A Critical Analysis of Bio-Hydrocarbon Production in Bacteria: Current Challenges and Future Directions


Abstract
1. Introduction
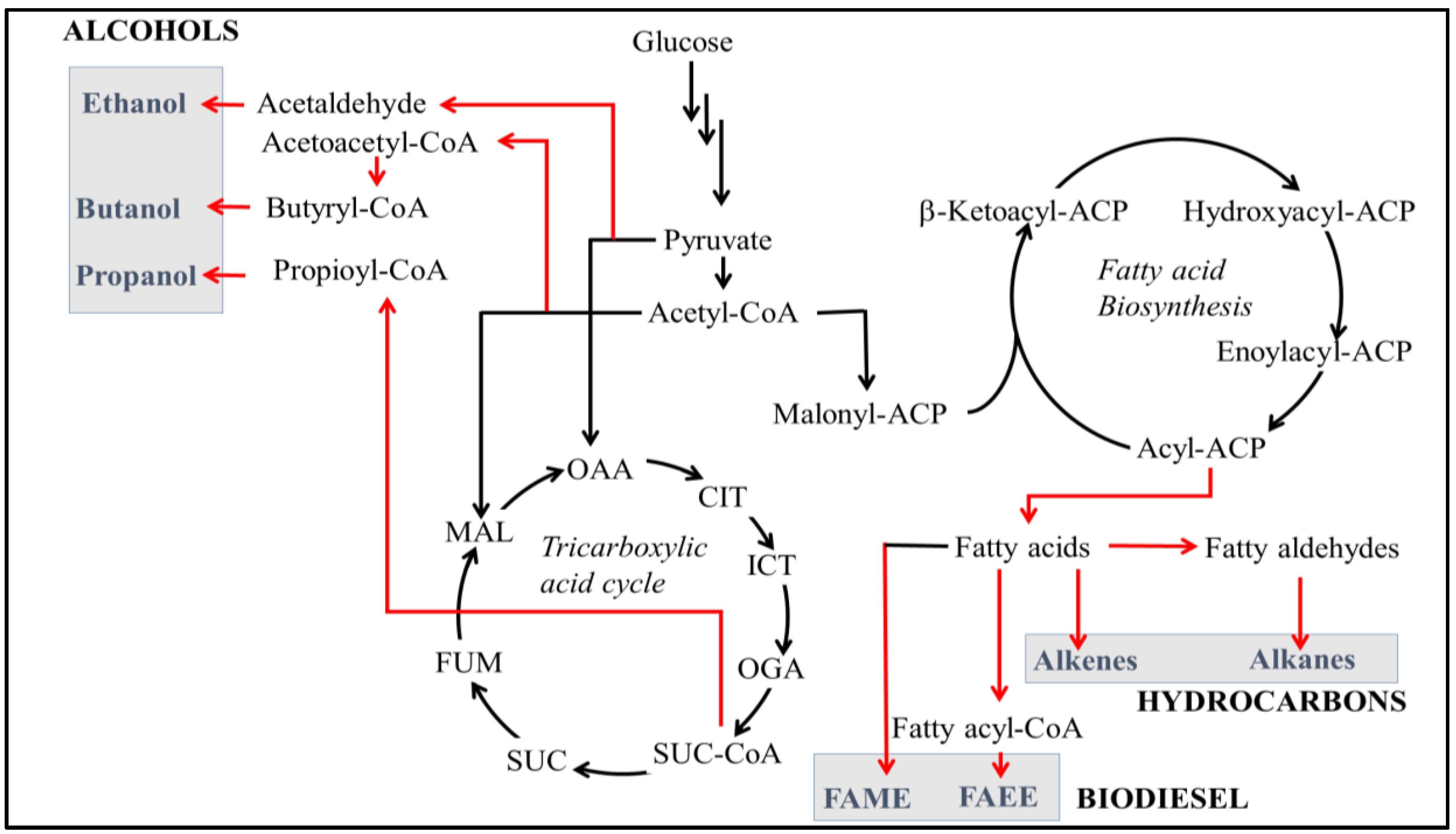
2. Engineering Metabolic Pathways for Hydrocarbon Production in Bacteria
2.1. Alcohols Production in Bacteria
2.2. Alkanes Production in Bacteria
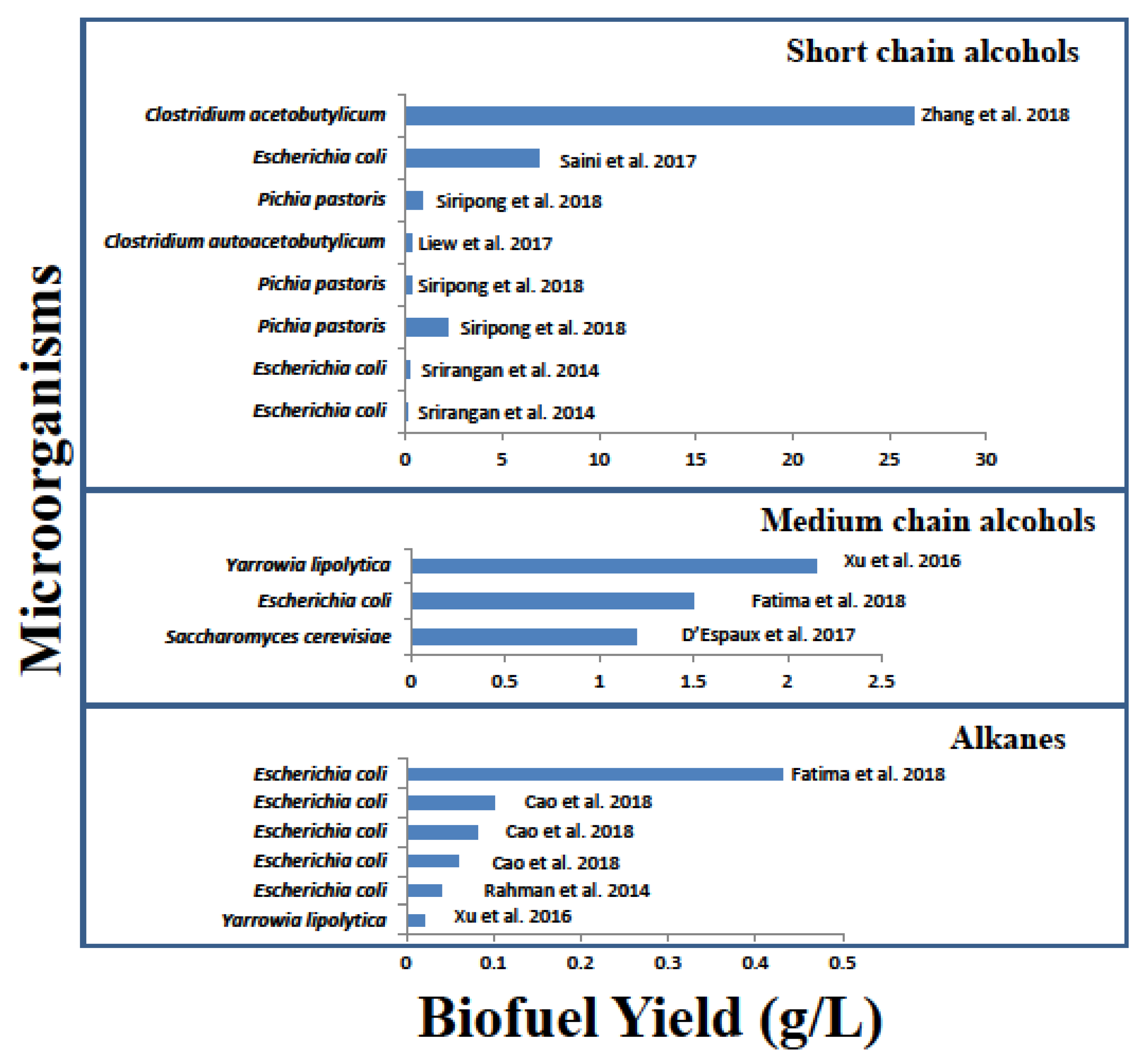
3. Comparative Analysis of Bio-Hydrocarbons Production in Bacteria
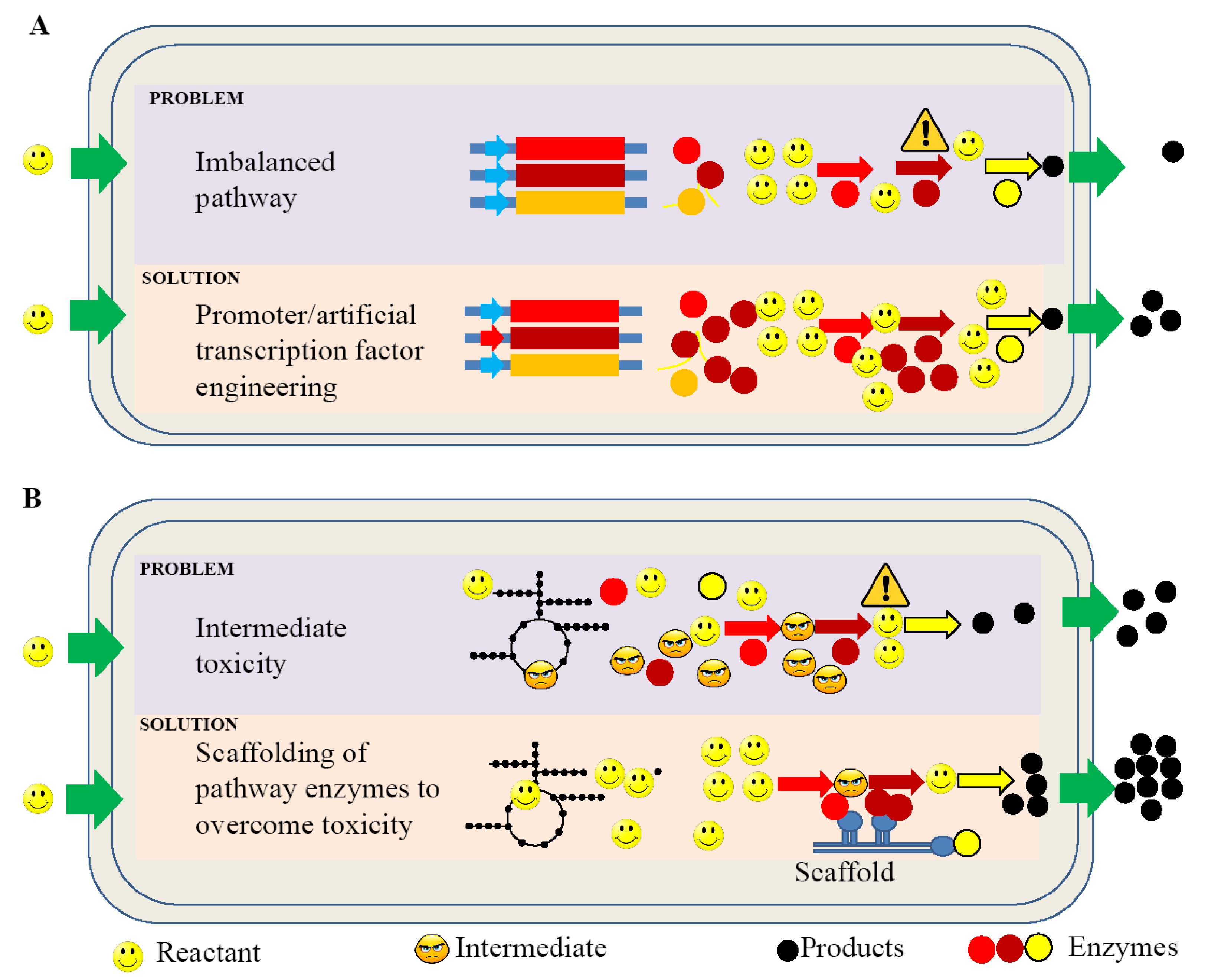
4. Current Challenges in Improving Hydrocarbon Production
4.1. Promoters
4.2. Gene Copy Number
4.3. Artificial Transcription Factor (ATF) Engineering
4.4. Ribosome Engineering
4.5. Transfer RNA Engineering
4.6. Cofactor Regeneration
4.7. Scaffold System
5. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goldthau, A. The G20 must govern the shift to low-carbon energy. Nature 2017, 546, 203–205. [Google Scholar] [CrossRef] [PubMed]
- Balch, J.K.; Nagy, R.C.; Archibald, S.; Bowman, D.M.; Moritz, M.A.; Roos, C.I.; Scott, A.C.; Williamson, G.J. Global combustion: The connection between fossil fuel and biomass burning emissions (1997–2010). Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371. [Google Scholar] [CrossRef] [PubMed]
- Den, W.; Sharma, V.K.; Lee, M.; Nadadur, G.; Varma, R.S. Lignocellulosic biomass transformations via greener oxidative pretreatment processes: Access to energy and value-added chemicals. Front. Chem. 2018, 6, 141. [Google Scholar] [CrossRef] [PubMed]
- Nejat, P.; Jomehzadeh, F.; Taheri, M.M.; Gohari, M.; Majid, M.Z.A. A global review of energy consumption, CO2 emissions and policy in the residential sector (with an overview of the top ten CO2 emitting countries). Renew. Sustain. Energy Rev. 2015, 43, 843–862. [Google Scholar] [CrossRef]
- Shields-Menard, S.A.; Amirsadeghi, M.; French, W.T.; Boopathy, R. A review on microbial lipids as a potential biofuel. Bioresour. Technol. 2018, 259, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Dresselhaus, M.; Thomas, I. Alternative energy technologies. Nature 2001, 414, 332. [Google Scholar] [CrossRef] [PubMed]
- d’Espaux, L.; Ghosh, A.; Runguphan, W.; Wehrs, M.; Xu, F.; Konzock, O.; Dev, I.; Nhan, M.; Gin, J.; Reider Apel, A.; et al. Engineering high-level production of fatty alcohols by Saccharomyces cerevisiae from lignocellulosic feedstocks. Metab. Eng. 2017, 42, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Fatma, Z.; Hartman, H.; Poolman, M.G.; Fell, D.A.; Srivastava, S.; Shakeel, T.; Yazdani, S.S. Model-assisted metabolic engineering of Escherichia coli for long chain alkane and alcohol production. Metab. Eng. 2018, 46, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Liew, F.; Henstra, A.M.; Kpke, M.; Winzer, K.; Simpson, S.D.; Minton, N.P. Metabolic engineering of Clostridium autoethanogenum for selective alcohol production. Metab. Eng. 2017, 40, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Demirbas, A. Biofuels sources, biofuel policy, biofuel economy and global biofuel projections. Energy Convers. Manag. 2008, 49, 2106–2116. [Google Scholar] [CrossRef]
- Pandey, R.K.; Tewari, L. Mycotechnology for lignocellulosic bioethanol production: An emerging approach to sustainable environment. In Microbial Biotechnology in Environmental Monitoring and Cleanup; IGI Glob: Hershey, PA, USA, 2018; pp. 28–43. [Google Scholar] [CrossRef]
- Babu, V.; Murthy, M. Butanol and pentanol: The promising biofuels for CI engines–A review. Renew. Sustain. Energy Rev. 2017, 78, 1068–1088. [Google Scholar]
- No, S.-Y. Application of hydrotreated vegetable oil from triglyceride based biomass to CI engines—A review. Fuel 2014, 115, 88–96. [Google Scholar] [CrossRef]
- Van Vliet, O.P.; Faaij, A.P.; Turkenburg, W.C. Fischer—Tropsch diesel production in a well-to-wheel perspective: A carbon, energy flow and cost analysis. Energy Convers. Manag. 2009, 50, 855–876. [Google Scholar] [CrossRef]
- Ohta, K.; Beall, D.S.; Mejia, J.P.; Shanmugam, K.T.; Ingram, L.O. Genetic improvement of Escherichia coli for ethanol production: Chromosomal integration of Zymomonas mobilis genes encoding pyruvate decarboxylase and alcohol dehydrogenase ii. Appl. Environ. Microbiol. 1991, 57, 893–900. [Google Scholar] [PubMed]
- Rahman, Z.; Rashid, N.; Nawab, J.; Ilyas, M.; Sung, B.H.; Kim, S.C. Escherichia coli as a fatty acid and biodiesel factory: Current challenges and future directions. Environ. Sci. Pollut. Res. Int. 2016, 23, 12007–12018. [Google Scholar] [CrossRef] [PubMed]
- Lu, X. A perspective: Photosynthetic production of fatty acid-based biofuels in genetically engineered Cyanobacteria. Biotechnol. Adv. 2010, 28, 742–746. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.J.; Buijs, N.A.; Siewers, V.; Nielsen, J. Fatty acid-derived biofuels and chemicals production in Saccharomyces cerevisiae. Front Bioeng. Biotechnol. 2014, 2, 32. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.L.; Blaschek, H.P. Butanol production by a butanol-tolerant strain of Clostridium acetobutylicum in extruded corn broth. Appl. Environ. Microbiol. 1983, 45, 966–973. [Google Scholar] [PubMed]
- Xu, P.; Qiao, K.; Ahn, W.S.; Stephanopoulos, G. Engineering Yarrowia lipolytica as a platform for synthesis of drop-in transportation fuels and oleochemicals. Proc. Natl. Acad. Sci. USA 2016, 113, 10848–10853. [Google Scholar] [CrossRef] [PubMed]
- Atsumi, S.; Cann, A.F.; Connor, M.R.; Shen, C.R.; Smith, K.M.; Brynildsen, M.P.; Chou, K.J.; Hanai, T.; Liao, J.C. Metabolic engineering of Escherichia coli for 1-butanol production. Metab. Eng. 2008, 10, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Rutter, C.D.; Rao, C.V. Production of 1-decanol by metabolically engineered Yarrowia lipolytica. Metab. Eng. 2016, 38, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Elgaali, H.; Hamilton-Kemp, T.; Newman, M.; Collins, R.; Yu, K.; Archbold, D. Comparison of long-chain alcohols and other volatile compounds emitted from food-borne and related gram positive and gram negative bacteria. J. Basic Microbiol. 2002, 42, 373–380. [Google Scholar] [CrossRef]
- Kohse-Hoinghaus, K.; Osswald, P.; Cool, T.A.; Kasper, T.; Hansen, N.; Qi, F.; Westbrook, C.K.; Westmoreland, P.R. Biofuel combustion chemistry: From ethanol to biodiesel. Angew. Chem. Int. Ed. Engl. 2010, 49, 3572–3597. [Google Scholar] [CrossRef] [PubMed]
- Siripong, W.; Wolf, P.; Kusumoputri, T.P.; Downes, J.J.; Kocharin, K.; Tanapongpipat, S.; Runguphan, W. Metabolic engineering of Pichia pastoris for production of isobutanol and isobutyl acetate. Biotechnol. Biofuels. 2018, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Saini, M.; Wang, Z.W.; Chiang, C.J.; Chao, Y.P. Metabolic engineering of Escherichia coli for production of n-butanol from crude glycerol. Biotechnol. Biofuels. 2017, 10, 173. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zong, W.; Hong, W.; Zhang, Z.T.; Wang, Y. Exploiting endogenous crispr-cas system for multiplex genome editing in Clostridium tyrobutyricum and engineer the strain for high-level butanol production. Metab. Eng. 2018, 47, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Rahman, Z.; Sung, B.H.; Yi, J.Y.; Bui le, M.; Lee, J.H.; Kim, S.C. Enhanced production of n-alkanes in Escherichia coli by spatial organization of biosynthetic pathway enzymes. J. Biotechnol. 2014, 192 Pt A, 187–191. [Google Scholar] [CrossRef]
- Srirangan, K.; Liu, X.; Westbrook, A.; Akawi, L.; Pyne, M.E.; Moo-Young, M.; Chou, C.P. Biochemical, genetic, and metabolic engineering strategies to enhance coproduction of 1-propanol and ethanol in engineered Escherichia coli. Appl. Microbiol. Biotechnol. 2014, 98, 9499–9515. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.X.; Xiao, W.H.; Zhang, J.L.; Xie, Z.X.; Ding, M.Z.; Yuan, Y.J. Heterologous biosynthesis and manipulation of alkanes in Escherichia coli. Metab. Eng. 2016, 38, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Garcia, H.G.; Brewster, R.C.; Phillips, R. Using synthetic biology to make cells tomorrow’s test tubes. Integr. Biol. 2016, 8, 431–450. [Google Scholar] [CrossRef] [PubMed]
- Smanski, M.J.; Zhou, H.; Claesen, J.; Shen, B.; Fischbach, M.A.; Voigt, C.A. Synthetic biology to access and expand nature’s chemical diversity. Nat. Rev. Microbiol. 2016, 14, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Yan, Q.; Jones, J.A.; Tang, Y.J.; Fong, S.S.; Koffas, M.A.G. Metabolic burden: Cornerstones in synthetic biology and metabolic engineering applications. Trends Biotechnol. 2016, 34, 652–664. [Google Scholar] [CrossRef] [PubMed]
- Jensen, P.R.; Hammer, K. Artificial promoters for metabolic optimization. Biotechnol. Bioeng. 1998, 58, 191–195. [Google Scholar] [CrossRef]
- Polli, F.; Meijrink, B.; Bovenberg, R.A.L.; Driessen, A.J.M. New promoters for strain engineering of Penicillium chrysogenum. Fungal. Genet. Biol. 2016, 89, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Lutz, R.; Bujard, H. Independent and tight regulation of transcriptional units in Escherichia coli via the lacr/o, the tetr/o and arac/i1-i2 regulatory elements. Nucleic. Acids. Res. 1997, 25, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Ho, K.M.; Lim, B.L. Co-expression of a prophage system and a plasmid system in Bacillus subtilis. Protein Expr. Purif. 2003, 32, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Dehli, T.; Solem, C.; Jensen, P.R. Tunable promoters in synthetic and systems biology. Subcell. Biochem. 2012, 64, 181–201. [Google Scholar] [PubMed]
- Song, Y.; Nikoloff, J.M.; Fu, G.; Chen, J.; Li, Q.; Xie, N.; Zheng, P.; Sun, J.; Zhang, D. Promoter screening from Bacillus subtilis in various conditions hunting for synthetic biology and industrial applications. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Rudge, T.J.; Brown, J.R.; Federici, F.; Dalchau, N.; Phillips, A.; Ajioka, J.W.; Haseloff, J. Characterization of intrinsic properties of promoters. ACS Synth. Biol. 2016, 5, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Stracquadanio, G.; Luo, J.; Boeke, J.D.; Bader, J.S. Biopartsbuilder: A synthetic biology tool for combinatorial assembly of biological parts. Bioinformatics 2016, 32, 937–939. [Google Scholar] [CrossRef] [PubMed]
- Stewart, G.S.; Lubinsky-Mink, S.; Jackson, C.G.; Cassel, A.; Kuhn, J. Phg165: A PBR322 copy number derivative of puc8 for cloning and expression. Plasmid 1986, 15, 172–181. [Google Scholar] [CrossRef]
- Atkinson, M.R.; Savageau, M.A.; Myers, J.T.; Ninfa, A.J. Development of genetic circuitry exhibiting toggle switch or oscillatory behavior in Escherichia coli. Cell 2003, 113, 597–607. [Google Scholar] [CrossRef]
- Lian, J.; Jin, R.; Zhao, H. Construction of plasmids with tunable copy numbers in Saccharomyces cerevisiae and their applications in pathway optimization and multiplex genome integration. Biotechnol. Bioeng. 2016, 113, 2462–2473. [Google Scholar] [CrossRef] [PubMed]
- Shizuya, H.; Birren, B.; Kim, U.J.; Mancino, V.; Slepak, T.; Tachiiri, Y.; Simon, M. Cloning and stable maintenance of 300-kilobase-pair fragments of human DNA in Escherichia coli using an f-factor-based vector. Proc. Natl. Acad. Sci. USA 1992, 89, 8794–8797. [Google Scholar] [CrossRef] [PubMed]
- Kasai, Y.; Oshima, K.; Ikeda, F.; Abe, J.; Yoshimitsu, Y.; Harayama, S. Construction of a self-cloning system in the unicellular green alga Pseudochoricystis ellipsoidea. Biotechnol. Biofuels. 2015, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Sera, T. Zinc-finger-based artificial transcription factors and their applications. Adv. Drug. Deliv. Rev. 2009, 61, 513–526. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Sung, B.H.; Yu, B.J.; Lee, J.H.; Lee, S.H.; Kim, M.S.; Koob, M.D.; Kim, S.C. Phenotypic engineering by reprogramming gene transcription using novel artificial transcription factors in Escherichia coli. Nucleic. Acids. Res. 2008, 36. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Yang, K.S.; Jang, S.A.; Sung, B.H.; Kim, S.C. Engineering butanol-tolerance in Escherichia coli with artificial transcription factor libraries. Biotechnol. Bioeng. 2011, 108, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Wei, X.; Sun, C.; Zhang, F.; Xu, J.; Zhao, X.; Bai, F. Improvement of acetic acid tolerance of Saccharomyces cerevisiae using a zinc-finger-based artificial transcription factor and identification of novel genes involved in acetic acid tolerance. Appl. Microbiol. Biotechnol. 2015, 99, 2441–2449. [Google Scholar] [CrossRef] [PubMed]
- Ochi, K. From microbial differentiation to ribosome engineering. Biosci. Biotechnol. Biochem. 2007, 71, 1373–1386. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.N.; Stephanopoulos, G. Combinatorial engineering of microbes for optimizing cellular phenotype. Curr. Opin. Chem. Biol. 2008, 12, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Kasahara, K.; Izawa, M.; Ochi, K. Applicability of ribosome engineering to vitamin B12 production by Propionibacterium shermanii. Biosci. Biotechnol. Biochem. 2017, 81, 1636–1641. [Google Scholar] [CrossRef] [PubMed]
- Kurosawa, K.; Hosaka, T.; Tamehiro, N.; Inaoka, T.; Ochi, K. Improvement of alpha-amylase production by modulation of ribosomal component protein S12 in Bacillus subtilis 168. Appl. Environ. Microbiol. 2006, 72, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Hosaka, T.; Xu, J.; Ochi, K. Increased expression of ribosome recycling factor is responsible for the enhanced protein synthesis during the late growth phase in an antibiotic-overproducing Streptomyces coelicolor ribosomal rpsl mutant. Mol. Microbiol. 2006, 61, 883–897. [Google Scholar] [CrossRef] [PubMed]
- Janosi, L.; Shimizu, I.; Kaji, A. Ribosome recycling factor (ribosome releasing factor) is essential for bacterial growth. Proc. Natl. Acad. Sci. USA 1994, 91, 4249–4253. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cai, L.; Fang, Z.; Tian, H.; Gao, X.; Yao, W. Site-specific incorporation of unnatural amino acids into urate oxidase in Escherichia coli. Protein Sci. 2008, 17, 1827–1833. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, K.J.; Gite, S. Trna-mediated protein engineering. Curr. Opin. Biotechnol. 1999, 10, 64–70. [Google Scholar] [CrossRef]
- Gladyshev, V.N.; Jeang, K.T.; Stadtman, T.C. Selenocysteine, identified as the penultimate c-terminal residue in human t-cell thioredoxin reductase, corresponds to tga in the human placental gene. Proc. Natl. Acad. Sci. USA 1996, 93, 6146–6151. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, T.; Ishii, R.; Fukunaga, R.; Kobayashi, T.; Sakamoto, K.; Yokoyama, S. Multistep engineering of pyrrolysyl-trna synthetase to genetically encode n(epsilon)-(o-azidobenzyloxycarbonyl) lysine for site-specific protein modification. Chem. Biol. 2008, 15, 1187–1197. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.A.; Lo, S.L.; Krzycki, J.A. Clustered genes encoding the methyltransferases of methanogenesis from monomethylamine. J. Bacteriol. 1998, 180, 3432–3440. [Google Scholar] [PubMed]
- Hara, K.Y.; Shimodate, N.; Hirokawa, Y.; Ito, M.; Baba, T.; Mori, H.; Mori, H. Glutathione production by efficient atp-regenerating Escherichia coli mutants. FEMS Microbiol. Lett. 2009, 297, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, L.; Ma, C.; Gao, C.; Tao, F.; Xu, P. Engineering of cofactor regeneration enhances (2S,3S)-2,3-butanediol production from diacetyl. Sci. Rep. 2013, 3, 2643. [Google Scholar] [CrossRef] [PubMed]
- Geier, M.; Brandner, C.; Strohmeier, G.A.; Hall, M.; Hartner, F.S.; Glieder, A. Engineering Pichia pastoris for improved NADH regeneration: A novel chassis strain for whole-cell catalysis. Beilstein. J. Org. Chem. 2015, 11, 1741–1748. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Tsuchiya, D.; Oyama, T.; Tsunaka, Y.; Morikawa, K. Structural basis for channelling mechanism of a fatty acid beta-oxidation multienzyme complex. EMBO J. 2004, 23, 2745–2754. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, K.; Rasmussen, A.V.; Morant, M.; Nielsen, A.H.; Bjarnholt, N.; Zagrobelny, M.; Bak, S.; Moller, B.L. Metabolon formation and metabolic channeling in the biosynthesis of plant natural products. Curr. Opin. Plant. Biol. 2005, 8, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, B.A.; Khosla, C. Biosynthesis of polyketides in heterologous hosts. Microbiol. Mol. Biol. Rev. 2001, 65, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.K.; Dandapani, H.; Thiel, K.; Jones, P.R. Microbial production of 1-octanol: A naturally excreted biofuel with diesel-like properties. Metab. Eng. Commun. 2015, 2, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.J.; Pitera, D.J.; Withers, S.T.; Newman, J.D.; Keasling, J.D. Engineering a mevalonate pathway in Escherichia coli for production of terpenoids. Nat. Biotechnol. 2003, 21, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Chen, X.; Li, P. Enhancing microbial production of biofuels by expanding microbial metabolic pathways. Biotechnol. Appl. Biochem. 2017, 64, 606–619. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.A.; Toparlak, O.D.; Koffas, M.A. Metabolic pathway balancing and its role in the production of biofuels and chemicals. Curr. Opin. Biotechnol. 2015, 33, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Peacock, A.F. Recent advances in designed coiled coils and helical bundles with inorganic prosthetic groups-from structural to functional applications. Curr. Opin. Chem. Biol. 2016, 31, 160–165. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Biofuel | Substrate | Microorganism | Metabolic Engineering | Yield (g/L) | References |
|---|---|---|---|---|---|
| Ethanol | Glycerol | Escherichia coli | Heterologous expression of sleeping beauty mutase operon | 0.20 a, 31 b | [29] |
| CO2 | Clostridium autoacetobutylicum | Allelic mutagenesis of aldehyde dehydrogenase (adhE) and expression of aldehyde ferredoxin reductase (AOR) | 0.28 a | [9] | |
| Propanol | Glycerol | Escherichia coli | Heterologous expression of sleeping beauty mutase operon | 0.15 a, 7 b | [29] |
| Butanol | CO2 | Clostridium acetobutylicum | Targeted deletion of sopA and cat1 (butyrate production) and integration of adhE gene by CRISPR-Cas9 | 26.2 a | [27] |
| Glycerol | Escherichia coli | Enhanced glycolytic flux, prevention of glycerol breakdown | 6.9 a | [26] | |
| 2-keto isovalerate | Pichia pastoris | Expression of keto acid degradation pathway | 0.28 a | [25] | |
| Glucose | Pichia pastoris | Overexpression of endogenous L-Valine biosynthesis pathway | 0.89 a | [25] | |
| Pichia pastoris | Fine tuning of expression levels using episomal plasmids | 2.22 a | [25] | ||
| Fatty alcohols | Glucose | Escherichia coli | In silico modelling of metabolic pathways for alkane production to remove bottlenecks identified in pentose phosphate pathway | 1.5 a, 12.5 b | [8] |
| Glucose | Yarrowia lipolytica | Expression of alkane-producing pathway, consisting of aldehyde reductase (AAR) and aldehyde deformylating oxygenase (FAD) | 2.15 a | [20] | |
| Lignocellulose | Saccharomyces cerevisiae | Expression of alkane-producing pathway Bottlenecks were identified by flux balance analysis and bacterial fatty aldehyde reductases (FARs) were replaced with mouse FARs | 1.2 a, 6 b | [7] | |
| Alkanes | Glucose | Escherichia coli | Heterologous expression of alkane-producing pathway (AAR & aldehyde-deformylating oxygenase (ADO)) from cyanobacteria, with no further modification | 0.06 a | [30] |
| Escherichia coli | Expression of alkane-producing pathway Deletion of adhE (endogenous reductase) | 0.08 a | [30] | ||
| Escherichia coli | Expression of alkane-producing pathway and modification of electron transfer system | 0.1 a, 1.3 b | [30] | ||
| Escherichia coli | Expression of alkane-producing pathway and DNA scaffolding of cyanobacterial AAR & ADO | 0.04 a | [28] | ||
| Yarrowia lipolytica | Expression of alkane-producing pathway (FAR & FAD), with no further modification | 0.02 a | [20] | ||
| Escherichia coli | In silico modelling of metabolic pathways for alkane production to help remove bottlenecks in pentose phosphate pathway | 0.43 a, 2.5 b | [8] |
| Biofuel | Microorganism | Experimental Conditions | Yield (g/L) | References | |||||
|---|---|---|---|---|---|---|---|---|---|
| Substrate | Substrate Concentration | pH | Temperature | Cell Density Initial-Final | Reaction Time | [29] | |||
| Ethanol | Escherichia coli | Glycerol | 30 g/L | 7.0 | 30 °C | 0.4–1.8 g/DCW/L | 28 h | 0.20 a, 31 b | [9] |
| Clostridium autoacetobutylicum | CO2 | 50 kPa | 5.8 | 37 °C | 0.2–1.8 O.D600 | 30 h | 0.28 a | [29] | |
| Propanol | Escherichia coli | Glycerol | 30 g/L | 7.0 | 30 °C | 1.8 g/DCW/L | 28 h | 0.15 a, 7 b | [27] |
| Butanol | Clostridium acetobutylicum | CO2 | - | 6.0 | 37 °C | - | 30 h | 26.2 a | [26] |
| Escherichia coli | Glycerol | 30 g/L | 7.0 | 37 °C | 0.2–7.0 O.D550 | 40 h | 6.9 a | [25] | |
| Pichia pastoris | 2-keto isovalerate | 4 g/L | - | 30 °C | 0.05–12 O.D550 | 72 h | 0.28 a | [25] | |
| Pichia pastoris | Glucose | 20 g/L | - | 30 °C | 0.05–1.2 O.D550 | 72 h | 0.89 a | [25] | |
| Pichia pastoris | Glucose | 20 g/L | - | 30 °C | 0.05–2.0 O.D550 | 72 h | 2.22 a | [8] | |
| Fatty alcohols | Escherichia coli | Glucose | 20 g/L | 7.0 | 30 °C | 0.02-O.D550 | 48 h | 1.5 a, 12.5 b | [20] |
| Yarrowia lipolytica | Glucose | 100 g/L | 5.5 | 28 °C | 1–80 O.D600 | 120 h | 2.15 a | [7] | |
| Saccharomyces cerevisiae | Lignocellulose | 20 g biomass | 5.0 | 30 °C | 1–25 O.D600 | 240 h | 1.2 a, 6 b | [30] | |
| Alkanes | Escherichia coli | Glucose | 20 g/L | 7.0 | 30 °C | 0.5–4.0 O.D600 | 50 h | 0.06 a | [30] |
| Escherichia coli | Glucose | 20 g/L | 7.0 | 30 °C | 0.5–4.0 O.D600 | 50 h | 0.08 a | [30] | |
| Escherichia coli | Glycerol | 140 g/L | 7.0 | 30 °C | 0.5–140 O.D600 | 50 h | 0.1 a, 1.3 b | [28] | |
| Escherichia coli | Glucose | 20 g/L | 7.0 | 30 °C | 0.4–3.0 O.D600 | 48 h | 0.04 a | [20] | |
| Yarrowia lipolytica | Glucose | 100 g/L | 5.5 | 28 °C | 1–80 O.D600 | 120 h | 0.02 a | [8] | |
| Escherichia coli | Glucose | 20 g/L | 7.0 | 37 °C | 0.02–102 O.D550 | 72 h | 0.43 a, 2.5 b | [29] | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, Z.; Nawab, J.; Sung, B.H.; Kim, S.C. A Critical Analysis of Bio-Hydrocarbon Production in Bacteria: Current Challenges and Future Directions. Energies 2018, 11, 2663. https://doi.org/10.3390/en11102663
Rahman Z, Nawab J, Sung BH, Kim SC. A Critical Analysis of Bio-Hydrocarbon Production in Bacteria: Current Challenges and Future Directions. Energies. 2018; 11(10):2663. https://doi.org/10.3390/en11102663
Chicago/Turabian StyleRahman, Ziaur, Javed Nawab, Bong Hyun Sung, and Sun Chang Kim. 2018. "A Critical Analysis of Bio-Hydrocarbon Production in Bacteria: Current Challenges and Future Directions" Energies 11, no. 10: 2663. https://doi.org/10.3390/en11102663
APA StyleRahman, Z., Nawab, J., Sung, B. H., & Kim, S. C. (2018). A Critical Analysis of Bio-Hydrocarbon Production in Bacteria: Current Challenges and Future Directions. Energies, 11(10), 2663. https://doi.org/10.3390/en11102663