Abstract
Hydrolysis and fermentation processes are key stages in xylitol production from lignocellulosic materials. In this study, pineapple cores, one of the wastes from the canned pineapple industry, were used as raw material for xylitol production. Two methods was used for hydrolysis: enzymatically using commercial enzyme Cellic HTec2, and acid hydrolysis using 4% H2SO4. In contrast, the fermentation process was carried out with two selected yeasts commonly employed in xylitol fermentation, Debaryomycess hansenii, and Candida tropicalis. Before these two processes, the pineapple cores were characterized using the Van Soest method to determine their lignocellulosic content. The hemicellulose content was 36.06%, the cellulose content was 14.20%, and the lignin content was 10.05%. This result indicates that the hemicellulose content of pineapple cores has the potential to be used as a raw material in the production of xylitol. The hydrolysis efficiency of enzymatic hydrolysis was 21% higher than that of acid hydrolysis. The highest xylitol and biomass yield of 0.371 gxylitol/gxylose and 0.225 gcell/gxylose were observed by C. tropicalis using an enzymatic hydrolysate.
1. Introduction
Pineapple, or Ananas comosus (L.) Merr is a popular tropical fruit from South America but has been cultivated widely worldwide [1,2]. The fruit has been processed into many kinds of food, but its byproducts such as pineapple peel, core, and crown, have not been utilized properly. Indonesia is the fourth biggest pineapple producer globally, producing 2,196,456 tonnes of pineapple per year. Therefore, pineapple is a vital commodity in the Indonesian economy [1]. However, using a massive amount of pineapples results in biomass waste that causes serious environmental issues if it is not handled correctly. The utilization of agricultural biomass waste, in addition to reducing the negative impact on the environment, has the potential to produce many bio-based products. For example, pineapple cores as byproducts have several advantages, such as high bromelain enzyme content and large lignocellulose fibers. The hemicellulose concentration of pineapple cores (28.53%) is sufficient for use as raw material for xylitol production [3].
Xylitol is a non-fermentable sugar alcohol with the same level of sweetness as sucrose and a low glycemic index. Because xylitol absorption in the body does not require insulin, it is safe to use as a food sweetener for people with diabetes. Moreover, xylitol has been widely employed as a sucrose sugar alternative in processed food goods, beverage industries, and diabetic health foods. Xylitol produces a cool sensation, heat-resistant characteristics, and slow absorption by the intestine [4,5].
Xylitol is produced industrially worldwide by catalytic hydrogenation of a pure d-xylose solution at high temperatures and pressure. Biotechnological manufacture of xylitol uses cheap industrial and agricultural waste sugar and softer process conditions [4]. The first step for biotechnological xylitol production is a hydrolysis process that converts polysaccharide molecules in lignocellulosic material into sugar monomers, such as glucose and xylose. Chemical and biological hydrolysis are two types of standard hydrolysis methods. Chemical hydrolysis uses acid or base as a catalyst at high temperatures and pressure [6]. Because this is a quick process, it must be closely monitored to avoid the co-production of numerous degradative compounds. However, the process conditions require utilizing particular reactor materials and accompanying equipment. In addition, pH must be neutralized after the reaction [7,8].
Biological or enzymatic hydrolysis employing lignocellulosic enzymes can be utilized instead of chemical hydrolysis. The use of enzymes in hydrolysis requires less energy and eliminates the use of hazardous and corrosive chemicals [9]. The utilized enzymes must be compatible with the polysaccharides to be hydrolyzed. Xylan is hydrolyzed into a D-xylose monomer containing five carbon atoms using the xylanase enzyme [9,10]. Following the extraction of the xylose from the xylan hydrolysis process, the biological reduction of xylose is executed by fermentation, mainly using yeast [11]. Candida tropicalis and Debaryomyces hansenii are the most employed yeast in the xylitol production process, which is environmentally friendly and cost-effective. These yeasts generate the xylose reductase (X.R.) enzyme that converts xylose to xylitol [8,12]. Fermentation conditions such as temperature, pH, aeration conditions, substrate content, and the presence of other sugars, such as glucose, affect the bioconversion of xylose to xylitol via fermentation [8].
Recently, xylitol has been produced from corn cobs (China) and hardwoods such as birch (US). Large amounts of industrially sourced corn cobs provide an abundant substrate for xylitol production in China, whereas birch hydrolyzate, a by-product of the paper and pulp industry, provides the substrate in the United States. A chemical catalytic reaction generally is still used to convert xylose in the hydrolyzed hemicellulose fraction to xylitol [13]. The chemical catalytic reaction in xylitol production is typically carried out at high temperatures and pressures, consuming significant energy and increasing production costs. As a result, the production of xylitol via fermentation, which can be carried out under mild conditions, is expected to meet market demand for xylitol at a lower cost production [14]. However, obstacles such as the high cost of traditional feedstock, low xylitol titer due to a lack of xylose-utilizing microbial cells, and repressed xylose metabolism in the presence of glucose, have all contributed to the slow progress of biotechnological xylitol synthesis. The potency of biochemical conversion of pineapple core waste to xylitol in Indonesia has yet to be studied. The goal of the current work is to employ cheap and plentiful lignocellulosic biomass to avoid paying high substrate expenses. It is also ideal for creating a straightforward yet affordable bioprocess that increases xylitol production. As a result, major commercial manufacturing initiatives have been related to the rise in interest in xylitol bioconversion. According to our knowledge, no attempts have been made to produce xylitol from pineapple core waste. Therefore, this paper focuses on optimizing hydrolysis processes using acid and enzymatic hydrolysis, and fermentation processes using C. tropicalis and D. hansenii.
2. Materials and Methods
2.1. Tools and Materials
The pineapples were sourced from an Indonesian pineapple orchard in Subang, West Java, Indonesia. The pineapple cores were mashed in a blender (Philips, Zhuhai, China) and dried in a dry oven(B-One, Messgerate Sukses Mandiri Ltd., Tangerang, Indonesia) at 60 °C for 24 h to make a powder sieved with a 60–60 mesh size (0.25–0.177 mm).
Approximately 75 I.U./mL of the Cellic HTec2 enzyme (Novozymes, Copenhagen, Denmark) was employed for the enzymatic hydrolysis of xylan. Debaryomyces hansenii ITB CC R85 and Candida tropicalis were collected from the ITB (Institut Teknologi Bandung, Bandung, Indonesia) Department of Chemical Engineering culture collection.
All of the additional chemicals received from Sigma Aldrich (St. Louis, MO, USA) were of analytical grade and were used immediately. Each solution was produced with distilled water.
2.2. Enzymatic Hydrolysis
The pineapple core powder was sterilized by submerging 20 g of the powder in 100 mL of acetate buffer at pH 5 and then autoclaving it for 15 min at 121 °C. Then, 50 I.U./g of biomass Cellic HTec 2 was added, and the mixture was shaken at 150 rpm for 96 h at 60 °C in a shaker incubator (N-Biotek, Seoul, Republic of Korea). The xylose hydrolyzate was obtained after 20 min of centrifugation at 5000 rpm. Every 24 h, sampling was conducted. At 70 °C, the liquid hydrolysate was concentrated threefold by evaporation.
2.3. Acid Hydrolysis
A 10 g powdered pineapple core was dissolved in 250 mL of 0.72 M or 4% (v/v) of 96% H2SO4 stock solution and autoclaved for 20 min at 121 °C in a 500 mL Erlenmeyer flask. The hydrolyzate was filtered to remove the dregs before being neutralized with 2 M NaOH to attain a pH of 7. It was then detoxified using 15 g of activated charcoal for 1 h at 30 °C and concentrated threefold by evaporation at 70 °C.
2.4. Fermentation
D. hansenii and C. tropicalis seeds were cultivated on growth media containing 50 mL of xylose (20 g/L) and 50 mL of nutritional medium containing 9.438 g/L (NH4)2SO4, 2.5 g/L KH2PO4, 0.05 g/L CaCl2·2H2O, 0.5 g/L MgSO4·7H2O, 0.5 g/L citric acid, 0.035 g/L FeSO4·7H2O, 0.0092 g/L MnSO4·7H2O, 0.011 g/L ZnSO4·7H2O, 0.001 g/L CuSO4·7H2O, 0.002 g/L CoCl2·6H2O, 0.0013 g/L Na2CoO4·2H2O, 0.002 g/L H3BO3, 0.0035 g/L KI, 0.0005 g/L Al2 (SO4)3, 0.1 g/L myo-inositol, 0.02 g/L calcium-pantothenate, 0.005 g/L thiamine hydrochloride, 0.005 g/L pyridoxal hydrochloride, 0.005 g/L nicotine acid, 0.001 g/L aminobenzoic acid, and 0.0001 g/L d-biotin. The seeds were cultured for 48 h at 30 °C with a stirring speed of 150 rpm in a shaker incubator.
The pineapple core hydrolyzate used in fermentation was generated via enzymatic and acid hydrolysis. The microaerobic condition was produced by combining nitrogen gas and air at a volume ratio of 1:5 in the Erlenmeyer head space. The volume ratio of the inoculum (106 CFU/mL), hydrolysate, and fermentation medium was 2:2:3.
2.5. Purification of Fermentation Product
The fermentation product was separated from cell biomass using centrifugation at 5000 rpm for 40 min. The supernatant was filtered using filter paper before being transferred to a 250 mL Erlenmeyer flask for further purification with 15 g/L of activated charcoal. The purification process was performed at 30 °C for 1 h using a magnetic stirrer (IKA, Selangor, Malaysia) for agitation. In the final step, the purified sample was filtered with filter paper for further analysis. The total working volume in a 250 mL Erlenmeyer flask was 100 mL. The microaerobic condition was produced by combining nitrogen gas and air in the Erlenmeyer head space at a volume ratio of 1:5.
2.6. Analysis Method
Using the Van Soest method [15], the biomass, cellulose, hemicellulose, and lignin contents were measured simultaneously to estimate the composition of lignocellulose. An HPLC system with UV and refractive index (R.I.) detectors was used to evaluate the hydrolysis and fermentation samples (Waters type 1515 pump; Autosampler type 2707, Waters, Milford, MA, USA.). The concentrations of xylose, glucose, ethanol, and xylitol were measured at 65 °C, with 5 mM H2SO4 as the mobile phase and 0.6 mL/min as the flow rate, using an Aminex HPX-87H column with a R.I. detector. Using a calibration curve, the optical density at 650 nm, which was used to quantify cell growth, was converted to dry cell weight (D.C.W.). The OPEFB surface was examined with a scanning electron microscope (JEOL, JSM-6330F; Tokyo, Japan) before and after several processes.
2.7. Data Interpretation
The hydrolysis yield measurement was calculated based on Equations (1)–(5)
Hemicellulose hydrolysis efficiency (%) is:
Maximum theoretical xylose from hydrolysis (Xylose (theo)):
The yield of biomass (YX/S) (g/g) is:
The product yield (YP/S) (g/g) is:
where xylose(t) is the xylose concentration (g/L) produced at time t, X is the biomass concentration (g/L), S is the substrate concentration (g/L), and P is the product (xylitol or ethanol) concentration (g/L).
Xylose or glucose Utilization (%) is:
Xylitol fermentation efficiency (%) is:
The theoretical yield of xylitol (xylitol (theo)) is 0.9 mol of xylitol per mol of xylose utilized [16].
The specific growth rate (µ) is:
2.8. Statistical Analysis
Student’s t-test was used to determine the statistical significance of each measurement. The information is given as a standard deviation from the mean. A p-value of less than 0.05 was regarded as statistically significant.
3. Results and Discussion
3.1. Lignocellulose Composition of Pineapple Core
The lignocellulose composition of the raw materials in the pineapple core was deteremined to estimate the quantity of xylose and glucose produced during hydrolysis. Table 1 summarizes the findings of the lignocellulose content analysis of the pineapple cores. The percentages of cellulose, hemicellulose, and lignin were 14.20 ± 0.31%, 36.06 ± 0.22%, and 10.05 ± 0.93%, respectively. The lignocellulose composition in this material was suitable for xylitol production by yeast fermentation. The high hemicellulose content will provides a great deal of xylose as the raw material for xylitol production [17]. The low cellulose content provides enough glucose to support microbial growth without inhibiting xylitol production [18]. The low lignin content facilitates the hydrolysis process, both acid and enzymatic, because the access of enzymes or acids to cellulose and hemicellulose is not much hindered by lignin [19,20].

Table 1.
Composition of Lignocellulosic Content of Pineapple Cores.
3.2. Impact of Hydrolysis Types on Hydrolysate Composition
In this study, two different hydrolysis methods were used to determine which resulted in the most xylose and hemicellulose hydrolysis efficiency, namely enzymatic hydrolysis and acid hydrolysis.
In the enzymatic hydrolysis of pineapple cores, auto-hydrolysis was used to break down lignocellulosic tissue and release hemicellulose from the material [9,21]. Because enzymes have specificity, their role in enzymatic hydrolysis is crucial; thus, enzyme performance is optimal if the substrate used is suitable and at the proper concentration [22]. The commercial enzyme cellic Htech2 was used in this study at a concentration of 50 IU/g biomass and a solid loading of 20%. This enzymatic hydrolysis yielded 23.792 g/L of xylose, with a hemicellulose hydrolysis efficiency of 37.550%. Furthermore, 2.73 g/L glucose was produced as a byproduct of hydrolysis (Table 2). The amount of glucose in the enzymatic hydrolysis process was low, since xylanase was more dominant than cellulase in the Cellic HTec enzyme blend composition, and the cellulose content in pineapple cores was lower than the hemicellulose content.
Table 2.
Components of pineapple core hydrolysate.
For acid hydrolysis, concentrations of up to 4% or 0.72 M H2SO4 were utilized. Acid hydrolysis with diluted acid is frequently employed for processing lignocellulosic material due to its efficiency and cost-effectiveness in producing sugars such as d-xylose and d-glucose [23]. However, the toxicity to microbial growth (furans, aliphatic acids, and phenolic components) complicates the fermentability of hydrolyzate and even stops the fermentation process [24]. A detoxification process, such as overliming, activated charcoal, pH adjustment, or enzyme treatment, can increase biomass and xylitol production by removing 4% to 95% of these inhibitory compounds [25]. Activated charcoal was used for detoxification in this study because Winkelhausen and Kuzmanova [5] reported that this approach is a simple detoxification treatment that can be combined with additional techniques, such as alkalinization and observation of the elimination of acetic acid, lignin phenolic compounds, furan (5-HMF and furfural), and clarity. The xylose obtained from the acid hydrolysis pretreatment was 9.844 g/L, or 31.074% of the theoretical hydrolysis yield. At the same time, the amount of glucose produced was only 1.22 g/L due to the low cellulose content in the pineapple core. The factors that affected the amount of xylose produced by acid hydrolysis were temperature, acid concentration, and reaction time [26]. More reducing sugar was obtained using a higher acid concentration and a longer time. However, the disadvantage of the high acid concentration was that it caused a decomposition reaction in which the sugar components dehydrated into furfural compounds [17].
Table 2 shows the results of enzymatic hydrolysis, which are significantly better than acid hydrolysis, including the value of hemicellulose hydrolysis efficiency, which is greater than 10%. The amount of solid loading is much higher, which can be applied to enzymatic hydrolysis to produce a high concentration of xylose.
3.3. Influence of Different Hydrolysate Types on Substrate Utilization by D. hansenii and C. tropcalis
Debaryomyces hansenii consumed xylose from enzymatic hydrolysate at a rate of 47.200%, and glucose was utilized at 98.890%. On the other hand, the consumption rate for reducing sugar obtained from acid hydrolysis was 41.860% for xylose and 97.540% for glucose (Table 3). This result implies that D. hansenii did not thoroughly consume all xylose for metabolite product formation or cell biomass synthesis. However, compared to xylitol production using OPEFB enzymatic hydrolysate by D. hansenii in a previous study [27], the xylose utilization in the current study was 57.3% higher. The use of xylose in pineapple core hydrolyzate and OPEFB may differ due to the complex medium of each hydrolysate, which contains other unknown substances consumed by the microorganisms [28]. On the other hand, inhibitory compounds such as acetic acid and furfural in acid hydrolysate can cause the slow conversion of xylose to xylitol during hydrolyzate fermentation [29]; therefore, the xylitol consumption in the acid hydrolysate was lower than in enzymatic one. A graph showing the decrease in xylose and glucose concentration during fermentation by D. hansenii is presented in Figure 1A,B.
Table 3.
Fermentation products of enzyme hydrolysate and acid hydrolysate by D. hansenii.
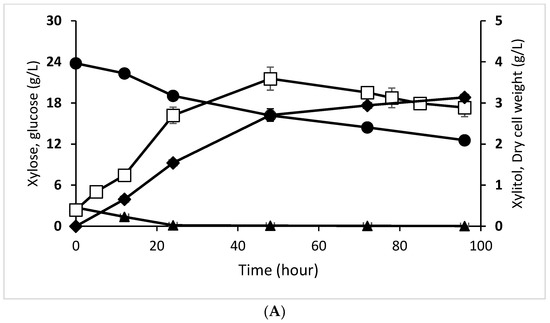
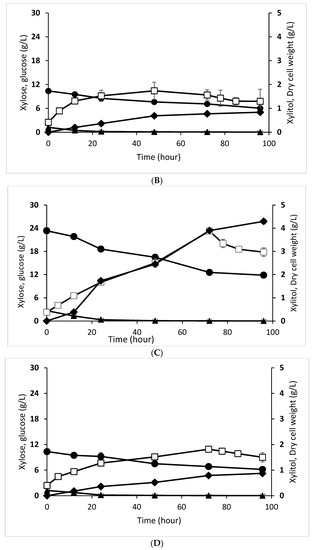
Figure 1.
Xylitol fermentation profiles of D. hansenii in enzymatic hydrolysate (A), D. hansenii in acid hydrolysate (B), C. tropicalis in enzymatic hydrolysate (C), and C. tropicalis in acid hydrolysate (D). Xylose concentration (●, closed circle), glucose concentration (▲, closed triangle), dry cell weight (□, open square), xylitol concentration (♦, closed diamond). The data are the means of three replicated investigations, and the error bars are standard deviations.
C. tropicalis is an excellent yeast for converting xylose to xylitol [30]. Like the xylose consumption pattern in D. hansenii, C. tropicalis also consumed more xylose in the enzymatic hydrolysate. The percentage of C. tropicalis that consumed xylose from enzymatic hydrolysate was 49.600%, while that which consumed glucose was 98.370%. The consumption of xylose from acid hydrolysate was 40.490%, while for glucose, it was 99.400%. In summary, the xylose consumption in enzymatic hydrolysate by C. tropicalis was higher than by D. hansenii, while there was no significant difference in xylose utilization in acid hydrolysate between the two yeasts. A graph of the decrease in xylose and glucose concentration during C. tropicalis fermentation is presented in Figure 1C,D.
3.4. Effect of Enzymatic and Acid Hydrolysate on Xylitol Production Profile by D. hansenii and C. tropicalis
Xylitol production by D. hansenii and C. tropicalis was evaluated using enzymatic and acid hydrolysate as the primary substrate (Figure 1). The two strains grew using glucose and xylose as a carbon source to build up cell biomass and xylitol. Glucose was consumed up to the first 24 h, since it is the most readily consumed substrate by the yeast and its concentration was low in the fermentation medium. However, the xylose consumption, cell growth, and xylitol production in C. tropicalis tended to be higher than that of D. hansenii in both enzymatic and acid hydrolysate. Moreover, these xylitol production parameters were higher using enzymatic hydrolysate than that acid hydrolysate for both yeasts.
D. hansenii converted 49.670% of xylose from enzymatic hydrolysate to produce 3.136 g/L of xylitol with 0.279 gxylitol/gxylose of xylitol yield and 0.221 gcell/gxylose of biomass yield (Table 3). In contrast, in the acid hydrolysate, the yeast converted 45.090% of xylose producing 0.837 g/L of xylitol with 0.193 gxylitol/gxylose of xylitol yield and 0.203 gcell/gxylose of biomass yield. The xylitol yield obtained in the current study was low when compared to other studies using other lignocellulose material. According to Parajo et al. [31], D. hansenii fermentation from sawdust hydrolysate produced approximately 0.5 to 9 g/L of xylitol from 18 g/L of xylose, resulting in a maximum yield of 0.79 gxylitol/gxylose.
C. tropicalis converted 49.600% of xylose from enzymatic hydrolysate resulting in 4.296 g/L of xylitol with a xylitol yield of 0.371 gxylitol/gxylose and a biomass yield of 0.225 gcell/gxylose, while in the acid hydrolysate, the yeast converted 40.490% of xylose producing 0.873 g/L of xylitol with 0.210 gxylitol/gxylose of xylitol yield and 0.211 gcell/gxylose of biomass yield (Table 4). Our method produced a greater concentration of xylitol than Tran et al. study [32] in the enzymatic hydrolysate of beechwood and walnut shell which obtained 2–3.5 g/L of xylitol after 72 h fermentation using C. tropicalis yeast.
Table 4.
Fermentation products of enzyme hydrolysate and acid hydrolysate by C. tropicalis.
Logarithmic growth of D. hansenii in both enzymatic and acid hydrolysate occurred for up to 48 h (Table 4). At this time, the cell concentration in the enzymatic hydrolysate was 3.59 g/L and 1.74 g/L in the acid hydrolysate, with a specific growth rate of 1.108 h−1 and 0.102 h−1, respectively. On the other hand, the logarithmic growth for C. tropicalis occurred at 72 h in both enzymatic and acid hydrolysate resulting in the highest cell concentration of 3.880 g/L and 1.82 g/L with specific growth rates of 0.088 h−1 and 0.082 h−1, respectively. Both yeast cells in acid hydrolysate grew slower than those in enzymatic hydrolysate due to the lower xylose concentration and, perhaps, toxic compounds remaining in the fermentation liquid. Compared to C. tropicalis growth with both hydrolysate, D. hansenii had a higher specific growth rate because this yeast reached logarithmic growth faster than C. tropicalis even though the xylitol yield and biomass yield were lower than C. tropicalis in the enzymatic hydrolysate.
3.5. Morphology Changes of Fresh and Hydrolyzed Pineapple Core
Morphological characterizations of the new and hydrolyzed pineapple cores were carried out by SEM analysis to determine the changes in the microstructure of the pineapple core during the enzymatic (xylanase) and acid hydrolysis process. The fresh pineapple core had a rigid surface and compact structure with fibers organized in bundles, as shown in Figure 2A. After hydrolysis, the system of the pineapple core was severely damaged, fractured, and roughened, and pores had formed. It was suggested that most of the lignin and hemicellulose were removed, resulting in a change of external and internal structure, and increased porosity. Because the enzyme or acid decomposed the lignin bonds, allowing it to break down the carbohydrates in the pineapple core inner structure, which contains hemicellulose and cellulose, the morphology structure was degraded after hydrolysis (Figure 2B,C), resulting in more and larger pores [33].

Figure 2.
Scanning electron microscope images of fresh OPEFB (A), after enzymatic hydrolysis (B), after acid hydrolysis (C).
Comparing fresh cores and post acid hydrolysis samples, the average pore area of enzymatically hydrolyzed pineapple core was much more significant. This was caused by the 96 h hydrolysis incubation period, whereas the disconnection of fiber chains into sugar monomers dissolved in the hydrolysate liquid enlarged the pores of fiber particles [9]. Acid hydrolysis caused damage to most of the surface of the pineapple core sample.
4. Conclusions
This study showed that pineapple cores are a potential source for xylitol-based products due to their high hemicellulose content, which is 36.060%. It was found that the enzymatic hydrolysis method resulted in over 20% higher xylose concentration than acid hydrolysis. It was also found that the highest concentration of xylitol production was 4.29 g/L, which was produced by Candida tropicalis. The ratio between product concentration and the substrate consumed (Yp/s) was 0.371 gxylitol/gxylose, and the ratio between product and growth cell (Yp/x) was 0.225 gcell/gxylose.
Author Contributions
Conceptualization: E.M., B.N., H.N.F. and A.T.H.; data curation: A.T.H., B.N., E.H. and H.N.F.; formal analysys: H.N.F. and R.A.E.; funding acquisition: E.M. and B.N.; investigation: B.N., E.H., H.N.F. and R.A.E.; methodology: E.M., B.N., E.H. and A.T.H.; project administration: E.M. and B.N.; resources: R.A.E. and A.T.H.; writing-original draft: E.M., A.T.H. and H.N.F.; writing—review and editing: B.N., H.N.F., E.H. and R.A.E. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Education, Research and Technology of Indonesia, Universitas Padjadjaran, and the National Research and Innovation Agency (BRIN).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We would like to thank Faculty of Agroindustrial Technology, Department of Agroindustrial Technology, Universitas Padjadjaran and the farmers in Subang, Indonesia, who provided pineapple samples for this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hikal, W.M.; Mahmoud, A.A.; Said-Al Ahl, H.A.H.; Bratovcic, A.; Tkachenko, K.G.; Kačániová, M.; Rodriguez, R.M. Pineapple (Ananas comosus L. Merr.), Waste Streams, Characterisation and Valorisation: An Overview. Open J. Ecol. 2021, 11, 610–634. [Google Scholar] [CrossRef]
- Zhang, Y.; Xia, X. Physicochemical Characteristics of Pineapple (Ananas Mill.) Peel Cellulose Prepared by Different Methods. Adv. Mater. Res. 2012, 554–556, 1038–1041. [Google Scholar] [CrossRef]
- Pardo, M.E.S.; Cassellis, M.E.R.; Escobedo, R.M.; García, E.J. Chemical Characterisation of the Industrial Residues of the Pineapple (Ananas comosus). J. Agric. Chem. Environ. 2014, 3, 53–56. [Google Scholar] [CrossRef]
- Dasgupta, D.; Bandhu, S.; Adhikari, D.K.; Ghosh, D. Challenges and Prospects of Xylitol Production with Whole Cell Bio-Catalysis: A Review. Microbiol. Res. 2017, 197, 9–21. [Google Scholar] [CrossRef]
- Winkelhausen, E.; Kuzmanova, S. Microbial Conversion of D-Xylose to Xylitol. J. Ferment. Bioeng. 1998, 86, 1–14. [Google Scholar] [CrossRef]
- Venkateswar Rao, L.; Goli, J.K.; Gentela, J.; Koti, S. Bioconversion of Lignocellulosic Biomass to Xylitol: An Overview. Bioresour. Technol. 2016, 213, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Suhartini, S.; Rohma, N.A.; Mardawati, E.; Kasbawati; Hidayat, N.; Melville, L. Biorefining of Oil Palm Empty Fruit Bunches for Bioethanol and Xylitol Production in Indonesia: A Review. Renew. Sustain. Energy Rev. 2022, 154, 111817. [Google Scholar] [CrossRef]
- Kresnowati, M.; Mardawati, E.; Setiadi, T. Production of Xylitol from Oil Palm Empty Friuts Bunch: A Case Study on Bioefinery Concept. Mod. Appl. Sci. 2015, 9, 206. [Google Scholar] [CrossRef]
- Mardawati, E.; Werner, A.; Bley, T.; Mtap, K.; Setiadi, T. The Enzymatic Hydrolysis of Oil Palm Empty Fruit Bunches to Xylose. J. Japan Inst. Energy 2014, 93, 973–978. [Google Scholar] [CrossRef]
- Mardawati, E.; Purwadi, R.; Setiadi, T. Evaluation of the Enzymatic Hydrolysis Process of Oil Palm Empty Fruit Bunch Using Crude Fungal Xylanase. ARPN J. Eng. Appl. Sci. 2017, 12, 5286–5292. [Google Scholar]
- Parajó, J.C.; Domínguez, H.; Domínguez, J.M. Biotechnological Production of Xylitol. Part 2: Operation in Culture Media Made with Commercial Sugars. Bioresour. Technol. 1998, 65, 203–212. [Google Scholar] [CrossRef]
- Azizah, N. Biotransformation of Xylitol Production from Xylose of Lignocellulose Biomass Using Xylose Reductase Enzyme: Review. J. Food Life Sci. 2019, 3, 103–112. [Google Scholar] [CrossRef]
- Ravella, S.R.; Gallagher, J.; Fish, S.; Prakasham, R.S. D-Xylitol; da Silva, S.S., Chandel, A.K., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; ISBN 9783642318863. [Google Scholar]
- Mardawati, E.; Febrianti, E.A.; Fitriana, H.N.; Yuliana, T.; Putriana, N.A.; Suhartini, S.; Kasbawati. An Integrated Process for the Xylitol and Ethanol Production from Oil Palm Empty Fruit Bunch (OPEFB) Using Debaryomyces hansenii and Saccharomyces cerevisiae. Microorganisms 2022, 10, 2036. [Google Scholar] [CrossRef] [PubMed]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Tamburini, E.; Costa, S.; Marchetti, M.G.; Pedrini, P. Optimized Production of Xylitol from Xylose Using a Hyper-Acidophilic Candida tropicalis. Biomolecules 2015, 5, 1979–1989. [Google Scholar] [CrossRef]
- Harahap, B.M. Degradation Techniques of Hemicellulose Fraction from Biomass Feedstock for Optimum Xylose Production: A Review. J. Keteknikan Pertan. Trop. dan Biosist. 2020, 8, 107–124. [Google Scholar] [CrossRef]
- Mardawati, E.; Wira, D.W.; Kresnowati, M.; Purwadi, R.; Setiadi, T. Microbial Production of Xylitol from Oil Palm Empty Fruit Bunches Hydrolysate: The Effect of Glucose Concentration. J. Japan Inst. Energy 2015, 94, 769–774. [Google Scholar] [CrossRef]
- Sugiharto, Y.E.C.; Harimawan, A.; Kresnowati, M.T.A.P.; Purwadi, R.; Mariyana, R.; Andry; Fitriana, H.N.; Hosen, H.F. Enzyme Feeding Strategies for Better Fed-Batch Enzymatic Hydrolysis of Empty Fruit Bunch. Bioresour. Technol. 2016, 207, 175–179. [Google Scholar] [CrossRef]
- Qi, B.; Chen, X.; Su, Y.; Wan, Y. Enzyme Adsorption and Recycling during Hydrolysis of Wheat Straw Lignocellulose. Bioresour. Technol. 2011, 102, 2881–2889. [Google Scholar] [CrossRef]
- Vázquez, M.J.; Alonso, J.L.; Domínguez, H.; Parajó, J.C. Production of Xylose-Containing Fermentation Media by Enzymatic Post-Hydrolysis of Oligomers Produced by Corn Cob Autohydrolysis. World J. Microbiol. Biotechnol. 2001, 17, 817–822. [Google Scholar] [CrossRef]
- Chen, H.Z.; Liu, Z.H. Enzymatic Hydrolysis of Lignocellulosic Biomass from Low to High Solids Loading. Eng. Life Sci. 2017, 17, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Mateo, S.; Roberto, I.C.; Sánchez, S.; Moya, A.J. Detoxification of Hemicellulosic Hydrolyzate from Olive Tree Pruning Residue. Ind. Crops Prod. 2013, 49, 196–203. [Google Scholar] [CrossRef]
- Purwadi, R.; Niklasson, C.; Taherzadeh, M.J. Kinetic Study of Detoxification of Dilute-Acid Hydrolyzates by Ca(OH)2. J. Biotechnol. 2004, 114, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Mussatto, S.I.; Roberto, I.C. Alternatives for Detoxification of Diluted-Acid Lignocellulosic Hydrolyzates for Use in Fermentative Processes: A Review. Bioresour. Technol. 2004, 93, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Ma, H.; Tian, Z.; Lyu, G.; Fang, G.; Chen, J.; Saeed, H.A.M. Production of Xylose from Diluted Sulfuric Acid Hydrolysis of Wheat Straw. BioResources 2017, 12, 7084–7095. [Google Scholar] [CrossRef]
- Mardawati, E.; Mandra Harahap, B.; Ayu Febrianti, E.; Try Hartono, A.; Putri Siahaan, N.; Wulandari, A.; Yudiastuti, S.; Suhartini, S.; Kasbawati, K. Integrated and Partial Process of Xylitol and Bioethanol Production from Oil Palm Empty Fruit Bunches. Adv. Food Sci. Sustain. Agric. Agroind. Eng. 2022, 5, 49–67. [Google Scholar] [CrossRef]
- Mohamad, N.L.; Kamal, S.M.M.; Abdullah, N.; Ismail, I. Evaluation of Fermentation Conditions by Candida tropicalis for Xylitol Production from Sago Trunk Cortex. BioResources 2013, 8, 2499–2509. [Google Scholar] [CrossRef][Green Version]
- Rahman, S.H.A.; Choudhury, J.P.; Ahmad, A.L. Production of Xylose from Oil Palm Empty Fruit Bunch Fiber Using Sulfuric Acid. Biochem. Eng. J. 2006, 30, 97–103. [Google Scholar] [CrossRef]
- Cheng, K.K.; Zhang, J.A.; Ling, H.Z.; Ping, W.X.; Huang, W.; Ge, J.P.; Xu, J.M. Optimization of PH and Acetic Acid Concentration for Bioconversion of Hemicellulose from Corncobs to Xylitol by Candida Tropicalis. Biochem. Eng. J. 2009, 43, 203–207. [Google Scholar] [CrossRef]
- Parajó, J.C.; Domínguez, H.; Domínguez, J.M. Biotechnological Production of Xylitol. Part 1: Interest of Xylitol and Fundamentals of Its Biosynthesis. Bioresour. Technol. 1998, 65, 191–201. [Google Scholar] [CrossRef]
- Tran, L.H.; Yogo, M.; Ojima, H.; Idota, O.; Kawai, K.; Suzuki, T.; Takamizawa, K. The Production of Xylitol by Enzymatic Hydrolysis of Agricultural Wastes. Biotechnol. Bioprocess Eng. 2004, 9, 223–228. [Google Scholar] [CrossRef]
- Soontornchaiboon, W.; Chunhachart, O.; Pawongrat, R. Ethanol Production from Pineapple Waste by Co-Culture of Saccharomycs cerevisiae TISTR 5339 and Candida shehatae KCCM 11422. Asia-Pacific J. Sci. Technol. 2016, 21, 347–355. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).