Nitrogen Deprivation in Fremyella diplosiphon Augments Lipid Production without Affecting Growth


 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cyanobacterial Strain and Culture Conditions
2.2. Growth of F. diplosiphon in Varying Sodium Nitrate Concentrations
2.3. Lipid Accumulation in F. diplosiphon Grown in Varying Levels of Sodium Nitrate
2.4. Simultaneous Transesterification and Lipid Extraction
2.5. Gas Chromatography-Mass Spectrometry in F. diplosiphon Grown in Varying Sodium Nitrate Levels
2.6. Statistical Analysis
3. Results and Discussion
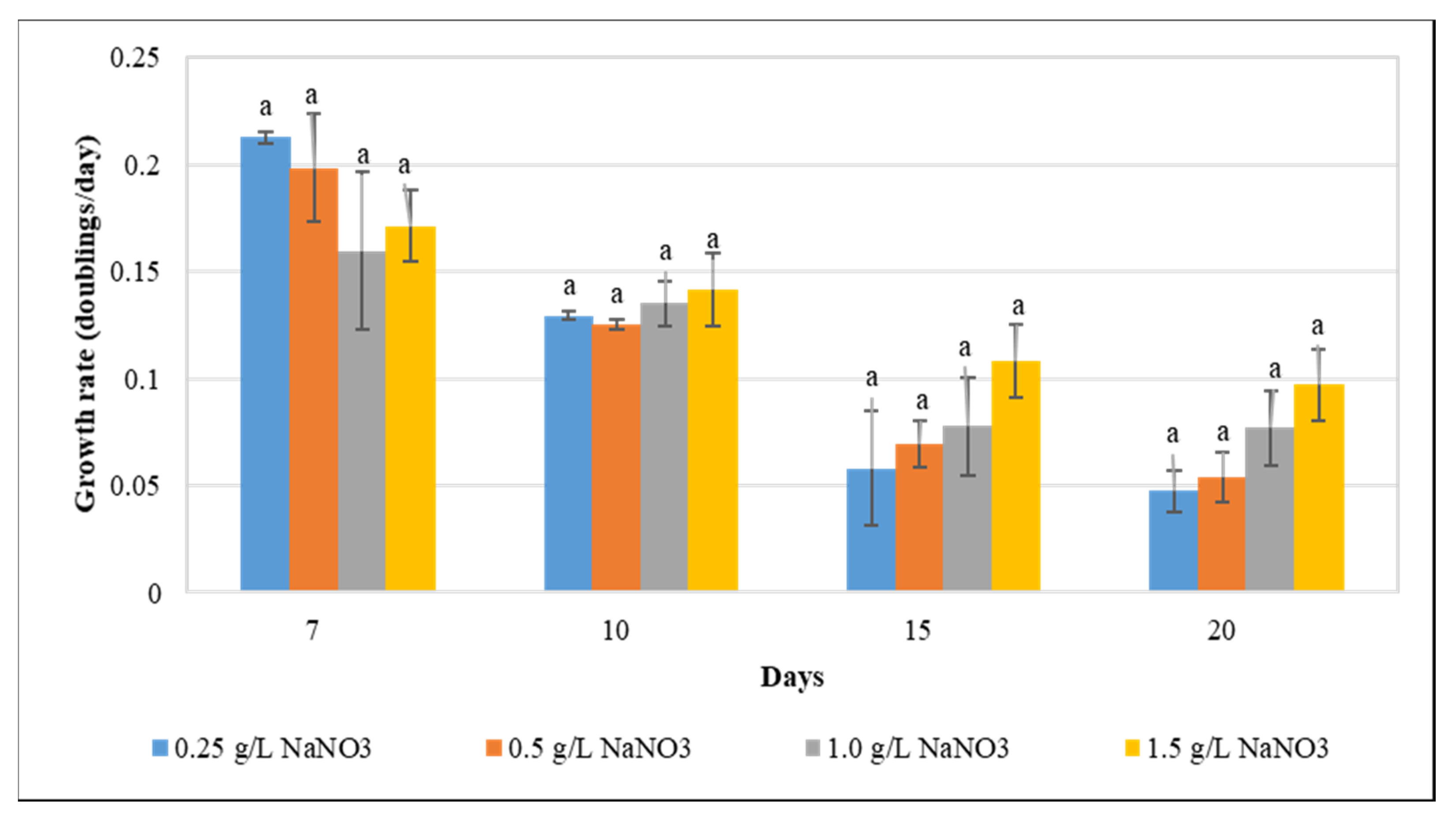
3.1. Effect of Nitrogen Deprivation on F. diplosiphon Growth
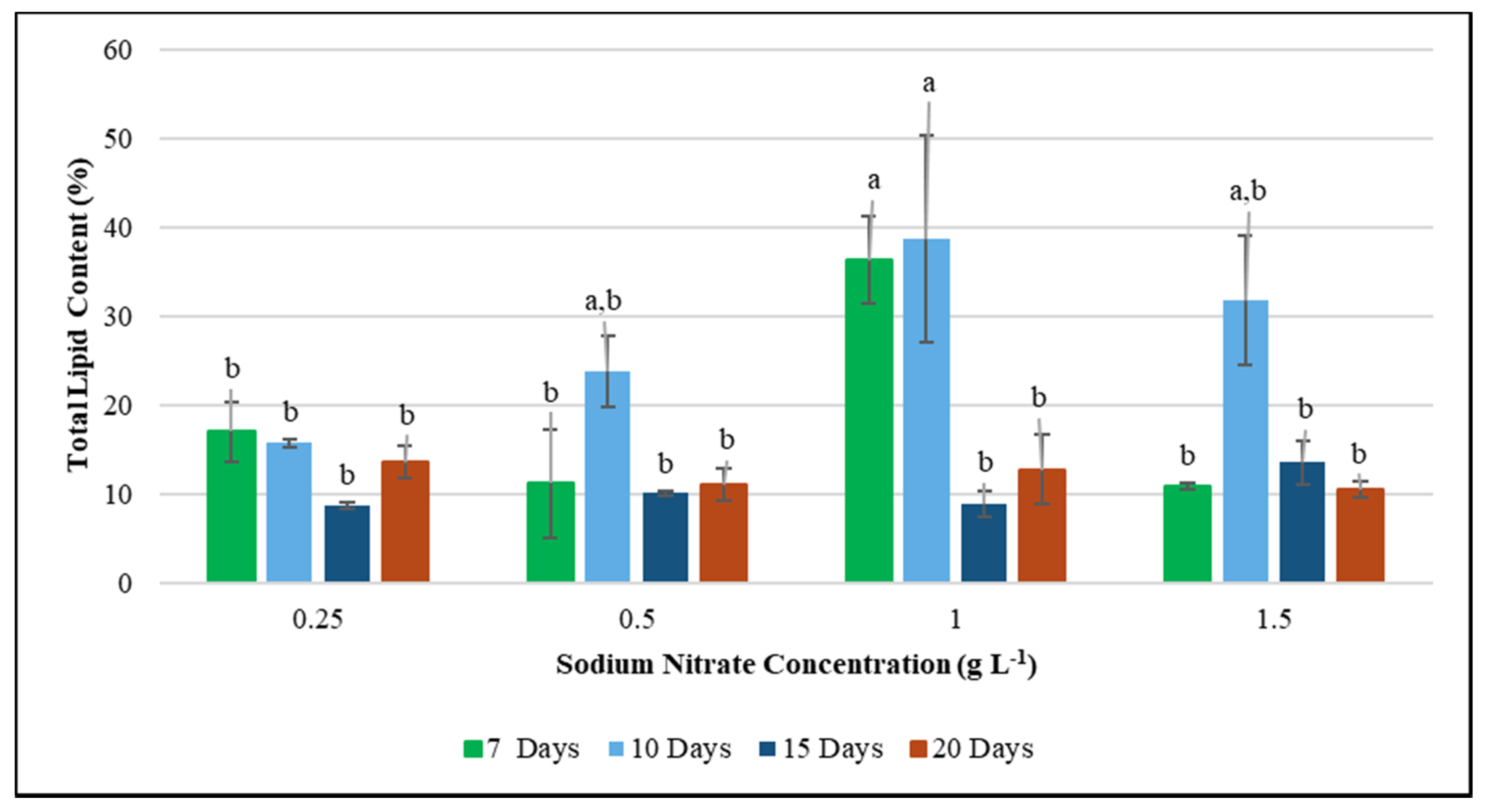
3.2. Effect of Nitrogen Deficiency on F. diplosiphon Total Lipid Content
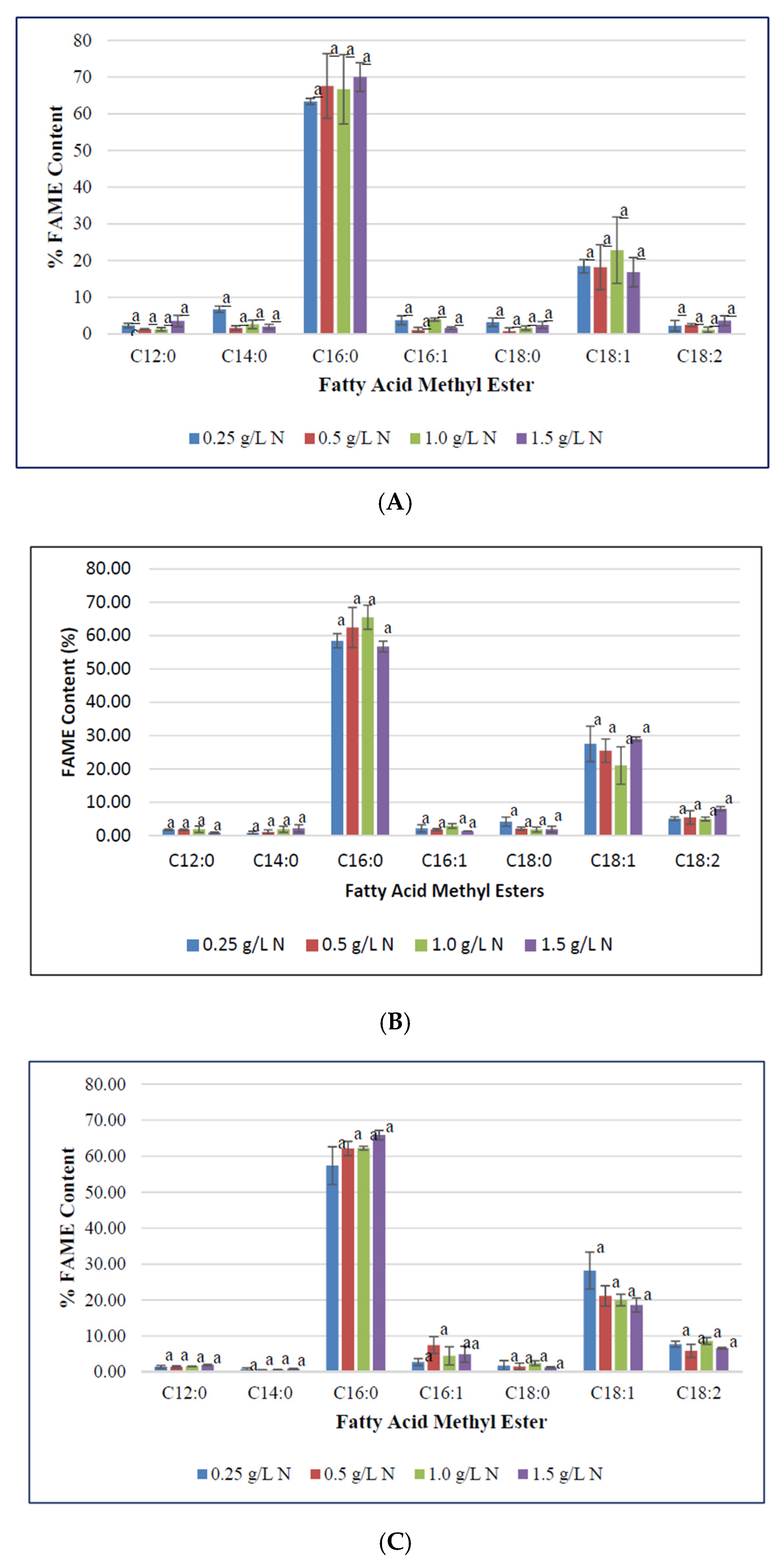
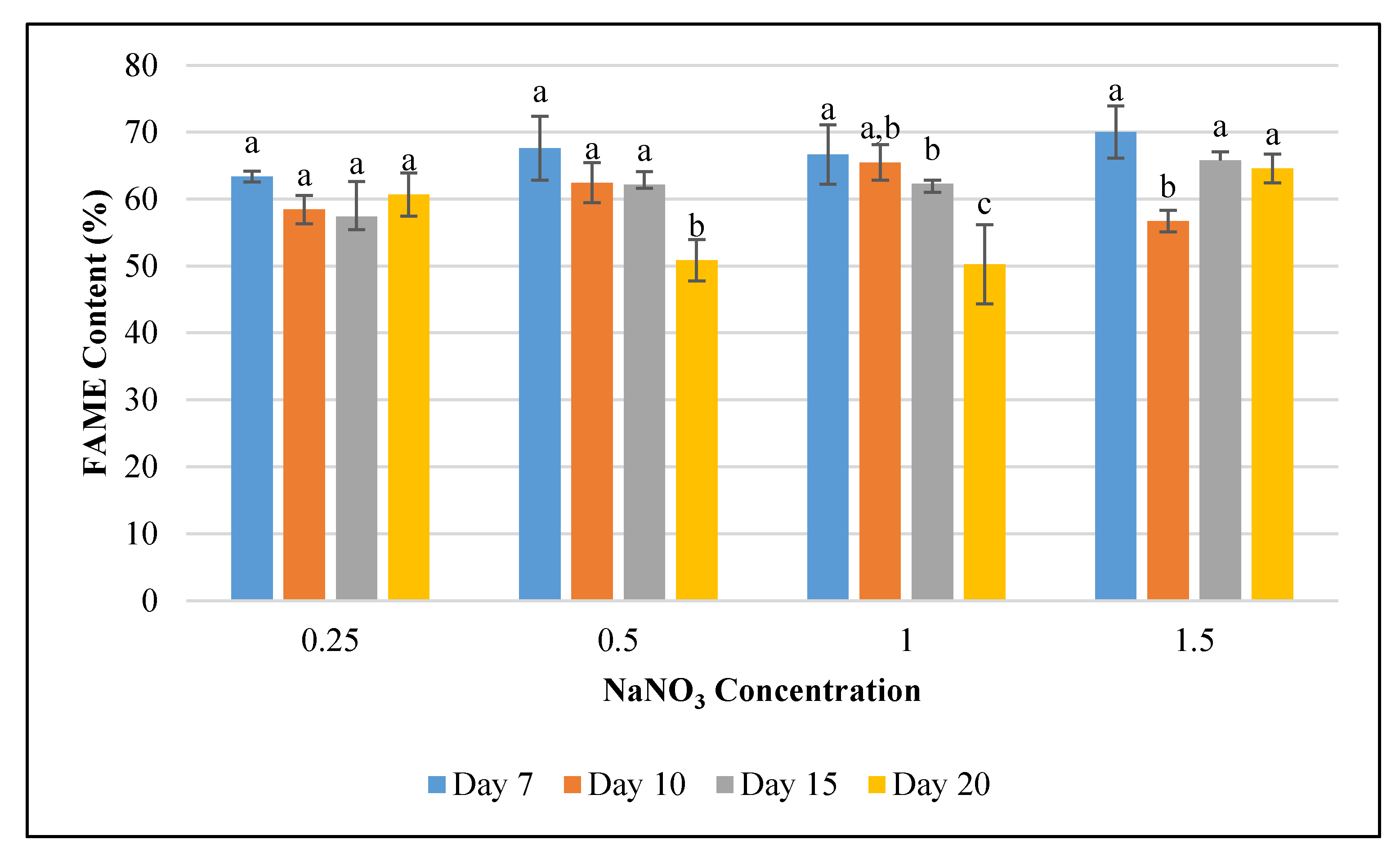
3.3. Effect of Nitrogen Deficiecny on F. diplosiphon Fatty acid Profile
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef] [PubMed]
- John, R.P.; Anisha, G.; Nampoothiri, K.M.; Pandey, A. Micro and macroalgal biomass: A renewable source for bioethanol. Bioresour. Technol. 2011, 102, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Benemann, J. Utilization of carbon dioxide from fossil fuel-burning power plants with biological systems. Energy Convers. Manag. 1993, 34, 999–1004. [Google Scholar] [CrossRef]
- Brennan, L.; Owende, P. Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sustain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Demirbas, A. Use of algae as biofuel sources. Energy Convers. Manag. 2010, 51, 2738–2749. [Google Scholar] [CrossRef]
- Atabani, A.; Silitonga, A.; Badruddin, I.A.; Mahlia, T.; Masjuki, H.; Mekhilef, S. A comprehensive review on biodiesel as an alternative energy resource and its characteristics. Renew. Sustain. Energy Rev. 2012, 16, 2070–2093. [Google Scholar] [CrossRef]
- Gouveia, L.; Oliveira, A.C. Microalgae as a raw material for biofuels production. J. Ind. Microbiol. Biotechnol. 2008, 36, 269–274. [Google Scholar] [CrossRef]
- Campbell, P.K.; Beer, T.; Batten, D. Life cycle assessment of biodiesel production from microalgae in ponds. Bioresour. Technol. 2011, 102, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Rouhany, M.; Montgomery, H. Global biodiesel production: The state of the art and impact on climate change: From production to combustion. In Biodiesel; Springer: Cham, Switzerland; 2019; pp. 1–14. [Google Scholar]
- Huang, G.; Chen, F.; Wei, D.; Zhang, X.; Chen, G. Biodiesel production by microalgal biotechnology. Appl. Energy 2010, 87, 38–46. [Google Scholar] [CrossRef]
- Khalid, K.; Khalid, K. Transesterification of palm oil for the production of biodiesel. Am. J. Appl. Sci. 2011, 8, 804–809. [Google Scholar] [CrossRef]
- Takeshita, T. Competitiveness, role, and impact of microalgal biodiesel in the global energy future. Appl. Energy 2011, 88, 3481–3491. [Google Scholar] [CrossRef]
- Lang, X.; Dalai, A.K.; Bakhshi, N.N.; Reaney, M.J.; Hertz, P.B. Preparation and characterization of bio-diesels from various bio-oils. Bioresour. Technol. 2001, 80, 53–62. [Google Scholar] [CrossRef]
- Aikawa, S.; Joseph, A.; Yamada, R.; Izumi, Y.; Yamagishi, T.; Matsuda, F.; Kawai, H.; Chang, J.-S.; Hasunuma, T.; Kondo, A. Direct conversion of Spirulina to ethanol without pretreatment or enzymatic hydrolysis processes. Energy Environ. Sci. 2013, 6, 1844–1849. [Google Scholar] [CrossRef]
- Dismukes, G.C.; Carrieri, D.; Bennette, N.; Ananyev, G.M.; Posewitz, M.C. Aquatic phototrophs: Efficient alternatives to land-based crops for biofuels. Curr. Opin. Biotechnol. 2008, 19, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Angermayr, S.A.; Hellingwerf, K.J.; Lindblad, P.; De Mattos, M.J.T. Energy biotechnology with cyanobacteria. Curr. Opin. Biotechnol. 2009, 20, 257–263. [Google Scholar] [CrossRef]
- Parmar, A.; Singh, N.K.; Pandey, A.; Gnansounou, E.; Madamwar, D. Cyanobacteria and microalgae: A positive prospect for biofuels. Bioresour. Technol. 2011, 102, 10163–10172. [Google Scholar] [CrossRef]
- Jones, C.S.; Mayfield, S.P. Algae biofuels: Versatility for the future of bioenergy. Curr. Opin. Biotechnol. 2012, 23, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Rodolfi, L.; Zittelli, G.C.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, G.; Tredici, M.R. Microalgae for oil: Strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112. [Google Scholar] [CrossRef]
- Lu, X. A perspective: Photosynthetic production of fatty acid-based biofuels in genetically engineered cyanobacteria. Biotechnol. Adv. 2010, 28, 742–746. [Google Scholar] [CrossRef]
- Quintana, N.; Van Der Kooy, F.; Van De Rhee, M.D.; Voshol, G.P.; Verpoorte, R. Renewable energy from cyanobacteria: Energy production optimization by metabolic pathway engineering. Appl. Microbiol. Biotechnol. 2011, 91, 471–490. [Google Scholar] [CrossRef]
- Rosgaard, L.; De Porcellinis, A.J.; Jacobsen, J.H.; Frigaard, N.-U.; Sakuragi, Y. Bioengineering of carbon fixation, biofuels, and biochemicals in cyanobacteria and plants. J. Biotechnol. 2012, 162, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Parkhey, P.; Joshi, K.; Mahilkar, A. Design of a microbial fuel cell and its transition to microbial electrolytic cell for hydrogen production by electro-hydro genesis. Indian J. Exp. Biol. 2013, 51, 860–865. [Google Scholar]
- Pruvost, J.; Van Vooren, G.; Le Gouic, B.; Couzinet-Mossion, A.; Legrand, J. Systematic investigation of biomass and lipid productivity by microalgae in photobioreactors for biodiesel application. Bioresour. Technol. 2011, 102, 150–158. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef]
- Mulders, K.J.M.; Lamers, P.P.; Martens, D.E.; Wijffels, R.H. Phototrophic pigment production with microalgae: Biological constraints and opportunities. J. Phycol. 2014, 50, 229–242. [Google Scholar] [CrossRef]
- González, L.E.; Díaz, G.C.; Aranda, D.A.G.; Cruz, Y.R.; Fortes, M.M. Biodiesel production based in microalgae: A biorefinery approach. Nat. Sci. 2015, 7, 358–369. [Google Scholar] [CrossRef]
- Yeesang, C.; Cheirsilp, B. Effect of nitrogen, salt, and iron content in the growth medium and light intensity on lipid production by microalgae isolated from freshwater sources in Thailand. Bioresour. Technol. 2011, 102, 3034–3040. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Yeh, K.-L.; Aisyah, R.; Lee, D.-J.; Chang, J.-S. Cultivation, photobioreactor design and harvesting of microalgae for biodiesel production: A critical review. Bioresour. Technol. 2011, 102, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Zhang, A.; Li, J.; He, B. A continuous process for biodiesel production in a fixed bed reactor packed with cation-exchange resin as heterogeneous catalyst. Bioresour. Technol. 2011, 102, 3607–3609. [Google Scholar] [CrossRef] [PubMed]
- Mairet, F.; Bernard, O.; Masci, P.; Lacour, T.; Sciandra, A. Modelling neutral lipid production by the microalga Isochrysis aff. galbana under nitrogen limitation. Bioresour. Technol. 2011, 102, 142–149. [Google Scholar] [CrossRef]
- Fuentes-Grünewald, C.; Garcés, E.; Alacid, E.; Sampedro, N.; Rossi, S.; Camp, J. Improvement of lipid production in the marine strains Alexandrium minutum and Heterosigma akashiwo by utilizing abiotic parameters. J. Ind. Microbiol. Biotechnol. 2011, 39, 207–216. [Google Scholar] [CrossRef]
- Sitther, V.; Tabatabai, B.; Fathabad, S.G.; Gichuki, S.; Chen, H.; Arumanayagam, A.C.S. Cyanobacteria as a biofuel source: Recent advances and applications. Adv. Cyanobacterial Biol. 2020, 269–289. [Google Scholar] [CrossRef]
- Tabatabai, B.; Arumanayagam, A.S.; Enitan, O.; Mani, A.; Natarajan, S.S.; Sitther, V. Overexpression of hlyB and mdh genes confers halotolerance in Fremyella diplosiphon, a freshwater cyanobacterium. Enzym. Microb. Technol. 2017, 103, 12–17. [Google Scholar] [CrossRef]
- Fathabad, S.G.; Arumanayagam, A.S.; Tabatabai, B.; Chen, H.; Lu, J.; Sitther, V. Augmenting Fremyella diplosiphon cellular lipid content and unsaturated fatty acid methyl esters via sterol Desaturase gene overexpression. Appl. Biochem. Biotechnol. 2019, 189, 1127–1140. [Google Scholar] [CrossRef]
- Tabatabai, B.; Chen, H.; Lu, J.; Giwa-Otusajo, J.; McKenna, A.M.; Shrivastava, A.K.; Sitther, V. Fremyella diplosiphon as a biodiesel agent: Identification of fatty acid methyl esters via microwave-assisted direct in situ Transesterification. BioEnergy Res. 2018, 11, 528–537. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Phys. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Rosenberg, J.N.; Kobayashi, N.; Barnes, A.; Noel, E.A.; Betenbaugh, M.J.; Oyler, G.A. Comparative analyses of three chlorella species in response to light and sugar reveal distinctive lipid accumulation patterns in the microalga C. sorokiniana. PLoS ONE 2014, 9, e92460. [Google Scholar] [CrossRef] [PubMed]
- Pinto, E.; Sigaud-Kutner, T.C.S.; Leitao, M.A.S.; Okamoto, O.K.; Morse, D.; Colepicolo, P. Heavy metal-induced oxidative stress in Algae1. J. Phycol. 2003, 39, 1008–1018. [Google Scholar] [CrossRef]
- Ip, P.-F.; Chen, F. Employment of reactive oxygen species to enhance astaxanthin formation in Chlorella zofingiensis in heterotrophic culture. Process. Biochem. 2005, 40, 3491–3496. [Google Scholar] [CrossRef]
- Sharma, K.K.; Schuhmann, H.; Schenk, P.M. High lipid induction in microalgae for biodiesel production. Energies 2012, 5, 1532–1553. [Google Scholar] [CrossRef]
- Illman, A.; Scragg, A.; Shales, S. Increase in Chlorella strains calorific values when grown in low nitrogen medium. Enzym. Microb. Technol. 2000, 27, 631–635. [Google Scholar] [CrossRef]
- Singh, S.C.; Sinha, R.P.; Hader, D.P. Role of lipids and fatty acids in stress tolerance in cyanobacteria. Acta Protozool. 2002, 41, 297–308. [Google Scholar]
- Hsieh, C.-H.; Wu, W.-T. Cultivation of microalgae for oil production with a cultivation strategy of urea limitation. Bioresour. Technol. 2009, 100, 3921–3926. [Google Scholar] [CrossRef]
- Yeh, K.-L.; Chang, J.-S. Nitrogen starvation strategies and photobioreactor design for enhancing lipid content and lipid production of a newly isolated microalga Chlorella vulgaris ESP-31: Implications for biofuels. Biotechnol. J. 2011, 6, 1358–1366. [Google Scholar] [CrossRef]
- Sun, X.; Cao, Y.; Xu, H.; Liu, Y.; Sun, J.; Qiao, D.R.; Cao, Y. Effect of nitrogen-starvation, light intensity and iron on triacylglyceride/carbohydrate production and fatty acid profile of Neochloris oleoabundans HK-129 by a two-stage process. Bioresour. Technol. 2014, 155, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Mallick, N. Microalga Scenedesmus obliquus as a potential source for biodiesel production. Appl. Microbiol. Biotechnol. 2009, 84, 281–291. [Google Scholar] [CrossRef]
- Nigam, S.; Rai, M.P.; Sharma, R. Effect of nitrogen on growth and lipid content of Chlorella pyrenoidosa. Am. J. Biochem. Biotechnol. 2011, 7, 124–129. [Google Scholar] [CrossRef]
- Li, Y.; Horsman, M.; Wu, N.; Lan, C.Q.; Dubois-Calero, N. Biofuels from microalgae. Biotechnol. Prog. 2008, 24, 815–820. [Google Scholar] [CrossRef] [PubMed]
- Suen, Y.; Hubbard, J.S.; Holzer, G.; Tornabene, T.G. Total lipid production of the green alga Nannochloropsis sp. QII under different nitrogen regimes. J. Phycol. 1987, 23, 289–296. [Google Scholar] [CrossRef]
- Stephenson, A.I.; Dennis, J.S.; Howe, C.J.; Scott, S.A.; Smith, A.G. Influence of nitrogen limitation regime on the production by Chlorella vulgaris of lipids for biodiesel feedstocks. Biofuels 2010, 1, 47–58. [Google Scholar] [CrossRef]
- Taher, H.; Al-Zuhair, S.; Al-Marzouqi, A.H.; Haik, Y.; Farid, M. Effective extraction of microalgae lipids from wet biomass for biodiesel production. Biomass Bioenergy 2014, 66, 159–167. [Google Scholar] [CrossRef]
- Scott, S.; Davey, M.P.; Dennis, J.S.; Horst, I.; Howe, C.J.; Lea-Smith, D.J.; Smith, A.G. Biodiesel from algae: Challenges and prospects. Curr. Opin. Biotechnol. 2010, 21, 277–286. [Google Scholar] [CrossRef]
- Becker, E.W. Microalgae: Biotechnology and Microbiology; Cambridge University Press: Cambridge, UK, 1994; ISBN 0-521-35020-4. [Google Scholar]
- Nagarkar, S.; Williams, G.A.; Subramanian, G.; Saha, S. Cyanobacteria-dominated biofilms: A high quality food resource for intertidal grazers. Hydrobiologia 2004, 512, 89–95. [Google Scholar] [CrossRef]
- Wahlen, B.D.; Morgan, M.R.; McCurdy, A.T.; Willis, R.M.; Dye, D.J.; Bugbee, B.; Wood, B.D.; Seefeldt, L.C. Biodiesel from microalgae, yeast, and bacteria: Engine performance and exhaust emissions. Energy Fuels 2012, 27, 220–228. [Google Scholar] [CrossRef]
- Tang, H.; Chen, M.; Garcia, M.; Abunasser, N.; Ng, K.S.; Salley, S.O. Culture of microalgae Chlorella minutissima for biodiesel feedstock production. Biotechnol. Bioeng. 2011, 108, 2280–2287. [Google Scholar] [CrossRef]
- Rittmann, B.E. Opportunities for renewable bioenergy using microorganisms. Biotechnol. Bioeng. 2008, 100, 203–212. [Google Scholar] [CrossRef] [PubMed]
- D’Oca, M.G.M.; Viêgas, C.V.; Lemões, J.S.; Miyasaki, E.K.; Morón-Villarreyes, J.A.; Primel, E.G.; Abreu, P.C. Production of FAMEs from several microalgal lipidic extracts and direct transesterification of the Chlorella pyrenoidosa. Biomass Bioenergy 2011, 35, 1533–1538. [Google Scholar] [CrossRef]
- Rai, M.P.; Gautom, T.; Sharma, N. Effect of salinity, pH, light intensity on growth and lipid production of microalgae for bioenergy application. Online J. Biol. Sci. 2015, 15, 260–267. [Google Scholar] [CrossRef]
- Moradi-Kheibari, N.; Ahmadzadeh, H.; Hosseini, M. Use of solvent mixtures for total lipid extraction of Chlorella vulgaris and gas chromatography FAME analysis. Bioprocess Biosyst. Eng. 2017, 40, 1363–1373. [Google Scholar] [CrossRef]
- Bowen, D. Effect of Fatty Acid Structure on Biodiesel. Biodiesel is Good. 2010. Available online: http://biodieselisgood.org/chemistry/effect-of-fatty-acid-structure-on-biodiesel/ (accessed on 3 November 2020).
- Knothe, G. Dependence of biodiesel fuel properties on the structure of fatty acid alkyl esters. Fuel Process. Technol. 2005, 86, 1059–1070. [Google Scholar] [CrossRef]
- Knothe, G. “Designer” biodiesel: Optimizing fatty ester composition to improve fuel properties. Energy Fuels 2008, 22, 1358–1364. [Google Scholar] [CrossRef]
- Griffiths, M.J.; Harrison, S.T. Lipid productivity as a key characteristic for choosing algal species for biodiesel production. Environ. Biol. Fishes 2009, 21, 493–507. [Google Scholar] [CrossRef]
- Wang, Q.; Sen, B.; Liu, X.; He, Y.; Xie, Y.; Wang, G. Enhanced saturated fatty acids accumulation in cultures of newly-isolated strains of Schizochytrium sp. and Thraustochytriidae sp. for large-scale biodiesel production. Sci. Total Environ. 2018, 994–1004. [Google Scholar] [CrossRef]
- Young, E.B.; Beardall, J. Photosynthetic function in Dunaliella tertiolecta (chlorophyta) during a nitrogen starvation and recovery cycle. J. Phycol. 2003, 39, 897–905. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 7-Day | NaNO3 Concentration | |||
|---|---|---|---|---|
| Fatty Acid Methyl Esters | 0.25 g L−1 | 0.5 g L−1 | 1.0 g L−1 | 1.5 g L−1 |
| methyl dodecanoate (12:0) | 2.28 ± 0.618 | 1.25 ± 0.136 | 1.25 ± 0.439 | 3.53 ± 1.55 |
| methyl myristate (14:0) | 6.7 ± 0.861 | 1.61 ± 0.599 | 2.59 ± 1.199 | 1.97 ± 0.651 |
| methyl palmitate (16:0) | 63.38 ± 0.809 | 67.59 ± 8.78 | 66.67 ± 9.46 | 70.03 ± 3.90 |
| methyl hexadecanoate (16:1) | 3.72 ± 1.21 | 1.06 ± 0.726 | 3.92 ± 0.415 | 1.62 ± 0.305 |
| methyl octadecanoate (18:0) | 3.21 ± 1.45 | 0.75 ± 0.370 | 1.60 ± 0.693 | 2.45 ± 1.35 |
| methyl octadecenoate (18:1) | 18.46 ± 1.82 | 18.19 ± 6.11 | 22.81 ± 9.04 | 16.80 ± 3.98 |
| methyl octadecadienoate (18:2) | 2.23 ± 1.16 | 2.41 ± 0.916 | 1.15 ± 0.567 | 3.60 ± 0.940 |
| 10-Day | ||||
| methyl dodecanoate (12:0) | 1.78 ± 0.204 | 1.76 ± 0.224 | 1.90 ± 0.969 | 0.86 ± 0.193 |
| methyl myristate (14:0) | 0.84 ± 0.444 | 1.07 ± 0.662 | 1.90 ± 0.910 | 2.14 ± 1.11 |
| methyl palmitate (16:0) | 58.45 ± 2.13 | 62.45 ± 5.99 | 65.49 ± 3.63 | 56.71 ± 1.59 |
| methyl hexadecanoate (16:1) | 2.17 ± 1.01 | 1.75 ± 0.332 | 2.90 ± 0.693 | 1.36 ± 0.047 |
| methyl octadecanoate (18:0) | 4.19 ± 1.35 | 2.07 ± 0.464 | 1.82 ± 0.754 | 1.92 ± 0.872 |
| methyl octadecenoate (18:1) | 27.51 ± 5.30 | 25.47 ± 3.52 | 21.02 ± 5.60 | 28.97 ± 0.623 |
| methyl octadecadienoate (18:2) | 5.06 ± 0.539 | 5.44 ± 2.02 | 4.98 ± 0.544 | 8.04 ± 0.640 |
| 15-Day | ||||
| methyl dodecanoate (12:0) | 1.44 ± 0.400 | 1.39 ± 0.278 | 1.55 ± 0.086 | 1.94 ± 0.202 |
| methyl myristate (14:0) | 0.76 ± 0.374 | 0.50 ± 0.117 | 0.63 ± 0.063 | 0.86 ± 0.108 |
| methyl palmitate (16:0) | 57.39 ± 5.25 | 62.14 ± 1.97 | 62.29 ± 0.523 | 65.80 ± 1.30 |
| methyl hexadecanoate (16:1) | 2.74 ± 0.957 | 7.46 ± 2.34 | 4.46 ± 2.52 | 4.92 ± 2.19 |
| methyl octadecanoate (18:0) | 1.75 ± 1.40 | 1.50 ± 0.931 | 2.44 ± 0.638 | 1.27 ± 0.170 |
| methyl octadecenoate (18:1) | 28.19 ± 5.135 | 21.15 ± 2.83 | 20.02 ± 1.61 | 18.62 ± 1.95 |
| methyl octadecadienoate (18:2) | 7.73 ± 0.815 | 5.85 ± 1.83 | 8.61 ± 0.924 | 6.59 ± 0.213 |
| 20-Day | ||||
| methyl dodecanoate (12:0) | 2.20 ± 1.61 | 2.83 ± 2.30 | 2.13 ± 0.958 | 2.41 ± 0.496 |
| methyl myristate (14:0) | 3.56 ± 2.45 | 8.13 ± 1.19 | 3.34 ± 1.65 | 3.69 ± 2.82 |
| methyl palmitate (16:0) | 60.71 ± 6.23 | 50.85 ± 6.11 | 50.25 ± 11.9 | 64.58 ± 4.13 |
| methyl hexadecanoate (16:1) | 2.24 ± 1.34 | 3.11 ± 1.65 | 4.94 ± 2.71 | 1.60 ± 0.472 |
| methyl octadecanoate (18:0) | 1.90 ± 0.923 | 3.60 ± 1.41 | 4.07 ± 1.41 | 2.31 ± 1.27 |
| methyl octadecenoate (18:1) | 25.03 ± 6.71 | 27.79 ± 4.91 | 30.88 ± 9.21 | 23.18 ± 5.26 |
| methyl octadecadienoate (18:2) | 4.36 ± 0.860 | 3.69 ± 2.54 | 4.39 ± 1.33 | 2.24 ± 1.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tabatabai, B.; Adusei, A.; Shrivastava, A.K.; Singh, P.K.; Sitther, V. Nitrogen Deprivation in Fremyella diplosiphon Augments Lipid Production without Affecting Growth. Energies 2020, 13, 5769. https://doi.org/10.3390/en13215769
Tabatabai B, Adusei A, Shrivastava AK, Singh PK, Sitther V. Nitrogen Deprivation in Fremyella diplosiphon Augments Lipid Production without Affecting Growth. Energies. 2020; 13(21):5769. https://doi.org/10.3390/en13215769
Chicago/Turabian StyleTabatabai, Behnam, Afua Adusei, Alok Kumar Shrivastava, Prashant Kumar Singh, and Viji Sitther. 2020. "Nitrogen Deprivation in Fremyella diplosiphon Augments Lipid Production without Affecting Growth" Energies 13, no. 21: 5769. https://doi.org/10.3390/en13215769
APA StyleTabatabai, B., Adusei, A., Shrivastava, A. K., Singh, P. K., & Sitther, V. (2020). Nitrogen Deprivation in Fremyella diplosiphon Augments Lipid Production without Affecting Growth. Energies, 13(21), 5769. https://doi.org/10.3390/en13215769