

Impact of 2,4-D and Glyphosate on Soil Enzyme Activities in a Resistant Maize Cropping System

Crop Production Systems Research Unit, USDA, Agricultural Research Service, Stoneville, MS 38776, USA
Agronomy 2022, 12(11), 2747; https://doi.org/10.3390/agronomy12112747
Submission received: 27 September 2022
/
Revised: 29 October 2022
/
Accepted: 2 November 2022
/
Published: 5 November 2022
(This article belongs to the Special Issue Impact of Agrochemicals on Soil)
Abstract
:Crop varieties resistant to multiple herbicides have been developed to provide better control of weed populations in row-crop fields where glyphosate resistance has become common. These new varieties include lines of maize (Zea mays) resistant to both glyphosate and 2,4-dichlorophenoxyacetic acid (2,4-D). As these herbicides have the potential to impact microbial communities in soil, there is concern that their co-application may have a greater effect on activities linked to soil nutrient cycling than if they were applied individually. To investigate this possibility, a field study was conducted on 2,4-D+glyphosate-resistant maize to determine the impact of 2,4-D alone and 2,4-D+glyphosate on extracellular enzyme activity in both bulk and rhizosphere soil. Maize was treated at the V2 and V8 developmental stages. Changes in soil activities were small in magnitude and inconsistent between timepoints. 2,4-D+glyphosate-treated plots had higher beta-glucosidase, cellobiohydrolase, and phosphatase activities, but only after the V2 application in bulk soil in the first year of the study, while no significant effects were observed in the rhizosphere. Enzyme activities were more impacted by soil organic matter than herbicide treatments. These results suggest that, when applied at label rates, 2,4-D+glyphosate application will not adversely affect soil microbial enzyme activities.
1. Introduction
Overreliance on glyphosate application for weed control in Roundup Ready® cropping systems over the last 25 years has led to the widespread prevalence of weeds resistant to this herbicide. In response, crop varieties have been developed stacked with multiple herbicide-resistance traits, in order to provide farmers with more weed-control options. Examples of this include glufosinate+glyphosate-resistant crops (LibertyLink®, BASF, Florham Park, New Jersey, USA), introduced in the mid-2000s, dicamba+glyphosate-resistant crops (Roundup Ready 2 Xtend®, Bayer, Whippany, New Jersey, USA), introduced in 2017, and 2,4-dichlorophenoxyacetic acid (2,4-D)+glyphosate-resistant crops (Enlist™, Corteva Agriscience, Indianapolis, Indiana, USA), introduced in 2019 [1]. The increased application of these herbicides during the growing season has led to questions about their secondary, non-target effects on the ecosystem, including the soil microbial community, which serves several functions in the field. One such critical function is the mediation of nutrient cycling and organic matter turnover in the soil.
Glyphosate application has the potential to alter soil microbial communities that carry out the above functions. Glyphosate inhibits 5-enolpyruvylshikimic acid-3-phosphate synthase (EPSP), which blocks aromatic amino acid synthesis in plants [2]. Some bacterial EPSP are also sensitive to glyphosate application [3,4]. Consistent with this, soil microbial biomass has been found to decrease in response to glyphosate application [5]. However, some soil microbes can degrade glyphosate as a nutrient source [6]. Therefore, application of glyphosate also has the potential to increase the growth of soil microorganisms capable of metabolizing it. As such, there are also multiple studies reporting enhanced microbial biomass levels in soils following glyphosate application [7,8]. However, other studies report no effect of glyphosate on microbial biomass in bulk or rhizosphere soils [9,10,11]. Of even greater importance is the effect that herbicide application may have on soil functions involved in nutrient cycling. These functions are often estimated in soil using assays of extracellular enzymes secreted by soil bacteria and fungi. Both short- and long-term glyphosate application has been found to enhance hydrolytic activities in soil, as indicated by increased rates of fluorescein diacetate (FDA) hydrolysis, which was observed in two Brazilian soils [12]. However, studies in Mississippi have reported no effects on FDA hydrolysis in maize-field soils following glyphosate application at recommended label rates [13]. Looking at specific activities, glyphosate has the potential to inhibit phosphatase [14,15], although this response is not universal, and phosphatase is often left unaffected after glyphosate application [8,11,13,16]. Glyphosate application has also been found to decrease the activity of N-acetylglucosaminidase, which is involved in C and N cycling, in the wheat rhizosphere [11], and lowers the abundance of ammonia oxidizing archaea and bacteria in bulk soil from sugarcane fields [17].
Application of 2,4-D in fields also has the potential to alter soil microbial community dynamics. As with glyphosate, some bacteria are capable of using 2,4-D as a carbon (C) source [18]. Conversely, this herbicide has also been found to have toxic effects on the growth of some common soil bacteria, including Azospirillum brasilense Cd [19] and some Rhizobium sp. [20], and exposure to high doses can decrease culturable bacteria in soils up to 15 days after spraying [21]. 2,4-D has the opposite effect on soil fungi, with high concentrations increasing the fungal population size [21]. In contrast to glyphosate, research on the effects of 2,4-D on soil nutrient-cycling activities is less extensive. In one study, 2,4-D was found to inhibit phosphatase activities in soil [8]. Meanwhile, 2,4-D has been found to inhibit nitrogenase activity in Klebsiella pneumoniae [22] and Nostoc muscorum Meg 1 [23], while stimulating it in Enterobacter agglomerans [22]. A similar trend was observed among nitrifying bacteria, where 2,4-D treatment inhibited autotrophic nitrifiers while stimulating heterotrophic nitrifiers [24]. The variability in bacterial sensitivity to 2,4-D could be a result of differences in antioxidative response pathways, with efficient hydrogen peroxide production being linked to tolerance to high levels of 2,4-D in certain Pseudomonas species [25]. As such, the response of the nutrient-cycling capabilities in soil may vary by location, based on differences in the microbial community.
There are fewer studies on the effects of the combined application of both 2,4-D and glyphosate. Given that both these herbicides have the potential to impact soil microbial activities, there is a possibility of synergistic effects when applied together. A greenhouse study indicated that 2,4-D + glyphosate had little to no effect on the activities of phosphatase, β-glucosidase, N-acetylglucosaminidase, and cellobiohydrolase in the soybean rhizosphere [26]. However, bulk soil and the rhizosphere of other crops may respond differently. These initial studies were conducted in the greenhouse to provide uniform conditions and minimize variation in enzyme response due to environmental factors. However, greenhouse studies cannot replicate all conditions experienced in the field, including exposure to wind, rain, insects, and other factors. Plant-associated microbial community composition has also been found to differ between greenhouse vs field grown plants [27]. In addition, to prevent herbicide contamination of other experiments, greenhouse-grown plants must be removed from their temperature-controlled environment and transported to alternate sites for herbicide application, introducing a potential stress to the plants that might impact results of rhizosphere assessments. As such, field-level studies are important to confirm the potential impacts of new herbicide-application regimes on soil microbial functions under real world conditions. Therefore, the purpose of the current study was to assess effects of glyphosate and 2,4-D application on microbial potential for nutrient cycling in both bulk and maize rhizosphere soil at the field scale in a multiyear study.
2. Materials and Methods
2.1. Site Description, Experimental Design, and Plot Management
The two-year field study was located on a research farm in Stoneville, MS. Weekly precipitation and air temperature during the study period were obtained from the Mississippi State University Delta Agricultural Weather Center [28] and are reported in Figure 1. Experimental plots were sown with maize resistant to 2,4-D and glyphosate (B14U78 SXE Brevant) and set up in a randomized complete block design with four blocks. Blocks contained one plot (32 m × 8.4 m) per treatment, for a total of three plots per block. Treatments were: 2,4-D alone (24D), 2,4-D in combination with glyphosate [N-(phosphonomethyl) glycine] (24DG), or no herbicide control (NH). Soil in the field is a Commerce very fine sandy loam [29]. Application rate for the 24D treatment was 2.3 L ha−1 Enlist One® (Corteva Agriscience, Indianapolis, IN, USA), and the application rate for 24DG was 2.3 L ha−1 Enlist One® and 2.3 L ha−1 Makaze® (Loveland Products, Greeley, Colorado, USA). These application rates translate to active-ingredient equivalent of 1 kg ha−1 of 2,4-D and 0.82 kg ha−1 of glyphosate. Herbicide treatments were applied at two timepoints: when maize reached the V2 and V8 developmental stages. The V2 timepoint was selected to assess effects of herbicide application during an early vegetative stage. The V8 vegetative stage was selected for the second application, as it is the latest developmental stage that Enlist One® can be applied according to label instructions. Timelines for maize sowing, herbicide treatments, and maize harvest are outlined in Table 1. The first year of the study was initiated in 2019, but the second year was delayed until 2021 due to labor shortage in 2020.
2.2. Sample Collection
Bulk soil samples were collected at sowing, one week after each herbicide application, and after maize harvest in the fall. Eight replicate soil cores were collected from each plot, divided into 0–5 and 5–15 cm depths, and mixed to form one composite soil sample per depth per plot. Rhizosphere soil was collected one week after each herbicide application. Briefly, five to six plants were randomly selected per plot and their root systems excavated, placed in Ziplock bags, and transported back to the laboratory on ice. The soil-root mass was gently massaged and shaken to dislodge loosely-adhered bulk soil. Then, the root systems were transferred to presterilized 500 mL beakers where they were submerged in approximately 100 mL of sterile deionized water and shaken to release rhizosphere soil from the roots. Plant debris was removed from the rhizosphere–water mixture by pouring through flame-sterilized 2 mm sieves. Rhizosphere soil was collected from the mixture by centrifuging at 3220× g for 15 min and discarding the supernatant.
2.3. Soil Sample Analyses
Bulk soils were assessed for soil moisture (SM) content by oven-drying aliquots of 5 g from each sample overnight at 105 °C, in triplicate, and calculated as the percent weight lost from drying. Soil organic matter (SOM) was determined by ashing oven-dried soil aliquots in a muffle furnace at 500 °C for 2 hr, and reported as the percent of weight lost due to ashing. Soil pH was determined by mixing equal parts of soil and water (1 g mL−1) on a magnetic stir plate and measuring using an electrode pH meter. Fluorescein diacetate (FDA) hydrolysis was determined in bulk soils using a 96-well plate protocol [30] and reported as nmole fluorescein produced per g dry weight of soil per hour. Activities of beta-glucosidase, cellobiohydrolase, N-acetyleglucosaminidase (NAGase), and phosphatase were assessed in both bulk and rhizosphere soils using p-nitrophenol (pNP)-linked substrates in a 96-well plate format [31], with modifications for rhizosphere soil. Briefly, pelleted rhizosphere soil was resuspended in 50 mM acetate buffer (pH 5) by vortexing to produce a soil slurry for assays, similar to those used in Jackson et al. [31]. The volume of buffer varied based on the developmental stage of the plant and the concomitant size of the rhizosphere pellet, with approximately 20 mL of buffer added to pellets from the post-V2 application samples and 50 mL of buffer added to the post-V8 application samples. All pNP-based enzyme activities were calculated as μmoles of substrate consumed per g dry weight per hour.
2.4. Statistics
Two-way analysis of variance (ANOVA) was used to determine significant differences between treatments and depth at each timepoint for soil chemical characteristics and enzyme activities. Differences between specific treatment effects detected by ANOVA were confirmed using a Tukey’s Honestly Significant Difference (HSD) post hoc test, with an alpha of 0.05. Relationships between soil chemical characteristics and enzyme activities were determined by Pearson correlations. All analyses were conducted in JMP version 15.0.0 (SAS Institute Inc., Carey, NC, USA).
3. Results and Discussion
3.1. Effects of 2,4-D and Glyphosate on Bulk Soil
Levels of SOM, pH, and SM all varied between timepoints throughout the study (Table 2). SOM was higher in 0–5 cm soil (p ≤ 0.0215), but did not differ between treatments at any timepoint (Table 2). Soil pH did not vary by depth or treatment at sowing or harvest timepoints either year, but was higher in 24DG than in NH control plots after the second year’s V2 application (p = 0.0416), and varied by depth at V2 in year 2 and V8 in both years of the study (Table 2; p ≤ 0.0027). SM did not vary by treatment and was greater in 5–15 than 0–5 cm soil in all but the sowing and V2 timepoints in year 1 (Table 2; p ≤ 0.0324).
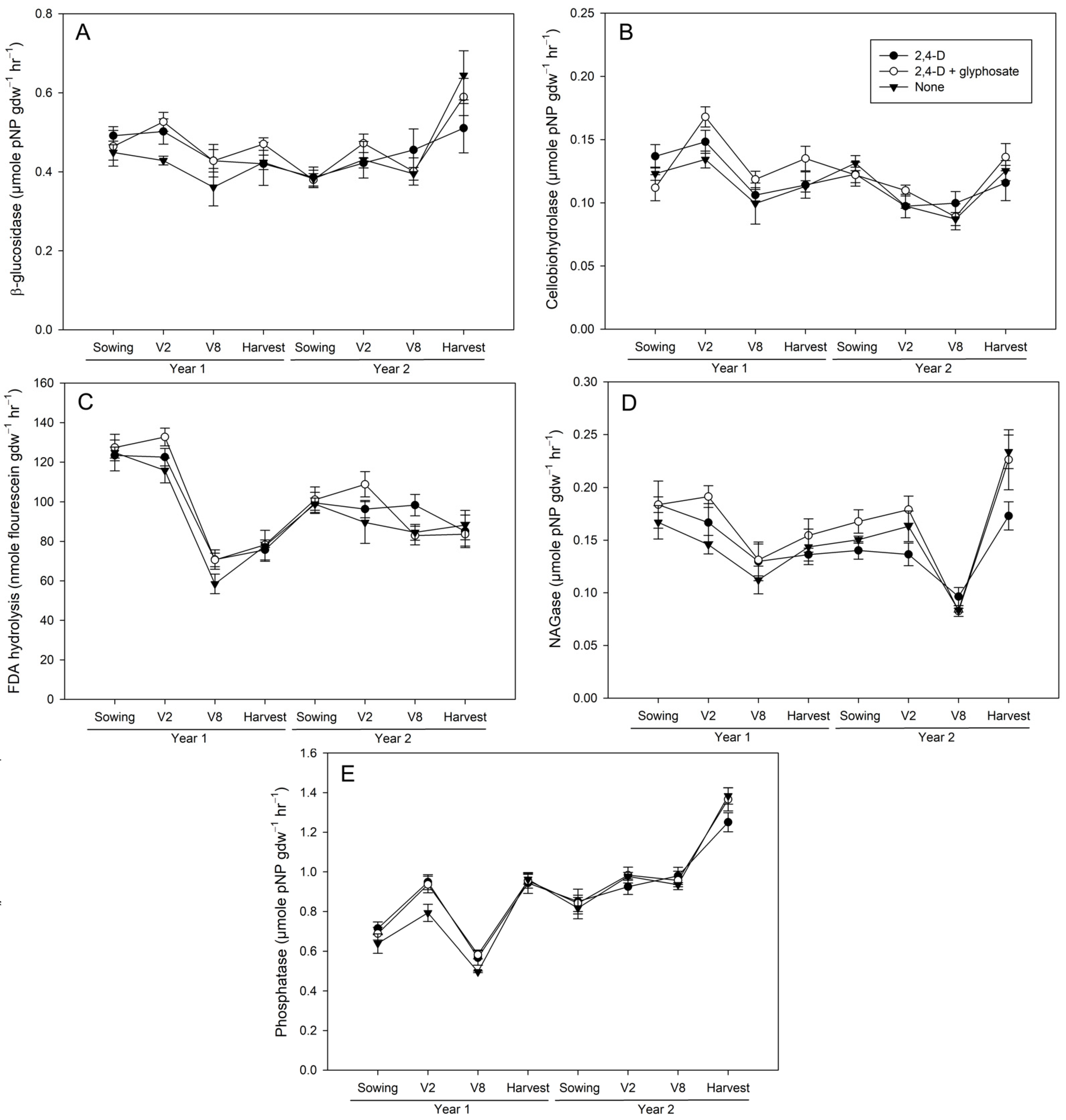
Activities of all enzymes assayed also varied across timepoint and year. Analyses of baseline soil samples collected at sowing the first year of the study found no significant differences in bulk soil activities from either 0–5 or 5–15 cm depths, indicating that variability at the plot level is not correlated with the treatment assigned to each plot. As such, subsequent differences were attributed to the result of herbicide treatments. Most differences in activity were observed in 0–5 cm soil during the first year of the study, following the V2 application. Two-way ANOVA indicated a significant interaction between depth and treatment (p ≤ 0.0373), where 24DG plots had significantly higher levels of beta-glucosidase (p = 0.04) and cellobiohydrolase (p = 0.0081) compared to NH plots in 0–5 cm soil at this timepoint (Figure 2A,B). In the case of phosphatase, there was no interaction with depth, but a treatment effect, indicating both 24D (p = 0.0142) and 24DG (p = 0.0286) had significantly higher phosphatase activities than NH plots (Figure 2E). Meanwhile, FDA hydrolysis only differed between treatments at the year 1 V8 timepoint, when two-way ANOVA indicated a treatment effect where 24D had higher activity than NH controls (p = 0.0302; Figure 2C and Figure 3C). While FDA hydrolysis in 24DG also appeared higher than NH controls in 0–5 cm soil at the V8 timepoint (Figure 2C), this difference was not significant at an alpha of 0.05. NAGase was the only enzyme to be significantly affected by herbicide treatment in the second year of the study at the V8 timepoint. There was no significant interaction with depth, but a treatment effect where 24D had higher activity than 24DG (p = 0.0304; Figure 2D and Figure 3D). Plots treated with 24D also appeared to have higher NAGase activity than NH controls, but this difference was not significant. While 24DG appears to have higher cellobiohydrolase activity at the year 1 harvest timepoint (Figure 2B and Figure 3B), Tukey’s HSD indicates this difference is not significant.
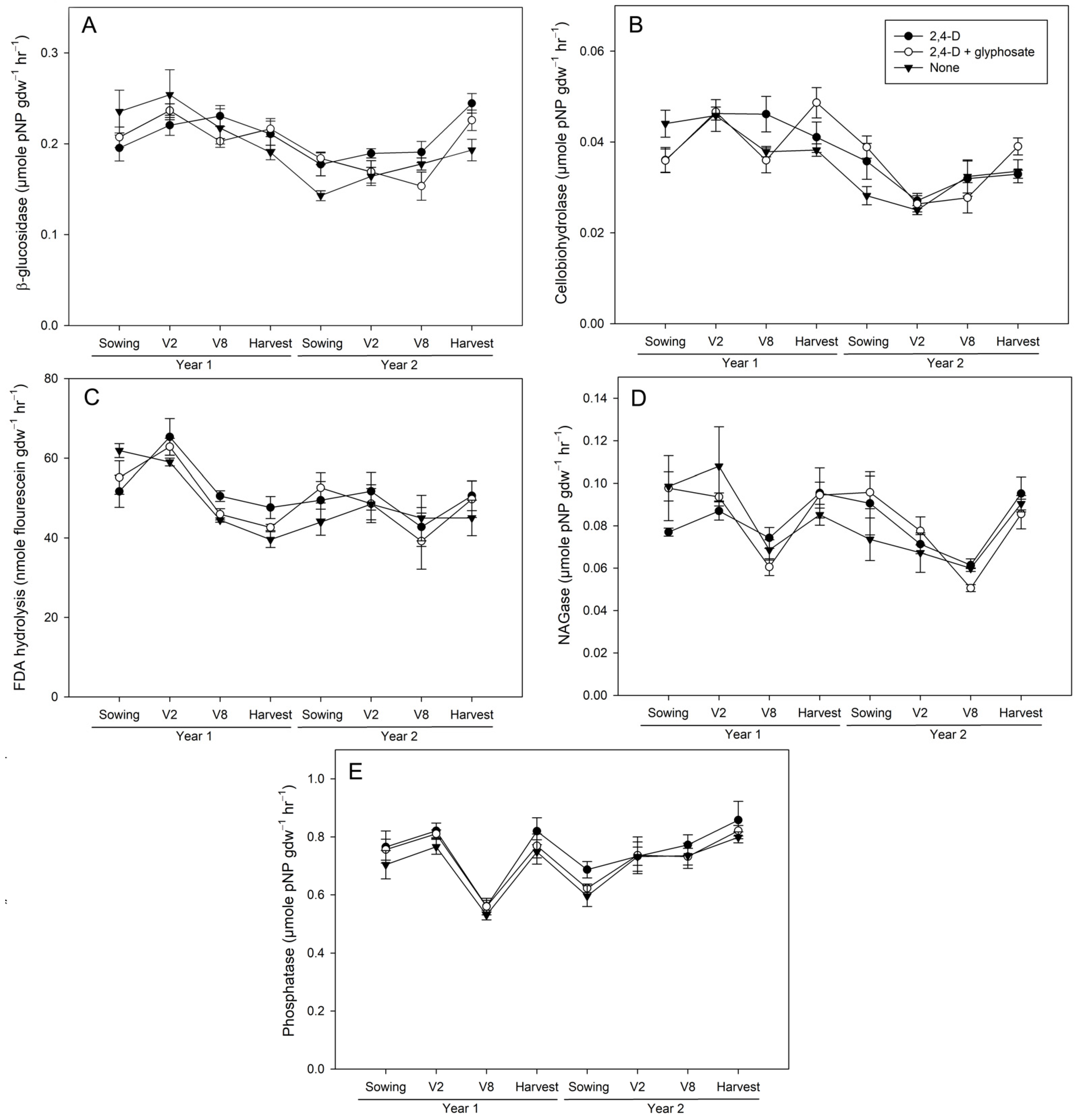
Activities in 5–15 cm soil were significantly lower and more variable than those observed in 0–5 cm soil across all timepoints, with the exception of phosphatase, which did not vary by depth at pre-sowing or post-V8 in year 1. As such, there were fewer treatment-related differences in this depth, all of which corresponded with treatment effects that were also observed in 0–5 cm soils. Most significant differences in the 5–15 cm depth occurred at the post-V8 timepoint, where 24D alone had higher FDA hydrolysis than NH control plots (p = 0.0302; Figure 3C) and higher NAGase than 24DG (p = 0.0304; Figure 3D). Only phosphatase was impacted following the V2 application at this depth, with higher activities in both herbicide treatments compared to NH control plots (p ≤ 0.0286; Figure 3E). The greater number of differences in 5–15 cm soils at the later V8 could potentially be due to transport of the herbicide through the soil profile as a result of subsequent precipitation events.
The observation of more differences occurring after the V2, rather than the V8, applications in the 0–5 cm depth was surprising, as the post-V8 application timepoints were expected to display greater differences between treatments, owing to the plots being exposed to an additional herbicide application at that point. One possible reason for this discrepancy could be due to seasonal weather differences, as the week prior to the V2 application received nearly double the amount of precipitation as the week prior to the V8 applications each year (Figure 1). However, correlations between SM and soil enzyme activities were low, suggesting other factors also contributed (Table 3). Soil pH only varied between treatments after the second year’s V2 application, being higher in 24DG than NH controls, which did not correspond with significant differences in soil activities at this time point. In addition, the correlations between soil pH and enzyme activities were relatively weak, only being significant with cellobiohydrolase (r = −0.1551) and phosphatase (r = 0.2219; Table 3). SOM appears to be a major factor contributing to enzyme activities, with all enzymes significantly correlating with SOM (Table 3). SOM did not differ by treatment during the study period, but was higher in 0–5 than 5–15 cm soil (Table 2), which is consistent with differences in activities observed between depths. Many of the enzymes assayed are involved in organic-matter breakdown, and organic matter and substrate availability are primary drivers of soil enzyme activities across a variety of ecosystems [32,33,34,35]. As such, having similar levels of SOM as substrate across all plots in the experiment may have masked herbicide treatment-related responses.
While only a few timepoints had significant differences between treatments, when differences were observed, there was a general trend for 24DG-treated plots to have higher activities than NH controls. This observation is consistent with the potential for these herbicides to be used as C, nitrogen (N), or phosphorus (P) sources. While these herbicides both have the potential to inhibit members of the soil microbial community, the concentrations usually used in field applications are not high enough to elicit toxic responses. In the case of 2,4-D butyl ester, Zhang et al. [36] found that high concentrations (1000 µg g−1) decreased the amount of culturable bacteria in soil, while a lower concentration (10 and 100 µg g−1) stimulated it 8 days after application. The rate of 2,4-D applied in the current study (10 µg cm−2) is comparable to the lower end of the range applied by Zhang et al. [36], who found that stimulation of bacterial abundance at this rate was lower and less persistent than the 100 µg g−1 rate. This might explain why responses in the current study were low and variable. Of the two herbicide treatments in the current study, 24DG tended to have higher activities than 24D, possibly due to additive effects of both herbicides. The general weak response of phosphatase observed in the current study is consistent with Nowak et al. [37], who found that herbicide formulations containing 2,4-D and dicamba did not significantly impact acid phosphatase activities in soil when applied at label rates, only impacting activities when applied at five and 25 times the label rate.
As previously mentioned, several enzymes assayed in the current study are associated with organic-matter breakdown, with beta-glucosidase and cellobiohydrolase each involved in the degradation of cellulose and considered a measure of organic-matter turnover [38], NAGase correlated with N mineralization [39], and phosphatase linked to P mineralization [40]. Meanwhile, FDA hydrolysis is considered a measure of heterotrophic activity, as it is a substrate for several hydrolytic enzymes, such as esterases, lipases, and proteases [41]. FDA hydrolysis correlated significantly with all other activities assayed in the current study, suggesting shifts in these enzymes were largely a result of changes in overall microbial activity. Dennis et al. [42] found that glyphosate did not significantly impact activities of FDA hydrolysis, beta-glucosidase, or the ability to use 15 other substrates 7 days after application, with the exception of a temporary inhibition of the ability to metabolize three organic acids (malic, oxalic, and citric acid). Within the same region where the current study was conducted, Zablotowicz et al. [43] found that glyphosate did not affect FDA hydrolysis in 0–5 cm soil at 14 days after a second application of glyphosate in a soybean field. Results of the current study varied slightly from Zablotowicz et al. [43], noting a small number of significant differences in FDA activity between treatments, but those may have been due to differences in sampling time (7 versus 14 days after herbicide application), or slight difference in soil texture as, Zablotowicz et al. [43] assessed a silt loam, while the current study was conducted in a field with very fine sandy loam.
In another Mississippi Delta field study on maize, Jenkins et al. [13] reported activities of FDA hydrolysis, beta-glucosidase, and NAGase were largely unresponsive to glyphosate application, with the few effects observed being temporary and inconsistent across years. These results are consistent with observation of 24DG in the current study, although the differences observed currently were greater in magnitude and across more enzymes than those observed by Jenkins et al. [13]. While each of these studies performed two applications, Jenkins et al. [13] performed their assessments at a single point after both applications, when maize was at the R2 developmental stage, while the current study assessed activities one week after each application, when maize was at earlier stages of development (V2 and V8), as well as post-harvest. However, similar inconsistencies were observed in the response of soil activities to 24DG application as those seen by Jenkins et al. [13] to glyphosate only, with most differences occurring closer to the time of herbicide application. Taken together, these observations suggest that combined application of 2,4-D and glyphosate does not have a substantially greater impact on microbial activities in soil than glyphosate alone.
There are still relatively few studies that have examined the effects of both 2,4-D and glyphosate on activities in bulk soil. In one such study, Zabaloy et al. [44] found that glyphosate alone, or glyphosate with 2,4-D, did not significantly impact substrate-induced respiration (SIR) or dehydrogenase activity (DHA) in soil. Earlier work found that application of these individual herbicides at 10 times the normal rate only resulted in minor and temporary shifts in activity, with 2,4-D inhibiting SIR and FDA while stimulating DHA [45]. Lupwayi et al. [46] found that pre-sowing application of 24DG decreased functional diversity of bulk soil collected in canola fields at flowering stage, but only in one out of two years. Glyphosate application has also been found to decrease nitrification, as well as nitrate- and nitrite-consumption rates, in soil form sugarcane fields [47]. Martens and Bremner [48] also noted inhibition of urea nitrification by 2,4-D and glyphosate. These observations of inhibition are inconsistent with the current study, where the few differences in activity indicated stimulation. In the case of Martens and Bremner [48], they found the inhibition occurred in coarse-, but not fine-textured, soils, and attributed the inhibition to low C content in coarse soil. Soil C in the current study ranged from approximately 18.2–23.4 kg g−1 (calculated from SOM), which is well above the level of the low-SOC soils assessed by Martens and Bremner [48]. As such, the lack of inhibition following herbicide application in the current study could be due to higher soil C levels. In addition, the current study examined activities involved in organic matter mineralization, rather than nitrification, with these activities significantly correlating with SOM, suggesting stimulation of activities due to substrate availability may have overcome any inhibitory effects of the herbicides.
3.2. Activities in Rhizosphere Soil from 2,4-D and Glyphosate Treated Maize
Activities in the maize rhizosphere tended to be elevated compared to those in bulk soils, ranging from 1.8- to 5.9-fold greater, in 0–5 cm, to 3.6- to 14.6-fold higher, in 5–15 cm soils (Table 4 vs. Figure 2 and Figure 3). A similar trend of higher activities in the rhizosphere compared to bulk soil was also observed by Jenkins et al. [13]. This difference is likely due to nutrients deposited into the rhizosphere from root exudates. None of the activities in the rhizosphere differed between treatments after the V2 or V8 applications in either year (Table 4). This lack of response in the maize rhizosphere is similar to observations of the soybean rhizosphere, where neither NAGase or cellobiohydrolase activities responded to 2,4-D or glyphosate up to a month after application [26]. While a small increase in beta-glucosidase and phosphatase activities was noted in the rhizosphere of soybeans treated with 24DG, it was only at a single timepoint for each enzyme (7- and 14-days post application, respectively) and not consistent across different soil-texture applications [26]. However, a study by Nandula and Tyler [26] was conducted on greenhouse-grown soybeans, where a more controlled environment would have introduced fewer environmental factors and variability in soil conditions, which may have made small, transient shifts in enzyme activity more easily discernable compared to field conditions in the current study. In a field study on maize, Jenkins et al. [13] also found that activities in the maize rhizosphere were largely unresponsive to glyphosate application. In the wheat rhizosphere, Lupwayi et al. [11] found glyphosate had no significant effects on beta-glucosidase or acid phosphatase, which is consistent with the current study. However, they did note NAGase activity responded to glyphosate application, although the response varied according to application rate, with both low and high rates inhibiting activity (1 and 8 kg ae ha−1, respectively), while middle application rates (2 and 4 kg ae ha−1) did not differ from untreated controls [11]. Lupwayi et al. [46] also observed no differences in functional diversity of the canola rhizosphere in response to 24DG application compared to each herbicide applied individually, although this study did not include an NH control. Similar to the current study assessing individual enzyme activities, they also noted that there were fewer significant differences in functional diversity between herbicide treatments than non-significant differences [46].
Newman et al. [49] reported a number of microbial metabolic pathways were either up- or down-regulated in the rhizosphere of glyphosate-treated maize and soybean, including those involved in amino acid synthesis and C metabolism, suggesting that glyphosate could affect bacterial activities in the rhizosphere. This observation contrasts with the observations of the current study, as well as Jenkins et al. [13] and Nandula and Tyler [26], where effects of glyphosate application were inconsistent and small in magnitude. However, Newman et al. [49] only examined RNA transcripts, and differences at this level may not be enough to translate into altered activities at the functional level. The differences they observed may not be involved in synthesis of the enzymes assayed in the current study. Additionally, high substrate availability from nutrients in root exudates may have also masked any herbicide induced shifts in activity.
4. Conclusions
Assessment of combined applications of glyphosate and 2,4-D in resistant cropping systems has been limited. The few studies that have examined the combined effects of these herbicides were conducted in greenhouse or microcosm settings, or assessed only pre-sowing application. To the author’s knowledge, the current study is the first to assess the effects of 24DG in field conditions during the growing season. The application of 24DG tended to have a greater impact on microbial activities in bulk soil than 24D. However, this response was highly variable between timepoints and years, relatively small in magnitude, and tended to be stimulatory in nature. Variation between timepoints is likely due to climate-induced changes in soil conditions. The stimulation seen in response to glyphosate and 2,4-D application could be the result of soil bacteria capable of utilizing these herbicides as an additional C or N source. However, the concentration of C and N from herbicide applications is small, relative to the levels of organic matter in the soils. In comparison to bulk soil, activities in the maize rhizosphere were unchanged, even after multiple applications. The lack of response in the rhizosphere was attributed to greater substrate availability from high nutrient concentration in root exudates, stimulating enzyme production by soil bacteria and fungi to a level that masked any herbicide-based effects. Given the results from this study, along with the inconsistent nature of responses seen in other studies of both glyphosate and 2,4-D, combined application 2,4-D and glyphosate is unlikely to have a significant impact on soil enzymes involved in SOM processing, especially when used at recommended field rates.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The author would like to thank Paige Goodlett for assistance in collecting and processing soil samples. This research was supported by the U.S. Department of Agriculture, Agricultural Research Service. Mention of trade names or commercial products is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the USDA. The findings and conclusions in this publication are those of the author and should not be construed to represent any official USDA or U.S. Government determination or policy. USDA is an equal-opportunity provider and employer.
Conflicts of Interest
The author declares no conflict of interest.
References
- Nandula, V.K. Herbicide Resistance Traits in Maize and Soybean: Current Status and Future Outlook. Plants 2019, 8, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sikorski, J.A.; Gruys, K.J. Understanding glyphosate’s molecular mode of action with EPSP Synthase: Evidence favoring an allosteric inhibitor model. Acc. Chem. Res. 1997, 30, 2–8. [Google Scholar] [CrossRef]
- Moorman, T.B.; Becerril, J.M.; Lydon, J.; Duke, S.O. Production of hydroxybenzoic acids by Bradyrhizobium japonicum strains after treatment with glyphosate. J. Agric. Food Chem. 1992, 40, 289–293. [Google Scholar] [CrossRef]
- Fischer, R.S.; Berry, A.; Gaines, C.G.; Jensen, R.A. Comparative action of glyphosate as a trigger of energy drain in Eubacteria. J. Bacteriol. 1986, 168, 1147–1154. [Google Scholar] [CrossRef] [Green Version]
- Gomez, E.; Ferreras, L.; Lovotti, L.; Fernandez, E. Impact of glyphosate application on microbial biomass and metabolic activity in a Vertic Argiudoll from Argentina. Eur. J. Soil Biol. 2009, 45, 163–167. [Google Scholar] [CrossRef]
- Dick, R.E.; Quinn, J.P. Glyphosate-degrading isolates from environmental samples: Occurrence and pathways of degradation. Appl. Microbiol. Biotechnol. 1995, 43, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Haney, R.L.; Senseman, S.A.; Hons, F.M. Effect of roundup ultra on microbial activity and biomass from selected soils. J. Environ. Qual. 2002, 31, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Joshi, N. Changes in microbial biomass and phosphatase activity exposed to 2,4-D and glyphosate. J. Environ. Res. Dev. 2009, 3, 663–669. [Google Scholar]
- Haney, R.L.; Senseman, S.A.; Hons, F.M.; Zuberer, D.A. Effect of glyphosate on soil microbial activity and biomass. Weed Sci. 2000, 48, 89–93. [Google Scholar] [CrossRef]
- Liphadzi, K.B.; Al-Khatib, K.; Bensch, C.N.; Stahlman, P.W.; Dille, J.A.; Todd, T.; Rice, C.W.; Horak, M.J.; Head, G. Soil microbial and nematode communities as affected by glyphosate and tillage practices in a glyphosate-resistant cropping system. Weed Sci. 2005, 53, 536–545. [Google Scholar] [CrossRef]
- Lupwayi, N.Z.; Blackshaw, R.E.; Geddes, C.M.; Dunn, R.; Petri, R.M. Multi-year and multi-site effects of recurrent glyphosate applications on the wheat rhizosphere microbiome. Environ. Res. 2022, 215, 114363. [Google Scholar] [CrossRef]
- Araújo, A.S.F.; Monteiro, R.T.R.; Abarkeli, R.B. Effect of glyphosate on the microbial activity of two Brazilian soils. Chemosphere 2003, 52, 799–804. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, M.B.; Locke, M.A.; Reddy, K.N.; McChesney, D.S.; Steinriede, R.W. Impact of glyphosate-resistant corn, glyphosate applications and tillage on soil nutrient ratios, exoenzyme activities and nutrient acquisition ratios. Pest Manag. Sci. 2017, 73, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Sannino, F.; Gianfreda, L. Pesticide influence on soil enzymatic activities. Chemosphere 2001, 45, 417–425. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, H.; Zhou, Q. Using soil available P and activities of soil dehydrogenase and phosphatase as indicators for biodegradation of organophosphorus pesticide methamidophos and glyphosate. Soil Sediment Contam. 2011, 20, 688–701. [Google Scholar] [CrossRef]
- Nakatani, A.S.; Fernandes, M.F.; De Souza, R.A.; Da Silva, A.P.; Dos Reis-Junior, F.B.; Mendes, I.C.; Hungria, M. Effects of the glyphosate-resistance gene and of herbicides applied to the soybean crop on soil microbial biomass and enzymes. Field Crops Res. 2014, 162, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, M.; Tang, L.; Che, R.; Chen, H.; Blumfield, T.; Boyd, S.; Nouansyvong, M.; Xu, Z. Long-term harvest residue retention could decrease soil bacterial diversities probably due to favouring oligotrophic lineages. Microb. Ecol. 2018, 76, 771–781. [Google Scholar] [CrossRef]
- Fournier, J.C. Enumeration of the soil micro-organisms able to degrade 2,4-D by metabolism or co-metabolism. Chemosphere 1980, 9, 169–174. [Google Scholar] [CrossRef]
- Rivarola, V.; Fabra, A.; Mori, G.; Balegno, H. In vitro protein synthesis is affected by the herbicide 2,4-dichlorophenoxyacetic acid in Azospirillum brasilense. Toxicology 1992, 73, 71–79. [Google Scholar] [CrossRef]
- Fabra, A.; Duffard, R.; De Duffard, A.E. Toxicity of 2,4-dichlorophenoxyacetic acid to Rhizobium sp. in pure culture. Bull. Environ. Contam. Toxicol. 1997, 59, 645–652. [Google Scholar] [CrossRef]
- Durga Devi, K.M.; Beena, S.; Abraham, C.T. Effect of 2,4-D residues on soil microflora. J. Trop. Agric. 2008, 46, 64–66. [Google Scholar]
- Haahtela, K.; Kilpi, S.; Kari, K. Effects of phenoxy acid herbicides and glyphosate on nitrogenase activity (acetylene reduction) in root-associated Azospirillum, Enterobacter and Klebsiella. FEMS Microbiol. Lett. 1988, 53, 123–127. [Google Scholar] [CrossRef]
- Sachu, M.; Kynshi, B.L.; Syiem, M.B. A biochemical, physiological and molecular evaluation of how the herbicide 2, 4-dichlorophenoxyacetic acid intercedes photosynthesis and diazotrophy in the cyanobacterium Nostoc muscorum Meg 1. Environ. Sci. Pollut. Res. 2022, 29, 36684–36698. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Zhang, J.; Fang, Y.; Zheng, X.; Zhang, Y.; Chen, D. Impact of herbicide 2,4-dichlorophenoxyacetic acid butyl ester on soil nitrogen-transforming bacterial populations in two soils. Int. J. Agric. Biol. 2017, 19, 812–816. [Google Scholar] [CrossRef]
- De Oliveira, E.P.; Rovida, A.F.D.S.; Martins, J.G.; Pileggi, S.A.V.; Schemczssen-Graeff, Z.; Pileggi, M. Tolerance of Pseudomonas strain to the 2,4-D herbicide through a peroxidase system. PLoS ONE 2021, 16, e0257263. [Google Scholar] [CrossRef]
- Nandula, V.K.; Tyler, H.L. Effect of new auxin herbicide formulations on control of herbicide resistant weeds and on microbial activities in the rhizosphere. Am. J. Plant Sci. 2016, 7, 2429–2439. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.; Hu, X.; Xu, X. Dispersal of Bacillus subtilis and its effect on strawberry phyllosphere microbiota under open field and protection conditions. Sci. Rep. 2016, 6, 22611. [Google Scholar] [CrossRef] [Green Version]
- Mississippi State University Delta Agricultural Weather Center. Available online: http://deltaweather.extension.msstate.edu/coop-stoneville (accessed on 24 May 2022).
- NRCS Web Soil Survey. Available online: https://websoilsurvey.sc.egov.usda.gov/App/WebSoilSurvey.aspx (accessed on 29 October 2020).
- Tyler, H.L. Winter cover crops and no till management enhance enzyme activities in soybean field soils. Pedobiologia 2020, 81-82, 150666. [Google Scholar] [CrossRef]
- Jackson, C.R.; Tyler, H.L.; Millar, J.J. Determination of microbial extracellular enzyme activity in waters, soils, and sediments using high throughput microplate assays. J. Vis. Exp. 2013, 80, e50399. [Google Scholar] [CrossRef] [Green Version]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Weintraub, S.R.; Wieder, W.R.; Cleveland, C.C.; Townsend, A.R. Organic matter inputs shift soil enzyme activity and allocation patterns in a wet tropical forest. Biogeochemistry 2013, 114, 313–326. [Google Scholar] [CrossRef]
- Štursová, M.; Baldrian, P. Effects of soil properties and management on the activity of soil organic matter transforming enzymes and the quantification of soil-bound and free activity. Plant Soil 2011, 338, 99–110. [Google Scholar] [CrossRef]
- Schnecker, J.; Wild, B.; Hofhansl, F.; Alves, R.J.E.; Bárta, J.; Čapek, P.; Fuchslueger, L.; Gentsch, N.; Gittel, A.; Guggenberger, G.; et al. Effects of Soil Organic Matter Properties and Microbial Community Composition on Enzyme Activities in Cryoturbated Arctic Soils. PLoS ONE 2014, 9, e94076. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Liu, X.; Dong, F.; Xu, J.; Zheng, Y.; Li, J. Soil microbial communities response to herbicide 2,4-dichlorophenoxyacetic acid butyl ester. Eur. J. Soil Biol. 2010, 46, 175–180. [Google Scholar] [CrossRef]
- Nowak, J.; Telesiński, A.; Szymczak, J. Comparison of herbicides containing isoproturon, 2.4-D and dicamba on phosphatase activity in the soil and in spring wheat (Triticum aestivum L.). Electron. J. Pol. Agric. Univ. 2006, 9, 17. [Google Scholar]
- Ljungdahl, L.G.; Eriksson, K.-E. Ecology of Microbial Cellulose Degradation. In Advances in Microbial Ecology; Marshall, K.C., Ed.; Springer: Boston, MA, USA, 1985; Volume 8, pp. 237–299. [Google Scholar]
- Tabatabai, M.A.; Ekenler, M.; Senwo, Z.N. Significance of enzyme activities in soil nitrogen mineralization. Commun. Soil Sci. Plant Anal. 2010, 41, 595–605. [Google Scholar] [CrossRef]
- Turner, B.L.; McKelvie, I.D.; Haygarth, P.M. Characterisation of water-extractable soil organic phosphorus by phosphatase hydrolysis. Soil Biol. Biochem. 2002, 34, 27–35. [Google Scholar] [CrossRef]
- Schnürer, J.; Rosswall, T. Fluorescein diacetate hydrolysis as a measure of total microbial activity in soil and litter. Appl. Environ. Microbiol. 1982, 43, 1256–1261. [Google Scholar] [CrossRef] [Green Version]
- Dennis, P.G.; Kukulies, T.; Forstner, C.; Orton, T.G.; Pattison, A.B. The effects of glyphosate, glufosinate, paraquat and paraquat-diquat on soil microbial activity and bacterial, archaeal and nematode diversity. Sci. Rep. 2018, 8, 2119. [Google Scholar] [CrossRef] [Green Version]
- Zablotowicz, R.M.; Reddy, K.N.; Weaver, M.A.; Mengistu, A.; Krutz, L.J.; Gordon, R.E.; Bellaloui, N. Cover crops, tillage, and glyphosate effects on chemical and biological properties of a lower Mississippi Delta soil and soybean yield. Environ. Res. J. 2010, 4, 227–251. [Google Scholar]
- Zabaloy, M.C.; Carné, I.; Viassolo, R.; Gómez, M.A.; Gomez, E. Soil ecotoxicity assessment of glyphosate use under field conditions: Microbial activity and community structure of Eubacteria and ammonia-oxidising bacteria. Pest Manag. Sci. 2016, 72, 684–691. [Google Scholar] [CrossRef] [PubMed]
- Zabaloy, M.C.; Garland, J.L.; Gómez, M.A. An integrated approach to evaluate the impacts of the herbicides glyphosate, 2,4-D and metsulfuron-methyl on soil microbial communities in the Pampas region, Argentina. Appl. Soil Ecol. 2008, 40, 1–12. [Google Scholar] [CrossRef]
- Lupwayi, N.Z.; Harker, K.N.; Clayton, G.W.; O’Donovan, J.T.; Blackshaw, R.E. Soil microbial response to herbicides applied to glyphosate-resistant canola. Agric. Ecosyst. Environ. 2009, 129, 171–176. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, W.; Tang, L.; Heenan, M.; Xu, Z. Effects of nitrification inhibitor and herbicides on nitrification, nitrite and nitrate consumptions and nitrous oxide emission in an Australian sugarcane soil. Biol. Fert. Soils 2018, 54, 697–706. [Google Scholar] [CrossRef]
- Martens, D.A.; Bremner, J.M. Influence of herbicides on transformations of urea nitrogen in soil. J. Environ. Sci. Health Part B Pestic. Food Contam. Agric. Wastes 1993, 28, 377–395. [Google Scholar] [CrossRef]
- Newman, M.M.; Lorenz, N.; Hoilett, N.; Lee, N.R.; Dick, R.P.; Liles, M.R.; Ramsier, C.; Kloepper, J.W. Changes in rhizosphere bacterial gene expression following glyphosate treatment. Sci. Total Environ. 2016, 553, 32–41. [Google Scholar] [CrossRef]
Figure 1.
Weekly precipitation (bars), minimum air temperature (open circle), and maximum air temperature (closed circle) during the first (A) and second (B) study years. Times of sowing, herbicide treatments, and harvest are indicating by arrows.
Figure 1.
Weekly precipitation (bars), minimum air temperature (open circle), and maximum air temperature (closed circle) during the first (A) and second (B) study years. Times of sowing, herbicide treatments, and harvest are indicating by arrows.

Figure 2.
Activity of beta-glucosidase (A), cellobiohydrolase (B), fluorescein diacetate (FDA) hydrolysis (C), N-acetylglucosaminidase (NAGase)(D), and phosphatase (E) in 0–5 cm soil from 2,4-D-only (closed circle), 2,4-D+glyphosate (open circle), and untreated control (closed triangle) plots for each timepoint during the two-year study. Values represent mean ± standard error (n = 4).
Figure 2.
Activity of beta-glucosidase (A), cellobiohydrolase (B), fluorescein diacetate (FDA) hydrolysis (C), N-acetylglucosaminidase (NAGase)(D), and phosphatase (E) in 0–5 cm soil from 2,4-D-only (closed circle), 2,4-D+glyphosate (open circle), and untreated control (closed triangle) plots for each timepoint during the two-year study. Values represent mean ± standard error (n = 4).

Figure 3.
Activity of beta-glucosidase (A), cellobiohydrolase (B), fluorescein diacetate (FDA) hydrolysis (C), N-acetylglucosaminidase (NAGase) (D), and phosphatase (E) in 5–15 cm soil from 2,4-D-only (closed circle), 2,4-D+glyphosate (open circle), and untreated control (closed triangle) plots for each timepoint during the two-year study. Values represent mean ± standard error (n = 4).
Figure 3.
Activity of beta-glucosidase (A), cellobiohydrolase (B), fluorescein diacetate (FDA) hydrolysis (C), N-acetylglucosaminidase (NAGase) (D), and phosphatase (E) in 5–15 cm soil from 2,4-D-only (closed circle), 2,4-D+glyphosate (open circle), and untreated control (closed triangle) plots for each timepoint during the two-year study. Values represent mean ± standard error (n = 4).


{kind=link}
{kind=link}
{kind=link}
Table 1.
Farm management timeline for experimental field plots *.
| Activity | Year 1 | Year 2 | Field Site |
|---|---|---|---|
| Sowing | 20 March 2019 | 5 April 2021 | All plots |
| Herbicide application 1 | 17 April 2019 | 7 May 2021 | Treatment-specific plots |
| Herbicide application 2 | 17 May 2019 | 26 May 2021 | Treatment-specific plots |
| Harvest | 11 September 2019 | 15 September 2019 | All plots |
* Herbicide applications were either 2,4-D alone, 2,4-D plus glyphosate, or no herbicide treatment. All herbicides were applied at label rates.
Table 2.
Soil characteristics in 0–5 and 5–15 cm depths for each timepoint during the study period *.
Table 2.
Soil characteristics in 0–5 and 5–15 cm depths for each timepoint during the study period *.
| Moisture Content (%) | pH | Organic Matter (%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Depth | Treatment | Sowing | V2 | V8 | Harvest | Sowing | V2 | V8 | Harvest | Sowing | V2 | V8 | Harvest |
| Year 1 | |||||||||||||
| 0–5 cm | 24D | 15.22 | 15.43 | 14.76 | 6.4 | 7.11 | 7.39 | 7.02 | 6.76 | 3.95 | 3.97 | 4 | 3.72 |
| glyphosate+24D | 14.55 | 15.74 | 15.2 | 5.37 | 6.9 | 7.07 | 7 | 6.7 | 3.86 | 4.02 | 3.99 | 3.78 | |
| none | 15.1 | 14.53 | 14.56 | 6.58 | 7.02 | 7.07 | 6.98 | 6.72 | 3.9 | 3.82 | 3.84 | 3.7 | |
| 5–15 cm | 24D | 14.49 | 19.4 | 17.1 | 9.65 | 7.24 | 7.13 | 6.53 | 6.86 | 3.52 | 3.29 | 3.54 | 3.13 |
| glyphosate+24D | 15.12 | 20.89 | 15.61 | 9.19 | 7.02 | 7.14 | 6.29 | 6.81 | 3.4 | 3.17 | 3.41 | 3.11 | |
| none | 15.11 | 15.98 | 16.07 | 9.69 | 7.12 | 6.97 | 6.81 | 6.74 | 3.4 | 3.2 | 3.42 | 3.13 | |
| 2-way ANOVA | |||||||||||||
| Depth | ns | ns | 0.005 | <0.0001 | ns | ns | 0.0027 | ns | 0.001 | <0.0001 | <0.0001 | <0.0001 | |
| Treatment | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | |
| Depth × Treatment | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | |
| Year 2 | |||||||||||||
| 0–5 cm | 24D | 12.92 | 14.35 | 11.16 | 11.93 | 6.48 | 6.79 ab | 7.29 | 7.59 | 3.92 | 4.15 | 3.94 | 3.91 |
| glyphosate+24D | 12.54 | 14.96 | 11.7 | 12.86 | 6.45 | 6.83 a | 7.38 | 7.51 | 3.86 | 3.98 | 3.54 | 3.82 | |
| none | 13.46 | 14.44 | 9.85 | 13.36 | 6.5 | 6.69 b | 7.31 | 7.47 | 3.72 | 3.92 | 3.77 | 3.83 | |
| 5–15 cm | 24D | 14.31 | 16.18 | 13.29 | 14.08 | 6.47 | 7.44 cd | 7.87 | 7.59 | 3.45 | 3.75 | 3.4 | 3.45 |
| glyphosate+24D | 14.83 | 15.85 | 12.49 | 13.76 | 6.61 | 7.57 c | 7.91 | 7.59 | 3.37 | 3.65 | 3.34 | 3.281 | |
| none | 16.18 | 15.99 | 12.36 | 14.41 | 6.6 | 7.26 d | 7.92 | 7.63 | 3.28 | 3.56 | 3.37 | 3.37 | |
| 2-way ANOVA | |||||||||||||
| Depth | 0.0008 | 0.001 | 0.0324 | 0.0226 | ns | <0.0001 | <0.0001 | ns | 0.0011 | 0.0002 | 0.0215 | <0.0001 | |
| Treatment | ns | ns | ns | ns | ns | 0.0493 | ns | ns | ns | ns | ns | ns | |
| Depth × Treatment | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | |
* Values represent mean from each treatment for each timepoint. Means followed by different letters are significantly different. Significant p-values from two-way analysis of variance between treatment and depth for each timepoint are indicated. ns (not significant).
Table 3.
Correlations between soil parameters during the study period *.
| SM | pH | SOM | Phos | BG | NAGase | Cello | FDA | |
|---|---|---|---|---|---|---|---|---|
| SM | 1 | 0.0976 | 0.0536 | −0.322 | −0.1889 | −0.0849 | −0.1996 | 0.0546 |
| pH | 1 | 0.0293 | 0.2219 | −0.0343 | −0.1118 | −0.1551 | −0.1179 | |
| SOM | 1 | 0.2048 | 0.6585 | 0.508 | 0.6598 | 0.6381 | ||
| Phos | 1 | 0.5895 | 0.5492 | 0.4467 | 0.3753 | |||
| BG | 1 | 0.8343 | 0.9197 | 0.7793 | ||||
| NAGase | 1 | 0.8223 | 0.7262 | |||||
| Cello | 1 | 0.8469 | ||||||
| FDA | 1 |
* Significant Pearson correlations are in bold (α = 0.05).
Table 4.
Soil enzyme activities in the rhizosphere of maize plants one week following herbicide treatment at the V2 and V8 developmental stages *.
Table 4.
Soil enzyme activities in the rhizosphere of maize plants one week following herbicide treatment at the V2 and V8 developmental stages *.
| Year 1 | Year 2 | |||
|---|---|---|---|---|
| Treatment | V2 | V8 | V2 | V8 |
| Beta-glucosidase | ||||
| 2,4-D | 1.74 ± 0.23 | 1.32 ± 0.17 | 1.33 ± 0.09 | 2.62 ± 0.42 |
| 2,4-D+Glyphosate | 2.42 ± 0.39 | 1.66 ± 0.36 | 1.21 ± 0.09 | 2.24 ± 0.1 |
| No herbicide | 1.98 ± 0.47 | 2.06 ± 0.3 | 1.76 ± 0.31 | 2.36 ± 0.33 |
| Cellobiohydrolase | ||||
| 2,4-D | 0.53 ± 0.09 | 0.22 ± 0.01 | 0.23 ± 0 | 0.26 ± 0.01 |
| 2,4-D+Glyphosate | 0.61 ± 0.07 | 0.27 ± 0.05 | 0.2 ± 0.01 | 0.26 ± 0.01 |
| No herbicide | 0.52 ± 0.08 | 0.31 ± 0.02 | 0.27 ± 0.03 | 0.27 ± 0.02 |
| NAGase | ||||
| 2,4-D | 0.72 ± 0.05 | 0.38 ± 0.05 | 0.34 ± 0.02 | 0.53 ± 0.05 |
| 2,4-D+Glyphosate | 0.56 ± 0.06 | 0.38 ± 0.07 | 0.33 ± 0.02 | 0.46 ± 0.05 |
| No herbicide | 0.53 ± 0.09 | 0.43 ± 0.04 | 0.35 ± 0.07 | 0.49 ± 0.05 |
| Phosphatase | ||||
| 2,4-D | 3.53 ± 0.17 | 2.03 ± 0.19 | 4.11 ± 0.22 | 4.98 ± 0.69 |
| 2,4-D+Glyphosate | 3.98 ± 0.59 | 2.24 ± 0.42 | 3.74 ± 0.25 | 4.13 ± 0.31 |
| No herbicide | 3.49 ± 0.59 | 2.66 ± 0.24 | 4.6 ± 0.54 | 4.17 ± 0.52 |
* Values represent mean ± standard error.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tyler, H.L. Impact of 2,4-D and Glyphosate on Soil Enzyme Activities in a Resistant Maize Cropping System. Agronomy 2022, 12, 2747. https://doi.org/10.3390/agronomy12112747
AMA Style
Tyler HL. Impact of 2,4-D and Glyphosate on Soil Enzyme Activities in a Resistant Maize Cropping System. Agronomy. 2022; 12(11):2747. https://doi.org/10.3390/agronomy12112747
Chicago/Turabian StyleTyler, Heather L. 2022. "Impact of 2,4-D and Glyphosate on Soil Enzyme Activities in a Resistant Maize Cropping System" Agronomy 12, no. 11: 2747. https://doi.org/10.3390/agronomy12112747
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.