
Evaluation of the Anticoccidial Activity of Sheep Bile against Eimeria stiedae Oocysts and Sporozoites of Rabbits: An In Vitro Study

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Bile
2.2. Infrared Spectroscopy
2.3. Isolation and Preparation of Eimeria stiedae
2.4. In Vitro Effect of Sheep Bile on Inhibition of Sporulated Oocysts
2.5. In Vitro Anti-Sporozoite Effect of Sheep Bile
2.6. Statistical Analysis
3. Results
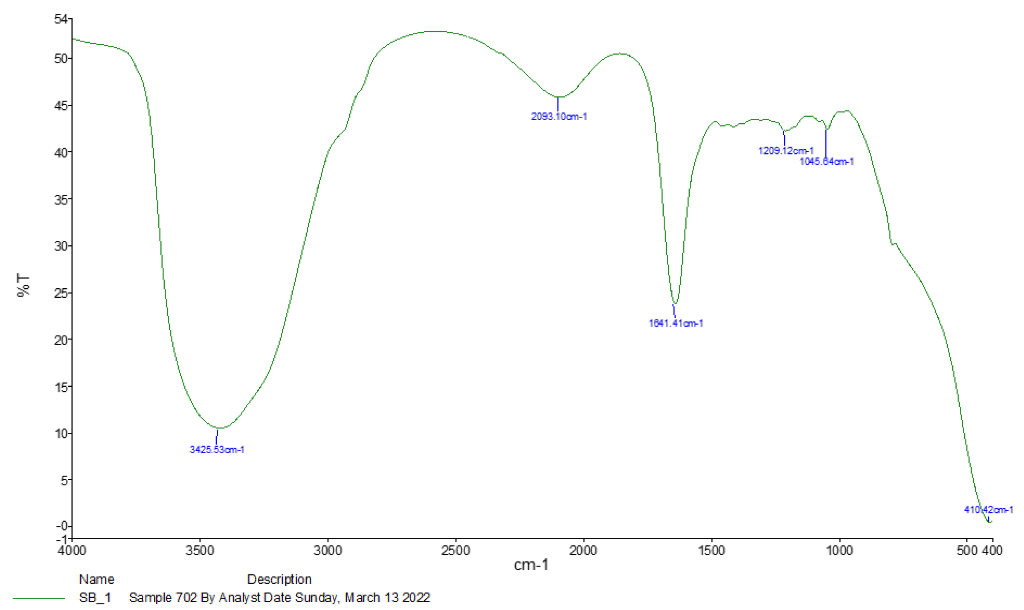
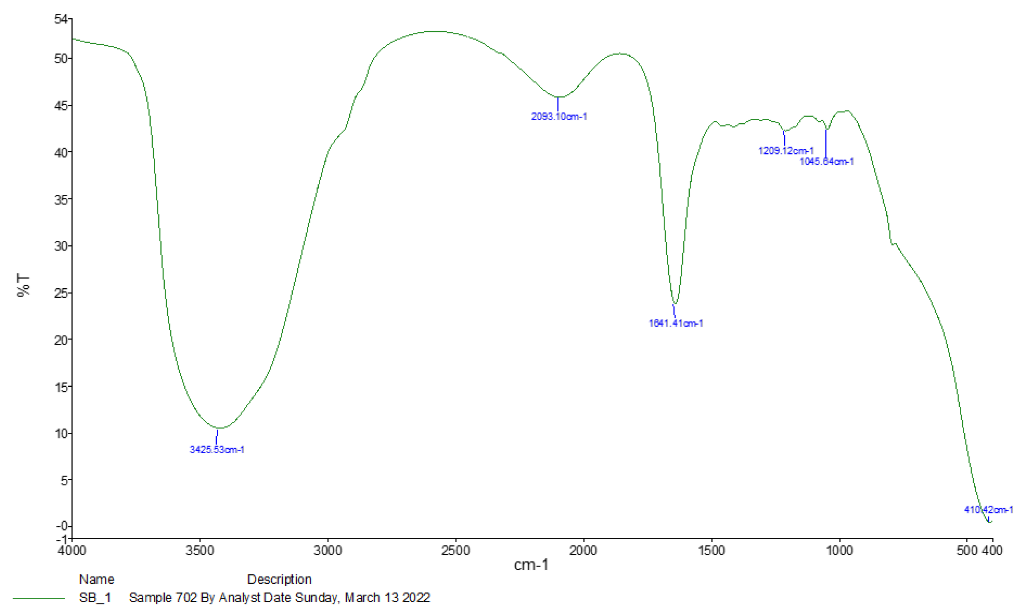
3.1. Spectroscopy Results
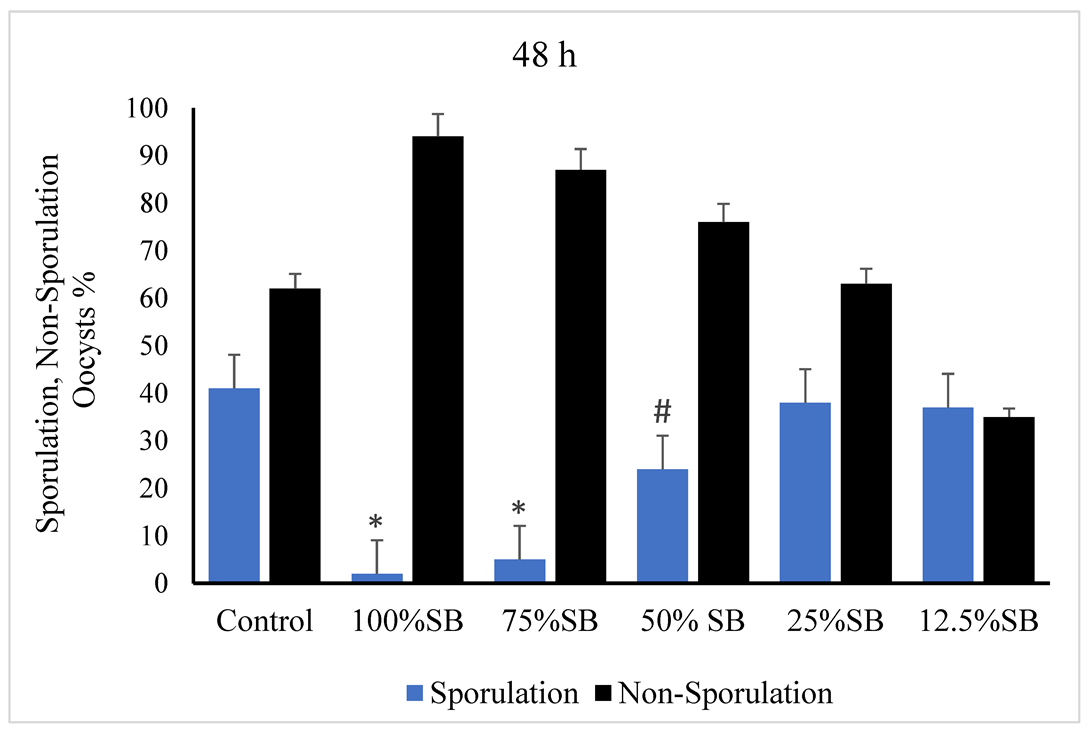
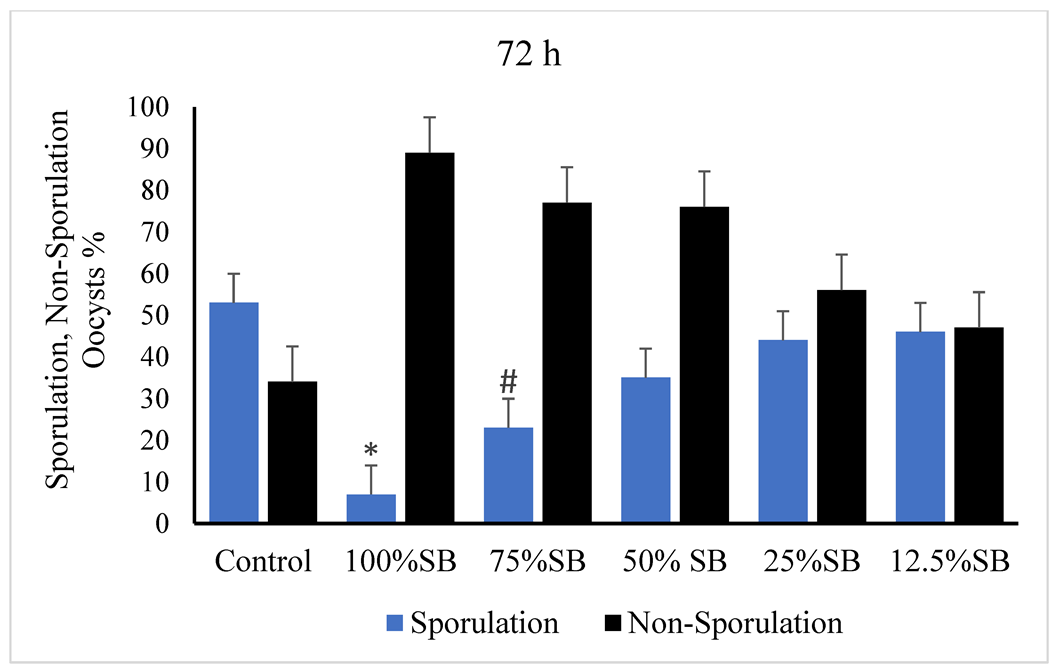
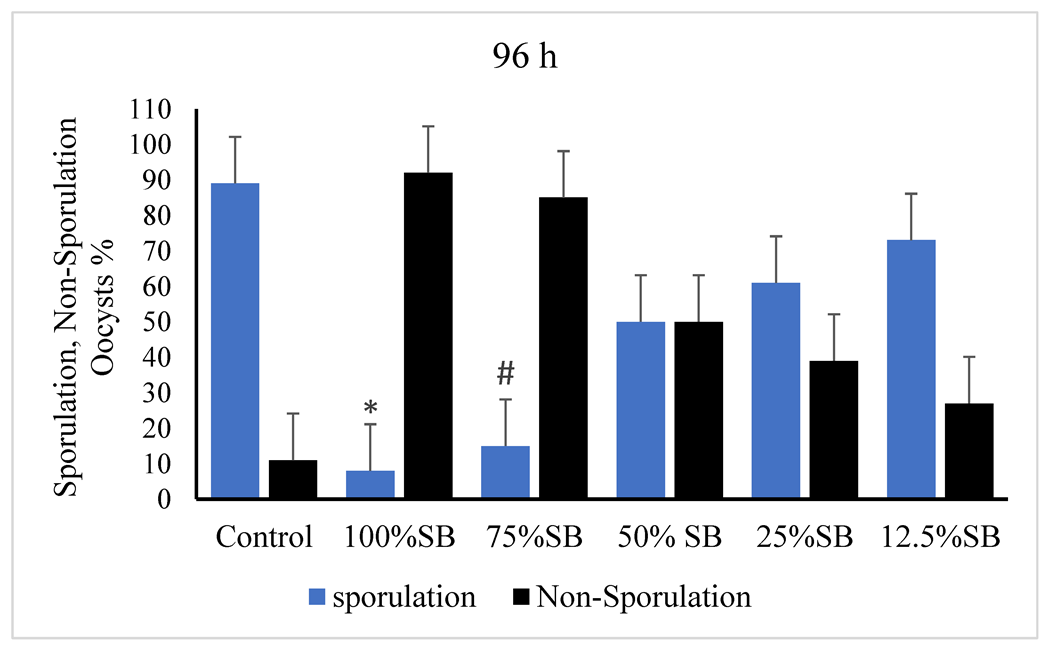
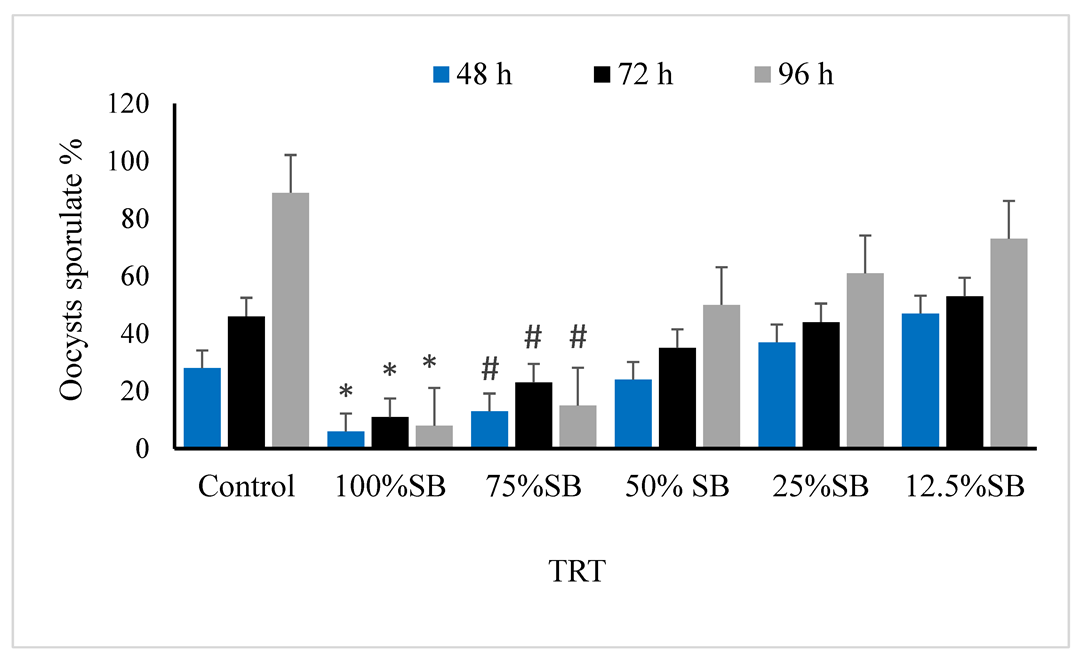
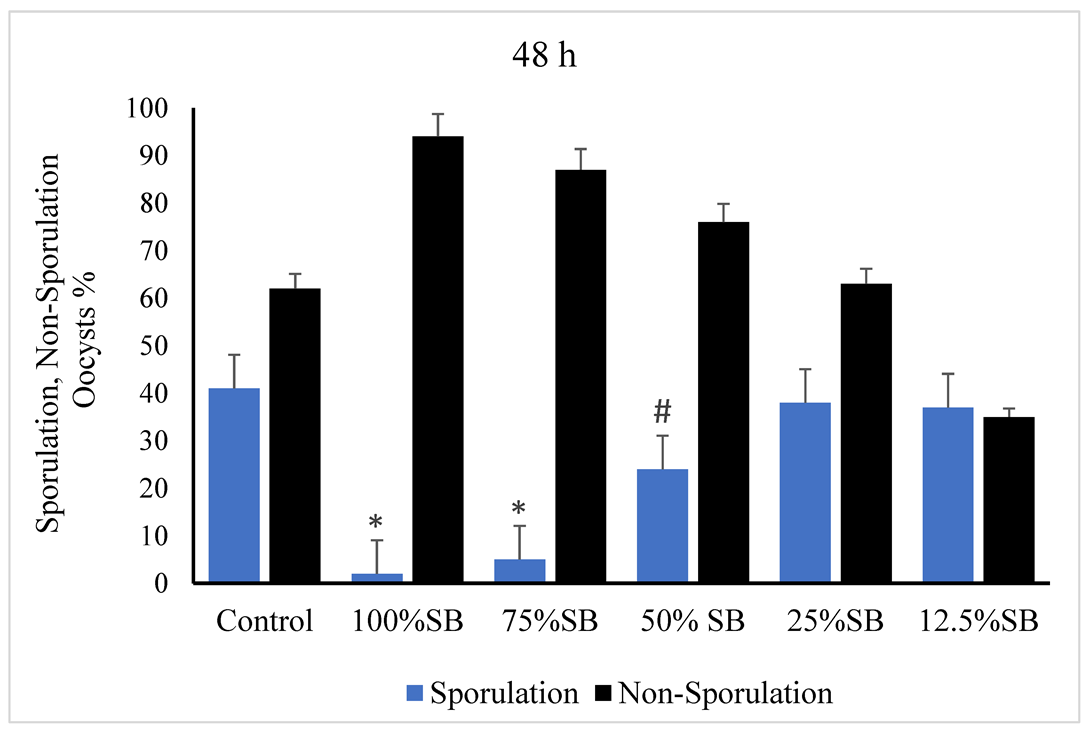
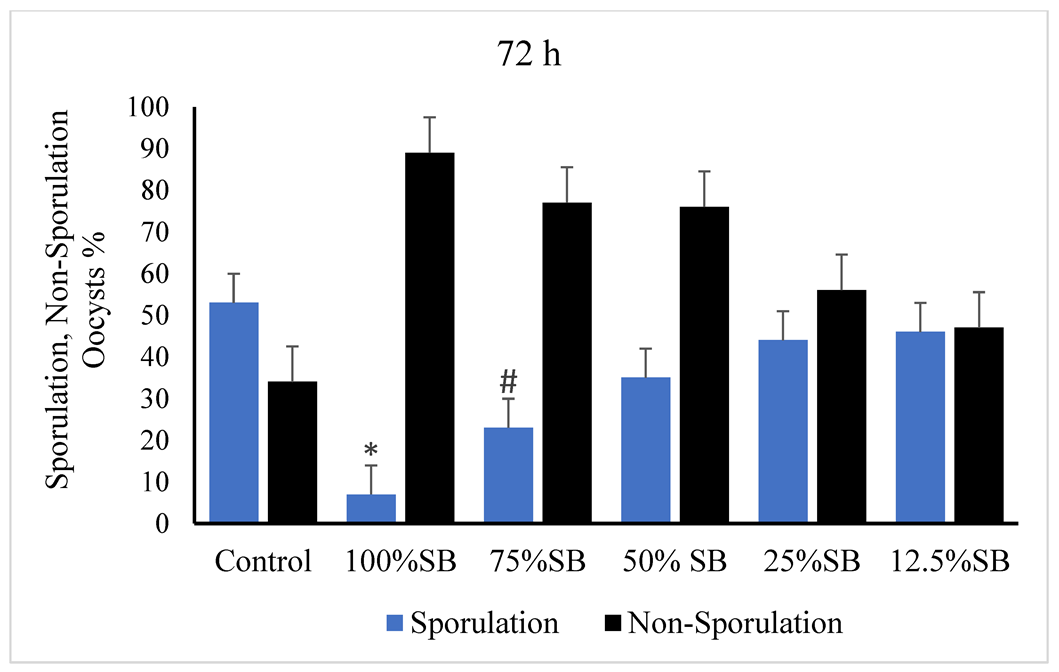
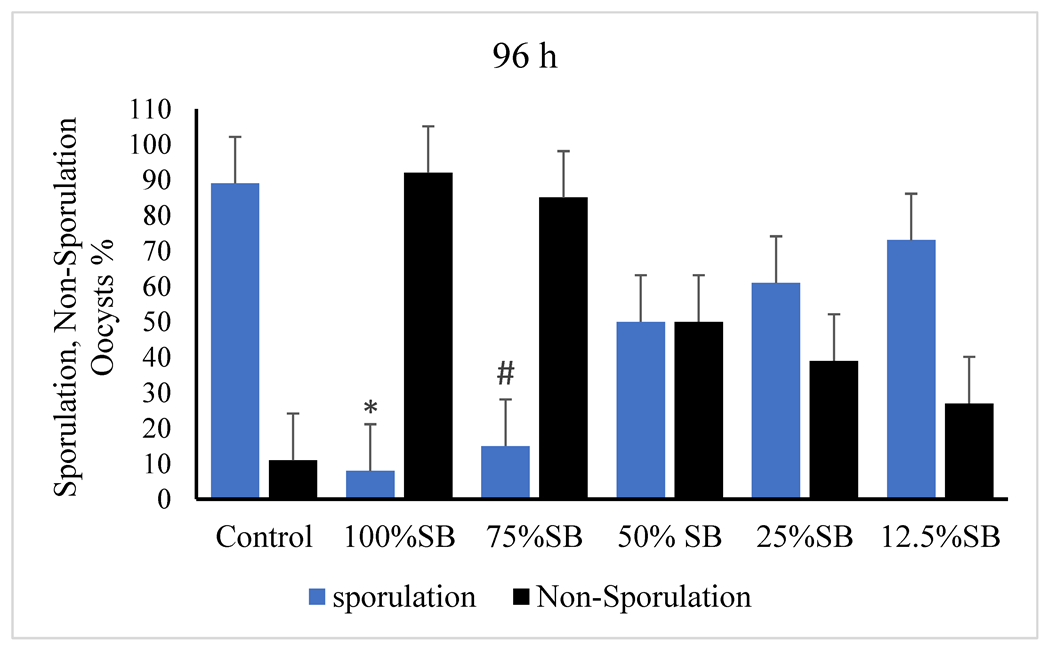
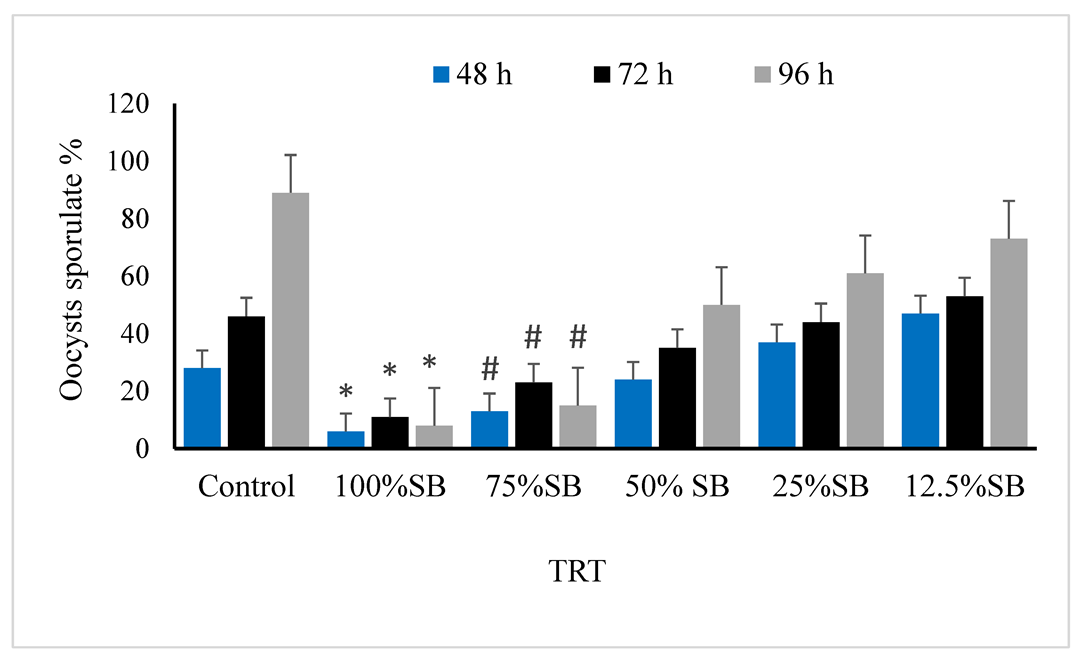
3.2. Oocysticidal Activities of Sheep Bile In Vitro
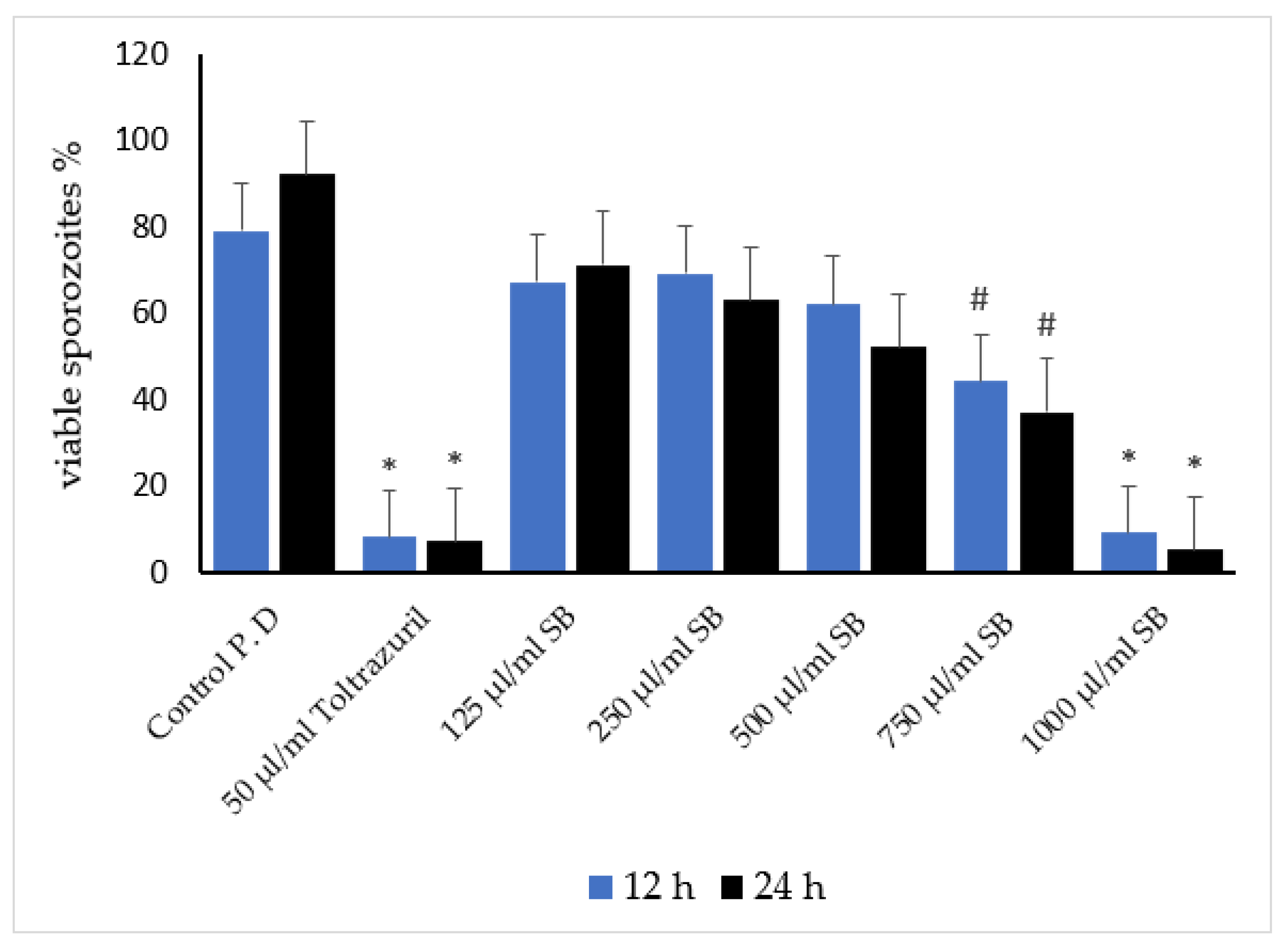
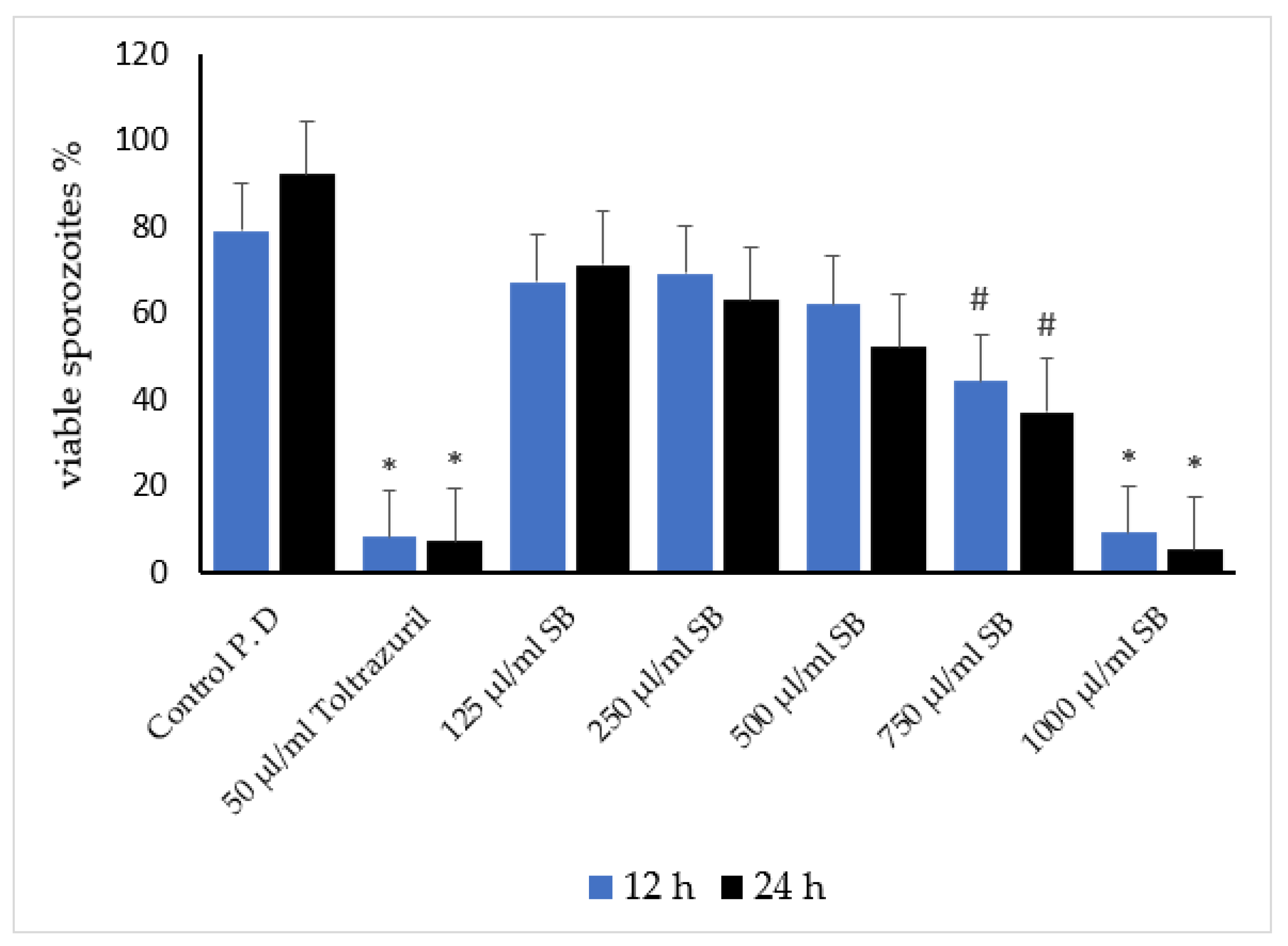
3.3. Antisporozoidal Activity of Sheep Bile In Vitro
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gill, B.; Ray, H. The coccidian of the domestic rabbit and the common field hare of India. Proc. Zool. Soc. 1960, 13, 128–143. [Google Scholar]
- Kvičerová, J.; Pakandl, M.; Hypša, V. Phylogenetic relationships among Eimeria spp. (Apicomplexa, Eimeriidae) infecting rabbits: Evolutionary significance of biological and morphological features. Parasitology 2008, 135, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Bull, P. Incidence of coccidia (Sporozoa) in wild rabbits, Oryctolagus cuniculus (L.) in Hawke's Bay, New Zealand. N. Z. J. Sci. 1958, 1, 289–328. [Google Scholar]
- Wang, J.; Tsai, S. Prevalence and pathological study on rabbit hepatic coccidiosis in Taiwan. Proc. Natl. Sci. Counc. Repub. China Part B Life Sci. 1991, 15, 240–243. [Google Scholar]
- Fang, S.; Gu, X.; El-Ashram, S.; Li, X.; Yu, X.; Guo, B.; Li, H.; Liu, N.; Liu, X.; Cui, P. Immune protection provided by a precocious line trivalent vaccine against rabbit Eimeria. Vet. Parasitol. 2019, 275, 108927. [Google Scholar] [CrossRef] [PubMed]
- Al-Mathal, E.M. Hepatic coccidiosis of the domestic rabbit (Oryctolagus cuniculus domesticus L.) in Saudi Arabia. World J. Zool. 2008, 3, 30–35. [Google Scholar]
- Hung, L.; Chen, F.; Lin, Y. Studies on the coccidia of rabbit, with a note on the endogenous stages and histopathology of Eimeria stiedae. Wuyi Sci. J. 1984, 4, 55–64. [Google Scholar]
- Vancraeynest, D.; De Gussem, M.; Marien, M.; Maertens, L. The anticoccidial efficacy of robenidine hydrochloride in Eimeria challenged rabbits. In Proceedings of the 9th World Rabbit Congress, Verona, Italy, 10–13 June 2008; pp. 10–13. [Google Scholar]
- Chapman, A.D. Numbers of Living Species in Australia and the World; Australian Government, Department of the Environment, Water, Heritage, and the Arts: Canberra, ACT, Australia, 2009.
- Abbas, R.; Colwell, D.; Gilleard, J. Botanicals: An alternative approach for the control of avian coccidiosis. World's Poult. Sci. J. 2012, 68, 203–215. [Google Scholar] [CrossRef]
- Shivaramaiah, C.; Barta, J.R.; Hernandez-Velasco, X.; Téllez, G.; Hargis, B.M. Coccidiosis: Recent advancements in the immunobiology of Eimeria species, preventive measures, and the importance of vaccination as a control tool against these Apicomplexan parasites. Vet. Med. Res. Rep. 2014, 5, 23. [Google Scholar]
- Garidel, P.; Hildebrand, A.; Knauf, K.; Blume, A. Membranolytic activity of bile salts: Influence of biological membrane properties and composition. Molecules 2007, 12, 2292–2326. [Google Scholar] [CrossRef] [Green Version]
- Zhai, C.; Tang, W.; Jang, X.; Lorenz, K. Studies of the safety of Chinese wild rice. Food Chem. Toxicol. 1996, 34, 347–352. [Google Scholar] [CrossRef]
- Begley, M.; Gahan, C.G.; Hill, C. The interaction between bacteria and bile. FEMS Microbiol. Rev. 2005, 29, 625–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binder, H.J.; Filburn, C.; Volpe, B.T. Bile salt alteration of colonic electrolyte transport: Role of cyclic adenosine monophosphate. Gastroenterology 1975, 68, 503–508. [Google Scholar] [CrossRef]
- de Aguiar Vallim, T.Q.; Tarling, E.J.; Kim, T.; Civelek, M.; Baldán, Á.; Esau, C.; Edwards, P.A. MicroRNA-144 regulates hepatic ABCA1 and plasma HDL following activation of the nuclear receptor FXR. Circ. Res. 2013, 112, 1602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullen, J.M.; Stalker, M.J. Liver and biliary system. Jubb Kennedy Palmer's Pathol. Domest. Anim. 2016, 2, 258. [Google Scholar]
- Li, S.; Tan, H.Y.; Wang, N.; Hong, M.; Li, L.; Cheung, F.; Feng, Y. Substitutes for bear bile for the treatment of liver diseases: Research progress and future perspective. Evid. Based Complement. Altern. Med. 2016, 2016, 4305074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zehua, L. Political and Intellectual Authority: The Concept of the “Sage-Monarch” and Its Modern Fate. Ideol. Power Power Ideol. Early China 2015, 273–300. [Google Scholar]
- Cohen, D.E.; Leonard, M.R.; Carey, M.C. In vitro evidence that phospholipid secretion into bile may be coordinated intracellularly by the combined actions of bile salts and the specific phosphatidylcholine transfer protein of liver. Biochemistry 1994, 33, 9975–9980. [Google Scholar] [CrossRef]
- Tan, D.-x.; Manchester, L.C.; Reiter, R.J.; Qi, W.; Hanes, M.A.; Farley, N.J. High physiological levels of melatonin in the bile of mammals. Life Sci. 1999, 65, 2523–2529. [Google Scholar] [CrossRef]
- Kou, G.; Zhang, C.; Wang, Z.; Xu, Q.; Wang, B. Study progress on pharmacology of bear bile powder. Chin. J. Drug Eval. 2014, 31, 142–145. [Google Scholar]
- Zhao, J.-Y.; Lin, W.; Zhuang, Q.-C.; Zhong, X.-Y.; Peng, J.; Hong, Z.-F. Bear bile powder inhibits angiogenesis in vivo and in vitro. Chin. J. Integr. Med. 2015, 21, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, S.; Romain, G.; Roux, M.; Chen, X.-L.; Brown, K.; Gibson, D.L.; Ramanathan, S.; Menendez, A. Bile acid administration elicits an intestinal antimicrobial program and reduces the bacterial burden in two mouse models of enteric infection. Infect. Immun. 2017, 85, e00942-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gumperz, J.E.; Litwin, V.; Phillips, J.H.; Lanier, L.L.; Parham, P. The Bw4 public epitope of HLA-B molecules confers reactivity with natural killer cell clones that express NKB1, a putative HLA receptor. J. Exp. Med. 1995, 181, 1133–1144. [Google Scholar] [CrossRef] [PubMed]
- Remmal, A.; Achahbar, S.; Bouddine, L.; Chami, F.; Chami, N. Oocysticidal effect of essential oil components against chicken Eimeria oocysts. Int. J. Vet. Med. 2013, 2, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.W.; Small, K.; Kashyap, S.; Deppen, J.G. Physiology, Gallbladder; StatPearls Publishing: Treasure Island, FL, USA, 2018. [Google Scholar]
- Haberkorn, A.; Stoltefuss, J. Studies on the activity spectrum of toltrazuril, a new anti-coccidial agent. VMR Vet. Med. Rev. 1987, 22–32. [Google Scholar]
- Zurliiski, P.; Vladiminova, A. Treatment of coccidiosis in rabbit with Baycox (tohrazuril). Vet. Sb. 1988, 86, 41. [Google Scholar]
- Singla, L.; Juyal, P.; Sandhu, B. Pathology and therapy in naturally Eimeria stiedae-infected rabbits. J. Protozool. Res. 2000, 10, 185–191. [Google Scholar]
- Schito, M.L.; Barta, J.R.; Chobotar, B. Comparison of four murine Eimeria species in immunocompetent and immunodeficient mice. J. Parasitol. 1996, 82, 255–262. [Google Scholar] [CrossRef]
- Heelan, J.S.; Ingersoll, F.W.; Ingersoll, F.W. Essentials of Human Parasitology; Delmar Publishers: Toebben Drive, KY, USA, 2002. [Google Scholar]
- Levine, N. The Protozoan Phylum Apicomplexa; Chemical Rubber Company Press, Inc.: Boca Raton, FL, USA, 1988; Volume 1. [Google Scholar]
- Gadelhaq, S.M.; Arafa, W.M.; Abolhadid, S.M. In vitro activity of natural and chemical products on sporulation of Eimeria species oocysts of chickens. Vet. Parasitol. 2018, 251, 12–16. [Google Scholar] [CrossRef]
- You, M.-J. Suppression of Eimeria tenella sporulation by disinfectants. Korean J. Parasitol. 2014, 52, 435. [Google Scholar] [CrossRef]
- Kubo, M.; Nakagami, T.; Yamasaki, N.; Ito, M.; Nogami, M.; Taji, S. Studies on Medicinal Resources from Livestock. I.: Anti-allergic Effects of Pig Bile. (1). Chem. Pharm. Bull. 1989, 37, 3409–3411. [Google Scholar] [CrossRef] [Green Version]
- Cedric, Y.; Payne, V.; Nadia, N.; Kodjio, N.; Kollins, E.; Megwi, L.; Kuiate, J. In vitro antioxidant and anticoccidial studies of pentaclethra macrophylla extracts against eimeria magna, eimeria flavescens, eimeria intestinalis eimeria stiedae oocysts and sporozoites of rabbits. J. Adv. Parasitol. 2018, 5, 38–48. [Google Scholar]
- Molan, A.L.; Liu, Z.; De, S. Effect of pine bark (Pious radiata) extracts on sporulation of coccidian oocysts. Folia Parasitol. 2009, 56, 1. [Google Scholar] [CrossRef]
- Jones, G.; McAllister, T.; Muir, A.; Cheng, K.-J. Effects of sainfoin (Onobrychis viciifolia Scop.) condensed tannins on growth and proteolysis by four strains of ruminal bacteria. Appl. Environ. Microbiol. 1994, 60, 1374–1378. [Google Scholar] [CrossRef] [Green Version]
- Hamad, K.K.; Iqbal, Z.; Abbas, R.Z.; Khan, A.; Muhammad, G.; Epperson, B. Combination of Nicotiana tabacum and Azadirachta indica: A Novel Substitute to Control Levamisole and Ivermectin-Resistant Haemonchus contortus in Ovine. Pak. Vet. J. 2014, 34, 24–29. [Google Scholar]
- Hofmann, A.F.; Hagey, L.R.; Krasowski, M.D. Bile salts of vertebrates: Structural variation and possible evolutionary significance [S]. J. Lipid Res. 2010, 51, 226–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, E.; Pozdeev, V.I.; Gatidis, S.; Qadri, S.M.; Häussinger, D.; Kubitz, R.; Herebian, D.; Mayatepek, E.; Lang, F.; Lang, K.S. Bile acid-induced suicidal erythrocyte death. Cell. Physiol. Biochem. 2016, 38, 1500–1509. [Google Scholar] [CrossRef] [PubMed]
- Liu, C. Review on the studies of unearthed Mawangdui medical books. Chin. Stud. 2016, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Carey, M. DUANE WC-Enterohepatic circulation. Liver: Biol. Pathobiol. 1994, 3, 719–786. [Google Scholar]
- Sjövall, J.; Setchell, K. Techniques for extraction and group separation of bile acids. In The Bile Acids: Chemistry, Physiology, and Metabolism; Springer: Berlin/Heidelberg, Germany, 1988; pp. 1–42. [Google Scholar]
- Lefebvre, P.; Cariou, B.; Lien, F.; Kuipers, F.; Staels, B. Role of bile acids and bile acid receptors in metabolic regulation. Physiol. Rev. 2009, 89, 147–191. [Google Scholar] [CrossRef] [Green Version]
- Murshed, M.; Al-Quraishy, S.; Qasem, M.A. In vitro: Anti-coccidia activity of Calotropis procera leaf extract on Eimeria papillata oocysts sporulation and sporozoite. Open Chem. 2022, 20, 1057–1064. [Google Scholar] [CrossRef]
- Khalafalla, R.E.; Müller, U.; Shahiduzzaman, M.; Dyachenko, V.; Desouky, A.Y.; Alber, G.; Daugschies, A. Effects of curcumin (diferuloylmethane) on Eimeria tenella sporozoites in vitro. Parasitol. Res. 2011, 108, 879–886. [Google Scholar] [CrossRef]
- Murshed, M.; Al-Quraishy, S.; Qasem, M. The Anticoccidial Effects of the Poultry Bile against Eimeria papillata Oocysts of Mice: In vitro Study. Indian J. Anim. Res. 2022, 1, 8. [Google Scholar] [CrossRef]
- Schubert, U.; Fuchs, J.; Zimmermann, J.; Jahn, D.; Zoufal, K. Extracellular calcium deficiency and ryanodine inhibit Eimeria tenella sporozoite invasion in vitro. Parasitol. Res. 2005, 97, 59–62. [Google Scholar] [CrossRef]
- Sárközi, S.; Almássy, J.; Lukács, B.; Dobrosi, N.; Nagy, G.; Jóna, I. Effect of natural phenol derivatives on skeletal type sarcoplasmic reticulum Ca2+-ATPase and ryanodine receptor. J. Muscle Res. Cell Motil. 2007, 28, 167–174. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Absorption (cm−1) | Appearance | Transmittance (%) | Group | Compound Class |
|---|---|---|---|---|
| 3425.53 | Medium | 12 | N-H stretching | Aliphatic primary amine |
| 2093.10 | Strong | 47 | N = C = S stretchy | Isothiocyanate |
| 1641.41 | Strong | 25 | C = C stretching | Alkene |
| 1209.12 | Strong | 37 | C-O stretching tertiary | Alcohol |
| 1045.64 | Strong and broad | 35 | CO-O-CO stretching | Anhydride |
| 410.42 | Strong | 3 | C-H bending | 1,2-disubstituted |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murshed, M.; Al-Quraishy, S.; Qasem, M.A. Evaluation of the Anticoccidial Activity of Sheep Bile against Eimeria stiedae Oocysts and Sporozoites of Rabbits: An In Vitro Study. Vet. Sci. 2022, 9, 658. https://doi.org/10.3390/vetsci9120658
Murshed M, Al-Quraishy S, Qasem MA. Evaluation of the Anticoccidial Activity of Sheep Bile against Eimeria stiedae Oocysts and Sporozoites of Rabbits: An In Vitro Study. Veterinary Sciences. 2022; 9(12):658. https://doi.org/10.3390/vetsci9120658
Chicago/Turabian StyleMurshed, Mutee, Saleh Al-Quraishy, and Mahmood A. Qasem. 2022. "Evaluation of the Anticoccidial Activity of Sheep Bile against Eimeria stiedae Oocysts and Sporozoites of Rabbits: An In Vitro Study" Veterinary Sciences 9, no. 12: 658. https://doi.org/10.3390/vetsci9120658