlands. Hydrobiologia 1990, 204, 211–217. [Google Scholar] [CrossRef]
- Milledge, J.J.; Nielsen, B.V.; Bailey, D. High-value products from macroalgae: The potential uses of the invasive brown seaweed, Sargassum muticum. Rev. Environ. Sci. Biotechnol. 2015, 15, 67–88. [Google Scholar] [CrossRef]
- Beacham, T.A.; Cole, I.S.; DeDross, L.S.; Raikova, S.; Chuck, C.J.; Macdonald, J.; Herrera, L.; Ali, T.; Airs, R.L.; Landels, A.; et al. Analysis of seaweeds from south west england as a biorefinery feedstock. Appl. Sci. 2019, 9, 4456. [Google Scholar] [CrossRef] [Green Version]
- Oyesiku, O.O.; Egunyomi, A. Identification and chemical studies of pelagic masses of Sargassum natans (Linnaeus) Gaillon and S. fluitans (Borgessen) Borgesen (brown algae), found offshore in Ondo State, Nigeria. Afr. J. Biotechnol. 2014, 13, 1188–1193. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Wang, Q.; Jiang, X.M.; Han, X.X.; Ji, H.S. Compositional analysis of bio-oil derived from pyrolysis of seaweed. Energy Conv. Manag. 2013, 68, 273–280. [Google Scholar] [CrossRef]
- Shekhar, S.H.S.; Lyons, G.; McRoberts, C.; McCall, D.; Carmichael, E.; Andrews, F.; McCormack, R. Brown seaweed species from strangford lough: Compositional analyses of seaweed species and biostimulant formulations by rapid instrumental methods. J. Appl. Phycol. 2012, 24, 1141–1157. [Google Scholar] [CrossRef]
- Gorham, J.; Lewey, S.A. Seasonal changes in the chemical composition of Sargassum muticum. Mar. Biol. 1984, 80, 103–107. [Google Scholar] [CrossRef]
- Tabassum, M.R.; Xia, A.; Murphy, J.D. Seasonal variation of chemical composition and biomethane production from the brown seaweed Ascophyllum nodosum. Bioresour. Technol. 2016, 216, 219–226. [Google Scholar] [CrossRef]
- Jury, M.R. Turks and Caicos Islands Climate and Its Impacts. Earth Interact. 2013, 17, 1–20. [Google Scholar] [CrossRef]
- Sezey, M.; Adun, P. Validation of mohr’s titration method to determine salt in olive and olive brine. JOTCSA 2019, 6, 329–334. [Google Scholar] [CrossRef]
- Addico, G.N.D.; deGraft-Johnson, K. Preliminary investigation into the chemical composition of the invasive brown seaweed sargassum along the west coast of ghana. Afr. J. Biotechnol. 2016, 15, 2184–2191. [Google Scholar] [CrossRef] [Green Version]
- Sivakumar, S.R.; Arunkumar, K. Sodium, potassium and sulphate composition in some seaweeds occurring along the coast of gulf of mannar, India. Asian J. Plant Sci. 2009, 8, 500–504. [Google Scholar] [CrossRef]
- McHugh, D.J. A Guide to the Seaweed Industry; FAO: Rome, Italy, 2003. [Google Scholar]
- Philippsen, A. Energy Input, Carbon Intensity, and Cost for Ethanol Produced from Brown Seaweed; University of Victoria: Victoria, BC, Canada, 2013. [Google Scholar]
- Williams, A.; Feagin, R. Sargassum as a natural solution to enhance dune plant growth. Environ. Manag. 2010, 46, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, V.; Gopakumar, K. Shellfish: Nutritive value, health benefits, and consumer safety. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1219–1242. [Google Scholar] [CrossRef] [Green Version]
- Immaculate, J.K.; Lilly, T.T.; Jamila, P. Macro and micro nutrients of seagrass species from gulf of mannar, India. MOJ Food Process Technol. 2018, 6, 391–398. [Google Scholar] [CrossRef]
- Chen, Q.; Pan, X.-D.; Huang, B.-F.; Han, J.-L. Distribution of metals and metalloids in dried seaweeds and health risk to population in southeastern China. Sci. Rep. 2018, 8, 3578. [Google Scholar] [CrossRef] [Green Version]
- Cherry, P.; O’Hara, C.; Magee, P.J.; McSorley, E.M.; Allsopp, P.J. Risks and benefits of consuming edible seaweeds. Nutr. Rev. 2019, 77, 307–329. [Google Scholar] [CrossRef] [Green Version]
- Azzazy, M.F.; Ayyad, M.S.; Nofal, A.M.; Abdelsalam, I.Z.; Abousekken, M.S.; Tammam, O.A.S. Ecological and phytochemical studies on brown algae sargassum muticum from marsa alam at red sea coast, egypt. Alex. Sci. Exch. J. 2019, 40, 743–753. [Google Scholar] [CrossRef] [Green Version]
- Yokoi, K.; Konomi, A. Toxicity of so-called edible hijiki seaweed (Sargassum fusiforme) containing inorganic arsenic. Regul. Toxicol. Pharmacol. 2012, 63, 291–297. [Google Scholar] [CrossRef]
- Rose, M.; Lewis, J.; Langford, N.; Baxter, M.; Origgi, S.; Barber, M.; MacBain, H.; Thomas, K. Arsenic in seaweed—Forms, concentration and dietary exposure. Food Chem. Toxicol. 2007, 45, 1263–1267. [Google Scholar] [CrossRef]
- Nachman, K.E.; Ginsberg, G.L.; Miller, M.D.; Murray, C.J.; Nigra, A.E.; Pendergrast, C.B. Mitigating dietary arsenic exposure: Current status in the United States and recommendations for an improved path forward. Sci.Total Environ. 2017, 581–582, 221–236. [Google Scholar] [CrossRef]
- Commission, E. Amending Regulation (EC) No 1881/2006 as Regards Maximum Levels of Inorganic Arsenic in foodstuffs; EU, Ed.; Official Journal of the European Union: Brussels, Belgium, 2015. [Google Scholar]
- Roleda, M.Y.; Marfaing, H.; Desnica, N.; Jónsdóttir, R.; Skjermo, J.; Rebours, C.; Nitschke, U. Variations in polyphenol and heavy metal contents of wild-harvested and cultivated seaweed bulk biomass: Health risk assessment and implication for food applications. Food Control 2019, 95, 121–134. [Google Scholar] [CrossRef]
- Adamse, P.; Van der Fels-Klerx, H.J.I.; de Jong, J. Cadmium, lead, mercury and arsenic in animal feed and feed materials—Trend analysis of monitoring results. Food Addit. Contam. Part A 2017, 34, 1298–1311. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, F.A.; Smith, S.R.; Alloway, B.J.; Carlton-Smith, C.; Chambers, B.J. An inventory of heavy metals inputs to agricultural soils in England and Wales. Sci. Total Environ. 2003, 311, 205–219. [Google Scholar] [CrossRef]
- Tarvainen, T.; Albanese, S.; Birke, M.; Poňavič, M.; Reimann, C. Arsenic in agricultural and grazing land soils of Europe. Appl. Geochem. 2013, 28, 2–10. [Google Scholar] [CrossRef]
- Centre for Process Innovation (CPI). The SeaGas Project. Available online: http://seagas.co.uk/ (accessed on 7 July 2016).
- Da Silva, E.B.; Mussoline, W.A.; Wilkie, A.C.; Ma, L.Q. Anaerobic digestion to reduce biomass and remove arsenic from as-hyperaccumulator Pteris vittata. Environ. Pollut. 2019, 250, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Taylor, V.F.; Li, Z.; Sayarath, V.; Palys, T.J.; Morse, K.R.; Scholz-Bright, R.A.; Karagas, M.R. Distinct arsenic metabolites following seaweed consumption in humans. Sci. Rep. 2017, 7, 3920. [Google Scholar] [CrossRef] [PubMed]
- Tejada-Tejada, P.; Rodriguez-Rodriguez, Y.; Rodriguez, L.E.; Perdomo, O.P.; Boluda, C.J. Quantification of heavy metals in Sargassum natans and Sargassum fluitans arrived to the coasts of dominican republic during 2019. In Proceedings of the Caribbean Science and Innovation Meeting 2019, Pointe-à-Pitre (Guadeloupe), France, 21 October 2019. [Google Scholar]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Tanniou, A.; Esteban, S.L.; Vandanjon, L.; Ibanez, E.; Mendiola, J.A.; Cerantola, S.; Kervarec, N.; La Barre, S.; Marchal, L.; Stiger-Pouvreau, V. Green improved processes to extract bioactive phenolic compounds from brown macroalgae using Sargassum muticum as model. Talanta 2013, 104, 44–52. [Google Scholar] [CrossRef]
- Swanson, A.K.; Druehl, L.D. Induction, exudation and the UV protective role of kelp phlorotannins. Aquat. Bot. 2002, 73, 241–253. [Google Scholar] [CrossRef]
- Le Lann, K.; Jegou, C.; Stiger-Pouvreau, V. Effect of different conditioning treatments on total phenolic content and antioxidant activities in two Sargassacean species: Comparison of the frondose Sargassum muticum (Yendo) fensholt and the cylindrical bifurcaria bifurcata R. Ross. Phycol. Res. 2008, 56, 238–245. [Google Scholar] [CrossRef]
- Koivikko, R.; Loponen, J.; Honkanen, T.; Jormalainen, V. Contents of soluble, cell-wall-bound and exuded phlorotannins in the brown alga Fucus vesiculosus, with implications on their ecological functions. J. Chem. Ecol. 2005, 31, 195–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powers, L.C.; Hertkorn, N.; McDonald, N.; Schmitt-Kopplin, P.; Del Vecchio, R.; Blough, N.V.; Gonsior, M. Sargassum sp. act as a large regional source of marine dissolved organic carbon and polyphenols. Glob. Biogeochem. Cycles 2019, 33, 1423–1439. [Google Scholar] [CrossRef] [Green Version]
- Connan, S.; Delisle, F.; Deslandes, E.; Gall, E.A. Intra-thallus phlorotannin content and antioxidant activity in Phaeophyceae of temperate waters. Bot. Mar. 2006, 49, 39–46. [Google Scholar] [CrossRef]
- Tanniou, A.; Vandanjon, L.; Incera, M.; Leon, E.S.; Husa, V.; Le Grand, J.; Nicolas, J.L.; Poupart, N.; Kervarec, N.; Engelen, A.; et al. Assessment of the spatial variability of phenolic contents and associated bioactivities in the invasive alga Sargassum muticum sampled along its European range from Norway to Portugal. J. Appl. Phycol. 2014, 26, 1215–1230. [Google Scholar] [CrossRef] [Green Version]
- Lapointe, B.E.; West, L.E.; Sutton, T.T.; Hu, C. Ryther revisited: Nutrient excretions by fishes enhance productivity of pelagic Sargassum in the western North Atlantic Ocean. J. Exp. Mar. Biol. Ecol. 2014, 458, 46–56. [Google Scholar] [CrossRef]
- Mæhre, H.K.; Dalheim, L.; Edvinsen, G.K.; Elvevoll, E.O.; Jensen, I.-J. Protein determination—Method matters. Foods 2018, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Angell, A.R.; Mata, L.; Nys, R.; Paul, N.A. The protein content of seaweeds: A universal nitrogen-to-protein conversion factor of five. J. Appl. Phycol. 2015, 28, 511–524. [Google Scholar] [CrossRef]
- González López, C.V.; Garcia, M.D.C.; Fernandez, F.G.A.; Bustos, C.S.; Chisti, Y.; Sevilla, J.M.F. Protein measurements of microalgal and cyanobacterial biomass. Bioresour. Technol. 2010, 101, 7587–7591. [Google Scholar] [CrossRef]
- Safi, C.; Charton, M.; Pignolet, O.; Pontalier, P.-Y.; Vaca-Garcia, C. Evaluation of the protein quality of Porphyridium cruentum. J. Appl. Phycol. 2013, 25, 497–501. [Google Scholar] [CrossRef] [Green Version]
- WHO/FAO/UNU. Protein and Amino Acid Requirements in Human Nutrition—Report of a Joint WHO/FAO/UNU Expert Consultation; WHO: Geneva, Switzerland, 2007. [Google Scholar]
- Streefland, M. Algae and Aquatic Biomass for a Sustainable Production of 2nd Generation Biofuels—Deliverable 1.5—Report on Biofuel Production Processes from Micro, Macroalgae and Other Aquatic Biomass; AquaFUELs: Brussels, Belgium, 2010. [Google Scholar]
- Lenstra, W.J.; Hal, J.W.v.; Reith, J.H. Economic aspects of open ocean seaweed cultivation. In Proceedings of the Alg’n Chem 2011, Montpellier, France, 7–10 November 2011. [Google Scholar]
- Van Ginneken, V.J.T.; Helsper, J.; de Visser, W.; van Keulen, H.; Brandenburg, W.A. Polyunsaturated fatty acids in various macroalgal species from north Atlantic and tropical seas. Lipids Health Dis. 2011, 10, 8. [Google Scholar] [CrossRef] [Green Version]
- Kumari, P.; Bijo, A.J.; Mantri, V.A.; Reddy, C.R.K.; Jha, B. Fatty acid profiling of tropical marine macroalgae: An analysis from chemotaxonomic and nutritional perspectives. Phytochemistry 2013, 86, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Bazes, A.; Silkina, A.; Douzenel, P.; Fay, F.; Kervarec, N.; Morin, D.; Berge, J.-P.; Bourgougnon, N. Investigation of the antifouling constituents from the brown alga Sargassum muticum (Yendo) Fensholt. J. Appl. Phycol. 2009, 21, 395–403. [Google Scholar] [CrossRef] [Green Version]
- Plouguerne, E.; Ioannou, E.; Georgantea, P.; Vagias, C.; Roussis, V.; Hellio, C.; Kraffe, E.; Stiger-Pouvreau, V. Anti-microfouling activity of lipidic metabolites from the invasive brown alga sargassum muticum (yendo) fensholt. Mar. Biotechnol. 2010, 12, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.P.; Rooker, J.R. Fatty acid composition of flora and fauna associated with sargassum mats in the gulf of Mexico. Mar. Biol. 2006, 149, 1025–1036. [Google Scholar] [CrossRef]
- Milledge, J.J. Commercial application of microalgae other than as biofuels: A brief review. Rev. Environ. Sci. Biotechnol. 2011, 10, 31–41. [Google Scholar] [CrossRef]
- Øverland, M.; Mydland, L.T.; Skrede, A. Marine macroalgae as sources of protein and bioactive compounds in feed for monogastric animals. J. Sci. Food Agric. 2019, 99, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef]
- Anastasakis, K.; Ross, A.B. Hydrothermal liquefaction of the brown macro-alga Laminaria Saccharina: Effect of reaction conditions on product distribution and composition. Bioresour. Technol. 2011, 102, 4876–4883. [Google Scholar] [CrossRef]
- Ross, A.B.; Jones, J.M.; Kubacki, M.L.; Bridgeman, T. Classification of macroalgae as fuel and its thermochemical behaviour. Bioresour. Technol. 2008, 99, 6494–6504. [Google Scholar] [CrossRef]
- Zhou, D.; Zhang, L.; Zhang, S.; Fu, H.; Chen, J. Hydrothermal liquefaction of macroalgae Enteromorpha prolifera to bio-oil. Energy Fuels 2010, 24, 4054–4061. [Google Scholar] [CrossRef]
- Vargas-Moreno, J.M.; Callejón-Ferre, A.J.; Pérez-Alonso, J.; Velázquez-Martí, B. A review of the mathematical models for predicting the heating value of biomass materials. Renew. Sustain. Energy Rev. 2012, 16, 3065–3083. [Google Scholar] [CrossRef]
- Barreiro-Vescovo, S.; de Godos, I.; Tomás-Pejó, E.; Ballesteros, M.; González-Fernández, C. Effect of microalgae storage conditions on methane yields. Environ. Sci. Pollut. Res. 2018, 25, 14263–14270. [Google Scholar] [CrossRef] [PubMed]
- Purcell, P.J.; O’Brien, M.; Boland, T.M.; O’Kiely, P. In vitro rumen methane output of perennial ryegrass samples prepared by freeze drying or thermal drying (40 °C). Anim. Feed Sci. Technol. 2011, 166–167, 175–182. [Google Scholar] [CrossRef]
- Adams, J.M.M.; Schmidt, A.; Gallagher, J.A. The impact of sample preparation of the macroalgae Laminaria digitata on the production of the biofuels bioethanol and biomethane. J. Appl. Phycol. 2015, 27, 985–991. [Google Scholar] [CrossRef] [Green Version]
- Milledge, J.J.; Nielsen, B.V.; Harvey, P.J. The inhibition of anaerobic digestion by model phenolic compounds representative of those from Sargassum muticum. J. Appl. Phycol. 2018, 31, 779–786. [Google Scholar] [CrossRef]
- Østgaard, K.; Indergaard, M.; Markussen, S.; Knutsen, S.H.; Jensen, A. Carbohydrate degradation and methane production during fermentation ofLaminaria saccharina (Laminariales, Phaeophyceae). J. Appl. Phycol. 1993, 5, 333–342. [Google Scholar] [CrossRef]
- Adams, J.M.M.; Ross, A.B.; Anastasakis, K.; Hodgson, E.M.; Gallagher, J.A.; Jones, J.M.; Donnison, I.S. Seasonal variation in the chemical composition of the bioenergy feedstock Laminaria digitata for thermochemical conversion. Bioresour. Technol. 2011, 102, 226–234. [Google Scholar] [CrossRef]
- Buffiere, P.; Loisel, D.; Bernet, N.; Delgenes, J.-P. Towards new indicators for the prediction of solid waste anaerobic digestion properties. Water Sci. Technol. 2006, 53, 233–241. [Google Scholar] [CrossRef] [Green Version]
- McKennedy, J.; Sherlock, O. Anaerobic digestion of marine macroalgae: A review. Renew. Sustain. Energy Rev. 2015, 52, 1781–1790. [Google Scholar] [CrossRef]
- Mao, C.; Feng, Y.; Wang, X.; Ren, G. Review on research achievements of biogas from anaerobic digestion. Renew. Sustain. Energy Rev. 2015, 45, 540–555. [Google Scholar] [CrossRef]
- Milledge, J.J.; Harvey, P.J. Anaerobic digestion and gasification of seaweed. In Grand Challenges in Marine Biotechnology; Rampelotto, P.H., Trincone, A., Eds.; Spinger: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Persson, S.P.E.; Bartlett, H.D.; Branding, A.E.; Regan, R.W. Agricultural Anaerobic Digesters; Pennsylvania State University: State College, PA, USA, 1979. [Google Scholar]
- Peu, P.; Picard, S.; Diara, A.; Girault, R.; Béline, F.; Bridoux, G.; Dabert, P. Prediction of hydrogen sulphide production during anaerobic digestion of organic substrates. Bioresour. Technol. 2012, 121, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Puyuelo, B.; Ponsá, S.; Gea, T.; Sánchez, A. Determining C/N ratios for typical organic wastes using biodegradable fractions. Chemosphere 2011, 85, 653–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Lu, X.; Li, F.; Yang, G. Effects of Temperature and Carbon-Nitrogen (C/N) Ratio on the Performance of Anaerobic Co-Digestion of Dairy Manure, Chicken Manure and Rice Straw: Focusing on Ammonia Inhibition. PLoS ONE 2014, 9, e97265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Este, M.; Angelidaki, I.; Alvarado-Morales, M. Algal Biomass for Bioenergy and Bioproducts Production in Biorefinery Concepts; Department of Environmental Engineering, Technical University of Denmark (DTU): Lyngby, Denmark, 2017. [Google Scholar]
- Chynoweth, D.P.; Ghosh, S.; Klass, D.L. Anaerobic digestion of kelp. In Biomass Conversion Processes for Energy and Fuels; Sofer, S.S., Zaborsky, O.R., Eds.; Springer USA: Boston, MA, USA, 1981; pp. 315–338. [Google Scholar] [CrossRef]
- Chynoweth, D.P. Review of Biomethane from Marine Biomass; Department of Agricultural and Biological Engineering, University of Florida: Gainesville, FL, USA, 2002. [Google Scholar]
- Ometto, F.; Steinhovden, K.B.; Kuci, H.; Lunnbäck, J.; Berg, A.; Karlsson, A.; Handå, A.; Wollan, H.; Ejlertsson, J. Seasonal variation of elements composition and biomethane in brown macroalgae. Biomass Bioenergy 2018, 109, 31–38. [Google Scholar] [CrossRef]
- Shakeri Yekta, S.; Svensson, B.H.; Björn, A.; Skyllberg, U. Thermodynamic modeling of iron and trace metal solubility and speciation under sulfidic and ferruginous conditions in full scale continuous stirred tank biogas reactors. Appl. Geochem. 2014, 47, 61–73. [Google Scholar] [CrossRef]
- Sierra-Alvarez, R.; Cortinas, I.; Yenal, U.; Field, J.A. Methanogenic inhibition by arsenic compounds. Appl. Environ. Microbiol. 2004, 70, 5688–5691. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Freire, L.; Moore, S.E.; Sierra-Alvarez, R.; Field, J.A. Adaptation of a Methanogenic Consortium to Arsenite Inhibition. Water Air Soil Pollut. 2015, 226, 414. [Google Scholar] [CrossRef] [Green Version]
- Molina-Alcaide, E.; Carro, M.D.; Roleda, M.Y.; Weisbjerg, M.R.; Lind, V.; Novoa-Garrido, M. In vitro ruminal fermentation and methane production of different seaweed species. Anim. Feed Sci. Technol. 2017, 228, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tabassum, M.R.; Xia, A.; Murphy, J.D. The effect of seasonal variation on biomethane production from seaweed and on application as a gaseous transport biofuel. Bioresour. Technol. 2016, 209, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Hierholtzer, A.; Chatellard, L.; Kierans, M.; Akunna, J.C.; Collier, P.J. The impact and mode of action of phenolic compounds extracted from brown seaweed on mixed anaerobic microbial cultures. J. Appl. Microbiol. 2013, 114, 964–973. [Google Scholar] [CrossRef]
- Pérez, M.J.; Falqué, E.; Domínguez, H. Antimicrobial action of compounds from marine seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, A.J.; Lewis, D.M.; Green, B. Anaerobic digestion of algae biomass: A review. Algal Res. Biomass Biofuels Bioprod. 2014, 5, 204–214. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, Z.; Bach, S.J.; McAllister, T.A. Effects of phlorotannins from Ascophyllum nodosum (brown seaweed) on in vitro ruminal digestion of mixed forage or barley grain. Anim. Feed Sci. Technol. 2008, 145, 375–395. [Google Scholar] [CrossRef]
- Monlau, F.; Sambusiti, C.; Barakat, A.; Quéméneur, M.; Trably, E.; Steyer, J.P.; Carrère, H. Do furanic and phenolic compounds of lignocellulosic and algae biomass hydrolyzate inhibit anaerobic mixed cultures? A comprehensive review. Biotechnol. Adv. 2014, 32, 934–951. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Moisture % ar | Ash % dw | TS % | VS % | |
|---|---|---|---|---|
| Ave | Ave | |||
| Mixed ‘Sargassum’ | 81.98 ± 0.89 | 46.94 ± 1.31 | 18.00 | 9.56 |
| S. natans VIII | 86.45 ± 0.10 | 34.26 ± 0.59 | 13.50 | 8.91 |
| S. natans I | 87.41 ± 0.23 | 35.71 ± 1.27 | 12.60 | 8.10 |
| S. fluitans | 86.32 ± 0.02 | 33.63 ± 4.14 | 13.70 | 9.08 |
| Salt % ash | Salt % dw | Salt % ww | |
|---|---|---|---|
| Ave | |||
| Mixed ‘Sargassum’ | 32.30 ± 5.09 | 15.2 | 2.7 |
| S. natans VIII | 56.87 ± 1.55 | 19.5 | 2.6 |
| S. natans I | 64.60 ± 2.80 | 23.1 | 2.9 |
| S. fluitans | 57.00 ± 0.00 | 19.2 | 2.6 |
| CaCO₃ | KCl | NaCl | MgO | K3Na (SO4)2 | CaSO4 | Na2SO4 | |
|---|---|---|---|---|---|---|---|
| % in Ash | |||||||
| Mixed ‘Sargassum’ | 42.07 | 23.93 | 19.03 | 4.88 | 3.76 | 4.98 | 1.36 |
| S. natans VIII | 17.99 | 9.43 | 57.02 | 7.04 | 1.38 | 5.67 | 1.27 |
| S natans I | 11.68 | 0.26 | 71.58 | 8.26 | 0.28 | 3.39 | 3.15 |
| S fluitans | 11.68 | 0.26 | 71.59 | 8.26 | 8.26 | 7.69 | 0.24 |
| Ash | C | H | N | S | O | Elemental Ratios | ||
|---|---|---|---|---|---|---|---|---|
| % Dry Weight | C:N | C:O | ||||||
| Mixed ‘Sargassum’ | 46.94 | 27.41 | 3.13 | 1.71 | 0.21 | 20.62 | 16.08 | 1.33 |
| S. natans VIII | 34.26 | 29.23 | 3.68 | 1.68 | 0.40 | 30.76 | 17.40 | 0.95 |
| S. natans I | 35.71 | 28.34 | 3.63 | 1.28 | 0.05 | 31.00 | 22.14 | 0.91 |
| S. fluitans | 33.63 | 29.23 | 3.78 | 1.57 | 0.00 | 31.79% | 18.62 | 0.92 |
| Mixed ‘Sargassum’ | S. Natans VIII | S. Natans I | S. Fluitans | ||
|---|---|---|---|---|---|
| Aluminium | mg kg−1 dw | 37.5 | 16.21 | 21.48 | 28.09 |
| Arsenic | mg kg−1 dw | 123.69 | 20.94 | 29.76 | 26.25 |
| Cadmium | mg kg−1 dw | 0.13 | 0.09 | 0.12 | 0.12 |
| Calcium | mg kg−1 dw | 70,305.77 | 26,019.69 | 28,879.26 | 33,196.4 |
| Chromium | mg kg−1 dw | <0.3 | 0.36 | ND | 0.43 |
| Copper | mg kg−1 dw | 2.51 | 1.25 | 2.71 | 2.91 |
| Iron | mg kg−1 dw | 3811.37 | 81.58 | 998.56 | 262.02 |
| Lead | mg kg−1 dw | 0.26 | 0.48 | 0.28 | 0.37 |
| Magnesium | mg kg−1 dw | 12,053.19 | 15,092.59 | 16,546.71 | 16,320.64 |
| Manganese | mg kg−1 dw | 30.15 | <3 | <3 | <3 |
| Mercury | mg kg−1 dw | 0.01 | ND | 0.01 | 0.01 |
| Phosphorus | mg kg−1 dw | 500.65 | 138.3 | 222.15 | 214.28 |
| Potassium | mg kg−1 dw | 69,359.39 | 7442.57 | 12,509.16 | 7771.73 |
| Zinc | mg kg−1 dw | 5.81 | 26.49 | 30.88 | 35.64 |
| Phenols (PG eq) | ||
|---|---|---|
| mg g−1 dw | mg g−1 VS. | |
| Ave | Ave | |
| Mixed ‘Sargassum’ | 29.5 ± 0.5 | 55.5 |
| S. natans VIII | 2.5 ± 0.2 | 3.8 |
| S. natans I | 6.6 ± 0.4 | 10.3 |
| S. fluitans | 3.7 ± 0.2 | 5.6 |
| Mixed ‘Sargassum’ | S. Natans VIII | S. Natans I | S. Fluitans | |
|---|---|---|---|---|
| Amino acid | % dw | |||
| Alanine | 0.34 | 0.13 | 0.26 | 0.19 |
| Arginine | 0.18 | 0.14 | 0.19 | 0.17 |
| Aspartic acid | 0.47 | 0.34 | 0.48 | 0.42 |
| Cystine | 0.09 | 0.11 | 0.09 | 0.09 |
| Glutamic | 0.85 | 0.35 | 0.58 | 0.46 |
| Glycine | 0.32 | 0.19 | 0.30 | 0.24 |
| Histidine | 0.06 | 0.05 | 0.07 | 0.07 |
| Iso-Leucine | 0.16 | 0.13 | 0.18 | 0.14 |
| Leucine | 0.27 | 0.18 | 0.28 | 0.23 |
| Lysine | 0.24 | 0.28 | 0.23 | 0.21 |
| Methionine | 0.10 | 0.14 | 0.10 | 0.09 |
| Phenylalanine | 0.18 | 0.14 | 0.19 | 0.17 |
| Proline | 0.18 | 0.06 | 0.14 | 0.11 |
| Serine | 0.22 | 0.19 | 0.22 | 0.20 |
| Threonine | 0.19 | 0.18 | 0.21 | 0.19 |
| Tryptophan | 0.04 | 0.05 | 0.04 | 0.04 |
| Tyrosine | 0.01 | 0.00 | 0.01 | 0.00 |
| Valine | 0.24 | 0.35 | 0.24 | 0.23 |
| Total Amino acids | 4.16 | 2.99 | 3.81 | 3.25 |
| % of TFA | ||||
|---|---|---|---|---|
| Mixed Sargassum | S. Natans VIII | S. Natans I | S. Fluitans | |
| C08:0 Caprylic | <0.05 | <0.05 | <0.05 | <0.05 |
| C10:0 Capric Acid | <0.05 | <0.05 | <0.05 | <0.05 |
| C11:0 Undecylic Acid | <0.05 | <0.05 | <0.05 | <0.05 |
| C12:0 Lauric Acid | 0.14 | <0.05 | 0.13 | 0.19 |
| C13:0 Tridecylic Acid | <0.05 | <0.05 | <0.05 | <0.05 |
| C14:0 Myristic Acid | 2.01 | 2.00 | 1.56 | 2.13 |
| C14:1 Myristoleic Acid | 0.43 | <0.05 | 0.15 | <0.05 |
| C15:0 Pentadecanoic Acid | 0.46 | 0.25 | 0.32 | 0.36 |
| C15:1 Pentadecenoic Acid | 0.39 | <0.05 | <0.05 | 0.37 |
| C16:0 Palmitic Acid | 26.68 | 40.71 | 23.61 | 24.12 |
| C16:1 Palmitoleic Acid | 4.03 | 8.28 | 3.54 | 4.13 |
| C17:0 Heptadecanoic Acid | 1.17 | 0.13 | 0.88 | 0.76 |
| C17:1 Heptadecenoic Acid | <0.05 | 0.19 | 0.63 | <0.05 |
| C18:0 Stearic Acid | 4.73 | 0.85 | 4.18 | 4.32 |
| C18:1 Oleic Acid | 12.71 | 10.71 | 13.31 | 15.23 |
| C18:2 Linoleic Acid | 5.32 | 7.90 | 6.92 | 6.02 |
| C18:3 Linolenic Acid | 4.4 | 3.52 | 5.9 | 3.48 |
| C18:4 Stearidonic Acid | 0.07 | 0.69 | 1.34 | 0.87 |
| C20:0 Arachidic Acid | 0.47 | 0.39 | 0.55 | 0.62 |
| C20:1 Gadoleic Acid | 0.18 | 0.76 | <0.05 | <0.05 |
| C20:4 Arachidonic Acid | 7.79 | 12.95 | 9.14 | 10.24 |
| C20:5 Eicosapentaenoic Acid | 3.75 | <0.05 | 2.77 | 1.49 |
| C22:0 Behenic Acid | 0.63 | 1.28 | 0.83 | 0.75 |
| C22:1 Erucic Acid | 1.59 | <0.05 | 1.56 | 2.11 |
| C22:4 Adrenic Acid | 1.17 | <0.05 | 0.77 | 0.78 |
| C22:5 Docosapentaenoic acid | 0.36 | <0.05 | 0.27 | 0.3 |
| C22:6 Docosahexaenoic Acid | 6.44 | <0.05 | 5.66 | 5.91 |
| C24:0 Lignoceric Acid | 0.42 | <0.05 | 0.35 | 0.44 |
| Monounsaturated Fatty Acids | 19.33 | 19.94 | 19.19 | 21.84 |
| Polyunsaturated Fatty Acids | 29.3 | 25.06 | 32.77 | 29.09 |
| Saturated Fatty Acids | 36.71 | 45.61 | 32.41 | 33.69 |
| Unidentified Fatty Acids | 14.66 | 9.39 | 15.63 | 15.38 |
| Lipid% | Total % AAs | Total % Fibre | Carbohydrate% | |
|---|---|---|---|---|
| Mixed ‘Sargassum’ | 3.88 ± 1.09 | 4.19 | 33.31 ± 0.90 | 11.68 |
| S. natans VIII | 3.58 ± 0.59 | 2.99 | 37.41 ± 0.43 | 21.76 |
| S natans I | 4.51 ± 0.90 | 3.81 | 37.00 ± 0.42 | 18.97 |
| S fluitans | 4.56 ± 0.90 | 3.25 | 31.15 ± 0.35 | 27.40 |
| HHV (kJ kg−1) | |||
|---|---|---|---|
| Measured | Calculated | ||
| Heaven et al. [56] | CHNSO | ||
| Mixed ‘Sargassum’ | 9.39 ± 0.27 | 9.8 | 10.7 |
| S. natans VIII | 10.23 ± 0.08 | 11.8 | 10.9 |
| S. natans I | 10.15 ±0.01 | 11.8 | 10.5 |
| S. fluitans | 10.26 ±0.11 | 11.9 | 10.8 |
| Methane Potential mL CH4 g−1 VS | |||||||
|---|---|---|---|---|---|---|---|
| Actual | Theoretical | Biodegradability Index | |||||
| CHNS | Heaven | Heaven ex Fibre | CHNS | Heaven | Heaven ex Fibre | ||
| Mixed ‘Sargassum’ | −24.0 | 496 | 461 | 195 | −5% | −5% | −12% |
| S. natans VIII | 145.1 | 395 | 449 | 207 | 37% | 32% | 70% |
| S. natans I | 65.8 | 392 | 460 | 187 | 17% | 14% | 35% |
| S. fluitans | 112.7 | 392 | 464 | 221 | 29% | 24% | 51% |
| Percentage of Total Amino Acids | |||||
|---|---|---|---|---|---|
| WHO | Mixed | S. Natans VIII | S. Natans I | S. Fluitans | |
| Recommendation | Sargassum | ||||
| Histidine | 1.50 | 1.55 | 1.57 | 1.75 | 2.02 |
| Isoleucine | 3.00 | 3.89 | 4.31 | 4.66 | 4.38 |
| Leucine | 5.90 | 6.48 | 5.88 | 7.29 | 7.07 |
| Lysine | 4.50 | 5.70 | 9.41 | 6.12 | 6.40 |
| Methionine | 1.60 | 2.33 | 4.71 | 2.62 | 2.69 |
| Cystine | 0.60 | 2.07 | 3.53 | 2.33 | 2.69 |
| Methionine + cysteine | 2.20 | 4.40 | 8.24 | 4.96 | 5.39 |
| Phenylalanine + tyrosine | 3.00 | 4.66 | 4.71 | 5.25 | 6.40 |
| Threonine | 2.30 | 4.66 | 5.88 | 5.54 | 5.72 |
| Tryptophan | 0.40 | 1.04 | 1.57 | 1.17 | 1.35 |
| Valine | 3.90 | 5.70 | 11.76 | 6.41 | 7.17 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milledge, J.J.; Maneein, S.; Arribas López, E.; Bartlett, D. Sargassum Inundations in Turks and Caicos: Methane Potential and Proximate, Ultimate, Lipid, Amino Acid, Metal and Metalloid Analyses. Energies 2020, 13, 1523. https://doi.org/10.3390/en13061523
Milledge JJ, Maneein S, Arribas López E, Bartlett D. Sargassum Inundations in Turks and Caicos: Methane Potential and Proximate, Ultimate, Lipid, Amino Acid, Metal and Metalloid Analyses. Energies. 2020; 13(6):1523. https://doi.org/10.3390/en13061523
Chicago/Turabian StyleMilledge, John James, Supattra Maneein, Elena Arribas López, and Debbie Bartlett. 2020. "Sargassum Inundations in Turks and Caicos: Methane Potential and Proximate, Ultimate, Lipid, Amino Acid, Metal and Metalloid Analyses" Energies 13, no. 6: 1523. https://doi.org/10.3390/en13061523
APA StyleMilledge, J. J., Maneein, S., Arribas López, E., & Bartlett, D. (2020). Sargassum Inundations in Turks and Caicos: Methane Potential and Proximate, Ultimate, Lipid, Amino Acid, Metal and Metalloid Analyses. Energies, 13(6), 1523. https://doi.org/10.3390/en13061523